Composição genética das populações do Sudeste Asiático: uma análise dos principais haplogrupos da região e uma contribuição para a compreensão das teorias sobre rotas migratórias / Fernanda Mathias de Miranda Barros. Composição genética das populações do Sudeste Asiático: uma análise dos principais haplogrupos da região e uma contribuição para a compreensão das teorias das rotas migratórias.
Migrações humanas para o Sudeste Asiático
Legenda: as setas vermelhas representam o caminho ao longo da costa do Sudeste Asiático e as setas azuis a dispersão em direção ao interior do continente. Finalmente, existe uma hipótese de migração mais complexa que propõe que ocorreram duas ondas descritas na população insular do Sudeste Asiático, em vez de apenas uma delas (JINAM et al., 2012).

Filipinas
Apesar da existência de outras línguas anteriores nas ilhas do Sudeste Asiático, o austronésio tornou-se dominante enquanto as línguas ancestrais das ilhas desapareceram. A família austronésica possui cerca de 1200 línguas e está amplamente representada em diversas ilhas do Sudeste Asiático (Figura 6), incluindo as Filipinas.

Vietnã
De acordo com o Censo Populacional e Habitacional do Vietname de 2009 (http://www.gso.gov.vn), este grupo étnico tem uma população de indivíduos que representa 85,7% da população total do Vietname. Os Tay são o segundo maior grupo étnico do Vietnã, com uma população de 1.626.392 pessoas (http://www.gso.gov.vn), a maioria das quais vive nas províncias montanhosas ao norte do Rio Vermelho (Grupos Étnicos no Vietnã Nam: Uma análise dos principais indicadores do Censo Populacional e Habitacional do Vietnã de 2009 / UNFPA, 2011).

Marcadores genéticos
STRs (Short Tandem Repeats), também conhecidos como microssatélites, são caracterizados por um menor número de bases formando o motivo repetitivo (geralmente 2–6 pb) (GYMREK et al., 2017). InDels (Polimorfismos de deleção de inserção) são variações de comprimento (na ordem de unidades) presentes no genoma, geradas por inserções ou deleções de nucleotídeos (LAPPALAINEN et al., 2019).
Cromossomo Y
Porém, com a descoberta de alta recombinação intracromossômica em NRY (SKALETSKY et al., 2003), um nome mais apropriado foi proposto: região específica do sexo masculino do cromossomo Y (MSY). Em estudos de genética populacional, os “marcadores do cromossomo Y” são atribuídos especificamente àqueles localizados na região MSY, excluindo aqueles em regiões pseudoautossômicas (PARs), que apresentam homologia com o cromossomo X (JORDE et al., 2000; UNDERHILL et al., 2001). Esses Multiplexes são encontrados na forma de kits, sendo utilizados em diferentes estratégias e diferenciados de acordo com o número de marcadores (Figura 10) (BELEZA et al., 2003; BUTLER, 2014; LI, 2015).
Portanto, é bastante aplicável a amostras do continente europeu, mas ainda não foi abordado no continente asiático (LESSIG et al., 2003). Desde então, milhares de outros polimorfismos bialélicos foram descobertos, com mais duas letras (S e T) adicionadas em 2008 (Tabela 2) (KARFET et al., 2008).

