Des hôtels apicoles sont implantés dans les différentes stations pour voir si l'urbanisation affecte l'implantation des abeilles dans celles-ci. Les résultats ont montré qu'il n'y avait pas d'influence de l'urbanisation sur l'implantation des abeilles dans les hôtels.
I NTRODUCTION
- Généralités
- Origine des abeilles
- Diversité des abeilles
- Modes de vie
- Importance des abeilles
- Interactions plantes-abeilles
- Importance économique
- Les communautés d’abeilles
- Déclin des abeilles
- Facteurs du déclin
- Moyens de conservation
- Les abeilles dans le paysage urbain
Les abeilles sauvages sont un moteur majeur de l'important service écosystémique de la pollinisation (Gallai et al., 2009). Deuxièmement, les espèces d'insectes envahissantes ont également des effets sur les abeilles sauvages.

O BJECTIFS
M ATÉRIEL ET MÉTHODES
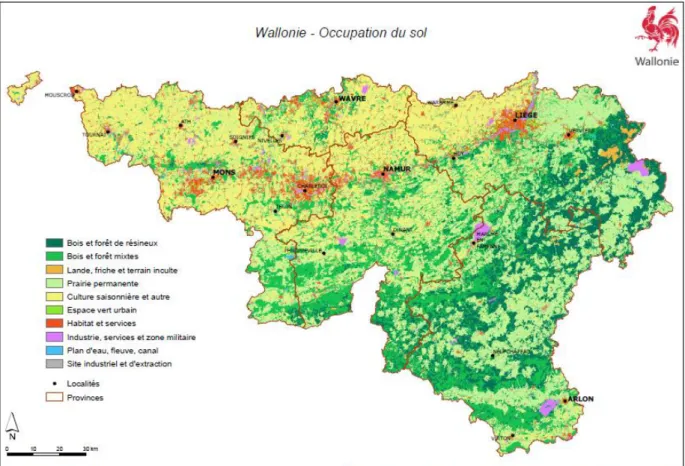
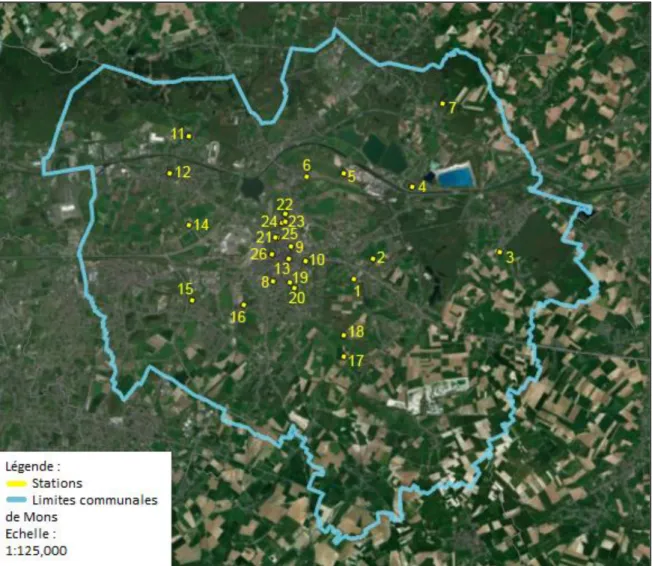
Présentation de la zone d’étude
De plus, la ville de Mons n'étant pas très grande, un rayon plus grand entraînerait un chevauchement important entre les tampons des différentes gares. Ces données proviennent de la base de données représentant les écotopes wallons v2.10 du géoportail LifeWatch Belgium.
Echantillonnage et récolte des données
Taille de la langue : s'agit-il d'une espèce à langue courte ou à langue longue. Nidification : c'est une espèce qui niche au sol ou sous le sol.
Analyses statistiques des données
- Caractérisation des communautés d’abeilles
- Analyses sur les hôtels
Lorsque l'ANOVA (valeur p < 0,05) montre qu'il existe une différence significative, un test de Tukey est effectué pour voir quelles catégories d'étanchéité diffèrent les unes des autres. Lorsque ce test montre qu'il existe une différence significative, un test de classement de Wilcoxon est effectué pour voir quelles catégories d'étanchéité diffèrent les unes des autres.
