We have observed that rhein inhibited RA-induced differentiation of SH-SY5Y cells and that 24 h treatment with rhein induced profound changes in the cellular proteome. SH-SY5Y cells were seeded in six-well plates and cultured for 6 days in the absence or presence of RA. Differentiation of SH-SY5Y cells to a neuronal phenotype was induced by RA following the previously described protocol (Ammer and Schulz 1994).
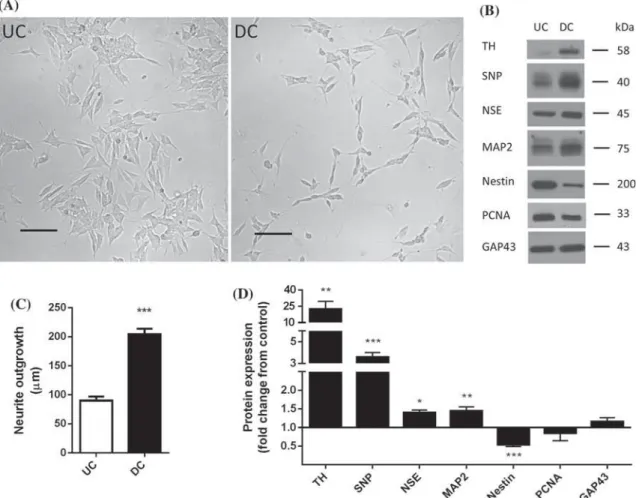
Prolonged exposure of SH-SY5Y cells to RA led to neuronal differentiation and most cells adopted a typical neuronal morphology. Our first set of proteomic experiments was designed to assess changes in the proteome of SH-SY5Y cells exposed to 10 lM RA for 6 days. In the next set of experiments, we explored the effect of 24 h treatment with 10 lM rhein on protein expression in UC and RA-DC SH-SY5Y cells.
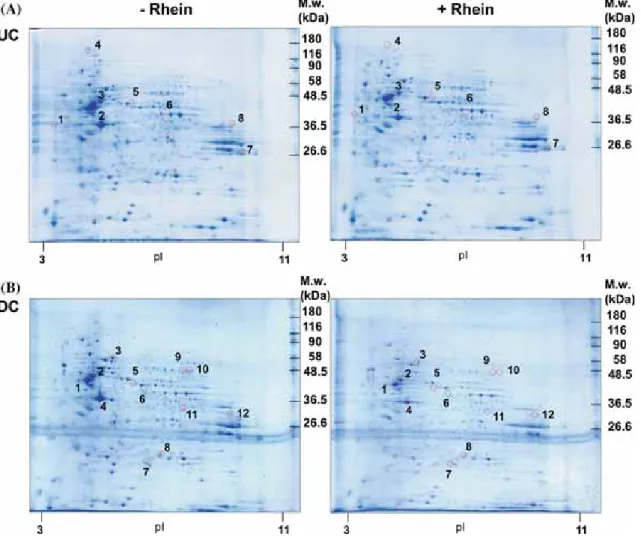
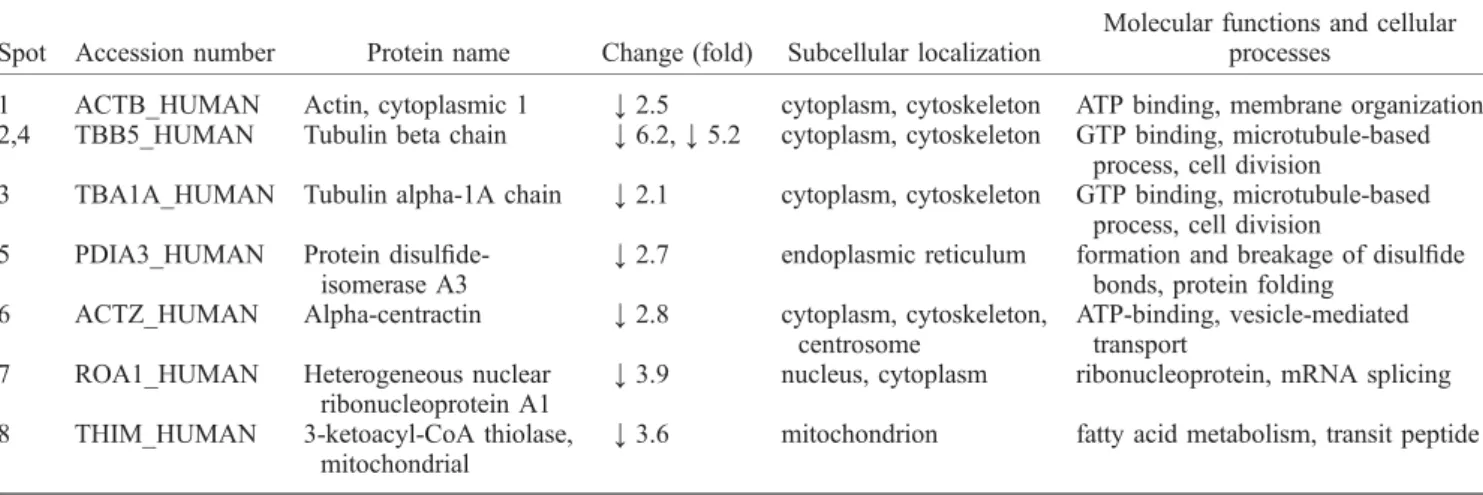
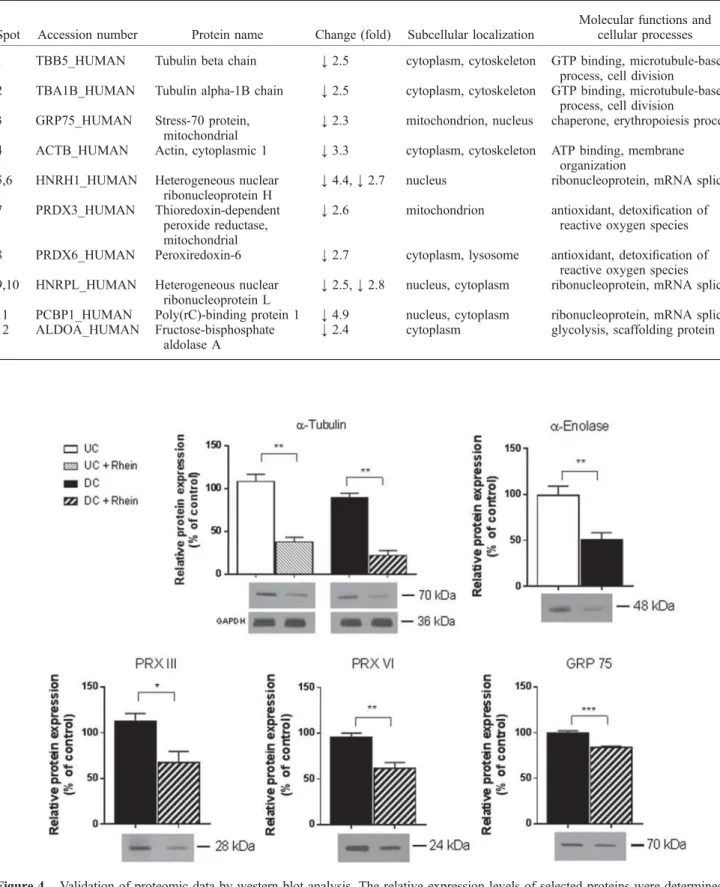
The comparative proteomic analysis indicated that rhein affected the expression of several proteins in UC and DC SH-SY5Y cells. The results of these experiments indicated that RA-induced differentiation of SH-SY5Y cells and/or treatment of these cells with rhein was associated with identical trends (downregulation) in the expression of these proteins. Rhein markedly reduced (by approximately 70%) atubulin expression in both UC and DC SH-SY5Y cells.
Effects of long-term RA treatment on the cellular morphology and expression of neuronal differentiation markers in SH-SY5Y cells. Subcellular localization and biological functions of proteins altered by rhein in undifferentiated SH-SY5Y cells. Subcellular localization and biological functions of proteins altered by rhein in neuronally differentiated SH-SY5Y cells.
