PAULA CAROLINA MONTAGNANA
AVALIAÇÃO DE DÉFICIT DE POLINIZAÇÃO EM
TOMATEIROS (Lycopersicum esculentum Mill.)
AVALIAÇÃO DE DÉFICIT DE POLINIZAÇÃO EM TOMATEIROS
(Lycopersicum esculentum Mill.)
Orientador: Profa. Dra. Maria José de Oliveira Campos
Co-orientador: Gleiciani Burger Patricio
Trabalho de Conclusão de Curso apresentado ao Instituto de Biociências da Universidade Estadual Paulista “Júlio de Mesquita Filho” - Câmpus de Rio Claro, para obtenção do grau de Ecólogo
ȱ
Primeiramente agradeço aos meus pais por todo o apoio e incentivo durante os anos de
graduação e por compartilharem comigo o sonho de ser uma Ecóloga. Obrigada!
Agradeço também a minha orientadora Maria José de Oliveira Campos (Zezé!) pelos
ensinamentos, dedicação e principalmente a amizade e carinho.
A minha co-orientadora, mas principalmente amiga, Gleiciani obrigada pelos socorros
prestados para a realização de muitos campos, pela ajuda em muitos deles, orientação, risadas,
conversas furadas, enfim pela amizade.
Ao Senhor Liberato, proprietário do sítio, por ceder os pés de tomate e permitir que
esse trabalho fosse realizado, além da sua companhia sempre divertida.
A Elizandra pela ajuda com a confecção dos mapas e análises da paisagem, esse
trabalho ficou melhor graças a sua ajuda.
Aos GEECAS (Socó, Siri, Audrei, Emily) pelas ajudas em campo, idéias trocadas,
trabalhos feitos em feriados (lembram do 21 de abril?), pela amizade... Logo dominaremos a
Unesp...
A toda a Turma de Ecologia de 2006 por momentos inesquecíveis, campos
inesquecíveis, pelas risadas, pela amizade... Saibam que foram os responsáveis pela minha
permanência no curso e por haver uma bióloga a menos no mundo!!! Em especial as
queridíssimas amigas Milene e Clarissa por todos os momentos que passamos (e passaremos)
juntas, pelos almoços com muita salsicha, pelo acolhimento e amizade. A Amanda, Lívia e
Carol, obrigada também pela amizade de vocês, pela muitas risadas, momentos
divertidíssimos que eu nunca esquecerei. Adoro todos vocês!!
Obrigada a todos que de alguma forma contribuíram para a minha formação, tanto
1 - INTRODUÇÃO ... 5
2 - OBJETIVOS ...14
3 - MATERIAIS E MÉTODOS... 15
3.1 – Caracterização da área de estudo...15
3.2 – Caracterização da cultura de tomate...18
3.3 – Duração das flores e período de receptividade dos estigmas e viabilidade dos grãos de pólen ... 18
3.4 – Avaliação da ocorrência de déficit de polinização ...19
3.5 – Caracterização da fauna de abelhas polinizadoras do cultivo de tomate... 21
4 - RESULTADOS E DISCUSSÃO ... 22
4.1 – Duração das flores e período de receptividade dos estigmas e viabilidade dos grãos de pólen ... 22
4.2 –Verificação da ocorrência de déficit de polinização...23
4.3 – Caracterização da fauna de abelhas polinizadoras do cultivo de tomate... 28
5 - CONCLUSÃO ... 31
1 - INTRODUÇÃO
Estima-se que em todo o mundo existam entre 25.000 e 30.000 espécies de abelhas
(BUCHMANN e NABHAN, 1996 apud FAO, 2008) principais agentes polinizadores de
muitas espécies de plantas selvagens e de plantas cultivadas; para muitas espécies a
produtividade aumenta significativamente após as suas visitas. As abelhas também são
utilizadas de forma indireta pelo homem através da comercialização de pólen apícola, mel,
própolis, geléia real e cera, para fins alimentícios, cosméticos e medicinais. Na região do
médio Araguaia, 77,55% dos moradores utilizam os produtos de origem apícola para fins
medicinais e também na fabricação de utensílios – 6,12% (MODRO et al. 2009).
Em sistemas agrícolas a presença de agentes polinizadores é de fundamental
importância visto que 35% da produção agrícola mundial é afetada de alguma forma pela
visita de polinizadores como pássaros, morcegos e, principalmente abelhas, sendo que nos
continentes africano, asiático e latino americano aproximadamente 40% da área total
cultivada é hoje ocupada por cultivos que possuem algum grau de dependência de
polinizadores animais (FAO, 2008). Estima-se que em um cenário de ausência de
polinizadores, as culturas que dependem desses agentes podem sofrer uma perda na produção
total em torno de 5% nos países desenvolvidos a 8% nos países em desenvolvimento (AIZEN
et al., 2009), tendo Gallai et al. (2009) estimado a taxa de vulnerabilidade da produção
agrícola mundial em 9,5% frente ao mesmo cenário.
Somente no ano de 2005 o valor da produção total de 46 culturas diretamente
dependentes da polinização por insetos foi de 625 bilhões de euros, o que representa 39% do
valor da produção agrícola mundial, e 153 bilhões de euros foi o valor estimado da
polinização realizada por insetos (GALLAI et al., 2009). Dessa forma, as perdas econômicas
decorrentes do declínio de populações de polinizadores são enormes e algumas regiões do
mundo podem sofrer mais que outras, pois muitos países se especializaram em culturas
dependentes de polinizadores como, por exemplo, os países do Leste Asiático que chegaram a
ser responsáveis por 52% dos vegetais produzidos no mundo, porém para esse grupo de
cultivo a vulnerabilidade foi estimada em 15% nessa região, sendo que a taxa mundial é de
12% (GALLAI et al., 2009). Curiosamente, os cultivos com maior valor por unidade de
produção foram os que apresentaram maiores índices de vulnerabilidade à perda de
São diversas as culturas que se beneficiam da polinização realizada por abelhas,
benefício esse que resulta em frutos de melhor qualidade, maiores e com elevado teor de
nutrientes, o que agrega maior valor econômico ao produto. Em cultivos de berinjela a
presença de abelhas pode aumentar em 36% o número de frutos formados, podendo chegar a
50% de aumento na produção (MONTEMOR e SOUZA, 2009). Gemmill-Herren e Ochieng
(2008) encontraram resultados semelhantes em cultivos de berinjela no Quênia, África, onde a
polinização por abelhas aumentou significativamente o número de sementes, assim como o
peso dos frutos, principalmente das parcelas que se encontravam próximas de fragmentos de
vegetação nativa resultando em maiores índices de visitação por abelhas.
Em cultivos de pepino, a visita de abelhas às flores não é obrigatória para a formação
de frutos já que muitas das variedades são partenocárpicas, porém a polinização por abelhas
promove um aumento na frutificação de 19,2% na variedade Japonês em estufa, segundo
estudo realizado por Nicodemo (2008). O autor também avaliou a qualidade da produção,
pelo peso, comprimento e diâmetro dos frutos, não tendo encontrado diferença entre aqueles
oriundos de flores visitadas ou não por abelhas. Resultados diferentes foram encontrados por
Santos et al. (2008) em estudo também realizado em estufas, onde se pretendeu estudar a
efetividade de duas abelhas sem ferrão como polinizadoras de pepino. Os autores encontraram
que, além de uma maior quantidade de frutos formados, as estufas que continham colônias das
abelhas estudadas apresentaram uma menor proporção de frutos imperfeitos, além de frutos
maiores, mais pesados e com sementes, o que não ocorreu com os frutos das estufas que não
continham as colônias.
Os pomares de laranja (Citrus sinensis) também têm a sua produção aumentada ao
serem polinizados por abelhas, entre outros benefícios (GAMITO e MALERBO-SOUZA,
2006). Esses autores mostraram que as abelhas Apis mellifera e Trigona spinipes foram
observadas coletando principalmente néctar nas flores e apresentaram uma frequência de
visita de 77,8% e 33,3%, respectivamente. Como efeito da polinização, 40% dos botões
florais utilizados no experimento formaram frutos, o que significa um aumento de 140% na
produção de frutos em flores livremente visitadas, sendo que nas flores impedidas de serem
visitadas essa porcentagem foi de apenas 16,7%; os frutos também foram mais pesados, mais
doces e com maior quantidade de vitamina C, características que agregam maior valor à
produção. Outro estudo que também visou avaliar a influência da polinização sobre a
frutificação de variedades de laranja-doce foi realizado por Domingues e Neto (1999), que
aberta a agentes polinizadores; esse tratamento resultou em maior prendimento dos frutos para
68% das variedades estudadas.
