Fiordaliso William (2020) Structure des communautés d'abeilles sauvages dans le paysage de Bergen : étude pour l'élaboration d'un plan d'action. De nombreuses études se sont penchées sur la question de l'écologie des abeilles sauvages en ville.
Introduction
Les abeilles sauvages
- Présentation
- Une fonction écologique : la pollinisation
- Diversité écologique
- Les communautés d’espèces
- Le déclin des abeilles
Commençons par nous attaquer au taxon Melittidae (3 genres et 9 espèces en Belgique, cf. Drossart et al., 2019). Chez les espèces du sol, soit 49 % des espèces belges (Drossart et al., 2019), les cellules larvaires seront construites sous terre.
L’urbanisation
- Les environnements urbains
- Effets négatifs supposés
- Effets positifs supposés
- Structuration écologique
Les fleurs de printemps apparaissent plus tôt, tandis que les variétés d'automne apparaissent plus tard (Neil et al., 2010). Premièrement, les milieux classés comme « urbains » englobent de nombreuses réalités différentes (McIntyre et al., 2000).
Objectifs et questions de recherche
Matériel et méthode
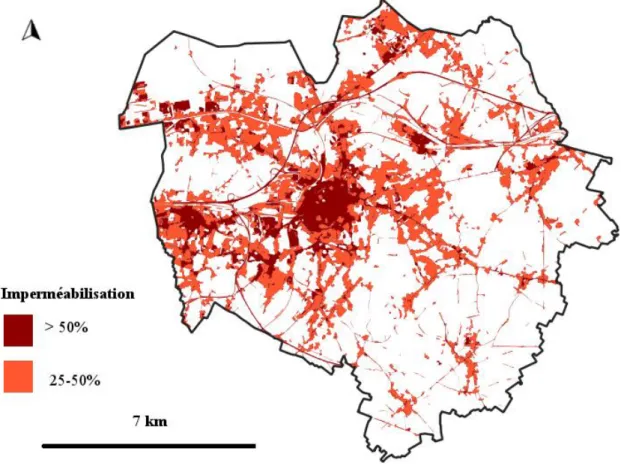
Site d’étude : le paysage montois
Pour cette raison, nous avons collecté plusieurs sources supplémentaires lors de la compilation de l'atlas et de l'inventaire. Il s'appuie sur les travaux du laboratoire de zoologie de l'Université de Mons (Pr. P. Rasmont) et de la faculté d'agronomie de l'Université de Gembloux (Pr. E. Haubruge).
Description des sites de collecte
- Choix des sites
- Relevés botaniques
- Analyse du paysage
Grâce à la Flore bleue de Belgique (Lambinon et al., 2012), nous avons également calculé la proportion de plantes indigènes parmi l'ensemble des ressources florales. TABLEAU 3 Typologie d'occupation du sol associée à la grille haute résolution (Radoux et al., 2017).
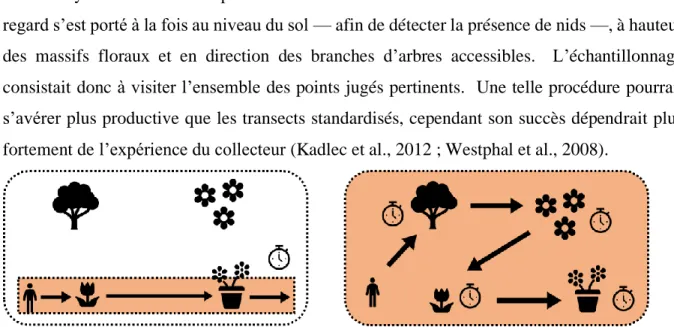
Méthode d’échantillonnage
De plus, les intervalles de collecte auraient gagné à être plus courts (une à deux semaines) pour atténuer les variations causées par les conditions météorologiques (William et al., 2001). Ces caisses en bois, remplies d'une centaine de tiges de bambou creuses, permettent d'enregistrer des espèces troglodytiques qui passeraient autrement inaperçues (Westphal et al., 2008).