R ÉSULTATS
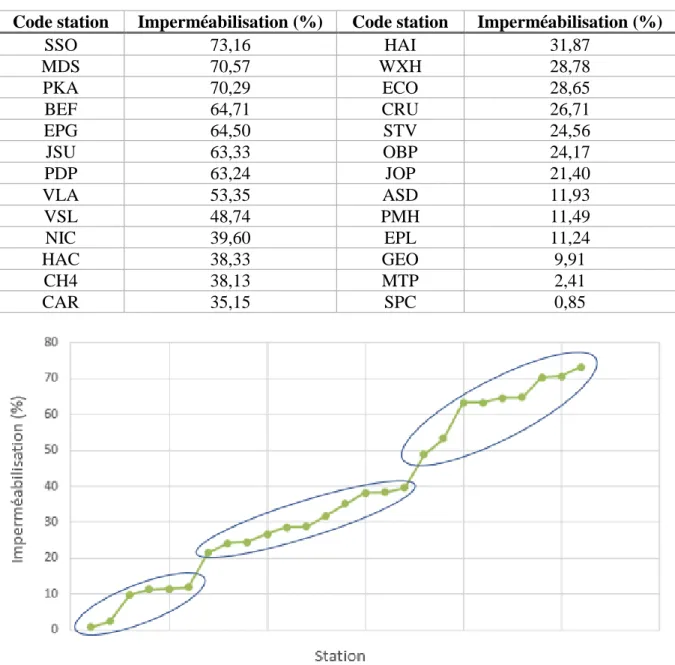
Pourcentage d’imperméabilisation des stations
La catégorie 1 représente alors les stations sous 20% d'étanchéité, la catégorie 2 représente les stations entre 20% et 45% d'étanchéité et la catégorie 3 représente les stations supérieures à 45%. Rq.1 : Les stations ECO, MDS et PKA n'ayant pas fait l'objet d'échantillonnage d'abeilles sauvages (cf. 3.2), la répartition des stations au sein des 3 catégories n'est pas la même pour les analyses sur les peuplements d'abeilles. Rq.2 : Les hôtels qui ont été volés (GEO, MTP, PMH, JOP, PKA et PDP) n'ont pas pu être inclus dans les analyses (cf. 3.3.2), la répartition des stations au sein des 3 catégories n'est pas la même pour les analyses sur les hôtels.
Par conséquent, la catégorie 1 a 3 stations, la catégorie 2 en a 9 et la catégorie 3 en a 7.
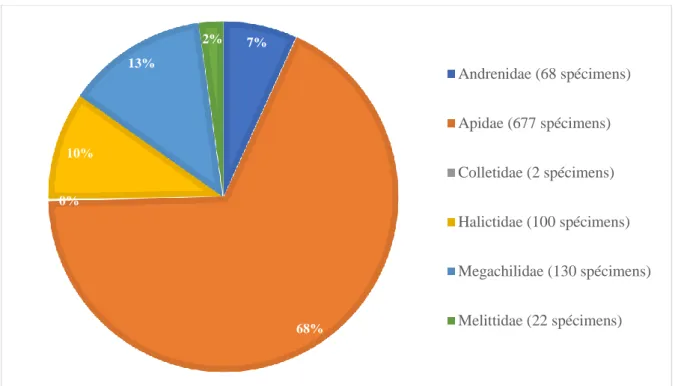
Caractérisation des communautés d’abeilles
- Commune de Mons
- Communautés d’abeilles en fonction de l’urbanisation
41 Le test PerMANOVA arrive à la conclusion qu'il n'y a pas de différences significatives entre les 3 catégories d'étanchéité concernant les communautés d'abeilles (R2 = 0,11, p-value = 0,11). La visualisation graphique du NMDS montre qu'aucune distinction n'est faite entre les différentes catégories d'étanchéité (Fig.16). L'analyse du dendrogramme (Fig.17) montre également que les communautés ne sont pas regroupées selon la catégorie d'étanchéité à laquelle elles appartiennent.