Ao estudarem os requerimentos de polinização do meloeiro Souza et al. (2009)
concluíram que esse cultivo requer a visita de polinizadores bióticos para alcançarem altos
índices de vingamento inicial, além de grandes melhorias nos parâmetros de qualidade dos
frutos, como peso e número de sementes.
A preocupação com o declínio das populações de polinizadores é crescente em todo o
mundo, porém a preocupação com a perda de biodiversidade deve ir além da preocupação em
nível de espécies e incluir a preocupação com a perda de funções e nichos, uma vez que é
essencial manter uma certa redundância funcional dos ecossistemas. Dessa forma, a
conservação de ecossistema ou paisagem pode ser mais efetivo devido ao fato de que uma
maior diversidade de espécies e de estoques de genes pode ser salva (ODUM e BARRETT,
2008).
Diversos são os fatores que ameaçam a manutenção das populações de abelhas, entre
eles a intensificação da agricultura que causa, entre outras coisas, uma simplificação da
paisagem devido à supressão da vegetação nativa e a implantação de monocultura, reduzindo
a diversidade local de espécies. Segundo Odum e Barrett (2008), 30% da superfície terrestre
não coberta de gelo é hoje ocupada por alguma forma de agricultura, com cerca de 20% dessa
área dedicada à produção de animais e 10% dedicada a cultivos. Ainda segundo os autores, as
melhores terras, as facilmente cultiváveis por meio da tecnologia existente, estão atualmente
todas em uso. Sendo assim, a intensificação agrícola, que implica na expansão da área
cultivada e na crescente utilização de agrotóxicos, é a principal ameaça aos polinizadores, em
especial às abelhas.
Dados da FAO de 2009 contidos no documento “State of the world’s forests 2009”
mostram que a agricultura, em suas diferentes abrangências, é responsável por 66% das
causas de desmatamento na América Latina (Figura 1). De acordo com esse documento ainda,
a conversão direta da terra em áreas agrícolas permanentes de larga escala, principalmente a
agricultura comercial focada na exportação, que incluí a pecuária, a soja, os cultivos voltados
à produção de biocombustíveis, frutas, vegetais e flores decorativas, respondem por 47% das
causas diretas pelas mudanças na cobertura florestal. Em proporção menor, mas responsável
por 13% da destruição da cobertura florestal, está a conversão dessas áreas florestais em áreas
agrícolas permanentes em pequena escala (propriedades familiares e pequenas propriedades),
porém a viabilidade dessas áreas está diminuindo devido a sua incorporação pelas grandes
cultivos em geral sobre as áreas de outras variedades (4%) e ao deslocamento de áreas
agrícolas sobre áreas florestais intactas (2%) (Figura 1).
A homogeneização da paisagem ao longo de extensas áreas agrícolas contribui para
uma diminuição na riqueza de espécies pela perda de heterogeneidade espacial (BEGON,
TOWNSEND e HARPER, 2007). Tais mudanças na estrutura da paisagem interferem na
relação planta-polinizador em nível de indivíduo, população e comunidade, porque alteram a
distribuição espacial e temporal de recursos florais, principalmente néctar e pólen, e de sítios
de nidificação (KREMEN et al., 2007).
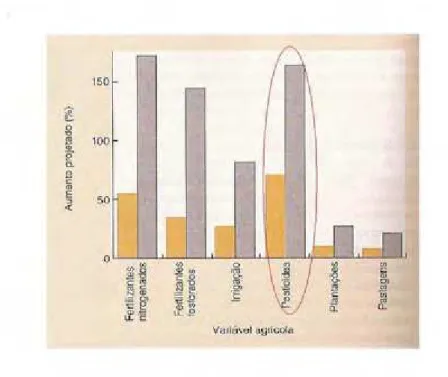
O aumento no uso de pesticidas também é um dos perigos decorrentes da expansão
agrícola e estima-se que nos próximos 50 anos esse aumento será mais de 150%, conforme
ilustra a Figura 2, adaptada de Begon, Townsend e Harper (2007). Em estudo realizado por
Baptista et al. (2009) foram avaliadas as toxicidades de inseticidas utilizados na cultura de
citros para Apis mellifera e os autores chegaram ao resultado de que o organofosforado
acefato é altamente tóxico para as operárias adultas quando pulverizado sobre os indivíduos,
quando alimento contaminado é ingerido (visto que há o acúmulo no néctar e no pólen) e
quando o inseto entra em contato com uma superfície contaminada. Skerl et al. (2009)
também avaliaram o nível de resíduos no pólen coletado por Apis mellifera carnica após a
aplicação dos inseticidas mais comumente utilizados em pomares de maçã, sendo eles o
Diazinon e Thiacloprid. Os resultados indicaram a presença de resíduos dos dois agentes no
pólen no primeiro dia após a aplicação no pomar e a concentração diminuiu somente entre o
6º e 10º dias. Tais produtos causam efeitos adversos em adultos e efeitos subletais causando
Figura 1 – Diferentes causas das mudanças de áreas florestais tropicais nos países da América Latina, 1990-2000. Modificado de FAO (2009).
Ganhos em áreas de floresta e cobertura de dossel
Conversão direta para agricultura permanente de grande escala
Intensificação agrícola sobre áreas cultivadas
Expansão agrícola sobre áreas de floresta
Conversão direta para agricultura permanente de pequena escala
Figura 2 - Aumento projetado no uso de pesticidas nos anos de 2020 (barra alaranjada) e 2050 (barra cinza). Modificado de Begon, Townsend e Harper (2007).
Valdovinos-Nunez et al. (2009) estudaram a toxicidade de pesticidas em três espécies
de abelhas Melipona beecheii, Trigona nigra e Nannotrigona perilampoides no México.
Segundo os autores, as abelhas estudadas se mostraram altamente suscetíveis aos compostos
se comparadas com Apis mellifera, sendo os químicos da classe nicotinode os mais tóxicos
com DL50 de apenas 0,001 μg no caso de N. perilampoides.
O desmatamento, intimamente relacionado com a intensificação agrícola, ameaça
também as Unidades de Conservação e dessa forma a presença de polinizadores nas áreas
adjacentes.
A abundância e a riqueza de espécies de Meliponini em áreas agrícolas estão
fortemente relacionadas com a cobertura florestal do entorno. Estudos realizados por Brosi
(2009) em uma paisagem fragmentada na Costa Rica e por Brown e Albrecht (2001) em
Rondonia mostraram que a abundância e a riqueza de espécies estão diretamente relacionadas
à porcentagem de cobertura de florestas primárias. Essa associação não é surpresa visto que
muitos meliponineos nidificam em cavidades de árvores presentes em florestas tropicais
(BLAUMGARTNER e ROUBIK, 1989).
Esses dados são preocupantes porque os meliponineos contribuem para a polinização
de mais de 60 cultivos tropicais, tais como o café, e são os polinizadores efetivos do abacate,
atividades de polinização é realizada por colônias selvagens estabelecidas em árvores de
grande porte em floresta primária (BROSI, 2009).
Outra conseqüência do desmatamento é o isolamento de fragmentos de vegetação
nativa circundados, muitas vezes, por uma matriz agrícola impermeável que pode reduzir ou
mesmo impedir a recolonização de outros fragmentos comprometendo, dessa forma, a
manutenção de sub-populações e de metapopulações, podendo levar a extinções locais tanto
de polinizadores como de plantas (KREMEN et al., 2007).
A introdução de espécies exóticas também se constitui um fator de risco para
polinizadores. Desde o momento em que o ser humano começou a domesticação de plantas e
animais há a introdução de diversas espécies ao longo de regiões onde elas naturalmente não
ocorriam, porém a partir do século XVI, período das grandes navegações, o transporte se deu
de forma mais rápida e em maior volume, aumentando desde então. No caso de introdução de
abelhas, sabe-se que a abelha do mel (Apis mellifera) foi domesticada pelo homem há 4.000
anos e tem sido introduzida em quase todos os países (GOULSON, 2003).
A introdução de espécies exóticas de abelhas, principalmente Apis mellifera, que
apresenta alta capacidade de ocupação de habitats e dieta muito diversificada não afeta apenas
as abelhas nativas, mas toda uma variedade de animais que se alimentam de néctar e/ou pólen
como pássaros, morcegos, mamíferos e outros insetos e de plantas com fecundação cruzada.
