Assim, os nutrientes que completam este farto prato, em ordem decrescente de quantidade, são carboidratos, lipídios, proteínas e sais minerais. Verdade seja dita, as vitaminas e os minerais não são decompostos durante a digestão. Portanto, a quebra dos macronutrientes libera gradativamente vitaminas e minerais para que as células intestinais possam se beneficiar desses nutrientes, tão necessários à nossa sobrevivência.
Desta forma, todos os monossacarídeos, ácidos graxos, aminoácidos, vitaminas e sais minerais podem ser absorvidos pelo organismo.

Meta da aula
Pré-requisito
Conhecendo os micronutrientes
O enxofre participa intensamente desse enovelamento, já que o enxofre do aminoácido cisteína (Figura 2.7) está na forma -SH. A vitamina B1 (tiamina) (Figura 2.20), por exemplo, participa diretamente de reações catalisadas pelas enzimas piruvato e -cetoglutarato desidrogenase. A riboflavina (vitamina B2) (Figura 2.21) fornece as moléculas reativas do sistema mononucleotídeo de flavina (FMN) e dinucleotídeo de flavina adenina (FAD).
A biotina, assim como o ácido fólico, participa de reações de transferência de carbono de uma molécula para outra dentro das células (Figura 2.23).

Metas da aula
Pré-requisitos
Aldeídos, cetonas e ésteres
Neral geranial Figura 3.3: Estrutura química do geranial e do neral usada para conferir odor cítrico aos perfumes. O famoso Chanel No5 (Figura 3.4), criado em 1925 por Ernest Beaux, foi o primeiro perfume famoso a utilizar aldeídos sintéticos em sua composição. Nestes casos, o nome cetona deve indicar a posição onde este grupo está localizado na cadeia de hidrocarbonetos (Figura 3.11).
Se o grupo aldeído estiver ligado a um anel hidrocarboneto, a nomenclatura é feita adicionando o termo carboxaldeído ao nome do hidrocarboneto (Figura 3.13). O sufixo ic é substituído pelo sufixo ato, acrescenta-se a palavra de e o nome do radical hidrocarboneto ligado ao oxigênio carboxílico (Figura 3.18). Os nomes comuns dos ésteres são formados da mesma forma, conforme mostrado na Figura 3.18, utilizando, porém, a nomenclatura comum do ácido carboxílico original (Figura 3.19).
Uma letra grega também é adicionada para se referir ao tamanho do anel ( para anéis de 4 membros, para anéis de 5 membros, para anéis de 6 membros) (Figura 3.20). As lactonas de ácido alifático podem ser nomeadas adicionando o sufixo olide ao nome do hidrocarboneto correspondente, indicando a posição da hidroxila que forma o anel lactona (Figura 3.21). As interações intermoleculares entre os compostos mostrados na Figura 3.22 e na Tabela 3.1 são de dois tipos: dipolo-dipolo, envolvendo carbonilas, e interações de van der Waals, envolvendo cadeias de hidrocarbonetos.
Conforme mostrado na Figura 3.22 e na Tabela 3.1, não há diferença significativa no ponto de ebulição entre aldeídos e cetonas de mesmo peso molecular, indicando que as interações intermoleculares entre eles são equivalentes. Os valores do ponto de ebulição dos ésteres são inferiores aos dos aldeídos e cetonas de mesmo peso molecular (Figura 3.22 e Tabela 3.1).

