Je tiens à exprimer ma gratitude à ceux qui ont rendu ce travail possible. Je remercie chaleureusement le Professeur Pierre RASMONT de m'avoir accueilli au sein du Département de Zoologie et de m'avoir fourni la logistique nécessaire au développement de ce projet. Je ne lui serai jamais assez reconnaissant de m'avoir initié à la cause de Macropidinus, de son soutien constant au cours de l'année écoulée.
NOTTON du département d'entomologie du Natural History Museum de Londres qui m'a généreusement prêté une partie du matériel de sa collection. Enfin, à toutes les personnes qui n'ont pas été citées et qui ont contribué de près ou de loin à la réalisation de ce travail, sachez que je les remercie du fond du cœur.
Introduction
- Systématique des abeilles
- La famille des Melittidae
- Phylogénie
- Biologie
- La tribu des Macropidini
- Paleomelitta nigripennis
- Evolution des choix floraux
- Oligolectisme vs polylectisme
- Les abeilles récolteuses d’huile
- Morphométrie géométrique
- Aperçu
- Définition et typologie des Landmarks
- Morphométrie géométrique appliquée aux ailes
- Objectifs
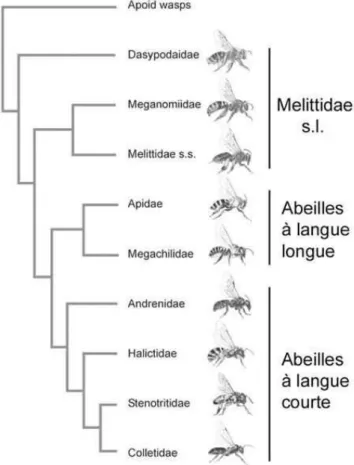
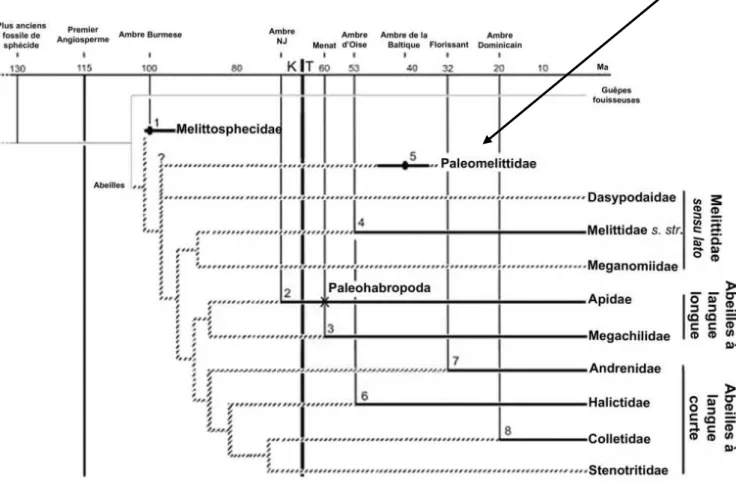
Les abeilles à langue courte ne sont plus considérées comme ancestrales (Danforth et al., 2006a, b) ; les caractéristiques de l'éblouissement seraient dues à une convergence évolutive. Une étude récente basée sur des données morphologiques et moléculaires (Michez et al., 2009b) définit les statuts tribaux au sein des Melittidae (Fig.4). On les trouve dans les régions tempérées, xériques et méditerranéennes de l'Ancien Monde et du Néarctique (Michener, 1979 ; Michez et al., 2009b).
Récemment, une révision phylogénétique basée sur des arguments morphologiques et moléculaires détermine le statut actuel de la tribu Macropidini (Michez et al., 2009b). De plus, un mâle a été prélevé chez un représentant de la famille des Convolvulaceae (Michez et al., 2007c). Certaines espèces sont représentées en Amérique du Sud et en Afrique (Hao et al., 2004).
