Ismênia Gurgel Martins
Padrão de atividades do sagüi
Callithrix jacchus numa área de
Caatinga
Divisão de Serviços Técnicos
Catalogação da Publicação na Fonte UFRN/Biblioteca Central Zila Mamede
Martins, Ismênia Gurgel.
Padrão de atividades do sagüi callithrix jacchus numa área de
caatinga / Ismênia Gurgel Martins. – Natal, RN, 2007. 56 f
Orientadora : Maria de Fátima Arruda.
Dissertação (Mestre). Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Sagüi – callithrix jacchus - Dissertação. 2. Comportamento
animal – Dissertação. 3.Caatinga – Dissertação. I. Arruda, Maria de Fátima. II. Título.
PADRÃO DE ATIVIDADES DO SAGUI
Callithrix jacchus
NUMA ÁREA DE CAATINGA
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre do Programa de Pós-Graduação em Psicobiologia.
Orientadora: Profª. Maria de Fátima Arruda ([email protected]).
NUMA ÁREA DE CAATINGA
Autora: Ismênia Gurgel Martins
Data da defesa: 11 de setembro de 2006
Banca Examinadora:
___________________________________________________ Profª Simone Almeida G. L. costa
Universidade do Estado do Rio Grande do Norte, RN.
___________________________________________________ Profº Arrilton Araújo
Universidade Federal do Rio Grande do Norte, RN.
___________________________________________________ Profª Maria de Fátima Arruda
Sou grata a Deus pelo profundo mecanismo da criação, da evolução e da prosperidade de todas as coisas e por ter me dado a permissão de ter chegado ate aqui.
A meus pais Rita Gurgel e Francisco Sales pela vida, em especial à minha mãe, por ser uma mulher maravilhosa, forte e presente em todos os momentos de minha vida.
A Fátima Arruda, por ter aceitado em me orientar no estudo do Comportamento Animal, obrigada pela confiança e ajuda na minha vida acadêmica.
Simone, grata estou com sua presença em mais uma etapa da minha vida acadêmica, pois tudo começou com o nosso trabalho na especialização. Você é responsável por este passo à frente e aqui estou concluindo mais um capítulo.
A você Arrilton, obrigada por mais estar mais uma vez contribuindo com o meu trabalho e no meu crescimento profissional.
Aos primatas Neotropicais que vivem no IBAMA-AÇU, que além de terem contribuído para a existência deste trabalho me proporcionou momentos maravilhosos de superação.
Ao Filósofo Damião Dantas e família e ao “irmão” José Fernandes e família pela confiança em ajudar-me sempre, pelo cuidado carinho e compreensão em todas as etapas da minha pesquisa. Aos funcionários Canindé e “Chiquinho” pela amizade e apoio.
Meus agradecimentos a Universidade Federal do Rio Grande do Norte, em especial aos Professores do Departamento de Fisiologia pelas informações valiosas e ao IBAMA pelo apoio e infra-estrutura na realização deste estudo.
Aos meus irmãos Dário e Érico que são tudo pra mim e sempre estiveram do meu lado, em especial a Dário que foi o maior incentivador desta jornada no meu aprimoramento acadêmico.
Ao meu noivo e amigo Paulo Henrique, pelo amor, otimismo e paciência nos momentos difíceis dessa longa caminhada.
Aos colegas de classe Fabiana, Yalkiria, Ernani, Fernando, que se mostraram verdadeiros amigos. A Val e a Eric pelas informações e ajudas valiosas, meu muito obrigada.
As minhas verdadeiras amigas da UERN Cristiane, Camila e Alininha, pela ajuda no trabalho de campo, força e companhia nos momentos de incertezas.
Callithrix jacchus é um primata da família Callitrichidae que vive em grupos sociais.