Concernant la richesse florale, l'ANOVA ainsi que les boîtes de dispersion montrent qu'il n'y a pas de différences significatives entre les différentes catégories d'imperméabilisation (Fig. 19f).
Hôtels
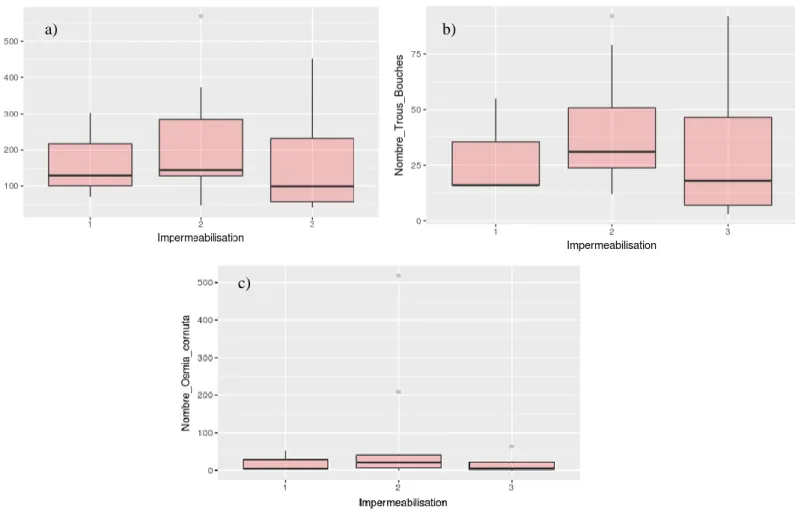
D'après le nombre total d'échantillons sortis de chaque hôtel (Fig.21a), aucune différence significative entre les 3 catégories d'étanchéité n'a été détectée. Selon le nombre de tiges bloquées pour chaque hôtel (Fig.21b), aucune différence n'est également enregistrée par rapport à l'étanchéité. Le test ANOVA à un facteur effectué sur l'abondance d'Osmia cornuta indique qu'il n'y a pas de différence significative entre les différentes catégories d'étanchéité.
La même observation peut être faite en regardant le nuage de points (Fig.21c).
D ISCUSSION
Echantillonnage
Communautés d’abeilles au sein de la commune de Mons
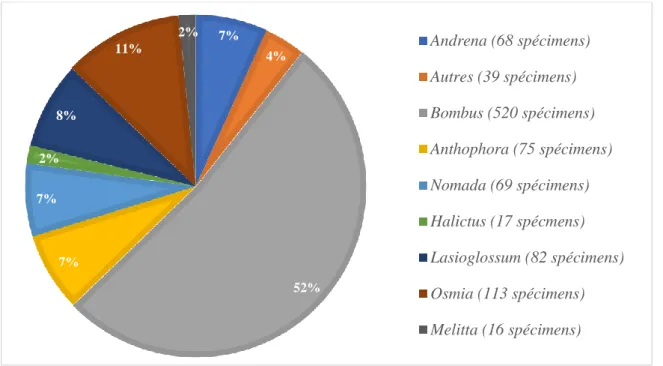
En tant que généralistes, ils peuvent butiner une large gamme de fleurs sans avoir de préférence (Potts et al., 2003). Bien que de nombreuses espèces soient en déclin, elle reste abondante et est pratiquement considérée comme une extension en Belgique (www.atlashymenoptera.net). Parmi les espèces récoltées en 2018, 3 sont inscrites dans la loi sur la protection de la nature (annexe IIb de l'arrêté du 6 décembre 2001) comme espèces protégées en Wallonie.
La présence de ces quelques espèces protégées, ainsi que l'importante diversité spécifique de la commune, font que Mons mérite une attention particulière par rapport à la préservation voire à l'enrichissement de ces communautés.