Goulson (2003) listou cinco possíveis efeitos indesejáveis da introdução de abelhas exóticas,
sendo (1) a competição com visitantes florais nativos por recursos alimentares, (2) a
competição com organismos nativos por locais de nidificação, (3) transmissão de parasitas ou
patógenos para os organismos nativos, (4) mudanças no padrão de produção de sementes de
plantas nativas (aumento ou diminuição) e (5) polinização de plantas daninhas exóticas.
A competição entre a abelha introduzida Apis mellifera e a abelha nativa Bombus
occidentalis foi estudada por Thomson (2004) no Estado da Califórnia, Estados Unidos. Os
resultados mostraram que a média de esforço de forrageamento de pólen, ou seja, a
quantidade de operárias de Bombus occidentalis que voltavam da viagem de forrageamento
com cargas de pólen nas corbículas, aumenta significativamente com a distância das colônias
de A. mellifera. Outro dado importante foi o aumento do sucesso reprodutivo observado em
colônias de Bombus, quanto mais distantes se encontravam de colônias de Apis.
Roubik e Villanueva-Gutiérrez (2009) avaliaram os efeitos da invasão de Apis
mellifera sobre as abelhas nativas na península de Yucatan, México, em um estudo com
duração de 20 anos onde foram utilizados ninhos armadilhas e amostragens de pólen para
comunidades de plantas e abelhas da península atribuída á invasão de A. mellifera,
principalmente no tocante a preferências florais, como o consumo de espécies de pólen
importantes para a reprodução das abelhas locais, porém, contraditoriamente, essa competição
não levou a um total declínio da população de abelhas nativas. Por isso, os autores
formularam as seguintes hipóteses: (1) as abelhas nativas ajustam seus comportamentos ou
sua fisiologia e alteram suas escolhas florais diante de uma nova espécie competidora e (2) as
abelhas melíferas, como polinizadoras, aumentam a disponibilidade de espécies de plantas
que seriam recursos básicos para as abelhas nativas devido ao seu comportamento generalista.
Déficit de polinização
O déficit de polinização pode ser entendido como uma série de eventos que limitam a
quantidade ou alteram a qualidade do pólen depositado sobre o estigma das flores
ocasionando em uma menor produção de frutos e sementes quando comparado com situações
onde há níveis adequados de recepção de pólen (KNIGHT et al., 2005). São vários os
distúrbios que podem causar uma limitação polínica, tais como a introdução de espécies
exóticas (tanto espécies de plantas como de polinizadores), fragmentação de habitats e
diminuição do tamanho populacional de plantas e de polinizadores (FREITAS et al., 2010). A
perda de polinizadores causa déficit na polinização, entre outros motivos, por diminuir o
índice de visitação às flores pelos insetos devido à abundância populacional e diversidade
diminuída, muitas vezes pelos fatores já citados anteriormente. Freitas et al. (2010)
apresentam uma revisão sobre a limitação polínica em plantas nativas da Mata Atlântica.
A ocorrência de déficit de polinização em plantas cultivadas pode ser devido a uma
recepção inadequada de pólen, de natureza tanto quantitativa (número insuficiente de visitas
recebidas por flor), o que resulta em uma baixa produção ou uma limitação qualitativa
(referentes a fatores como a qualidade, distribuição e/ou origem do pólen) o que afeta as
características do fruto ou das sementes, por exemplo. O déficit qualitativo da polinização
pode ser o resultado de condições como ausência de abelhas silvestres altamente eficientes
como polinizadores ou de uma baixa diversidade de polinizadores, acarretando em menores
níveis de polinização (VAISSIÈRE et al., 2009).
A condução de experimentos envolvendo suplementação de pólen permite a
verificação de ocorrência de déficit de polinização em uma população natural ou cultivada.
Nesses experimentos se compara o sucesso reprodutivo de plantas controle, ou seja, expostas
a polinização natural, com plantas que receberam suplemento de pólen. A suplementação de
como corre em áreas agrícolas, por meio de enriquecimento da área com polinizadores,
quando colméias são transportadas para os campos de cultivos (THOMSON, 2001). Se os
resultados indicarem uma maior produção de frutos ou sementes nas plantas suplementadas
quando comparadas com as plantas controle, concluí-se que há um déficit de polinização na
população estudada (KNIGHT et al., 2005). Siqueira et al. (2009) ao estudarem a ecologia da
polinização do maracujá-amarelo (Passiflora edulis f. flavicarpa Deg.) verificaram a
ocorrência de déficit na polinização ao compararem a qualidade dos frutos provenientes dos
testes de polinização cruzada manual e polinização natural. Os frutos apresentaram maior
número de sementes bem formadas, maior concentração de açúcares e maior espessura de
casca quando submetidos à polinização cruzada manual. Os autores observaram também o
florescimento de um número elevado de plantas na vegetação em torno dos cultivos no
período chuvoso e a grande atratividade dessas espécies sobre as abelhas e levantaram a
possibilidade de que esse poderia ser um fator adicional contribuindo para o déficit de
polinização.
A cultura do tomate
O tomate (Lycopersicum esculentum Mill) é a segunda hortaliça mais cultivada no
mundo em área plantada e a primeira em volume industrializado (FILHO et al., 1994).
Segundo dados do IBGE de 2002, no Brasil a cultura do tomate ocupou naquele ano uma área
de plantio de 62.647 ha, produzindo 3.652.923 toneladas e gerando um valor de R$
1.429.412. Somente no Estado de São Paulo a área plantada no ano de 2002 chegou a 11.930
ha que produziram 765.990 toneladas e totalizaram R$ 323.248. Originário da região andina
da América do Sul, o tomateiro é uma planta anual com uma inflorescência em forma de
cacho produzindo de 6 a 12 flores, sendo elas bissexuais com o tubo do cálice curto e coberto
de pelos. Em geral há 6 pétalas de cor amarela e recurvadas quando maduras, mesma cor das
anteras poricidas que estão dispostas ao redor do estilete provido de uma ponta alongada
estéril (NAIKA et al., 2006).
Por possuir anteras poricidas, o tomateiro possui como característica a associação a
melitofilia, especialmente por abelhas capazes de realizar polinização por vibração (VIANNA
et al., 2007), como exemplificado no experimento realizado por Greenleaf e Kremen (2006)
em plantações de tomate no Nordeste da Califórnia, onde constataram que os tomateiros do
grupo submetidos à polinização aberta, sendo visitados especialmente por abelhas silvestres,
produziram tomates maiores que as plantas dos demais grupos (grupo controle,
2 - OBJETIVOS
O presente trabalho tem como objetivos:
• Verificar a existência de déficit de polinização em tomateiros cultivados em uma propriedade rural no município de Rio Claro/São Paulo e, em caso afirmativo, quais
fatores contribuem para isso, tais como:
- paisagem altamente modificada por uma agricultura intensiva;
- ausência de remanescentes florestais;
- ausência de vegetação ruderal no entorno dos canteiros cultivados com tomate;
- baixa frequência de visitas de abelhas nas flores.
• Caracterizar a fauna de abelhas polinizadoras do cultivo de tomate.
3 - MATERIAIS E MÉTODOS
3.1 – Caracterização da área de estudo
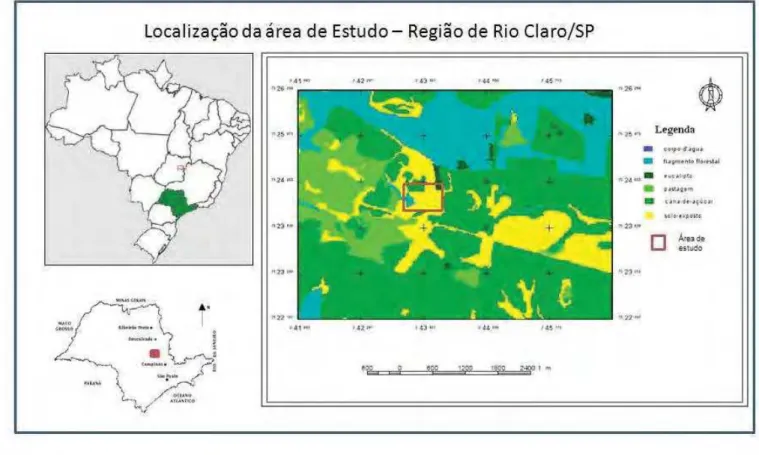
O estudo foi realizado na propriedade “Bom Jesus” localizada no município de Rio
Claro/São Paulo (22º22’14.03”S e 47º29’8.88”O) situado ás margens da Rodovia Wilson
Finardi (SP 191) na altura do kilômetro 59, sentido Araras-Rio Claro (Figura 3). O sítio
possui aproximadamente 48 hectares, sendo classificado como pequena propriedade de acordo
com a Lei 8.629/93. A produção é voltada para o cultivo de hortaliças (verduras e legumes em
geral), incluindo o tomate. Nos arredores dos canteiros de tomate são cultivadas cana de
açúcar, árvores frutíferas e hortaliças diversas, como ilustra a Figura 4.