Pode ser encontrado em vários tipos de habitat, cujas fisionomias e comunidades florísticas distintas estão intrinsecamente relacionadas com a ecologia comportamental e relações sociais. Nosso objetivo foi descrever as relações sociais e hábitos alimentares do sagüi Callithrix jacchus no Bioma Caatinga. Observamos um grupo de
sagüi na Floresta Nacional de Açu-IBAMA/RN, composto no início do estudo por cinco animais adultos, 2 fêmeas e 3 machos. Foi registrado o nascimento de 5 filhotes ao longo do estudo. A técnica de amostragem foi o animal focal, ao longo de 11 meses, uma vez/semana, com registro das seguintes categorias comportamentais: catação social, contato, proximidade, forrageio, ingestão, locomoção e descanso. O forrageio foi a atividade mais realizada pelos animais, sendo significativamente mais freqüente na seca. Já a catação social, foi significativamente mais elevada no período chuvoso, e com os maiores índices entre os reprodutores. Esse mesmo padrão foi observado com relação a proximidade. O alimento mais ingerido pelo grupo foi o exsudado, sendo mais expressivo no período chuvoso. A espécie vegetal mais procurada foi o limão Cirus limon e a jurema branca Pitecolobium foliolosum. Quando
Callithrix jacchus, as the other species in the family Callitrichidae, lives in social
groups. The groups cam be found in different habitats, whose distinct floristic physiognomies and communities are intrinsically related to their behavioral ecology and social relations. Our objective was to describe the social relations and feeding behavior of Callithrix jacchus in the Bioma Caatinga. We observed a group at the
FLONA (National Forest – IBAMA), in Açu-RN, in northeastern Brazil, compose of five adults (2 females and 3 males) at the beginning of the study. The birth of five animals was registered along the study. The following behavioral categories were registered along eleven months, once a week, through instantaneous focal animal sampling: social grooming, contact, proximity, foraging, feeding, locomotion and rest. Foraging presented the highest levels comparing to other activities, and was more frequent in the dry season. Social grooming was the second more frequent activity, with higher levels in the rainy season, and between the reproductive couple. We found similar results for proximity. The most explored feeding item was the gum, specially in the rainy season. The most explored species for exudates feeding were Cirus limon
(limão) and Pitecolobiun foliolosum (jurema branca). The comparision of fruit and
1. INTRODUÇÃO...
1.1
- Padrão de atividade...1.2
- Comportamento alimentar...1.3
- Interações sociais...1.4
- O bioma caatinga...1 3 4 6 8
2. OBJETIVOS... 2.1 – Geral... 2.2 – Específicos...
10 10 10
3. MÉTODO... 3.1 - Local de estudo... 3.2 - Grupo de Estudo... 3.3 - Captura e Marcação dos Animais... 3.4 - Observação Comportamental... 3.5 - Registro de Utilização da Área... 3.6 - Análise Estatística dos Dados...
11 11 14 16 16 18 18
4. RESULTADOS... 4.1 - Padrão de Atividade do Grupo... 4.2 - Padrão de ingestão do grupo... 4.3 - Utilização da área... 4.4 - Interação Social...