Communautés d’abeilles le long d’un gradient d’urbanisation
52 les abeilles sont relativement bien préservées en milieu urbain contrairement à Ahrné et al. 2009), qui a conclu que la diversité des espèces était négativement affectée par l'urbanisation croissante. En fait, il a été démontré que la connectivité joue un rôle important dans la richesse des abeilles sauvages (Gonçalves et al., 2014). En effet, ces ressources de nidification sont connues pour jouer un rôle important dans la structure des sociétés apicoles (Potts et al., 2005).
Il en va de même pour l'abondance et la diversité des fleurs, qui jouent un rôle dans la structure de la communauté (Potts et al., 2003).
C ONCLUSION ET PERSPECTIVES
B IBLIOGRAPHIE
Interactions plantes-pollinisateurs : caractérisation de la qualité pollinique de deux cucurbitacées au cours de l'ontogenèse, apparition et transport dans le corps de l'abeille, 191p. Baisse de l'abondance, augmentation de la diversité et modification de la structure communautaire des abeilles sauvages (Hyménoptères : Anthophiles) le long a. Gembloux : Unité de Zoologie Générale et Appliquée de la Faculté des Sciences Agronomiques de Gembloux.
Pollen hosts of western Palearctic bees of the genus Colletes (Hymenoptera: Colletidae): The Asteraceae paradox.
A NNEXES
Spécialisation = Spécialisation alimentaire (polylectique, oligolectique ou cleptoparasite), Langue = langue courte ou langue longue, Mode de vie = Solitaire, Social ou cleptoparasite, Nidification = Mode de nidification souterraine ou aérienne ou mixte. Langue de spécialisation Mode de vie Nidification Andrena Angustior Court solitaire polylectique chez Anrena Barbilabris Court solitaire polylectique chez Andrena Bicolor Court solitaire polylectique chez Andrena Chrysoscelis Court solitaire polylectique chez Andrena cineraria Court polylectique chez Andrena dorsata -court polylectique polylectique fula flavipes Polylectique Polylectique chez les polylectiques Polylectique Polylectique Polylectique Enlena Dorsata Fula Fe Solitaire chez Andrena fulvago Solitaire court polylectique chez Andrena gravida Solitaire court polylectique chez Andrena haemorrhoa Solitaire court polylectique chez Andrena humilis Solitaire court polylectique chez Andrena minutula Solitaire court polylectique chez Andrena nigroaenea Solitaire court polylectique chez Andrena nigroaenea Solitaire court polylectique chez Andrena sc. Andrena tibialis Polylectique Court Solitaire Ci-dessous Andrena vaga Oligolétique Court Solitaire Ci-dessous Andrena ventralis Oligolétique Court Solitaire Ci-dessous Anthidiellum strigatum Polylectique Long Solitaire Ci-dessous Anthidium manicatum Polylectique Long Solitaire Ci-dessus Anthophora plumipes Polylectique Grand Solitaire Long Solitaire Long Solitaire Mixte Anthopulata ci-dessus Mix toparasite Long Cleptoparasite Ci-dessous Bombus campestris Clepto parasite Grand Cleptoparasite Mixte Bombus hortorum Polylectique Grand Social Sous Bombus humilis Polylectique Grand Social Sous Bombus hypnorum Polylectique Grand Social Sous Bombus lapidarius Polylectique Grand Social Sous Bombus lucorum Polylectique Grand Social Sous Bombus pascuorum Polylectique Grand Social Social A Clepris Grand Social A BClep parasite Grand Kleptoparasite Mixte Bombus terrestris Polylectic Long Social Parmi. Chalicodoma ericetorum Oligolectique Long Solitaire Dessus Chelostoma florisomne Oligolectique Long Solitaire Dessus Chelostoma rapunculi Oligolectique Long Solitaire Dessus Coelioxys aurolimbata Kleptoparasite Grand Kleptoparasite Dessus