Functionally highly diverse enzymes that modify fatty acyl cells from insects, one of the most common animals in the world, represent an important resource for modern cell factory biotechnology. We then functionally characterized the corresponding enzymes in yeast and assessed their participation in the biosynthesis of fatty alcohols observed in the pheromone-producing marker tissue. In addition, we studied the effect of different residues in the substrate tunnel on the specificity of horn FADs.
We exchanged the polar residues in PTC for their nonpolar homologues from soluble methane monooxygenase (MMO). When talking about individual lipid compounds of fatty acyl origin, the abbreviation consists of the number of carbon atoms in the fatty chain followed by a colon (eg 14: for tetradecanoyl/tetradecyl).

Sequence and structure
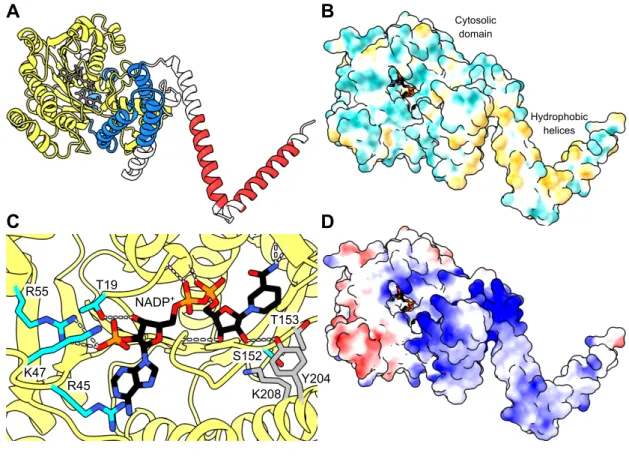
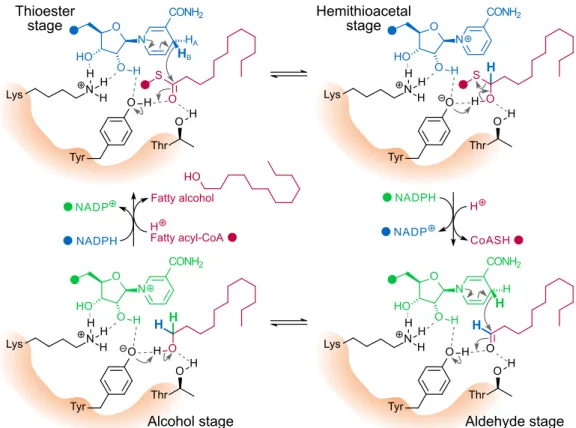
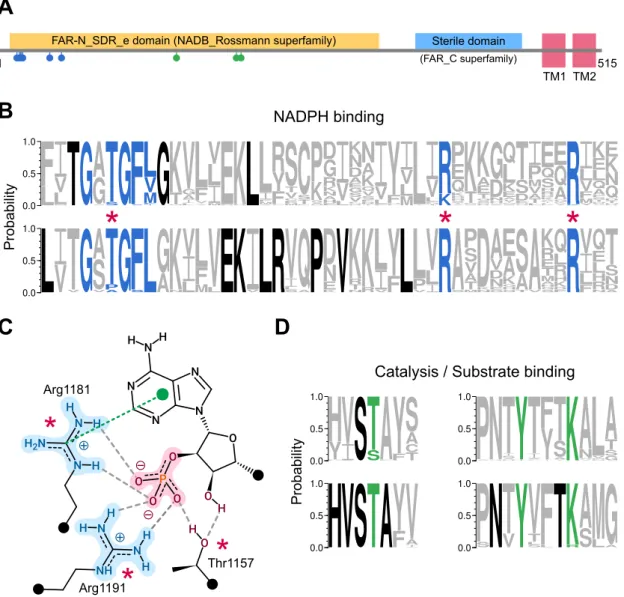
In the models, the C-terminal region coils with part of the Rossmann fold, potentially forming the substrate-binding cavity of the enzyme. A detailed overview of the putative active site of FAR2 based on the structure of the SauMxaA-R active site. There is no information on the binding pattern of the fatty acyl-CoA substrate to the enzyme.
However, the predominantly electropositive portion of the Sterile domain may play a role in such binding ( Fig. 3D ), or may mediate interaction with the negatively charged phospholipid membrane. A decrease in activity occurs when the critical micellar concentration of the substrate is exceeded, suggesting that the enzyme uses only the free form of the fatty acyl.
Function and engineering
In flagellate Euglena gracilis, the co-expression of its FAR and wax synthase in yeast results in the production of medium chain wax esters (C28-C30) [89]. Since then they have been studied extensively, mainly in moths, because their pheromone is composed of oxygenated compounds derived from fatty acid reduction - the alcohols, aldehydes and acetates. In silk moth (Bombyx mori), a pheromone gland (pg)-specific FAR is able to convert 10E12Z-16:2 precursor into the corresponding alcohol bombykol [30], the main component of the female sexual pheromone.