Mise en collection et identification
Par conséquent, le jeu de données relatif à ces hôtels à insectes reste incomplet et ne peut faire l'objet d'une étude statistique.
Description des communautés d’abeilles sauvages
- Complétude de l’échantillonnage
- Indices de biodiversité
- Raréfaction
- Diversité bêta
La mesure la plus simple pour décrire la biodiversité d'un lieu est celle de la "richesse en espèces", c'est-à-dire le nombre total d'espèces qui y sont recensées. Pour cette raison, il peut être judicieux de préférer une autre mesure d'exhaustivité : la couverture.
Structuration des communautés
- Autocorrélation spatiale
- Modélisation des effets de l’urbanisation sur la diversité
- Sélection du meilleur modèle et model averaging
- Diversité bêta et urbanisation
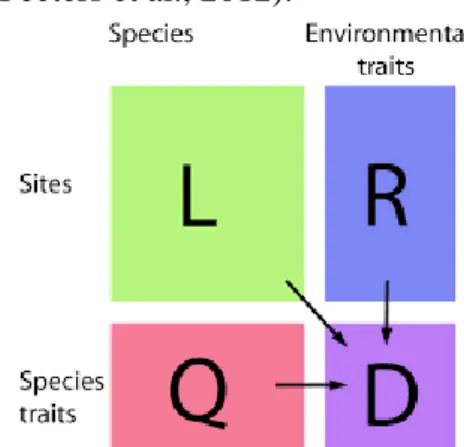
- Analyse de la relation traits-environnement
Il présente la valeur de l'indice de Moran en fonction de la valeur du « décalage ». Cela correspond aux classes « spécialisées » et « opportunistes » de la liste rouge belge (Drossart et al., 2019).
Résultats
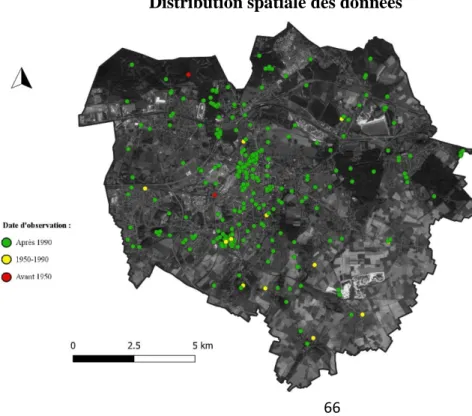
Inventaire et atlas des abeilles de la région de Mons
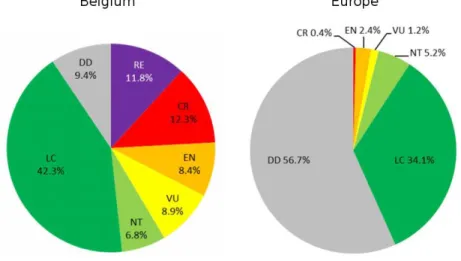
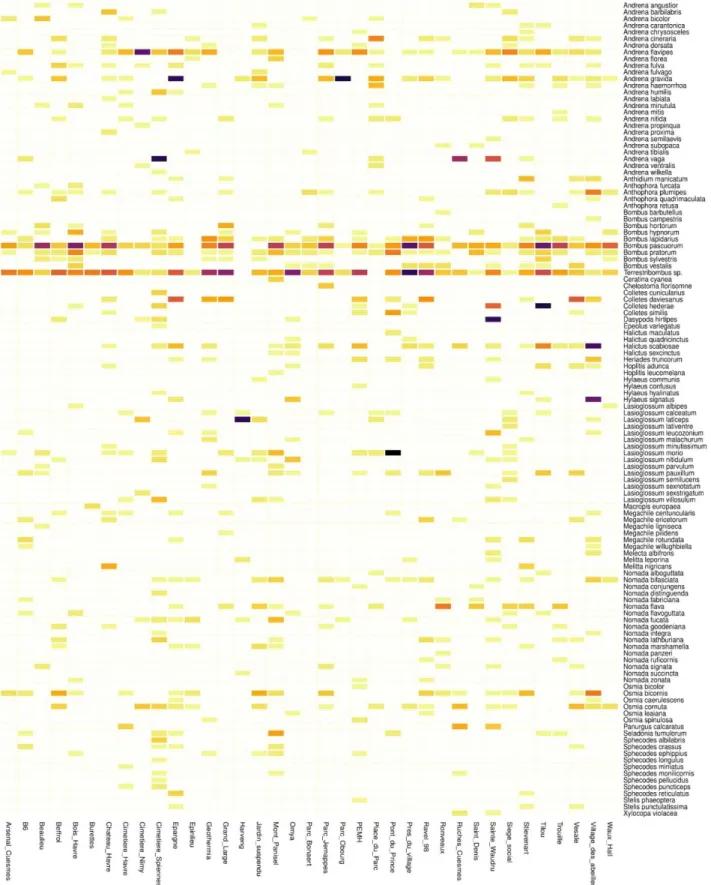
Une densification des observations est perceptible en centre-ville et à proximité du campus universitaire (amas de points centraux), et autour du terril de l'Héribus (sud-ouest de l'amas central). Les résultats de l'inventaire, en nombre d'espèces et d'observations, sont présentés dans le tableau ci-dessous. On peut alors décrire la composition taxonomique de cette collection, en précisant la proportion d'espèces menacées pour chaque famille.