Figura 4 – Vista geral do entorno dos canteiros de tomate. (A) Áreas de solo exposto preparados para o plantio com os canteiros de tomate ao fundo, (B) Borda de cultivo de cana-de-açúcar, (C) Canteiros de tomate próximos à espécies arbóreas frutíferas e (D) Cultivo de hortaliças.
O clima da região é classificado como sendo do tipo “Cwa” segundo a classificação de
Koeppen, que caracteriza clima subtropical de inverno seco e frio e verão quente e úmido. As
médias de temperatura e precipitação nos meses de realização do estudo foram,
respectivamente, 17,9º C e 26,9 mm (julho), 19,7º C e 28,8 mm (agosto) e 21,2ºC e 66,8 mm
(setembro) (CEPAGRI). Porém, de acordo com o Boletim de Informações Climáticas do
CPTEC/INPE de 20 de agosto de 2010, o fenômeno La Nina se estabeleceu no período de
final de julho e início de agosto, o que ocasionou temperaturas acima da média e baixos
valores de umidade relativa do ar na região de estudo.
Tendo como objetivo descrever a paisagem em torno da área de estudo, para obter
subsídios para a discussão de aspectos relacionados a ocorrência de déficits de polinização,
foi feita uma caracterização do uso e ocupação do solo no entorno da área de estudo, para o
qual foram utilizadas imagens CBERS, adquiridas gratuitamente junto a catálogos online do
Instituto Nacional de Pesquisas Espaciais (INPE). As imagens CBERS utilizadas foram do
sensor CCD, órbita 155, com data de imageamento 09/12/2006.
Todas as etapas de processamento digital de imagem foram realizadas no aplicativo
SPRING (CÂMARA et al., 1996). Após o registro, as imagens CCD foram realçadas com a
aplicação de contraste, modelo linear, e segmentadas (BINS, 1996), sendo empregada a
B
A
composição R2G4B3 e os seguintes parâmetros: método crescimento de regiões e valor de
similaridade e área iguais a 20.
As imagens segmentadas foram classificadas com o uso do algoritmo de agrupamento não
supervisionado Isoseg, com limiar de aceitação de 95%. Os temas identificados, 48 no total,
foram agregados conforme as classes presentes na área de estudo, sendo transformadas em
imagem temática. A imagem classificada referente a cada classe de uso e ocupação do solo foi
exportada para o aplicativo Fragstat (McGARIGAL and MARKS 1995), onde foram
realizadas rotinas de cálculo da paisagem (Tabela 1) os quais foram analisados.
Tabela 1 - Métricas de Ecologia de Paisagem calculadas para a área de estudo Área Total da Classe (CA)
Porcentagem da Paisagem (PLAND)
Número de Fragmentos (NP)
Índice de formato da Paisagem (LSI) Métricas de Classe (CLASS)
Índice de coesão de Fragmentos
(COHESION)
As classes de uso e ocupação do solo presentes na área de estudo foram medidas em
uma paisagem com 5 kilômetros de raio a partir da propriedade onde se localizavam os
canteiros de tomate e tiveram as seguintes métricas de classe calculada: Área Total da Classe
(CA) que mede a composição da paisagem, sendo que a soma de todas as áreas de todos os
fragmentos de um tipo correspondente aproximando-se de zero quanto mais raro o fragmento
for na paisagem. Porcentagem da Paisagem (PLAND) é a porcentagem da paisagem composta
por um tipo específico de fragmento, ou seja, da classe. Número de Fragmentos (NP), como o
próprio nome diz, mede a quantidade de fragmentos existentes na paisagem referentes à classe
medida. Índice de Formato de Paisagem (LSI) é uma medida de agregação de classes, onde se
LSI é igual a 1 a paisagem consiste em um único quadrado de fragmentos, aumento sem
limites à medida que o tipo de fragmento torna-se menos agregado. Índice de Coesão de
Fragmentos (COHESION) mede a conectividade física de uma classe, aproximando-se de
zero à medida que a proporção da paisagem referente à classe focal diminui e torna-se mais
3.2 – Caracterização da cultura de tomate
O estudo foi realizado em tomateiros do cultivar “Perinha”. Os canteiros de tomate
foram formados pelo proprietário, sem a interferência do pesquisador, com aproximadamente
40 plantas na área de estudo.
3.3 – Duração das flores, período de receptividade dos estigmas e viabilidade dos grãos de pólen
Esse estudo foi conduzido para a identificação dos melhores períodos para a realização
dos testes de polinização e do levantamento da fauna de abelhas com potencial de polinização
das flores de tomateiro.
Para a determinação da duração média de uma flor de tomateiro foram marcados 5
botões com linha colorida e acompanhados desde a sua abertura até o murchamento.
Os períodos de receptividade do estigma e viabilidade do pólen foram avaliados pelo
acompanhamento de 50 botões marcados com linhas coloridas, que foram ensacados em fase
de pré-antese; a cada dia, 10 flores eram coletadas e levadas ao laboratório onde se realizavam
os testes para a determinação da receptividade estigmática e viabilidade de pólen. Esse
procedimento foi feito para a fase de botão, e para flores abertas, 24, 48, 72 e 96 horas após a
antese; o objetivo foi o de verificar qual o tempo de duração da flor, para posteriormente
realizar os testes de polinização.
Para a determinação da receptividade do estigma foi utilizado o corante Sudan IV, de
acordo com Dafni (1992), onde a receptividade é indicada pela mudança de cor do estigma,
que quando receptivo se colore de vermelho. A viabilidade dos grãos de pólen foi avaliada
com o emprego de carmim acético que colore os grãos de pólen viáveis de vermelho
permitindo assim a sua diferenciação dos grãos de pólen não viáveis (DAFNI, 1992). Para a
coloração por carmim acético as anteras das flores eram retiradas com o auxílio de uma pinça
e com uma espátula eram abertas de forma que os grãos de pólen fossem depositados sobre
uma lâmina que em seguida recebia duas gotas de carmim acético. Depois da colocação de
uma lamínula e após 5 minutos, as lâminas eram analisadas em microscópio óptico. A análise
da viabilidade dos grãos de pólen era feita através da contagem do número de grãos viáveis
em 100 grãos escolhidos aleatóriamente.
Os testes realizados mostraram um período de duração da flor de 3 dias, de forma que
suficiente para a flor completar o seu ciclo e ser exposta somente quando não houvesse a
possibilidade de receber pólen através de outras vias.
3.4 – Avaliação da ocorrência de déficit de polinização
Para a verificação da ocorrência de déficit de polinização foram realizados testes que
avaliaram a taxa de formação de frutos, o peso médio e o número médio de sementes por fruto
a partir de flores que receberam os seguintes tratamentos:
a) Exclusão de polinizadores:
Procurou-se com este teste avaliar os efeitos da ausência de visitas de abelhas
polinizadoras nas flores de tomate, e para isso foram marcados 40 botões com linhas na cor
laranja e estes tiveram a visita por polinizadores impedida usando-se sacos de papel
perfurados com agulha para evitar o acúmulo de umidade (Figura 5). Depois de uma semana
os sacos de papel foram retirados e as flores foram acompanhadas para verificação de
formação de frutos.
Figura 5 – Teste de exclusão de polinizadores. (A) Botão marcado com linha laranja indicando o tipo de teste, (B) Botão ensacado para impedir a visita de polinizadores.
b) Polinização manual cruzada:
O teste de polinização manual foi realizado de forma a simular uma polinização
“perfeita” onde a flor receberia uma grande quantidade de pólen proveniente de flores de
outros indivíduos. Assim, 40 botões em fase de pré antese foram marcados com linhas na cor
azul e ensacados com sacos de papel perfurados para impedir a visita por polinizadores ao
abrirem. No dia seguinte, depois de abertas, as flores foram emasculadas com o auxílio de
uma pinça, para impedir a autopolinização. A escolha de flores com até 24 horas pós antese
para a realização destes testes deve-se ao fato de que, embora os estigmas estejam receptivos
e os grãos de pólen viáveis em flores ainda em botão, as anteras estão fechadas até cerca de 48
horas após a antese. Dessa forma a autopolinização na fase de botão até 24 horas após a
antese fica impedida. As anteras extirpadas foram abertas e o pólen foi retirado e utilizado
para a polinização cruzada (xenogamia). O transporte do pólen até o estigma foi realizado
com o auxílio de um pincel. Depois de realizada a polinização cruzada, cada flor foi
novamente ensacada (Figura 6) e ao final de uma semana os sacos foram retirados e as flores
acompanhadas para a verificação de formação de frutos.