Química dos aminoácidos e proteínas
No experimento (Figura 4.1), Miller capturou os gases que Oparin propôs em sistema fechado. Os aminoácidos possuem quatro grupos químicos diferentes em sua estrutura, ligados a um átomo de carbono central ou carbono (Figura 4.2). iv) um grupo R, que varia de aminoácido para aminoácido. Como podemos ver na Figura 4.3, os aminoácidos possuem dois pólos: um com carga positiva e outro com carga negativa.
O gráfico da Figura 4.4 mostra uma curva ácido-base para um aminoácido. A Figura 4.4 mostra que em valores de pH zero ambos os grupos ionizáveis de glicina são totalmente protonados. A curva na Figura 4.4 mostra aminoácidos que não possuem grupo R ionizável, mas apenas grupos carboxila e amina ionizáveis.
De acordo com esta propriedade, os 20 aminoácidos essenciais são classificados da seguinte forma (Figura 4.6): i) aqueles que possuem o grupo R alifático apolar; ii) com o grupo R apolar aromático; iii) com grupo R polar não carregado; iv) com o grupo polar R com carga positiva ou negativa. As mãos têm duas propriedades muito importantes: (i) cada mão é a imagem espelhada da outra e (ii) essas imagens espelhadas não se sobrepõem (Figura 4.13). A primeira coisa que deve acontecer para que um aminoácido seja utilizado como combustível para gerar energia é a remoção do grupo amina (Figura 4.16).
Assim, o -cetoácido, que é o resultado da remoção de um grupo amino de um aminoácido, pode entrar nas vias de produção de energia como intermediários no ciclo do ácido cítrico (Figura 4.17). Todas essas sete moléculas entrarão no ciclo do ácido cítrico para produzir energia (Figura 4.17).

Os carboidratos e sua importância metabólica
Esta reação forma derivados chamados hemiacetais ou hemicetais (Figura 5.7), que possuem um carbono extra assimétrico e podem, portanto, existir em duas formas estereoisoméricas. De acordo com a Figura 5.7, para a forma cíclica da glicose, notamos que o grupo aldeído no carbono 1 e o grupo hidroxila no carbono 5 reagiram para formar o hemiacetal cíclico. Os dissacarídeos, como maltose, lactose e sacarose, consistem em dois monossacarídeos ligados covalentemente por uma ligação O-glicosídica, que é formada quando um grupo hidroxila em um açúcar reage com o carbono anomérico de outro açúcar (Figura 5.10).
Os carbonos anoméricos de ambos os monossacarídeos estão envolvidos em uma ligação glicosídica e, portanto, a sacarose não é um açúcar redutor (Figura 5.11). No final do processo, as células produzem duas moléculas de ATP, duas moléculas de NADH e duas moléculas de piruvato a partir de uma molécula de glicose (Figura 5.14). Existem também células em nosso corpo que podem fermentar a glicose, mas o produto resultante é o lactato (Figura 5.14).
Essas transferências são realizadas por diversas proteínas transportadoras de elétrons, na membrana mitocondrial interna, até o aceptor final, que é o O2 (Figura 5.15). A transferência de elétrons através de transportadores gera um gradiente eletroquímico de H+ através da membrana interna mitocondrial (Figura 5.16). O retorno do H+ ao interior mitocondrial, realizado através de uma proteína chamada F0/F1 ATP sintase, é o que permite a síntese do ATP (Figura 5.16).
É através deles que os elétrons migram de pares redox com potenciais de redução padrão mais negativos para potenciais mais positivos (Figura 5.17). Os carboidratos também podem ser encontrados na forma de unidades simples – monossacarídeos – ou combinados em outra unidade simples – dissacarídeos.

