La région éthiopienne est la seule région où les trois familles (Dasypodaidae, Melittidae s.str. et Meganomiidae) sont présentes. Les Meganomiidae ont un comportement de nidification intermédiaire entre les Dasypodaidae et les Melittidae s.str. Révision du genre d'abeille Capicola (Hymenoptera : Apoidea : Melittidae) distribué en Afrique du Sud-Ouest.

Introduction
Michener (2000) included Capicola in Hesperapis, where Capicola and Capicoloides are two subgenera among seven other Nearctic subgenera of Hesperapis. 2007a) demonstrated the monophyly of Capicola and recognized Capicola as a genus. This monographic revision is based on our own publications (Appendices I-VIII) and a synthesis of the literature on melittid bees.
Systematics and biogeography of the Melittidae s.l
Family Dasypodaidae Börner 1919
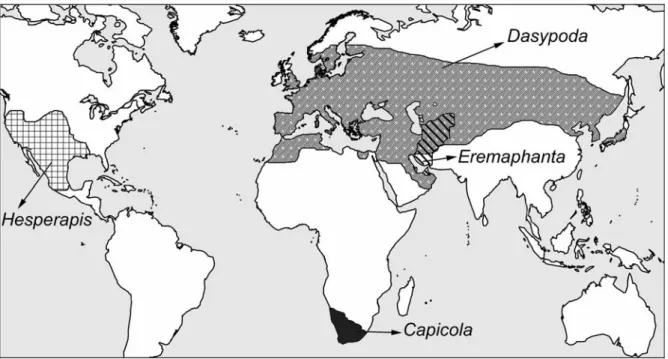
N1= number of species included in the genus (described spp.. undescribed spp.); N2= number of species included in the subgenus (described spp. The taxonomic assignment of Afrodasypoda definitely requires further samples and a global phylogenetic study of Dasypodaidae. Hesperapis and Capicola share synapomorphies, especially the shape of the stigma, the two sub-marginal cells ( the first longer than the other), the galea crest inserted in front of the maxillary palpus insertion and the scopa restricted to the outside of the hind tibia and basitarsus.
Rather, both genera differ in the shape of the male S6 and the female pygidial plate, which shows a strong longitudinal median relief in Capicola females. The shape of the female pygidial plate is also diagnostic for many species, which in Melittidae s.l. very unusual. 15 The characteristics that confirm the special status of the nine species of Eremaphanta are: the ratio between the length of the palpi and the glossa, the development of the apical bands on the terga and the appearance (sculpture, coloration) of the propodeal triangle.
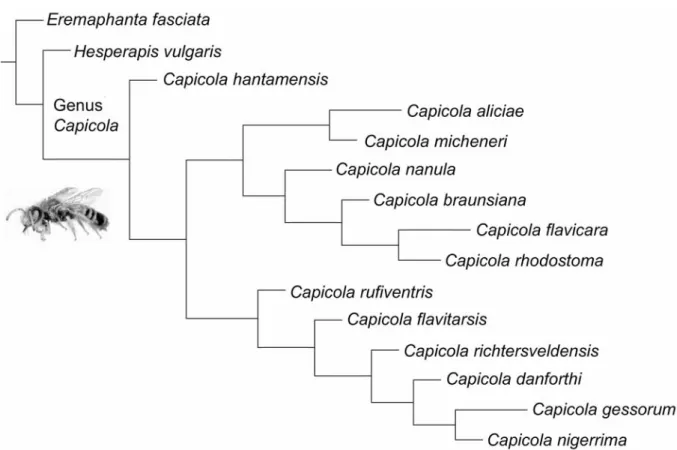
This genus is characterized by the flat abdomen, the inner marginal margin of the gonostyle, and the flat pygidial plate of the female. Michener (2000) considered nine subgenera in the genus Hesperapis, but Michez et al. 2007a) proposed placing the South African species in a separate monophyletic genus (Capicola).