Figura 6 – Teste de polinização manual cruzada. (A) Botão marcado com linha azul indicando o tipo de teste, (B) Flor ensacada depois de emasculada e polinizada para impedir a visita de polinizadores.
c) Polinização aberta:
As flores utilizadas neste teste foram expostas a polinização por abelhas. Quarenta
botões foram marcados com linhas de cor rosa e acompanhados para a verificação de
formação de frutos (Figura 7).
Figura 7 – Teste de polinização aberta com botão marcado com linha rosa indicando o tipo de teste.
Para a comparação do peso médio e número médio de sementes por fruto formado a
partir de cada tratamento foi realizada análise de variância (ANOVA) ao nível de
significância de p<0,05.
3.5 – Caracterização da fauna de abelhas polinizadoras do cultivo de tomate
A coleta das abelhas foi realizada de forma a caracterizar a fauna visitante das flores
de tomate e registrar a sua presença na área de estudo. Esta etapa do estudo foi realizada em
setembro de 2010, período correspondente ao pico de florescimento dos tomateiros na área.
Foram coletados todos os indivíduos que pousavam nas flores no período das 7h às 17h, em
condições climáticas de dia ensolarado e pouco vento. O esforço amostral foi equivalente a
100 horas de coletas divididas entre 5 coletores que percorriam as linhas do cultivo durante os
primeiros 20 minutos de cada hora, coletando os indivíduos que pousavam nas flores de
4 - RESULTADOS E DISCUSSÃO
4.1 – Duração das flores, período de receptividade dos estigmas e viabilidade dos grãos de pólen
Foi observado que a duração média de uma flor de tomateiro é de 3 dias, sendo que a
partir do 4º dia ocorre o murchamento das flores que apresentam então alteração da coloração
das pétalas e escurecimento do ápice das anteras e do estigma que se curvam para baixo. A
figura 8 apresenta uma seqüência de fases de desenvolvimento da flor de tomateiro, desde
botão até o início da formação de frutos.
Figura 8 – Ciclo de desenvolvimento da flor de tomate (Lycopersicum esculentum Mill.), de botão até o fruto formado. (A) Botão em pré-antese, (B) Botão a um dia da antese – fase em que eram ensacados para a realização dos testes, (C) Flor em início de antese, (D) Flor aberta, (E) Flor em senescência e (F) Início da formação do fruto.
O estigma manteve-se receptivo desde a fase de botão (a um dia da antese) até o
terceiro dia de duração da flor aberta, sendo que por volta do quarto dia quando a flor começa
a murchar o estigma perde a sua receptividade (Tabela 2).
A
B
C
Tabela 2 – Porcentagem média de botões e flores com estigmas receptivos e grãos de pólen viáveis em diferentes estágios de desenvolvimento.
Estágio de desenvolvimento
da flor
Porcentagem média de
flores com estigma receptivo
(N=50)
Porcentagem média de
grãos de pólen viáveis por
flor (N=50)
Botão em pré antese 100 91
Flor aberta há 24 horas 80 89,5
Flor aberta há 48 horas 90 94,8
Flor aberta há 72 horas 100 83,2
Flor aberta há 96 horas 20 81
É possível encontrar grãos de pólen viáveis durante todo o período de duração da flor,
com um leve aumento da proporção de grãos inviáveis na medida em que a flor se aproxima
da senescência. O mesmo acontece com a proporção de estigmas receptivos, mas a perda de
receptividade é mais marcante que a perda de viabilidade.
4.2 –Verificação da ocorrência de déficit de polinização
Para este estudo foram marcadas 120 botões (40 flores para cada tratamento) tendo
havido perda de 16 flores devido à predação por lagartas e ao manuseio do proprietário nos
cuidados com o cultivo. A Tabela 3 apresenta o número e a porcentagem de frutos formados a
partir de cada tratamento.
Tabela 3 – Produção de frutos formados a partir dos diferentes tipos de tratamentos
Tratamento Número de flores Número de frutos
formados
% de frutos formados
Exclusão de visitantes
florais
30 22 73,33
Polinização manual
cruzada
38 30 78,94
Polinização aberta 36 16 44,44
O tratamento “Polinização aberta” apresentou a menor porcentagem de frutos
um fruto. Stephenson (1981), em uma revisão sobre o assunto, afirma que há espécies de
plantas que adotam uma estratégia de reprodução onde são produzidas flores em excesso,
podendo ocorrer o aborto de frutos excedentes, ou seja, daqueles que a planta não conseguirá
nutrir. Frutos danificados também podem ser abortados como forma de realocação de recursos
para o amadurecimento de outros que contribuam para uma melhor formação das próximas
gerações, como também podem ocorrer abortos de frutos danificados como forma de evitar a
contaminação por patógenos.
Stephenson (1981) também relata a ocorrência de frutos abortados devido a fatores
ligados à polinização, como variações na quantidade de pólen depositado sobre o estigma que
também leva a uma variação no número de sementes formadas por fruto. Quando isso ocorre,
os frutos com menor quantidade de sementes tem maior probabilidade de sofrer aborto,
principalmente quando há a competição por recursos limitados. No caso das flores submetidas
ao tratamento de polinização aberta é possível que os frutos abortados tenham sido
polinizados inadequadamente pelas abelhas que ocorrem na região, podendo (1) o número de
visitas às flores ter sido insuficiente para levar uma quantidade adequada de pólen ao estigma
ou (2) a população local de abelhas não é em número suficiente para visitar todas as flores do
cultivo, podendo dessa forma sinalizar a existência de um déficit de polinização. O fato de
estes frutos terem ficado desprotegidos durante o período de estudo podem tê-los tornado
mais susceptíveis a pressão de predação, dessecamento e outras intempéries.
A tabela 4 mostra os resultados das métricas de classe aplicadas na área de estudo,
onde juntamente com as visitas ao local e através de imagens de satélite (ver Figura 3) foi
possível identificar a ocorrência de solo exposto, plantações de cana-de-açúcar, pastagem,
plantações de eucalipto, fragmentos florestais e corpos d’água. Através de uma análise da
paisagem podem ser obtidos indícios de que o entorno da área de estudo não seja propício
para o estabelecimento de populações de abelhas em tamanho suficiente para atuarem como
Tabela 4 – Valores obtidos para os diferentes parâmetros descritores da paisagem em torno da área de estudo. CA – Área Total da Classe, PLAND – Porcentagem da Paisagem, NP – Número de Fragmentos, LSI – Índice de Formato da Paisagem, COHESION – Índice de Coesão de Fragmentos.
ÁREA DE ESTUDO
TIPO CA PLAND NP LSI COHESION
Solo exposto 89,49 Km2 17,48% 276 5,58 99,05
Cana-de-açúcar
178,29 Km2 34,83% 362 23,87 99,71
Pastagem 68,69 Km2 13,42% 161 9,06 99,56
Eucalipto 7,17 Km2 1,40% 120 0,12 93,91
Fragmento
florestal
55,07 Km2 10,76% 161 3,63 98,59
Corpo d’água 2,95 Km2 0,57% 32 0,11 94,88
A cana-de-açúcar foi a classe que apresentou a maior área, aproximadamente 178,29
Km2 correspondendo a 34,83% da composição da paisagem, com 362 fragmentos
identificados e sendo a classe com maior índice de agregação. A segunda classe com maior
área foi a de solo exposto, provavelmente preparado para o plantio de cana, com
aproximadamente 89,49 Km2, cerca de 17,48% da composição da paisagem e 276 fragmentos.
Porém quando é analisado o LSI, a classe que possui o segundo maior índice de
agregação é a pastagem (LSI=9,06) e não o solo exposto (LSI=5,58). Dentro da classe
fragmento florestal as áreas mais representativas são a Mata São José e a Floresta Estadual
Edmundo Navarro de Andrade, que dentro da paisagem analisada são os maiores fragmentos
florestais remanescentes.