Colletes daviesanus Oligolérique Daviesanus Oligolérique Mixte Soliget Court sypoda hirtipes Oligolectique Court Solitaire Sous Halictus confusa Short Polylectic Social Under Halictus rubicundus Short Polylectic Solitaire au-dessous de Halictus scabiosae Court Polylectique Social au-dessous de Halictus tumulorum Court Polylectique Social au-dessous de Heriades truncorum Long Oligoltique Solitaire au-dessus de Hoplitis adunca Long Oligoltique Solitaire au-dessus de Hoplitis leucomelana Grand Polylectique Hyla Solitaire lectique Court Solitaire Mixte Lasioglossum calceatum Polylectique Court Solitaire au-dessous de Lasioglossum fulvicorne Polylectique Court Sol Itary Ci-dessous Lasioglossum laticeps Solitaire court polylectique ci-dessous Lasioglossum lativentre Solitaire court polylectique ci-dessous Lasioglossum leucozonium Solitaire court polylectique ci-dessous Lasioglossum malachurum court Solitum court Polylectica ci-dessous Solitum court Solitum Minut io court Solitaire Polylectica court Lasioglossum nitidiusculum Solitaire court Polylectica ci-dessous Polylectica Solitaire Ci-dessous Lasioglossum pallens Court Polylectica Solitaire Ci-dessous Lasio glossum parvulum Polylectic Solitaire court Ci-dessous Lasioglossum pauxillum Court Polylectica Solitaire Ci-dessous Lasioglossum punctatissimum Court Polylectica Solitaire Ci-dessous Court Polylectica Solitaire um villosulum Court Polylectic Solitaire Ci-dessous Lasioglossum zonulum Court Polylectic Solitaire Ci-dessous Macropis europaea Oligoletic Court Solitaire Ci-dessous Megachile alpicola Polylectic Long Solitaire Ci-dessous Mega chili centuncularis Grand Solitaire polylectique au-dessus de Megachile circumcincta Grand Solitaire polylectique mixte Megachile lagopoda Grand Solitaire polylectique au-dessus de Grand Solitaire polylectique au-dessus de rotation Megachile au-dessus de Megachile willughbiella Solitaire polylectique long au-dessus de Melecta albifrons albifrons Cleptoparasite Grand cleptoparasite mixte.
71 Melecta luctuosa Cleptoparasite Longue Cleptoparasite En dessous Melitta leporina Oligolectique Courte Solitaire En dessous Cleptoparasite Longue Cleptoparasite En dessous Nomada fabriciana Cleptoparasite dess Longue Cleptoparasitee ssous Nomada flavoguttata Cleptoparasite Longue Cleptoparasite En dessous Nomada fucata Cleptoparasite Longue Cleptoparasite En dessous Nomada goodeniana Cleptoparasite Longue Cleptoparasite En dessous Nomada lathburiana Cleptoparasite Longue Cleptoparasite En dessous Cleptoparasite Longue Cleptoparasite En dessous Nomada Longue Cleptoparasite En dessous Nomada panzeri Cleptoparasite sous Nomada sheppardana Cleptoparasite Longue Cleptoparasite En dessous Nomada signata Cleptoparasite Longue Cleptoparasite En dessous Nomada succincta Cleptoparasite Longue Kleptoparasite En dessous Osmia caerulescens Polylectique Longue Solitaire Au-dessus Osmia hornuta Polylectique Longue Solitaire Au-dessus. Panurgus calcaratus Oligolectique Courte Solitaire En dessous Sphecodes crassus Cleptoparasite Courte Cleptoparasite En dessous Sphecodes ferruginatus Cleptoparasite Courte Cleptoparasite En dessous Sphecodes miniatus Cleptoparasitee Courte Cleptoparasitele Cleptoparasitee ptoparaziti En dessous Sphecodes reticulatus Cleptoparasite Courte Cleptoparasite En dessous Stelis breviscula Cleptoparasite Longue Cleptoparasite sipër Stelis punctulatissima Kleptoparazit Longue Kleptoparazit Au-dessus Thyreus orbatus Kleptoparazit Longue Kleptoparazit Mixte Xylocopa violacea Polylectique Longue Solitaire Au-dessus.