Family Meganomiidae Michener 1981
Family Melittidae s.str. Schenk 1860
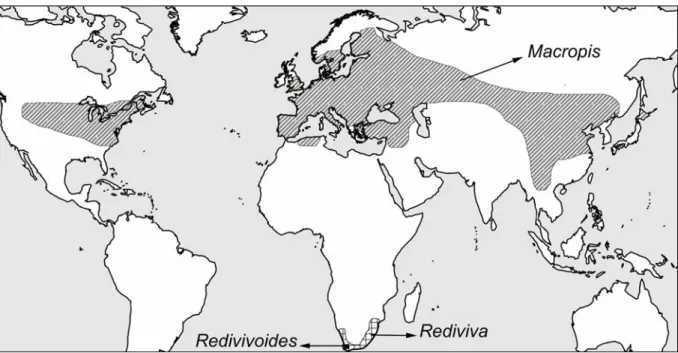
As written above, most Dasypodaidae (Afrodasypoda, Capicola, Dasypoda, Eremaphanta and Promelitta) and all Meganomiidae are restricted to the xeric regions of the Old World (Michener 1981, Michez & Patiny 2006, Michez et al. 2007a, b, c) . Among Melittidae s.str. the genus Macropis is particularly characterized by the yellow markings of the male (fig. 25), the two submarginal cells (while the other Melittidae s.str. have three) and the well-developed pygidial plate of both sexes (Michener 1981; Annex II). Each species can be defined by a unique combination of some features: shape of male yellow facial mark, pubescence of propodeal triangle, puncture of terga, shape of pygidial plate, shape of hidden male sterna, and shape of male genitalia (Michener 1981, Wu & Michener 1986, Michez & Patiny 2005).
At the species level, Melitta is unambiguously characterized by a combination of several morphological structures, such as the proportions of the proboscis, the separation of the clypeus and mesonotum, the sculpture of the propodeal triangle, the shape of the hidden male sternum (S6-8) and the male genitalia (Michez & Eardley in press). Melitta appears to be more diverse in the temperate and Mediterranean climate regions of the Old World region, such as the Mediterranean Basin, the Central European Valley, Sichuan, China, and the southwestern region of Africa (Michez & Eardley in press). Diagnostic characters are numerous at the species level: color of metasoma, shape of labrum, length of malar space, body dress, apical margin of terga, length of foreleg of female, and shape of hidden sternum of male (Whitehead & Steiner 2001, Whitehead et. al. in press).
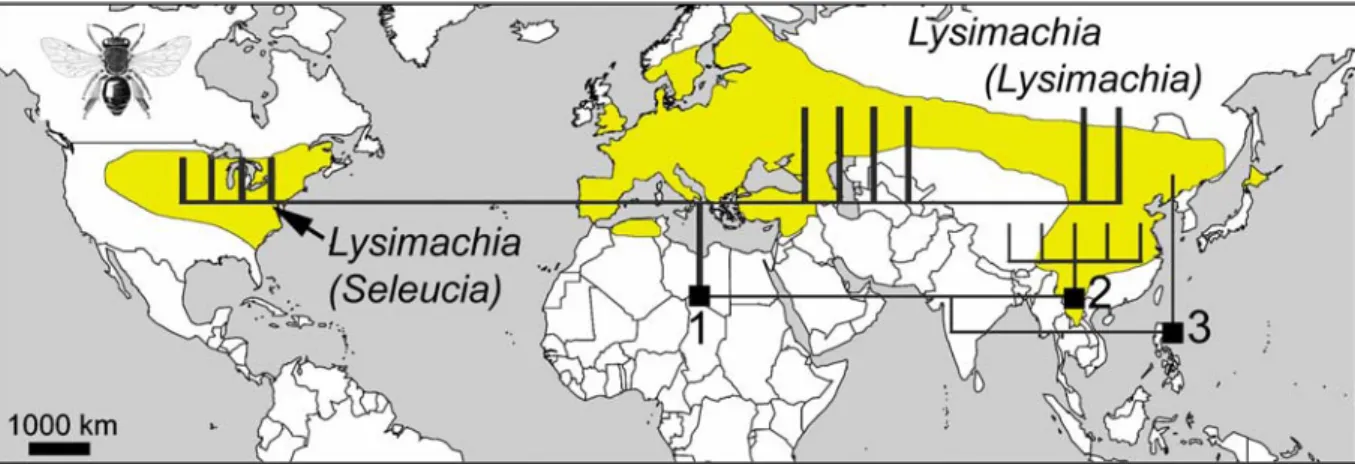
Worldwide distribution of the genus Melitta (from Snelling & Stage 1995, Eardley & Kuhlmann 2006, Michez & Eardley in press). Redivivoides show some plesiomorphies such as the shape of the volsella and the pygidial plate present in the male.