A tabela 4 e as figuras 3 e 4 mostram que a área de estudo está inserida em um
contexto de agricultura intensiva caracterizada, principalmente, pela atividade canavieira, com
grandes áreas de um cultivo que não oferece nenhum recurso para polinizadores e alguns
poucos fragmentos florestais. Uma paisagem fragmentada pode causar o rompimento de
interações planta-polinizador acarretando um declínio na reprodução das plantas nativas e/ou
cultivadas (WILCOCK e NEILAND, 2002). Em experimento que procurou demonstrar o
efeito do isolamento de habitats (o que ocorre em uma paisagem fragmentada), cultivos de
rabanete e mostarda (ambos auto-incompatíveis) foram cultivados a diferentes distâncias, de
campos ricos em espécies de polinizadores. Os resultados mostraram que à medida que o grau
abelhas às flores por hora era reduzido bem como a diversidade taxonômica dos visitantes e a
taxa de formação de frutos e número de sementes, indicando que o isolamento de habitats
também resulta em limitação da polinização e no fluxo de pólen (STEFFAN-DEWENTER e
TSCHARNTKE, 1999).
Espécies nativas de abelhas estão intimamente relacionadas à presença de áreas de
vegetação nativa, visto que muitas dessas espécies nidificam e se abrigam nessas áreas. Essa
relação foi encontrada por Greenleaf e Kremen (2006) em cultivos de tomate no norte da
Califórnia, Estados Unidos, onde os principais visitantes do cultivo foram abelhas nativas,
principalmente Bombus vosnesenskii, cuja taxa de visitação dependia da proximidade de
habitats naturais, ou seja, a espécie de Bombus foi mais abundante quanto mais perto o cultivo
se encontrava de vegetação nativa. A mesma relação foi encontrada por Klein et al. (2003) em
plantações de café na Indonésia, por Kremen et al. (2004) em cultivos de melancia nos
Estados Unidos e por Kim et al. (2006) em plantações de girassol nos Estados Unidos.
A comparação da qualidade de frutos formados (peso médio e número médio de
sementes por fruto) a partir dos diferentes tratamentos neste estudo permite discutir aspectos
adicionais sobre uma possível ocorrência de déficit de polinização. Os resultados obtidos a
partir dessa análise constam da tabela 5.
Tabela 5 – Peso médio e número médio de sementes de frutos de tomateiro (Lycopersicum esculentum Mill) formados a partir dos diferentes tratamentos.
Tratamento Número de frutos Peso médio (g)* Número médio de
sementes*
Exclusão de
polinizadores
22 14,43±6,27 a 36,5±15,64 a
Polinização manual
cruzada
30 7,98±3,82 b 26,3±16,30 a
Polinização aberta 16 14,96±7,85 a 50,12±16,47 b
*Coluna 3 e 4: Média±Desvio Padrão
Valores seguidos pelas mesmas letras na coluna não diferem entre si a P < 0,05, pelo teste de Tukey.
Os resultados mostram que em relação ao peso médio dos frutos não há diferenças
significativas entre os testes de exclusão de polinização e polinização aberta. Esses resultados
uma planta agrícola que vem sofrendo seleção artificial desde o início de século XVI, a
autogamia, forçada pela ausência de polinizadores, própria da região andina de onde esta
espécie se origina, conduziram a uma fixação rápida dos genes o que levou a um aumento da
uniformidade da espécie, de forma que uma das características que provavelmente se fixou foi
a autogamia (NUEZ, 1995).
Contudo, o tratamento “polinização aberta” resultou em um número de sementes
significativamente maior se comparado aos demais tratamentos indicando que, apesar de o
tomateiro ser autocompatível, a polinização por abelhas pode melhorar a qualidade dos frutos,
pela deposição de uma maior quantidades de grãos de pólen sobre o estigma, o que aumenta a
quantidade de sementes produzidas. Sabe-se que as sementes desempenham um papel
importante na produção de hormônios que atuam no desenvolvimento dos frutos
(STEPHENSON, 1981). Também foi observado no presente estudo que a polinização aberta
produziu frutos que possuíam a polpa mais consistente e avermelhada que os outros dois
tratamentos, por exemplo, os frutos da exclusão de polinizadores eram ocos, com poucas
sementes e com a polpa menos avermelhada. Macias-Macias et al. (2009) avaliaram a
contribuição das abelhas como polinizadoras de flores de tomate e encontraram resultados
similares. Segundo os autores, a qualidade dos frutos foi maior no tratamento com flores
polinizadas por abelhas do que nas flores não polinizadas, inclusive com a formação de um
maior número de sementes por fruto. Estudo realizado com tomates por Santos et al. (2009)
também comprovaram a eficiência das abelhas na melhora da qualidade dos frutos em estufas
que abrigavam colônias de Meliponini em relação àquelas sem colônias.
Souza et al. (2009) estudaram os requerimentos de polinização em culturas de
meloeiro cultivados em pomares a céu aberto aplicando quatro tratamentos, sendo (1)
polinização cruzada manual, (2) polinização aberta com a introdução de abelhas Apis
mellifera, (3) polinização livre e (4) restrição à visita de polinizadores. Em relação a todas as
variáveis analisadas (taxa de vingamento inicial de frutos – com até 7 dias de formação -,
produção de frutos, peso dos frutos e número de sementes nos frutos) o tratamento
“polinização manual cruzada” apresentou os melhores resultados comparado aos demais
tratamentos, provavelmente devido a uma maior quantidade de pólen depositado na região dos
estigmas nesse tratamento. Ao se analisar os resultados dos tratamentos polinização aberta
com a introdução de abelhas e polinização livre, com o primeiro obteve-se melhores
resultados em todas as variáveis se comparado ao segundo, pois com a introdução de colônias
A Tabela 6 apresenta os resultados de tratamentos de polinização aberta e exclusão de
polinizadores para diferentes cultivos em relação ao número médio de sementes por fruto.
Tabela 6 – Comparação do número médio de sementes de frutos formados a partir de polinização aberta e a partir da exclusão de visitantes florais em diferentes cultivos.
Espécie Referência Polinização aberta Exclusão de
polinizadores
Lycopersicum
esculentum (tomate)
Psidium guajava L.
(goiaba)
Capsicum frutescens
(pimenta)
Macias-Macias et al.,
2009
Freitas e Alves, 2008
Cruz, D. O. 2009
162,1±36,3 413,56±17,10 11,21±6,51 52,5±14,2 187,00±33,66 11,68±6,76
Cucumis melo L.
(melão)
Souza et al., 2009 545,0±31,6 500,4±59,8
Colunas 3 e 4: Média±Desvio Padrão
É importante salientar que alguns fatores referentes à metodologia empregada neste
estudo podem ter contribuído para os resultados encontrados. Como os experimentos de
polinização cruzada foram realizados em flores recém abertas (com 24 horas pós antese), que
foram ensacadas a seguir, pode-se sugerir que a quantidade de pólen depositado poderia ter
sido insuficiente para a fecundação de todos os óvulos. As flores abertas à visitação podem ter
recebido pólen por um período muito mais longo. Ao mesmo tempo, pode-se sugerir também
que uma grande quantidade de pólen depositada manualmente sobre o estigma pode ter
gerado uma competição entre os grãos para a formação do tubo polínico, comprometendo a
fecundação dos óvulos. Apesar de o estigma estar receptivo já no primeiro dia da flor aberta -
momento em que foi realizada a polinização manual cruzada -, não há dados na literatura que
mostrem que a taxa de polinização seja igual em todos os dias de duração da flor.
4.3 – Caracterização da fauna de abelhas polinizadoras do cultivo de tomate
A lista de espécies de abelhas coletadas em flores de tomateiro neste estudo está
apresentada na tabela 7. Foram coletadas abelhas pertencentes às famílias Halictidae, Apidae
gêneros Augochlora e Augochloropsis. Entre as abelhas da família Apidae três foram os
gêneros identificados, sendo eles Exomalopsis, Trigona e Apis. Psaenythia foi o único gênero
da família Andrenidae identificado.
Tabela 7 – Identificação em nível de gênero das abelhas coletadas nas flores de tomate (Lycopersicum esculentum Mill).