Enfin, les Melittidae forment le groupe le moins diversifié avec seulement 5 espèces et ne présentent aucune espèce menacée. Cela permet notamment de localiser les taxons les plus rares et les sites d'intérêt où se concentrent les espèces menacées (cimetière de Spiennes, terril de l'Héribus, campus de Nimy, carrières d'Harmignies.

Composition des communautés
- Complétude de l’échantillonnage
- Description de l’assemblage
- Nombres de Hill raréfiés
Le tableau de distribution des traits (TAB 7) nous donne la proportion d'individus et d'espèces avec un état de trait. Les abeilles solitaires ici sont beaucoup plus nombreuses que les abeilles eusociales en nombre d'espèces, mais la relation est inversée en ce qui concerne l'abondance. La proportion d'espèces représentées par le groupe des kleptoparasites est également relativement élevée compte tenu du nombre d'individus collectés.
Le nombre d'espèces présentes sur chaque site peut être comparé en fonction de l'abondance à l'aide des courbes de dilution de la Figure 17. En regardant le graphique, la plupart des sites se caractérisent par un fort couvert (à l'exception du site n°15 : arsenal de Cuesmes) augmentent leur nombre d'espèces assez rapidement, en fonction de l'effort d'échantillonnage.
Facteurs structurant l’assemblage
- Modélisation de la biodiversité alpha et de l’abondance
- Diversité bêta et urbanisation
- Relation traits-environnement
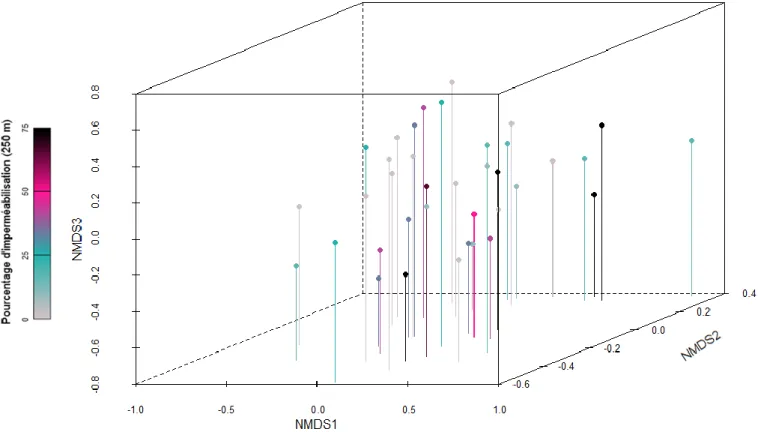
La qualité de la représentation fournie par le NMDS peut être appréciée à l'aide de la figure 20. Le test visant à déterminer si l'un des niveaux de la variable « Société » présentait un degré d'étanchéité différent des autres rejetait l'hypothèse d'une urbanisation. Une corrélation négative (p-value) est également détectée entre la durée de la période d'activité et la fermeture des terres.