Biology of the Melittidae s.l
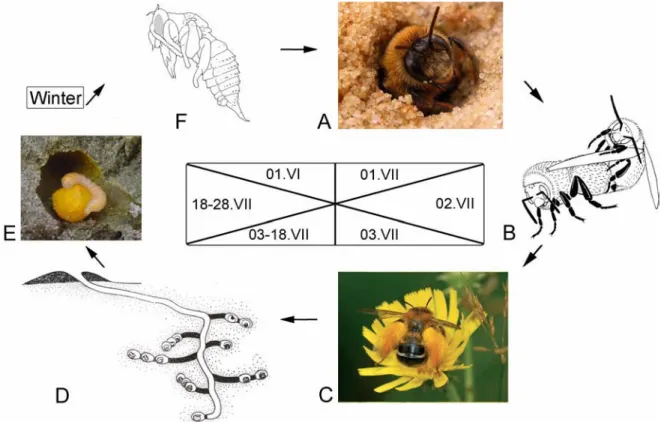
General cycle of development
24 Mating behavior is described only for Dasypoda hirtipes (Dasypodaidae) (review of bee mating behavior see Ayasse et al. 2001). The main tunnel penetrates the surface at a low angle and descends vertically after a few decimetres. Cells are single as in Haplomelitta and Hesperapis (except H. larrae) or arranged in linear arrays as in Capicola and Dasypoda.
The tunnels and cells are unlined and waterproof, but the walls are stronger than the foundation. The stocks are formed in various shapes, spherical in Capicola, Haplomelitta and Hesperapis or with basal cones in Dasypoda. The female lays one egg on top of the pollen ball and closes the cell with a small finger.
The main tunnel is about 20 to 40 cm in depth, clearly less deep than Dasypodaidae. Macropis uses oil flower as cell covering (Cane et al. 1983), whereas Melitta uses Dufour's gland secretion (Celary 2006).

Host-plants
In parentheses, percentage of main host plant family; pollen counts are corrected for volume; *=Hundred percent of data. Taxon Field Main host plants Taxon (continued) Field Main host plants Genus Haplomelitta Michener H. Amblyapis) arida 3;3 Zygophyllaceae (66%) H.(Hesperapis) elegantula - Boraginaceae*. The field observations recorded in the literature and our studies of the available series suggest a strong orientation of flower selection towards different plant taxa in Euasteridae.
The pattern of host plant use is different in other parts of the Hesperapis clade. Females feed on pollen and oil on Lysimachia, while nectar is usually collected from a number of host plants (Fig. However, only 13 of these plant families account for more than 5% percent of visits for at least one Melitta species (Michez . & Eardley in press ).
The main host plants are Diascia (Scrophulariaceae), but some Rediviva forage on oil from other Scrophulariaceae (Hemimeris and Bowkeria), Iridaceae and Orchidaceae (tab. Oil-gathering bees of the genus Rediviva and South African oilseeds display rare plant-pollination relationships, with some plant species depend solely on one Rediviva species for their pollination (Steiner & Whitehead Pauw 2006).