Família Subfamília Tribo Gênero Nº de indivíduos
coletados
Halictidae Halictinae Augochlorini Augochlora 19
Augochloropsis 8
Apidae Apinae Apini Exomalopsis 6
Trigona 6
Apis 2
Andrenidae Panurginae Protandrenini Psaenythia 1
As abelhas que podem ser consideradas polinizadoras efetivas das flores do tomate são
as que possuem a capacidade de vibrar as anteras poricidas para que haja a liberação dos
grãos de pólen. Entre as abelhas amostradas neste estudo esse comportamento é observado
entre espécies dos gêneros Augochlora, Augochloropsis, Exomalopsis e Psaenythia. Essas
espécies são frequentemente relatadas como visitantes florais e polinizadores importantes de
diversas espécies de Solanaceae como Solanum palinacantum (CARVALHO et al., 2001),
Solanum stramonifolium (BEZERRA e MACHADO, 2003), Solanum lycocarpum
(OLIVEIRA-FILHO e OLIVEIRA, 1988), Solanum paniculathum (FORNI-MARTINS et al.,
1998) e Solanum viarum (PATRICIO, 2007; MONTAGNANA et al., 2010). Psaenythia
(Andrenidae) foi registrada visitando as flores de Solanum rostratum, em estudo de Bowers
(1975). Abelhas do gênero Trigona e Apis mellifera, apesar de visitarem as flores do tomate
não possuem a capacidade de realizar a polinização por vibração, sendo consideradas abelhas
pilhadoras, ou seja, que coletam o pólen espalhado nas anteras e pétalas pela atividade de
outras abelhas (CARVALHO et al., 1998; BEZERRA e MACHADO, 2003). As abelhas
foram observadas visitando as flores do tomate entre às 8h e 16h, com pico das atividades
entre às 9h e 10h, principalmente as pertencentes a família Halictidae que consistiram nas
mais abundantes. A partir das 13h houve uma queda na frequência de visitas, cessando às 17h.
Padrão semelhante foi encontrado por Montagnana et al. (2010) em flores de juá (Solanum
As abelhas dos gêneros Augochlora e Augochloropsis prendem-se ao cone de anteras
utilizando tanto as pernas como as mandíbulas, em seguida curvavam o abdômen sobre o cone
e nessa postura realizam movimentos vibratórios para a liberação do pólen. Em seguida,
prendendo-se às anteras através das pernas medianas e posteriores, realizam a “limpeza” da
superfície ventral levando o pólen até as escopas (Figura 9). Comportamentos semelhantes
também foram observados por Forni-Martins et al. (1998) em Solanum paniculathum L. e por
Oliveira-Filho e Oliveira (1988) em Solanum lycocarpum.
Figura 9 – Abelha da família Halictidae na flor do tomate (Lycopersicum esculentum Mill). (A) Indivíduo com o abdômen curvado realizando vibração das anteras, (B) Realizando “limpeza” da superfície ventral do corpo.
Macias-Macias et al. (2009) ao estudarem a polinização de tomate no México, também
encontraram as abelhas da família Halictidae e gênero Exomalopsis (Apidae) como as abelhas
nativas mais comuns no cultivo. Os autores avaliaram a eficiência da polinização por
Augochloropsis, Exomalopsis e Apis mellifera através do aumento do peso dos frutos e do
número médio de sementes por fruto. Os resultados encontrados por esses autores mostraram
que as flores polinizadas por Augochloropsis e Exomalopsis produziram frutos de melhor
qualidade se comparadas as flores polinizadas por Apis mellifera. Assim, os autores concluem
que as abelhas solitárias (por exemplo, espécies da família Halictidae) são polinizadores mais
eficientes que as espécies sociais (por exemplo, Apis mellifera), mas os autores argumentam
que para uma polinização adequada dos cultivos é necessário que se mantenha comunidades
de abelhas diversificadas e populações abundantes, sendo para isso imprescindível a
preservação de habitats de nidificação e alimentação adequados para estas abelhas.
5 - CONCLUSÃO
A área de estudo está inserida numa paisagem completamente alterada devido,
principalmente, a atividade agrícola canavieira, o que resultou em uma paisagem homogênea
onde pouquíssimos foram os remanescentes de vegetação nativa encontrados. Uma paisagem
como a encontrada representa uma grande ameaça às populações locais de abelhas, uma vez
que grandes monoculturas ou extensas áreas de pastagens apresentam-se como barreiras ao
deslocamento desses indivíduos. Muitas espécies, notadamente aquelas da família Halictidae
apresentam raio de vôo restrito a poucas centenas de metros em torno de seus ninhos.
O cultivo de tomate estudado apresentou uma baixa porcentagem de frutos formados
por polinização aberta, quando comparado aos demais tratamentos, e as características da
paisagem de entorno, como a ausência de fragmentos florestais em maior quantidade e mais
próximos, são um dos motivos do déficit na polinização desse cultivo, corroborando com a
hipótese levantada. Mesmo em condições adversas, os resultados de qualidade dos frutos, que
nas flores sujeitas a polinização aberta foram maiores, formando frutos mais avermelhados,
com maior número de sementes e polpa mais consistente, indicando os benefícios que a
polinização por essas abelhas pode trazer ao cultivo.
É importante que estudos complementares sejam realizados em outros contextos de
paisagens, onde outros testes sejam empregados como, a suplementação de pólen e a
suplementação de polinizadores na área do cultivo, bem como estudos mais detalhados em
nível de paisagem, com caracterizações das abelhas encontradas nos fragmentos florestais,
6 - BIBLIOGRAFIA
AIZEN, M. A. et al. How much does agriculture depend on pollinators? Lessons from long-term trends in crop production, Annals of Botany, n. 103, p. 1579-1588, 2009.
BAPTISTA, A. P. M. et al., Toxicidade de produtos fitossanitários utilizados em citros para Apis mellifera, Ciência Rural, v. 39, n. 4, p. 955-961, 2009.
BINS, L. S.; FONSECA, L. M. G.; ERTHAL, G. J.; II, F. M. Satellite imagery segmentation: a region growing approach. In: Simpósio Brasileiro de Sensoriamento Remoto. 8, Salvador. Anais... Salvador: INPE, 1996. p.221-223, 1996.
BEGON, M.; TOWNSEND, C. R. e HARPER, J. L. Ecologia: de indivíduos a ecossistemas. Tradução de A. S. Melo et al.; Revisão Técnica de P. L. de Oliveira. 4 ed. Artmed. Porto Alegre. 752 p. 2007.
BEZERRA, E. L. S. e MACHADO, I. C. Biologia floral e sistema de polinização de Solanum stramonifolium Jacq. (Solanaceae) em remanescente de mata atlântica, Pernambuco. Acta. Bot. Bras., v. 17, n. 2, p. 247-257, 2003.
BLAUMGARTNER, D. L. e ROUBIK, D. W., Ecology of Necrophilous and Filth-Gathering stingless bees (Apidae:Meliponae) of Peru, Journal of the Kansas Entomological Society, vol. 62, n. 1, p. 11-22, 1989.
BOWERS, K. A. W. The pollination ecology of Solanum rostratum (Solanaceae). American Journal of Botany, v. 62, n. 6, p. 633-638, 1975.
BRASIL, Produção agrícola municipal: culturas temporárias e permanentes 2002, Instituto Brasileiro de Geografia e Estatística – IBGE, v. 29, 85p, 2002.
BROSI, B. J., The complex responses of social stingless bees (Apidae:Meliponini) to tropical deforestation, Forest Ecology and Management, vol. 258, p. 1830-1837, 2009.
BUCHMANN, S. L. e NABHAN, G. P. The Forgotten Pollinators, Island Press,
Washington, D. C. Shearwater Books, Covelo, California, 1996.
CÂMARA, G.; SOUZA, R. C. M.; FREITAS, U. M.; GARRIDO, J. SPRING: integrating remote sensing and GIS by object-oriented data modelling. Comput Graph, v. 20. p. 395-403,1996.
CARVALHO, C. A. L. et al. Comportamento forrageiro de abelhas (Hymenoptera, Apoidea) em flores de Solanum palinacanthum Dunal (Solanaceae), Revista Brasileira de Zoociências, v. 3, n. 1, p. 35-44, 2001.
CEPAGRI – Centro de Pesquisas Meteorológicas e Climáticas Aplicadas à Agricultura, Clima dos Municípios Paulistas, Campinas. Disponível em:<www.cpa.unicamp.br/outras-informacoes/clima_muni_494.html> Acesso em: 5 ago. 2010.
CRUZ, D. O. Biologia floral e eficiência das abelhas Apis mellifera L. (Campo aberto) e Melipona quadrifasciata Lep. (Ambiente protegido) na cultura da pimenta malagueta (Capsicum frutescens L.) em Minas Gerais, Brasil. 2009. 83f. Tese (Doctor Scientiae) – Universidade Federal de Viçosa, Viçosa, 2009.
DAFNI, A. Pollination ecology. Oxford University Press, Oxford, 738p., 1992.
DOMINGUES, E.T. e NETO, A. T. Influência da polinização e da morfologia floral na frutificação de variedades de laranja-doce, Scientia Agrícola, v. 56, n. 1, p. 163-170, 1999.
FAO, A Contribution to the International Iniciative for the Conservation and Sustainable Use of Pollinators, Rapid Assessment of Pollinators’ Status, 52 p., 2008.