Le mois moyen d'occurrence et la durée moyenne de la période d'activité selon le degré d'urbanisation (250m). Le graphique du bas montre la durée moyenne de la période d'activité des espèces, pondérée par l'abondance de chacune d'elles.
Discussion
Inventaire et atlas des abeilles montoises
Cependant, la comparaison entre ces villes est risquée, notamment en raison de la distance géographique qui les sépare (Ropars et al., 2018) et de la différence entre les zones étudiées. Si la ville de Louvain présente un ratio similaire de 3,8% (calculé grâce à Drossart et al., 2019, sans tenir compte de l'espèce DD20), une localité moins anthropisée comme Han-sur-Lesse présente 26,4% des taxons menacés ( également converti). En effet, l'utilisation massive d'apports d'azote dans les cultures intensives a favorisé le développement d'une flore plus homogène, adaptée à la présence d'une grande quantité d'azote assimilable dans le sol (Carvalheiro et al., 2020 ; Carvell et al., 2006 ; Kleijn et al., 2009 ; Rasmont, 2006).
Pour étayer cette idée, nous pouvons nous concentrer sur les fleurs sur lesquelles le plus grand nombre d'individus ont été capturés lors de nos collectes (TAB 10). ONGLET 10 Ces six types de fleurs sont celles dont le plus grand nombre d'individus a été collecté.
Caractéristiques des communautés d’abeilles sauvages dans la région de Mons
Il est intéressant de noter que Bombus hypnorum, habituellement surreprésenté en milieu urbain (Folschweiller et al., 2020 ; Fowler, 2015), ne fait pas partie des espèces les plus abondantes dans la commune. Les deux dernières espèces dominantes sont Bombus pratorum (commun en ville cf. Folschweiller et al., 2020) et Andrena vaga. En comparaison, les données collectées à Paris semblent très différentes : dans la capitale française, les espèces les plus communes appartenaient toutes au genre Lasioglossum (Ropars et al., 2018).
Les chiffres obtenus ici correspondent à la fourchette fournie par l'auteur concernant les milieux tempérés (Danforth et al. 2019). Une telle hétérogénéité des communautés peut surprendre, mais ce ne serait pas un cas isolé dans la littérature (Normandin et al., 2017 ; Williams et al., 2001).
Effets de l’urbanisation sur les communautés d’abeilles
- Biodiversité alpha et abondance
- Diversité bêta
- Traits fonctionnels
Nous allons illustrer ce phénomène par un exemple tiré de notre étude : les sites du clocher et de la collégiale Sainte-Waudru, bien que voisins, sont radicalement différents l'un de l'autre. Dans ce cas, les valeurs observées sont assez faibles (environ 30%), ce qui signifie qu'une part importante de l'écart zéro reste inexpliquée. L'analyse de la diversité et de l'abondance alpha à l'aide de modèles linéaires généralisés a donné des résultats assez surprenants.
Ces chiffres suggèrent qu'une augmentation de la couverture urbaine se traduirait par une augmentation de la diversité plus importante que celle produite par une augmentation équivalente des terres agricoles. Par conséquent, nous soupçonnons que le couvert forestier joue un rôle négatif en termes de biodiversité.