One FAR putatively involved in the biosynthesis of fatty alcohol precursors to wax was identified in Drosophila [91], and another six FARs were found in the honey bee (Apis mellifera) [35]. The honey bee FAR1, which is preferentially expressed in the head, has been shown to reduce hydroxylated fatty acyl precursors apart from the common saturated C16-C22 substrates.
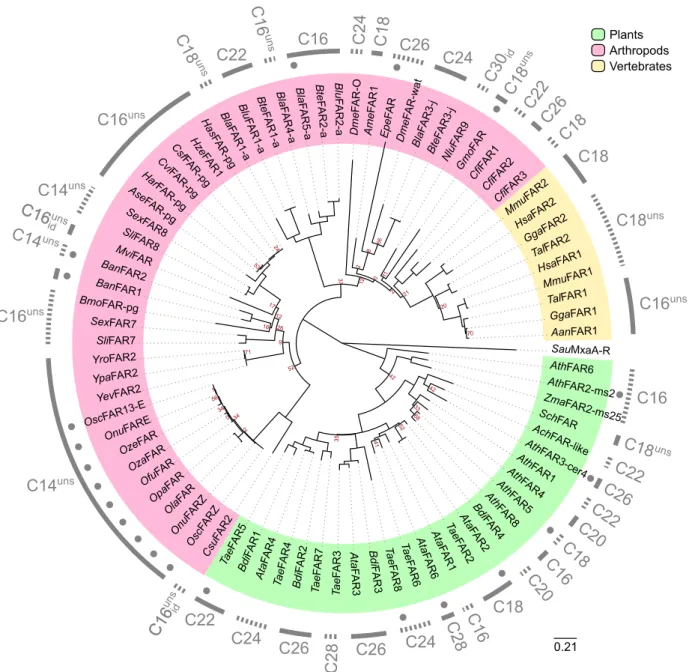
Fatty acyl desaturases
Fatty acyl-CoA desaturases
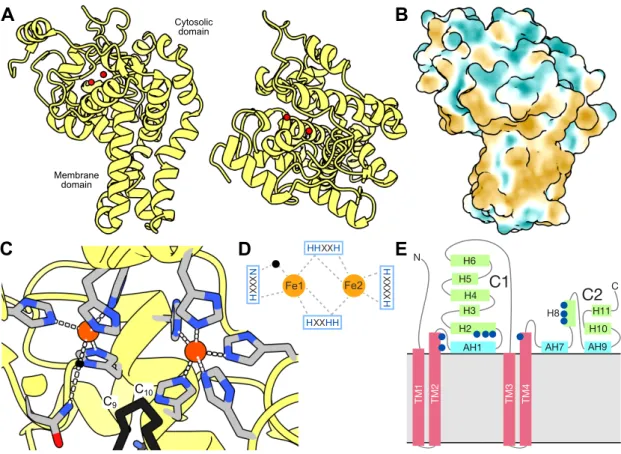
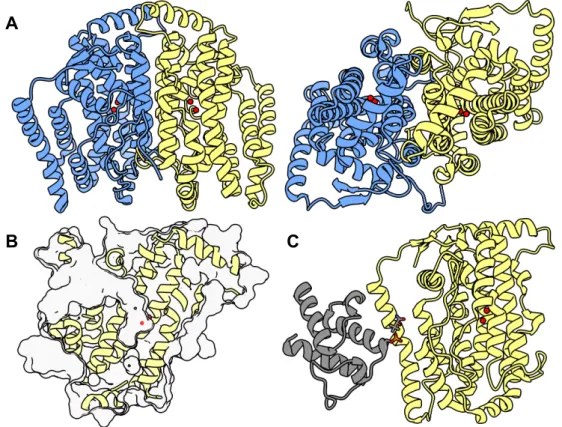
A narrow tunnel starts at the surface of the cytosolic domain and continues deeper into the protein (Fig. 9A). In the structures, both metals are approx. 5 Å apart from C9 and C10 atoms of the fatty acyl-CoA substrate, i.e. the coenzyme A portion of the substrate interacts with the predominantly electropositive surface adjacent to the tunnel entrance (Fig. 9B) [120,137].
The substrate tunnel accommodates the acyl fatty chain of the substrate, presumably by the hydrophobic effect [120,137]. In the canonical Δ9 desaturases, the acyl of the substrate forms a hydrogen bond with a tryptophan residue at the beginning of the substrate tunnel ( Fig. 9E ) which acts as an acyl register.