De lipídeos de reserva a combustíveis metabólicos
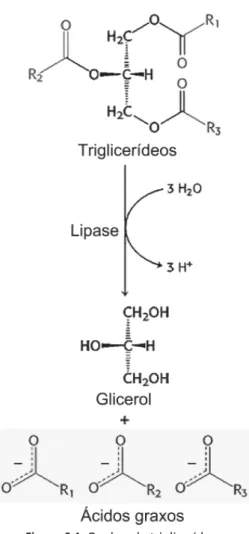
Para lembrar, os triglicerídeos são formados a partir de três cadeias de ácidos graxos (representados por R1, R2 e R3 na figura) ligadas à molécula central de glicerol. Os ácidos graxos e o glicerol absorvidos pelas células intestinais são reunidos e reconstituem os triglicerídeos. Os adipócitos absorverão ácidos graxos e glicerol do sangue e construirão triglicerídeos novamente.
Lembremos que as células do nosso corpo absorvem ácidos graxos na forma de ácidos graxos livres. A -oxidação dos ácidos graxos ocorre quando eles são absorvidos pelas células dos diversos tecidos que constituem o corpo. A segunda etapa na oxidação dos ácidos graxos é o transporte do acil-CoA para a mitocôndria.
Isso ocorre porque a carnitina é a substância que transporta os ácidos graxos ativados do citoplasma para o interior das mitocôndrias. Durante a hibernação, a oxidação dos ácidos graxos derivados da quebra dos triglicerídeos gera a energia (ATP) e a água necessárias à sobrevivência desses animais. No peroxissomo, o processo de degradação e oxidação dos ácidos graxos é semelhante ao que ocorre nas mitocôndrias.
A oxidação dos ácidos graxos leva à produção de corpos cetônicos para armazenar o excesso de acetil-CoA. Antes dos ácidos graxos serem transportados para as mitocôndrias, os ácidos graxos devem ser ativados.
Métodos de detecção de carboidratos,
Reação de Antrona
A reação de Antrona deve ser realizada com muito cuidado porque seu reagente contém antranol em meio concentrado de H2SO4. O hidroximetilfurfural, a partir da desidratação das hexoses, produz uma substância verde-azulada, relativamente estável; enquanto o furfural, a partir da desidratação das pentoses, dá um produto de cor semelhante, mas que se torna amarelado ou vermelho rubi alguns minutos depois (Figura 7.8). O teste Antrona é muito sensível e pode ser usado com soluções padrão de carboidratos para quantificar carboidratos.
Os resíduos de palha e cana-de-açúcar representam uma fonte abundante, barata e prontamente disponível de biomassa lignocelulósica renovável. Para transformar essa biomassa em etanol, a celulose deve estar acessível às enzimas que degradam a celulose. A lignina, por outro lado, forma uma camada protetora para a celulose e a hemicelulose, protegendo os polissacarídeos da degradação enzimática.
Portanto, é fundamental remover a lignina, tornando a celulose vulnerável a enzimas, permitindo que a levedura, um microrganismo, converta a glicose em etanol durante a fermentação. Contudo, durante esta hidrólise podem surgir derivados indesejados para o posterior processo de fermentação da levedura. Os derivados de furano, furfural e 5-hidroximetilfurfural, que são produzidos principalmente durante a desidratação de pentoses e hexoses durante a hidrólise em hidrolisados lignocelulósicos, estão entre os inibidores mais fortes do processo de fermentação.
Assim, a reduzida capacidade da levedura em realizar o processo de produção de etanol a partir de material lignocelulósico está relacionada principalmente às altas concentrações de furfural e 5-hidroximetilfurfural. Portanto, apesar de desejarmos aumentar a produção de etanol combustível através da utilização de biomassa lignocelulósica, é de grande importância o desenvolvimento de novos métodos de remoção de lignina, sem produção de derivados tóxicos.
Reação de Seliwanoff
Reação de Bial
O método do Biureto é amplamente utilizado para determinar a concentração de proteínas em vários materiais biológicos, como soro ou plasma sanguíneo, líquido cefalorraquidiano (LCR), urina, alimentos, saliva, fibrinogênio e tecido animal. Apesar de ser rápida e utilizar reagentes de baixo custo, esta metodologia apresenta baixa sensibilidade em comparação com outros métodos de quantificação da concentração proteica. Um dos métodos colorimétricos mais utilizados na quantificação de proteínas é o método Folin-Lowry, que acopla o método Biuret a uma redução do ácido fosfomolíbdico fosfotúngstico (reagente Ciocalteau).
O método Lowry tem alta sensibilidade a seu favor e, portanto, é usado para determinar a concentração de proteína. A identificação e determinação de proteínas pelo método Bradford é um método que utiliza o corante azul Coomassie (BG-250). Os grupos amino e R (cadeia lateral) podem sofrer reações químicas (exemplo: ninidrina, Sakaguchi, Pauly, nitroprussiato) que permitem a sua identificação.
Por outro lado, os hidratos de carbono possuem propriedades (desidratação e poder redutor) que permitem a sua identificação e quantificação. As proteínas, apesar de muito diferentes entre si, possuem peculiaridades químicas que possibilitaram o desenvolvimento de diferentes metodologias de identificação e quantificação. Os grupos amino e R são grupos escolhidos para identificação e quantificação de aminoácidos.
O DNS determinaria a concentração de açúcar na amostra, enquanto o reagente Biureto determinaria a quantidade de proteínas. As propriedades dos carboidratos de sofrer desidratação e sua capacidade de reduzir alguns sais (como o cobre) têm sido estudadas há muitos anos para identificar e quantificar esses nutrientes.

UFRJ