Conclusion
La deuxième caractéristique pour laquelle une corrélation significative a été démontrée est la durée de la période active. Premièrement, cela ne pourrait être qu'une conséquence de la baisse du nombre de bourdons en ville, ces abeilles ont généralement une longue période d'activité par rapport aux espèces solitaires. Deuxièmement, les zones urbaines sont caractérisées par des périodes de floraison partiellement artificielles.
En général, il y aura des pics à des moments bien précis de l'année, entrecoupés de périodes moins denses.
Perspectives
Localisation : Réserve des Prés du village (réserve naturelle périurbaine) et autour des carrières de calcaire d'Harmignies. Nomada integra (VU) : uniquement au cimetière de Spiennes et sur le campus de Nimy Hôtes : Andrena humilis, Andrena fulvago. Ce type de milieu (très présent au sud de la commune), alors qu'il fait déjà l'objet d'une certaine forme de protection, devrait intégrer des objectifs de conservation liés aux espèces d'abeilles ici répertoriées.
Enfin, on aura remarqué que plusieurs lieux reviennent systématiquement dans la liste : le cimetière de Spiennes, la carrière de craie d'Harmignies, le campus de Nimy, etc.
Bibliographie
The neonicotinoid clothianidin disrupts solitary navigation in Osmia cornuta in a lab test. Species-specific foraging strategies of the syntopic and synchronous bees Panurgus calcaratus (Scopoli, 1763) and Dasypoda hirtipes (Fabricius, 1793) (Hymenoptera: Apidae). Atlas of the Human Planet - Mapping the human presence on Earth with the global human settlement layer.
The Relative Performance of Native Bee Sampling Methods: An Empirical Test and Literature Review. Urban bumblebees are unaffected by the proportion of intensively developed land in urban environments of the US industrial Midwest.
Annexes
Atlas
Inventaire
Lasioglossum minutissim Halictidae LC Lasioglossum morio Halictidae LC Lasioglossum nitidiusculum Halictidae LC Lasioglossum nitidulum Halictidae LC Lasioglossum pallens Halictidae LC Lasioglossum parvulum Halictidae LC Lasioglossum pauxillum Halictidae LC Lasioglossum punctatissimum Halictidae LC Lasioglossum semilucentus Halictidae LC Lasioglossum sexnotatum Halictidae LC Lasioglossum sexstrigatum Halictidae LC Lasioglossum um villosulum Halictidae LC Lasioglossum zonulum Halictidae LC Macropis europaea Melittidae LC Macropis fulvipes Melittidae LC Megachile centuncularis Megachilidae LC Megachile circumcincta* Megachilidae EN Megachile ericetorum Megachilidae LC Megachile lagopoda Megachilidae CR Megachile ligniseca Megachilidae LC Megachile pilidens Megachilidae CR Megachile rotundata Megachilidae LC Megachile versicolor Megachilidae LC Megachile willughbiella Megachilidae LC Melecta albifrons Apidae NT Melecta luctuosa** Apidae CR Melitta leporina Melittidae LC Melitta nigricans Melittidae LC Nomada alboguttata Apidae LC Nomada bifasciata Apidae LC Nomada conjungens Apidae LC Nomada onderscheidbare Apidae EN Nomada fabriciana Apidae LC. Osmia bicolor Megachilidae LC Osmia bicornis Megachilidae LC Osmia caerulescens Megachilidae LC Osmia cornuta Megachilidae LC Osmia leaiana Megachilidae LC Osmia spinulosa Megachilidae NT Panurgus calcaratus Andrenidae LC Seladonia confusa Halictidae VU Seladonia tumulorum Halictidae LC Sphecodes albilabris Halictidae LC S phecodes crassa Halictidae LC Sphecodes ehippius Halictidae LC Sphecodes ferruginatus Halictidae. .
Calendrier de récolte
Exemple de relevé botanique
Sites et variables associées
Matrice des traits écologiques
Autocorrélation spatiale
Choix du rayon d’analyse
Diagnostic des modèles linéaires généralisés
Effets interannuels