Objetivo geral
Objetivos específicos
Analisar os resultados obtidos em relação às linhagens masculinas presentes nas populações do Sudeste Asiático, levando em consideração diferentes hipóteses sobre a colonização desta região, a fim de esclarecer aspectos relacionados à história dessas populações.
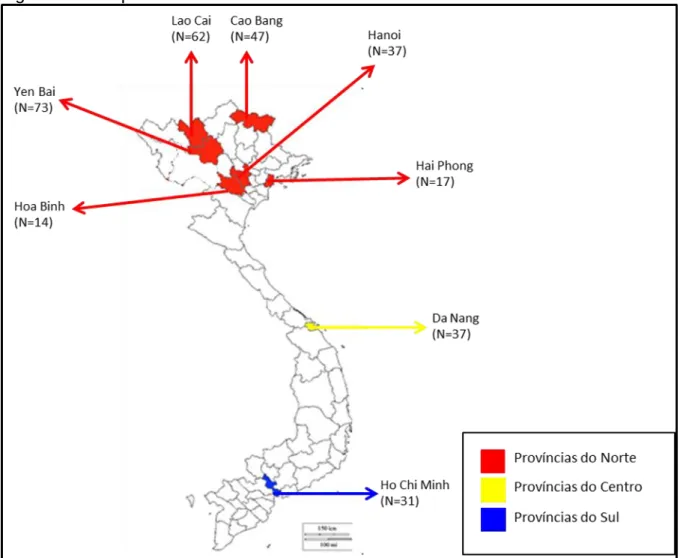
Amostragem e obtenção das amostras de DNA
Legenda: Em destaque estão as províncias utilizadas neste trabalho, bem como seus respectivos números amostrais. As amostras do Vietname foram obtidas numa colaboração já existente com os investigadores Carlos Vullo (Laboratorio de Genética Forense, Equipo Argentino de Antropología Forense_EAAF), Nguyen Duc Nhu (Instituto de Medicina Forense do Ministério da Saúde de Hanói, Vietname) e Tran Duc Phan. (Instituto de Medicina Legal, Ministério da Saúde, Hanói, Vietnã). As amostras foram coletadas mediante consentimento informado e enviadas à LDD-UERJ de forma criptografada, após retirada dos dados pessoais.
Extração de DNA
A extração de DNA das amostras filipinas foi realizada utilizando o método de extração orgânica (fenol/clorofórmio) por funcionários da Faculdade de Ciências Biológicas da Universidade Complutense de Madrid, Espanha. Nas amostras vietnamitas, a extração foi realizada pelo método de extração com fenol/clorofórmio orgânico ou pelo método Chelex®. Na extração com fenol/clorofórmio, 1 µl de 1x SSC (sal-citrato de sódio) foi adicionado ao tubo contendo a mancha de sangue e após 15 minutos o papel foi macerado para liberar o sangue nele contido.
No dia seguinte, foram adicionados 200 µL de Fenol/Clorofórmio/Isoamil (25:24:1) para separar as fases orgânica e aquosa. Como esta imersão em água não é suficiente para remover o sangue contido no papel, foram adicionados 400 µL de tampão de extração (citado acima no protocolo fenol/clorofórmio) e incubados em temperatura ambiente por 30 minutos.
Genotipagem de marcadores Y-STRs
Genotipagem de Y-SNPs
Protocolos utilizados com Multiplex Um e Dois
- Seleção das sequências alvo e desenho dos primers
- Multiplex Um e Dois
- Análise estatística dos resultados
O programa Primer3 (//primer3.ut.ee/) foi utilizado para seleção de PCR e sequenciamento de primers. Após a seleção dos primers para a construção do Multiplex 1 e 2, a ausência de dímeros e grampos foi verificada utilizando o programa AutoDimer (https://strbase.nist.gov//AutoDimerHomepage/AutoDimerProgramHomepage.htm). Em ambos os casos, as reações de PCR foram realizadas em um volume final de 5 µL contendo 2,5 µL da mistura mestre 2X incluída no kit QIAGEN Multiplex PCR, 0,5 µL da mistura de primers 10X (concentração de 2 µM de cada primer direto e reverso) , 1 μL de água milliQ e 1 μL de DNA a 2 μM.
Antes do sequenciamento com SNaPshot, os produtos de PCR foram purificados adicionando 0,5 µl da enzima ExoSAP_itTM (ThermoFisher Scientific) a 1 µl de produto amplificado e incubando por 15 minutos a 37ºC, seguido de 15 minutos de desnaturação a 85ºC em termociclador. Após a reação SNaPshot, a purificação foi realizada adicionando 1 µl de SAP (fosfatase alcalina de camarão) ao produto final por 1 hora a 37ºC e 15 minutos a 85ºC.
Composição genética do Vietnã
Análise da subestrutura genética das províncias
Após aplicar a correção de Bonferroni para testes múltiplos (nível de significância de 0,00179), a província de Yen Bai mostrou uma distância genética significativa de Cao Bang nas análises Y-STR usando FST. No entanto, esta distância genética não foi significativa para análises de Y-STRs com RST e Y-SNPs, sugerindo a necessidade de aumentar o número de amostras de Yen Bai. Lao Cai apresentou uma distância genética elevada, na ordem de 10 a 37% (Tabela 2), em comparação com as demais províncias, sendo esta distância significativa nas análises para Y-STRs com RST (exceto Hai Phong).
Assim, esse isolamento geográfico dos Mong contribuiu para a grande distância genética entre eles e os demais grupos étnicos estudados. As amostras deste estudo também foram comparadas com outras da literatura (PURPS et al., 2014; TREJAUT et al., 2014) e não foi observada distância genética significativa entre elas.