Evolution of the Melittidae s.l
Phylogeny and host-plants of the Melittidae s.l
In the fixed-specialist pattern, the ancestral host plant appears to have been inherited through the evolution of the genus. 1= shift from oligolecty on Asteraceae to polylecty with strong preference of Cistaceae; 2= shift from oligolecty on Asteraceae to oligolecty on Dipsacaceae. Phylogeny and host plants of Capicola (from Michez & Kuhlmann 2007; Michez & Timmermann unpublished data).
1= shift from Campanulaceae to Aizoaceae; 2= shift from Aizoaceae to Fabaceae; 3= shift from Aizoaceae to Campanulaceae; 4= shift from Aizoaceae to Asteraceae. However, the ancestral host plant appears to have a strong influence on the current host plants visited by contemporary species. The potential alternative host plants appear to be limited to plant taxa with similar morphology.
In contrast, choice of alternative host plants is not atavistic (i.e., related to the previous ancestral host plant) within lineages that exhibit a sequence specialist pattern. This hypothesis is supported by the recent discovery of a fossil bee, Paleomacropis eocenicus from the early Eocene (~53 myBP) (Michez et al. 2007c).
Origin and diversification of the Melittidae s.l
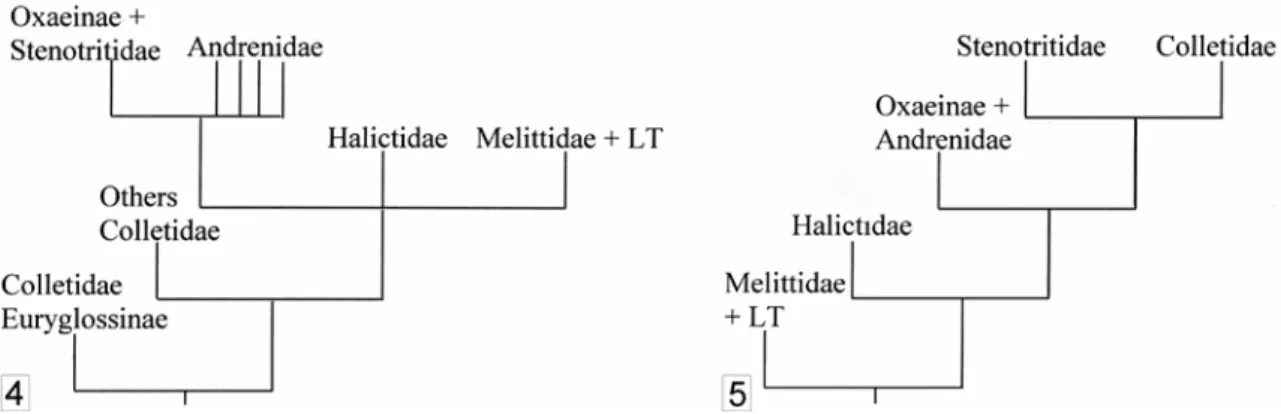
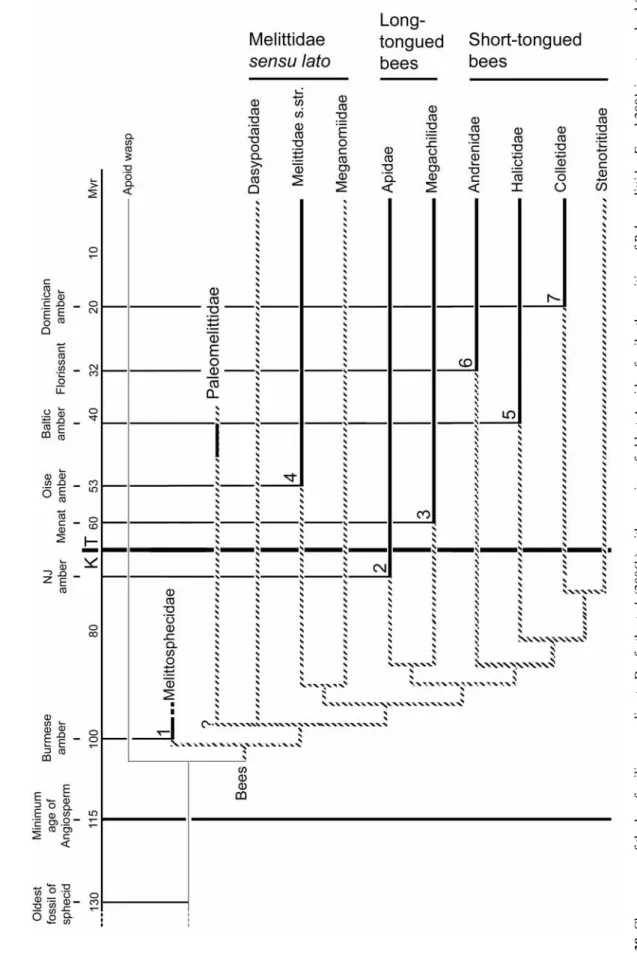
Michez – Monographic revision of the Melittidae s.l. Melittid basal” topology might indicate an African origin of bees, but Danforth et al. 2006b) have not developed a comprehensive phylogenetic or biogeographical argument (see next chapter). These studies allow us to envision a scenario of early bee diversification. 2004) explored the molecular phylogeny of Halictidae with extensive biogeographical analysis. The results in Ascher (2004) and Almeida (2007): (i) confirmed the monophyly of the Andrenidae, Colletidae and Stenotritidae; (ii) suggested that the Andrenidae probably originated from western continents, rather from South America, as suggested by the distribution of the genera Alocandrena, Euherbsia and Orphana (Patiny 2001); (iii) indicated that colletid bees probably appeared in Australia before the separation of Antarctide (i.e. the paleocontinent lumping Australia + Antarctica + South America) but after the separation between Africa and South America.