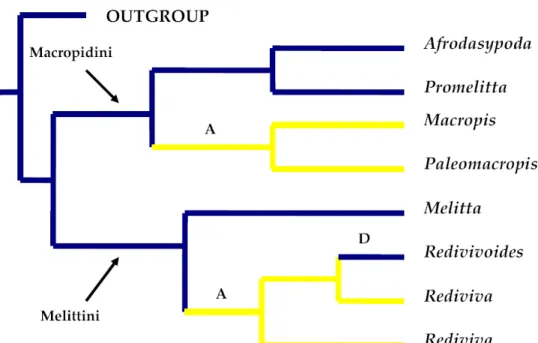
Distribution, phylogénie et associations de plantes hôtes chez Macropis (d'après Michez & Patiny, 2005 et Michez et al., 2008).
Matériel et méthode
Morphométrie géométrique
- Matériel
- Méthode
- Photographie de l’aile
- Positionnement des landmarks
- Analyses statistiques
Nous avons tenté de collecter 20 échantillons pour chaque espèce ainsi que pour le groupe de comparaison dans la mesure de la disponibilité des données. Les différentes manipulations effectuées lors de l'étude de la forme de l'aile peuvent être divisées en trois phases : photographie de l'aile, placement des repères et analyse statistique. Toute déformation de l'aile lors de la prise de vue constitue un biais ; le placer entre la lame et la lamelle permet d'obtenir une image qui reflète au mieux la forme réelle de l'aile (Fig. 11).
Positionnement des 15 repères sur l'aile antérieure gauche d'un Macropidini (d'après une illustration d'une aile d'Eomacropis glaesaria, Engel, 2001). 1 Extrémité interne gauche de la cellule marginale II 2 Extrémité interne la plus à droite de la cellule marginale II 3 Intersection de la veine marginale avec la première veine. 9 La côte ulnaire croise la côte basale I 10 La côte basale croise la côte discoïde I 11 La côte discoïde croise la première côte.
14 Pointe interne inférieure gauche de la deuxième cellule discoïde II 15 Intersection de la veine anale avec la nervure médiane. La numérisation des coordonnées cartésiennes bidimensionnelles des amers a été réalisée à l'aide du logiciel tps-DIG (Rohlf, 2006a). La première phase de l'analyse consiste à superposer les coordonnées des amers selon la méthode procuste GLS (« Generalized Laids-squares ») (Bookstein, 1991).
Ces dernières manipulations permettent d'éliminer toutes les variations qui ne concernent pas la forme de l'objet. Techniquement, une « gauchissement relatif » correspond à une analyse en composantes principales (ACP) basée sur les coordonnées alignées des points de repère (c'est-à-dire les procustes) (Michez et al., 2009a). 2001), les « warps relatifs » correspondent à la composante principale et définissent une forme d'espace dans laquelle les individus se replacent (Aytekin et al., 2007). Un histogramme représentant le nombre d'individus en fonction de la distance au centre de gravité du groupe Macropis est produit par le logiciel R 2.9.2.

Phylogénie moléculaire
- Matériel
- Méthode
- Extraction de l’ADN
- Gènes amplifiés
- PCR
- Séquençage
- Analyses phylogénétiques
Ces gènes sont nucléaires et se caractérisent par un faible taux de mutation (Danforth et al., 2006a ; Michez et al., 2009b). Ce pigment protéique est impliqué dans la phototransduction au niveau de la membrane des cellules photosensibles. Ce gène a été utilisé dans de nombreuses études phylogénétiques, notamment chez les arthropodes et les myriapodes, grâce à sa facilité d'amplification et d'adaptation (Danforth et al., 2006a).
Ce gène a une résolution phylogénétique pouvant aller jusqu'au sous-genre (Simon et al., 1994). Les amorces ainsi que les différents paramètres PCR sont détaillés dans l'étude de Michez et al. L'alignement des séquences a été effectué à l'aide du logiciel MAFFT ver.6 (Katoh et al., 2002) en utilisant les paramètres par défaut.
Six puces ont été construites puis analysées : une pour chaque gène (y compris l'exon et l'intron d'EF-1α) ainsi qu'une puce qui inclut tous les gènes. Les analyses du maximum de vraisemblance (ML) ont été réalisées à l'aide du logiciel GARLI 2.0 (Zwickl, 2011). Pour le gène EF-1α, nous avons considéré séparément la partie codante (EF1Ex) de l'intron (EF1In).