Análise da diversidade das províncias
Este resultado confirma os resultados de frequência de haplogrupos, portanto há uma diferença genética entre Lao Cai e outras províncias. Ao correlacionar os marcadores com os grupos étnicos, o subhaplogrupo O2-M95 permaneceu o mais comum em todos os grupos étnicos, exceto na etnia Mong, onde o subhaplogrupo O3-P201 foi o mais comum, confirmando novamente a diferença entre os Mong e outras etnias. Devido aos elevados valores da diversidade de haplótipos identificados, a hipótese de deriva genética foi descartada na maioria dos grupos estudados.
No entanto, a diversidade de haplótipos foi ligeiramente menor nas amostras de Lao Cai e Yen Bai. O número de haplótipos partilhados nas populações destas províncias também foi elevado, sendo 35,5% em Yen Bai, 45,1% em Lao Cai comparado com apenas 14% em Cao Bang e zero nas restantes.
Análise da subestrutura genética por etnias
A etnia Mong apresentou uma alta distância genética (da ordem de um nível de significância de 0,00139 (após aplicação da correção de Bonferroni para testes múltiplos)) com todas as etnias nas análises do FST, exceto tailandês (Tabela 4). Com base na classificação linguística e na distância genética resultados, Kinh e Muong poderiam ser agrupados com Thai, Tay, Nung e Cao Lan, pois não apresentam diferenças significativas entre eles. Portanto, alguns grupos separados por classificação linguística poderiam ser geneticamente unidos, e outros que foram classificados como o mesmo grupo , eles seriam separados.
Nota: Acima da diagonal (*) estão os valores de probabilidade de não diferenciação e abaixo deles estão os valores de FST e RST. Com nível de significância de 0,00139, as amostras que apresentaram distância genética significativa estão destacadas em vermelho e as amostras que apresentaram alto valor de distância genética estão destacadas em laranja.
Composição genética das Filipinas
Análise da subestrutura genética
O resultado mostrou em todas as análises que não há distância genética significativa (p<0,01666) para os grupos de ilhas Filipinas nas amostras deste trabalho, confirmando o resultado anterior. Isso permite uni-los para análises comparativas com outras amostras das Filipinas e até mesmo com outros países do Sudeste Asiático. O segundo subhaplogrupo mais comum nas amostras foi O1a*-M119, com frequência de 21% em Luzon e 28,9% em Mindanao/Visayas.
Nas amostras filipinas estudadas por Trejaut et al.(TREJAUT et al., 2014) este foi o sub-haplogrupo mais frequente. Neste trabalho, foi encontrada uma frequência menor deste sub-haplogrupo em Luzon do que a descrita por Trejaut e colegas (TREJAUT et al., 2014).

Comparação com outras amostras das Filipinas na literatura
- Comparação com 21 Y-STRs
- Comparação com 17 Y-STRs
O subhaplogrupo O1a*-M119 representa altas frequências em todas as populações do Sudeste Asiático (com exceção da Indochina) e tem sido amplamente citado em estudos de expansão austronésica anteriores à entrada na Polinésia, uma vez que nesta região a frequência deste haplogrupo diminuiu , mostrando-se baixo (MIRABAL et al., 2012). Na análise de distância genética do FST, os valores de probabilidade de não diferenciação foram estatisticamente significativos apenas entre as amostras de Luzon estudadas por Trejaut et al.(TREJAUT et al., 2014) e as demais amostras das Filipinas. Legenda: FST da distância genética entre amostras da população filipina (abaixo da diagonal) e o correspondente valor de probabilidade de não diferenciação (acima da diagonal).
Legenda: Comparação pareada de distâncias genéticas RST entre amostras da população filipina (abaixo da diagonal) e valor de p correspondente à não diferenciação (acima da diagonal) (a) Purps e colegas (PURPS et al., 2014); (b) estudo atual; (c) Trejaut e colegas (TREJAUT et al., 2014).; Número de permutações: 50175; s.e.

Comparação entre as populações do Sudeste da Ásia
- Análise das rotas de migração continentais
- Escalonamento multidimensional entre as amostras populacionais das ilhas
Portanto, estes resultados são consistentes com a teoria da existência de duas rotas migratórias no Sudeste Asiático. O Vietnã e as Filipinas apresentaram um perfil típico do Sudeste Asiático, já que o haplogrupo O foi o mais frequente em ambos. 34;Tamanho pequeno" no registro fóssil humano das Filipinas: é importante para uma melhor compreensão da história evolutiva dos Negritos.
Every microsatellite is different: The intrinsic properties of DNA dictate the mutagenesis of common microsatellites present in the human genome. Presidential Address: "The Peoples of Asia"—Science and Policy in the Classification of Ethnic Groups in Thailand, China, and Vietnam. Investigation of the HUMTH01 STR locus using PCR and two electrophoresis formats: UK and Galician Caucasian population surveys and utility in paternity investigations.
Structure: a comparison of the spatial patterns of home gardens of different ethnic groups in Thailand and Vietnam.