The resulting temporal distribution of the fossil record for bees provides additional evidence for the hypothesis of Alexander & Michener (1995) and Danforth et al. Taken together, the four latter sources of argument allow us to present a global picture of the origin (temporal and geographic) and early diversification of bees. This fossil is logically younger than the oldest fossil of the common ancestor (the Apoidea Spheciformes) recorded from ~130 myBP.
Michez – Monographic review of Melittidae s.l. 40 Phylogeny and biogeography of modern taxa of Melittidae s.l. seem to indicate that bees appeared in Africa. Finally, all the oldest fossil bee records indicate that the derived clade of bees LT and Melittidae s.str. were present in the late Cretaceous or early Cenozoic.
How to become a bee?
LT bees” was probably rapid given their basal position and the limitation of angiosperm diversification in the Middle Cretaceous. Diversification of derived ST bees would have occurred much more recently, in the Cenozoic, which explains the absence of old fossil records from this group. In this hypothesis, the emergence of high-level Apoidea Apiformes taxa could be "explosive" (Grimaldi 1999).
The short-tongued bees may have been very rare during the Cretaceous, which explains their absence in fossil records. They could have benefited from the K/T boundary, which was detrimental to the specialized group such as Melittidae s.l., basal Megachilidae and basal Apinae (Labandera et al. 2002).
Future research
Notes on the autumnal display of the spring desert bumblebee, Hesperapis fulvipes Crawford (Hymenoptera, Apoidea). Molecular phylogeny of the large carpenter bees, Genus Xylocopa (Hymenoptera: Apidae), based on mitochondrial DNA sequences. The major opsin gene is useful for inferring the higher-level phylogenetic relationships of corbicular bees.
World revision of the oil-harvesting bee genus Macropis Panzer 1809 (Hymenoptera, Apoidea, Melittidae) with description of a new species from Laos. Review of the bee genus Eremaphanta Popov 1940 (Hymenoptera: Melittidae), with description of a new species. Molecular phylogeny of Old World stingless bees (Hymenoptera: Apidae: Meliponini) and non-monophyly of the large genus Trigona.
Biology and larvae of the cleptoparasitic beetle Townsendiella pulchra and nest biology of its host Hesperapis larrae (Hymenoptera: . Apoidea). Molecular phylogenetics of the exoneurin allodapine bees reveal an ancient and enigmatic distribution from Africa to Australia.