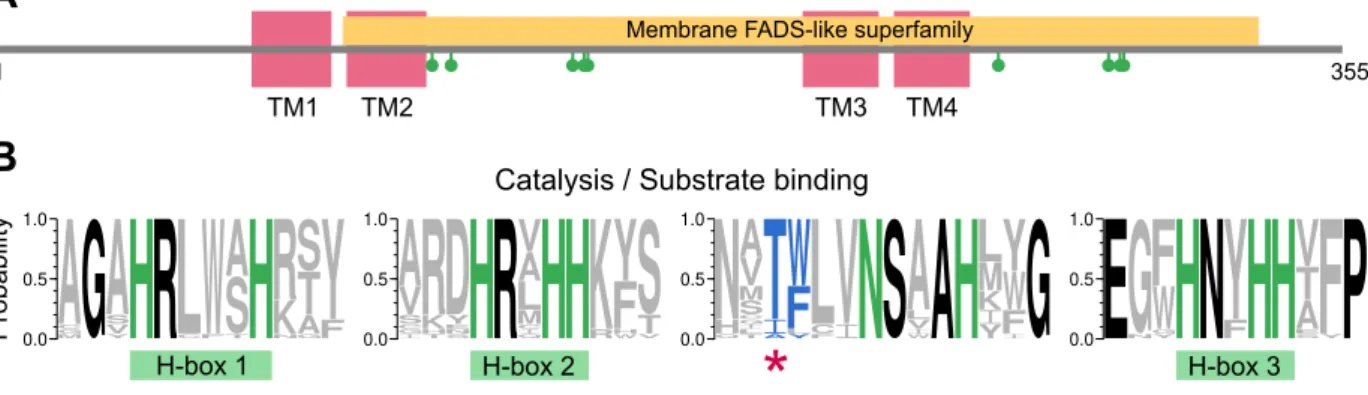
Fatty acyl-ACP desaturases
The fatty acyl is attached to the 4'-phosphopantetheine moiety, which is attached to the serine residue of ACP. Fdis of reduced ferredoxin (two molecules). The ACP portion of the substrate interacts with the outer surface of sFAD, thus keeping the fatty acyl chain in place. The reaction cycle is first initiated by the binding of the fatty acyl-ACP substrate, followed by the binding of reduced ferredoxin [188].
Part of the phosphopantetheine group at the interface of desaturase and ACP, carrying the fatty acyl chain (not visible in this crystal structure), is depicted as sticks. Similar biochemical reactions are usually catalyzed by members of the same superfamily of enzymes (reviewed in [18,208]).
Publication II: A single specificity switch in moth desaturases
In other words, they show 11Zdesaturase and 10,12-conjugase activities on 16: and 11Z-16: substrates, respectively (i.e. the native specificity of MseD2), with less bulky residues alanine and threonine (Fig. 17A). In contrast, with the increase in side chain volume to valine and isoleucine, the enzyme specificity gradually shifts to Δ14-desaturase activity which manifests on 10E12E-16: substrate as a production of trines (i.e. the specificity inherent to MseD3). In this case, we also noticed that the enzymatic specificities among the studied enzymes are more relaxed and broader when it comes to variable residues at position 224 (Fig. 17A).
For the C14 substrate, desaturation also occurs with glycine residues (in contrast to the Δ14 desaturation of the C16 substrate, Fig. 17A) or phenylalanine (in contrast to the Δ11 desaturation / 10,12-conjugation of C16 substrates). When we mutated Thr227 in BmoD1 (homologous to position 224 in MseD2/D3) to the larger isoleucine present in wild-type MseD3, we observed a decrease in 11Z-16: production and a complete loss of conjugase activity (i.e. diene17 enzyme production) in Fig. The complete absence of MseD3 simulations did not pose a problem as both MseD2 and MseD3 share similar functional patterns in all mutations (Fig. 17A) and they probably behave similarly at the MD level.
First, we simulated complexes of MseD2 and five mutants at position 224 (glycine, threonine, valine, isoleucine and phenylalanine) with 16:CoA, which is a native substrate for the A11 desaturation reaction catalyzed by MseD2. We found the highest number of contacts (more than 1,000) for wild-type MseD2 and MseD2-224T mutant (Fig. 17B), followed by MseD2-224V, which is consistent with the data from functional characterization in yeast (Fig. 17A); we did not detect contacts in glycine, isoleucine and phenylalanine mutants. In addition, the enzymes with alanine, threonine, valine and isoleucine at position 224 show lower mean distances (Fig. 17B) and lower standard deviations of distances between zinc and C11-C12 binding, in contrast to the MseD2-224F mutant.