Les analyses par méthodes bayésiennes (MB) ont été réalisées à l'aide du logiciel MR BAYES 3.1.2 (Ronquist & Huelsenbeck, 2003). L'analyse est arrêtée après l'observation dans le graphique de la constance des valeurs de probabilité à travers les générations. Les topologies soutenues par des probabilités ultérieures supérieures ou égales à 0,95 sont considérées comme significatives (Wilcox et al., 2002).
Analyses palynologiques
Résultats
Morphométrie géométrique
- Relative warps analysis
- Histogramme
- Dendrogramme
L'"analyse des déformations relatives" est réalisée à partir d'une matrice de 130 individus pour 30 variables (15 repères en coordonnées (x, y)). Les deux premiers axes de l'analyse effectuée sur le jeu de données sont présentés dans la figure 14. Afin de visualiser une variance maximale de l'ensemble de données, les axes 1 et 3, ainsi qu'une représentation tridimensionnelle de la distribution des points sont présentés dans les figures 15 et 16 respectivement.
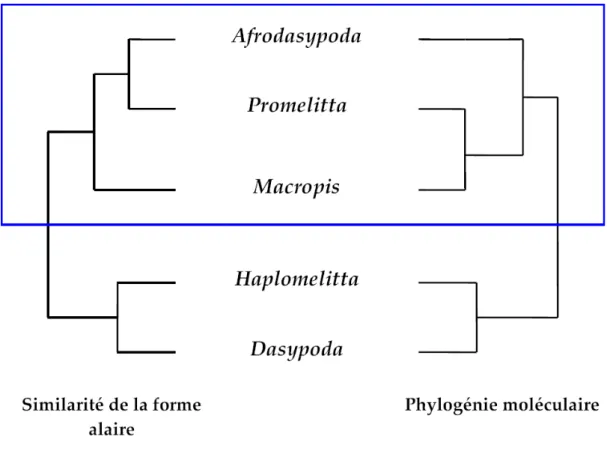
Notez que dans la description de ces résultats, la formule "est plus proche de" fera référence à une similitude dans la forme de l'aile, et non à une proximité phylogénétique. 76 % de la variance totale de l'ensemble de données est expliquée par les deux premiers axes de l'"analyse des distorsions relatives". 71 % de la variance totale dans l'ensemble de données est expliquée par l'axe 1 et l'axe 3.
83% de la variance totale de l'ensemble de données est expliquée par les trois premiers axes de. L'histogramme de la figure 17 montre le nombre d'individus en fonction de la distance au centre du groupe Macropis. L'histogramme est divisé en deux parties : un grand pic correspondant à Macropis et une zone plus étendue correspondant aux autres individus de l'ensemble de données.
Un continuum de déformation semble exister entre Afrodasypoda, Promelitta et Paleomacropis entraînant un déplacement du repère 10 (c'est-à-dire l'intersection de la côte basale avec la côte discoïde) vers la gauche. De la même manière, une déformation progressive apparaît au niveau du repère 15 (i.e. intersection de la veine anale avec la nervure médiane transversale) entre Macropis, Paleomelitta et Eomacropis. Quant aux ailes d'Haplomelitta et de Dasypoda, elles présentent un rétrécissement dans le sens de la largeur, qui s'explique par la fusion des repères 7 et 8 (i.e. intersection de la côte cubitale avec la première côte récurrente et intersection de la côte ulnaire avec la première côte ulnaire transversale, respectivement).

Phylogénie moléculaire
Les probabilités postérieures sont indiquées en noir ; les valeurs bootstrap sont affichées en rouge. L'arbre basé sur le gène 28S (Fig.19) montre que la tribu Macropidini est bien supportée, à la fois en bayésien et en maximum de vraisemblance. Les relations entre les espèces de l'exogroupe (Dasypoda hirtipes, Rediviva mcgregori, Meganomia binghami, Melitta leporina) ne sont pas significatives.
L'arbre obtenu par analyse combinée des quatre gènes (Fig. 20) montre que les genres sont correctement séparés et significativement soutenus par les deux méthodes. Par rapport à l'arbre obtenu par l'analyse du gène 28S, la tribu Macropidini comprend Afrodasypoda plumipes, qui occupe une position basale dans la tribu.