FAO, State of the world’s forests 2009, 152f. Roma, 2009.
FERREIRA, B. O efeito do contexto da paisagem e da estrutura de habitat sobre abelhas e vespas silvestres em fragmentos de cerrado. 2010. 96f. Dissertação (Mestrado em Ciências Biológicas) – Instituto de Biociências, Universidade Estadual Paulista, Rio Claro, 2010.
FORNI-MARTINS, E. R. et al. Biologia floral e reprodução de Solanum paniculatum L. (Solanaceae) no estado de São Paulo, Brasil, Revista Brasileira de Botânica, v. 21, n. 2, p. 117-124, 1998.
FREITAS, L. et al. Ocorrência de limitação polínica em plantas de Mata Atlântica, Oecologia Australis, v. 14, n. 1, p. 251-265, 2010.
FREITAS, B. M. e ALVES, J. E. Efeito do número de visitas florais da abelha melífera (Apis mellifera L.) na polinização da goiabeira (Psidium guajava L.) cv. Paluma, Rev. Ciên. Agron., v. 39, n. 1, p. 148-154, 2008.
GALLAI, N. et al. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline, Ecological Economics, n. 68, p. 810-821, 2009.
GAMITO, L. M. e MALERBO-SOUZA, D. T. Visitantes florais e produção de frutos em cultura de laranja (Citrus sinensis L. Osbeck), Acta Scientiarum. Animal Sciences, v. 28, n.
4, p. 483-488, 2006.
GEMMILL-HERREN, B. e OCHIENG, A. O. Role of native bees and natural habitats in eggplant (Solanum melongena) pollination in Kenya, Agriculture, Ecosystems and Environment, n. 127, p. 31-36, 2008.
GOULSON, D., Effects of introduced bees on native ecosystems, Annual Review of Ecology, Evolution and Systematics, v. 34, p. 1-26, 2003.
GREENLEAF, S. S. e KREMEN, C., Wild bee species increase tomato production and respond differently to surrounding land use in Northern California, Biological Conservation, v. 33, p. 81-87, 2006.
INFOCLIMA, Boletim de Informações Climáticas do CPTEC/INPE, MCT/INPE/CPTEC, ano 17, n. 18, 2010.
KIM, J. et al. Effects of Cultivation and Proximity to Natural Habitat on Ground-nesting Native Bees in California Sunflower Fields, Journal of the Kansas Entomological Society, v. 74, n. 4, p. 309-320, 2006.
KNIGHT, T. M. et al. Pollen limitation of plant reproduction: Pattern and Process, Annual Review of Ecology, Evolution and Systematics, n. 36, p. 467-497, 2005.
KREMEN, C. et al. The area requirements of an ecosystem service: crop pollination by native bee communities in California, Ecology Letters, n. 7, p. 1109-1119, 2004.
KREMEN, C. et al. Pollination and other ecosystem services produced by mobile organisms: a conceptual framework for the effects of land-use change, Ecology Letters, n. 10, p. 299-314, 2007.
MACIAS-MACIAS, O. et al. Contribution of native bees and africanized honey bees (Hymenoptera:Apoidea) to Solanaceae crop pollination in tropical México, J. Appl. Entomol., n. 133, p. 456-465, 2009.
MCGARIGAL K., MARKS B.J. FRAGSTATS: Spatial Pattern Analysis Program for Qunatifying Landscape Structure. Forest Science Department, Oregon State University, Carvallis, 1995.
MODRO, A. F. H. et al. Conhecimento dos moradores do médio Araguaia, Estado do Mato Grosso, sobre a utilidade de produtos de abelhas (Hymenoptera, Apidae), Acta Scientiarum. Biological Sciences, v. 31, n. 4, p. 421-424, 2009.
MONTAGNANA, P. C. et al. Comportamento forrageiro de abelhas visitantes das flores de juá (Solanum viarum Dunnal., Família Solanácea). In: IX Encontro sobre Abelhas, n. 9, 2010, Ribeirão Preto, Anais do IX Encontro sobre Abelhas, Ribeirão Preto: FUNPEC-Editora, 2010. CD
MONTEMOR, K. A. e SOUZA, D. T. M. Biodiversidade de polinizadores e biologia floral em cultura de berinjela (Solanum melongena), Zootecnia Tropical, v. 27, n. 1, p. 97-103, 2009.
NAIKA, S. et al. A cultura do tomate: produção, processamento e comercialização; Fundação Agromisa e CTA, Wageningen, 2006.
NICODEMO, D. Características florais e dependência por polinizadores de cinco cultivares de pepino e manejo de colméias em estufas. 2008. 89f. Tese (Doutorado em Zootecnia) – Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2008.
ODUM, E. P. e BARRETT, G. W. Fundamentos de Ecologia. 5 ed. Tradução de Pégasus Sistemas e Soluções. São Paulo, 2008.
OLIVEIRA-FILHO, A. T. e OLIVEIRA, L. C. A. Biologia floral de uma população de Solanum lycocarpum St. Hil. (Solanaceae) em Lavras, MG, Revista Brasileira de Botânica, v. 11, p. 23-32, 1988.
PATRICIO, G. B. Efeito da heterogeneidade de habitats sobre as interações planta-polinizador em uma região da bacia do Rio Corumbataí-SP. 2007. 67f. Dissertação (Mestrado em Ciências Biológicas) – Instituto de Biociências, Universidade Estadual Paulista, Rio Claro, 2007.
ROUBIK, D. W. e VILLANUEVA-GUTIÉRREZ, R. Invasive Africanized honey bee impact on native solitary bees: a pollen resource and trap nest analysis, Biological Journal of the Linnean Society, n. 98, p. 152-160, 2009.
SANTOS, S. A. B. et al. Pollination of tomatoes by the stingless bee Melipona quadrifasciata and the honey bee Apis mellifera (Hymenoptera, Apidae), Genetics and Molecular Research, n. 8, v. 2, p. 751-757, 2009.
SANTOS, S. A. B. et al. Pollination of Cucumber, Cucumis sativus L. (Cucurbitales: Cucurbitaceae), by the Stingless Bees Scaptotrigona aff. depilis Moure and Nannotrigona testaceicornis Lepeletier (Hymenoptera: Meliponini) in Greenhouses, Neotropical Entomology, v. 37, n. 5, p. 506-512, 2008.
SOUZA, R. M. et al. Requerimentos de polinização do meloeiro (Cucumis melo l.) no município de Aracajú – CE – Brasil, Revista Caatinga, v. 22, n. 1, p. 238-242, 2009.
SIQUEIRA, K. M. M. et al. Ecologia da polinização do maracujá-amarelo, na região do vale do submédio São Francisco, Revista Brasileira de Fruticultura, v. 31, n. 1, p. 1-12, 2009.
SKERL, M. I. S. et al. Residues of Pesticides in Honeybee (Apis mellifera carnica) Bee Bread and in Pollen Loads from Treated Apple Orchards, Bull Environ Contam Toxicol, v. 83, p. 374-377, 2009.
SOUSA, R. M. et al. Requerimentos de polinização do meloeiro (Cucumis melo l.) no município de Acaraú – CE – Brasil, Revista Caatinga, v. 22, n. 1, p. 238-242, 2009.
STEPHENSON, A. G. Flower and fruit abortation: proximate causes and ultimate functions, Annual Review of Ecology and Systematics, v. 12, p. 253-279, 1981.
THOMSON, D. Competitive interactions between the invasive European honey bee and native bumble bees, Ecology, v. 85, n. 2, p. 458-470, 2004.
THOMSON, J. D. Using pollination deficits to infer pollinator declines: Can theory guide us? Conservation Ecology, v. 5, n. 1, 2001. Disponível em <http://www.consecol.org/vol5/iss1/art6/> Acesso em 11/Mar/2010.
VALDOVINOS-NUNEZ, G. R. et al., Comparative Toxicity of Pesticides to Stingless Bees (Hymenoptera: Apidae: Meliponini), Journal of Economic Entomology, v. 102, n. 5, p. 1737-1742, 2009.
VIANNA, M. R., et al. Manejo de polinizadores e o incremento da produtividade agrícola: uma abordagem sustentável dos serviços dos Ecossistemas, Revista Brasileira de Agroecologia, v. 2, n. 1, p. 144-147, 2007.
VAISSIÈRE. B. E. et al. Protocol to Detect and Asses Pollination Deficits in Crops, Versão 1, Global Action on Pollination Services for Sustainable Agriculture, 26f. FAO, Roma, 2009.