Second, we studied the complexes of MseD2 and five position 224 mutants (see above) with 10E12E-16:CoA which is a native substrate used by MseD3 during Δ14 saturation reaction. MseD2- 224V shows the highest number of contacts (above 10,000) followed by glycolinF, followed by much lower number and 000. 17B); the C14-C15 bond of 10E12E-16:CoA exhibits no contact with active-site zinc ions in MseD2, and MseD2-224T and -224F mutants. Similar to 16:CoA substrate, the mutantMseD2-224F manifests the largest distance between the active site and the substrate C-C bond during the simulations. The conversion is based on a change in side chain volume of a single amino acid at the substrate tunnel bend (Fig. 17C).
Publication III: Proton transfer chain disruption in castor desaturase
The amino acid residues in MMO at positions structurally homologous to PTC A in Δ9D are quite non-polar (isoleucine, valine, phenylalanine) [232], so they can significantly hinder the proton transfer efficiency to the active site. We found that all the desaturases are active (Fig. 18C), they 9Z-desaturate 18:CoA to oleoyl-CoA, but the accumulation of PTC-disrupting mutations gradually reduces the activity, i.e. 8 (facing page): Impacts of proton transfer chain mutations in a soluble desaturase A) Detailed view of the Δ9D active site (PDB 1afr [163]) with proposed proton transfer chains (PTCs), A (light green) and B (light purple), which can direct protons to it during catalysis [191].
The residues constituting the PTCs are colored yellow, the residues chelating the active site iron ions (red) are colored pink, except for the proton transfer target residue (red). B) The shared architecture of the active site with four helix bundles in NHFe2 proteins: soluble Δ9desaturase from R. Production was quantified by GC-MS of derivatized reaction mixtures. wild-type A9D), which were identified by synthetic standards as a co-eluting mixture of mainly 9-, 10- and 11-monohydroxyl-18:-acyl groups (Fig. 18C). The hydroxylation activity is much lower than the desaturation activity in absolute numbers, the most active mutant Δ9D (hydroxylation-wise) produces almost 0.3%, but when we calculate the ratio between hydroxylation and desaturation, there is a clear shift in specificity from almost exclusively desaturation-specific wild-type Δ9D to more hydroxylation-sensitive mutant Δ9D-101I203I206V22 2F. (Fig. 18C; there is an almost 100-fold difference in absolute values between the wild-type and the 4-fold mutant).
We found that wild-type A9D and the mutants exhibit activities against dihydroxylation and epoxidation (Fig. 18D). Because the diol, epoxide, and allyl alcohol products were consistently present in reaction mixtures mainly as a result of oleoyl-CoA oxidation, we decided to investigate the ability of the desaturase to secondarily desaturate and/or hydroxylate the already hydroxylated products, 9-, 10-, or 11-OH-18:CoA. In contrast, with 11-OH-18:CoA acting as a substrate, we detected both diol and allyl alcohol products in all enzymes [234].
On the other hand, the diols are represented by two products, erythro-entreo-9,11-diOH-18: acyls (i.e. the desaturase introduced a new hydroxyl group at C9), and their production is increased in mutant enzymes compared to wild type. Finally, the disruption of PTC A in Δ9D by mutagenesis does not completely shut down the desaturation channel. This may be due to the existence of partial PTC B or due to the access of protons to the active site from other sources.
Publication IV: Insects as a source of enzymes for lipid biotechnology?
In general, LG fatty alcohols partially correspond to the specificity determined for the abundantly expressed FARs, i.e., the phylogenetic analysis shows an expansion of the FAR-A group only among species from small bees (Bombini) and stingless bees (Meliponini). Notably, transposable elements have been detected specifically in the vicinity of FAR-A genes in the two bee genomes.
Thus, the TE-mediated duplications could play an important role in FAR evolution in Hymenoptera. The effects of introducing identical residues at position 224 in Manduca sexta desaturases D2 and D3 on their specificity are similar. The switching mechanism between Δ11/conjugase specificity observed in wild-type D2 and Δ14 specificity observed in D3 is most likely determined by residue size and hydrophobic effect.
After the introduction of larger and more hydrophobic residues, valine and isoleucine, the specificity of both enzymes shifts towards Δ14desaturation of the substrate 10E12E-16:. Interestingly, in the Δ11desaturase/conjugase D1 of the evolutionarily more distant lepidopteran, Bombyx mori, the specificity derived from the residues of the tunnel bend that goes from Δ11/conjugase (with a threonine residue) to Δ14 (with an isoleucine residue) is preserved. Disruption of the proton transfer chain (PTC) in soluble Δ9desaturase from Ricinus communis by introducing homologous residues from methane monooxygenase, which has a structurally similar active site architecture, leads to increased substrate hydroxylation.
The desaturase accepts not only saturated and monounsaturated fatty acyls as substrates, but also monohydroxylated fatty acyls. The effect of PTC disruption can be explained by limited access of protons to the active site necessary for the release of water during the desaturation reaction.