Palynologie
Discussion
- Méthode de phylogénie moléculaire
- Morphométrie géométrique vs phylogénie moléculaire
- Systématique des Macropidini
- Eomacropis glaesaria
- Paleomacropis eocenicus
- Paleomelitta nigripennis
- Origines multiples des structures adaptées à la collecte d’huile
- Biogéographie des Macropidini
- La transition Afrique-Eurasie
- Connexions Paléarctique-Néarctique au début du Tertiaire
- Apparition et dispersion du genre Macropis
- Oligolectisme des Macropis
En se référant à la littérature, la forme des ailes des abeilles n'en est probablement pas une. De plus, la morphométrie géométrique regroupe correctement les Macropidini, suggérant que la forme des ailes forme une synapomorphie partagée par les lignées de cette tribu. Cela signifie que l'appartenance ou non aux Macropidini peut être déterminée en fonction de la forme de l'aile.
Eomacropis glaesaria est inclus dans la définition actuelle de la tribu Macropidini (Michez et al., 2009b). La forme de l'aile d'Eomacropis glaesaria présente une grande similitude avec celle de Paleomelitta nigripennis. Eomacropis glaesaria montre secondairement une similitude dans la forme des ailes avec le genre Macropis.
Paleomacropis eocenicus a des appendices qui peuvent être associés à la collecte d'huile (c.f. suggérant qu'il pourrait former un groupe frère du genre Macropis (Michez et al., 2007b). Paleomacropis eocenicus ne partage aucune affinité de forme d'aile avec les deux autres fossiles, Eomacropis glaesaria et Paleomelitta eocenicus L'hypothèse d'un ancêtre non oléicole et la survenue indépendante d'adaptations morphologiques appuient l'hypothèse formulée par Michez et al.
Cette connexion semble avoir été peu utilisée pour la répartition de la faune et de la flore (Sanmartin et al., 2001). Les connexions transpaléarctiques ont été entravées par la mer de Turgai du Jurassique moyen (-180 Ma) à l'Oligocène (-30 Ma) (Sanmartin et al., 2001). Le cas du polylectisme chez les abeilles du genre Meganomia en est un exemple (Michez et al., 2010).
Conclusion
Perspectives
Bibliographie
Marmoot garaa keelloo (Marmota flaviventris) ‘bakka boca qaban’ (Rodentia, Sciuridae): diimoorfiizimii saalaa, guddinaa fi aloomeetirii mandible, Zoomorphology. Faayiloojinii beeylada orkidee (Hymenoptera: Apinae: Euglossini): DNA fi morfoloojiin moodeelota walqixa ta’an, faayiloojineetiksii molakiyuulaa fi jijjiirama argamsiisu. Irra deebiin gosa beeyladaa Kaapiikoolaa (Hymenoptera: Apoidea: Melittidae), Joornaalii Awurooppaa Entomology.
Ti kadaanan a fossil ti melittid nga uyokan (Hymenoptera: Apiformes) manipud idi nasapa nga Eoseno ti Oise (Pransia), Zoological Journal ti Linnean Society. Di Ninamnama a Polilekty iti Uyokan a Genus a Meganomia (Hymenoptera: Apoidea: Melittidae), Ti Pagwarnak ti Entomologo a Kagimongan ti Kansas. Monograpiko a rebision ti henero ti uyokan a Melitta Kirby 1802 (Hymenoptera: Apoidea: Melittidae), Annals ti Entomologo a Kagimongan ti Pransia (n. s.
Phylogeny of the bee family Melittidae (Hymenoptera: Anthophila) based on combined molecular and morphological data, Systematic Entomology. Pollen hosts of western Palearctic bees of the genus Colletes (Hymenoptera: Colletidae): the Asteraceae paradox, Biological Journal of the Linnean Society. Phylogeny and biogeography of bees of the tribe Osmiini (Hymenoptera: . Megachilidae), Molecular Phylogenetics and Evolution.
Global stingless bee phylogeny supports ancient divergence, variability and long-distance dispersal, Biological Journal of the Linnean Society. Studies of the phylogeny and classification of long-tongued bees (Hymenoptera: Apoidea), The University of Kansas Science Bulletin. Patterns of animal distribution, variability and diversification in the Holarctic, Biological Journal of the Linnean Society.