The relative expression levels of selected proteins were determined in samples from both UC and DC SH-SY5Y cells unaffected by rhein. Effect of long-term rhein treatment on α-enolase expression and activity in SH-SY5Y cells. Untreated (UC and RA) DC SH-SY5Y cells were prepared as described in Materials and Methods.
Our analysis identified a total of 15 down-regulated proteins in samples from SH-SY5Y cells treated with rhein for 24 h. In the present study we observed that nestin expression was reduced in RA-DC SH-SY5Y cells and that the presence of rhein prevented this change. The present study aimed to compare the proteomic profiles of SH-SY5Y cells exposed to rhein.
Ammer H and Schulz R 1994 Retinoic acid-induced differentiation of human neuroblastoma SH-SY5Y cells is associated with changes in protein abundance G.J.
DISCUSSION
On the other hand, the FTO inhibitor MO-I-500 did not affect the expression of these proteins and did not reverse the STZ effect (Supplementary Figure S4). Here, we observed that pretreatment of cells with the FTO inhibitor MO-I-500 did not result in increased AMPK phosphorylation, which was the case in the STZ group. Their results suggested that at a biochemical level, the apoptosis and cellular stress response to STZ proceeds in a relatively uniform manner, regardless of cell type. 25 Here, we found that the cytotoxic effect of STZ in CCF-STTG1 cells is associated with impaired metabolism. of energy, mitochondrial damage, elevated ROS, astrocyte reactivation, and apoptotic and necrotic cell death.
RNA methylation and neurodegenerative diseases, including AD.26−30 In genome-wide association studies, a gene polymorphism in the m6A FTO demethylase has been reported to be a risk factor in AD.31,32m6A RNA methylation is elevated in the cortex and hippocampus of a transgenic AD mouse model. The expression of the m6A methyltransferase METTL3 was increased and the m6A demethylase FTO was decreased in AD mice.30 Altered expression of METTL3 in AD was described in another study.29 Interestingly, METTL3 was down-regulated in human AD brain after death. In contrast to both studies, our study shows that in STZ-treated human astrocytoma cells, FTO protein expression is elevated, while METTL3 remains unaffected.
The function of FTO in relation to AD was further investigated in neurons isolated from 3xTg AD mice. In their work, FTO knockdown reduced the levels of tau phosphorylation (Ser 262), whereas FTO overexpression resulted in the opposite effect. apparently missing. Interestingly, inhibition of FTO led to lower GFAP levels along with reduced oxidative stress and generally mitigated the cytotoxic effect of STZ.
Interestingly, the addition of FTO inhibitor to warm astrocytes caused a dramatic increase in mitochondrial mass and was followed by increased ATP levels. In the work of Wang and colleagues, muscle cells deprived of FTO expressed reduced levels of PGC-1α, which was further accompanied by reduced mitochondrial mass and mtDNA content.44 Another study showed that overexpression of FTO up-regulation of PGC-1α induced through the direct influence of the m6A level of PGC-1α transcription.45 Among other factors, insulin action may be an important contributor to the modified mitochondrial metabolism via mTOR signaling. Numerous studies have indicated the important role of AMPK activation in the pathogenesis of AD, but whether its effects are beneficial or detrimental remains controversial.
Several groups found reduced AMPK activity in the brains of ICV-STZ rats.46,47 However, some researchers reported conflicting results.48 Our findings showed that exposure to STZ increased the phosphorylation level of AMPK. We believe that in our case AMPK could be activated in response to starvation or oxidative stress of cells, but this activation was insufficient to elicit the protective effect against STZ-induced damage. Together, these results are consistent with the hypothesis that FTO could participate in the control of energy homeostasis.
CONCLUSION
Cell culture. The human astrocytoma cell line CCF-STTG1 was purchased from the European Collection of Authenticated Cell Cultures (ECACC). Cells were treated with the inhibitor 24 h before the addition of STZ and then incubated in the presence of both drugs for the next 48 h. After treatment, the culture medium was replaced with fresh DMEM supplemented with 0.5% FBS, and cells were incubated at 37°C in the presence of 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) at a concentration of 0.2 mg/ml for 3 hours.
Cell cytotoxicity assay. The integrity of the plasma membrane of dead cells is impaired, which leads to the release of cytoplasmic lactate dehydrogenase (LDH) into the environment. Detection of apoptosis. The degree of apoptosis or necrosis was assessed by Annexin V/Hoechst 33258 staining, followed by flow cytometric analysis. Samples were then treated with 5 μL of Dyomics 647-labeled Annexin V and incubated in the dark for 30 min on ice.
Approximately 105 experimentally treated cells in wells of a 24-well microplate were incubated with both MitoTracker probes (40 nM) in the dark for 30 min at 37°C. For measurement of maximal respiratory rate, cells were treated with 1.5μM carbonyl cyanide-4-(trifluoromethoxy)phenylhydrazone (FCCP) before the addition of oil. Measurement of intracellular ATP. Intracellular ATP content was determined according to the ATP Bioluminescence Assay Kit CLS II instructions with minor adjustments.
Gel Electrophoresis and Western Blot Analysis. The relative levels of selected proteins were determined by Western blotting of whole cell lysates. After centrifugation (1000g, 10 min, 4. °C), the cell pellet was resuspended in RIPA buffer (50 mM Tris, 150 mM NaCl, 1 mM ethylenediaminetetraacetic acid, 0.5% sodium deoxycholic acid, 0.1% sodium dodecyl sulfate, 1% sulfate NP- 40; pH 7.4) in the presence of protease and phosphatase inhibitors and homogenized with a syringe and 22-gauge needle (20 strokes). Samples in the presence of cupric ions, BCA and alkaline environment were incubated for 30 min at 37°C, followed by measurement of absorbance at 562 nm on BioTek synergy HT microplate reader.
A common variant in the FTO gene is associated with body mass index and predisposes to obesity in children and adults.Science. Genetic variants in the fat and obesity-associated (FTO) gene and risk of Alzheimer's disease. PLoS One2012.7, e50354. Epitranscriptomics: Correlation of N6-Methyladenosine RNA Methylation and Pathway Dysregulation in the Hippocampus of HIV-Transgenic Rats. PLoS One2019,14, e0203566.
A commonly carried allele of the obesity-related gene FTO is associated with reduced brain volume in healthy elderly. Proc. Important role of FTO in the survival of rare pan-resistant triple-negative inflammatory breast cancer cells facing a severe metabolic challenge. PLoS One2016,11, e0159072.