21 21 23 27 28
5. DISCUSSÃO... 30
6. CONCLUSÕES... 43
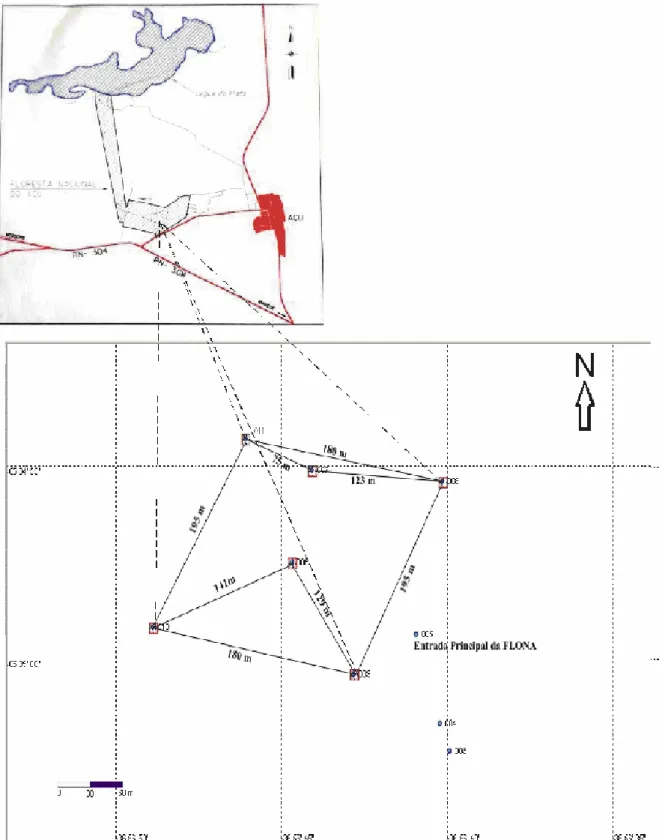
Figura 01 Mapa da região oeste do Estado do Rio Grande do Norte, destacando a área da Flona-Açu (IBAMA) no município de Açu-RN...12
Figura 02 Representação da área utilizada no estudo (IBAMA/Flona-Açu)...13
Figura 03 Grupo Bosque: B1, fêmea subordinada forrageando, com a metade da cauda inferior pintada; B2, fêmea reprodutora alimentando-se com inseto, com os tufos tingidos; B3, macho reprodutor com colar de identificação (seta); B4, macho adulto subordinado com colar de identificação (seta); B5, macho juvenil (início do estudo) em repouso, com as patas posteriores e metade da cauda superior tingida...15
Figura 04 Representação da área de uso de Callithrix jacchus, na FLONA-Açu. O
Ponto 005 representa a Entrada Principal da FLONA. O Ponto 006 representa o limite direito inferior da área de uso. O Ponto 007 representa o final da trilha 1. O Ponto 008 representa o final da trilha 2. O Ponto 009 representa o limite esquerdo inferior da área de uso. O Ponto 010 representa o limite esquerdo superior da área de uso. O Ponto 011 representa o limite direito superior da área de uso. Toda essa área é composta de vegetação nativa da caatinga. POMAR: área com árvores frutíferas e pouca vegetação nativa da caatinga, situada nos limites inferior dos pontos 006 a 011...20
Figura 05 Padrão de atividade de Callithrix jacchus em ambiente natural (Bioma
Caatinga) no período de março/05 a fevereiro/06...22
Figura 06 Padrão de atividade de Callithrix jacchus em ambiente natural (Bioma
níveis significativamente mais elevados...22 Figura 07 Perfil alimentar de Callithrix jacchus em ambiente natural (Bioma
Caatinga), no período de mar/2005 a jan/2006...24 Figura 08 Perfil alimentar de Callithrix jacchus em ambiente natural (Bioma
Caatinga), durante a estação chuvosa (mar-jul/2005) e seca (ago/2005-jan/2006)...24 Figura 09 Árvores de exsudado e fruto utilizados na dieta de Callithrix jacchus
em ambiente natural (Bioma Caatinga), no período de mar/2005 a jan/2006...26 Figura 10 Espécies vegetais utilizadas para a gomivoria de Callithrix jacchus em
ambiente natural (Bioma Caatinga), no período de março/05 a janeiro/06...26 Figura 11 Árvores de frutos utilizados na dieta de Callithrix jacchus em ambiente
natural (Bioma Caatinga), no período de mar/2005 a jan/2006...27 Tabela I Composição do Grupo Callithrix jacchus estudado (FEV/2006)...14
1. INTRODUÇÃO
Os aspectos comportamentais e morfológicos dos primatas são produtos do processo evolutivo que selecionou as características que beneficiaram a aptidão dos indivíduos (Strier, 1999). A aptidão caracteriza-se pela representação gênica que os membros de uma geração parental contribuem para a descendência futura.
Cada indivíduo empenha-se em elevar ao máximo sua contribuição genética às gerações seguintes, empenho que pode ser medido através do esforço reprodutivo individual (Strier, 1999). Diante dessa condição, fêmeas e machos desenvolvem ações que visam aumentar a probabilidade de sobrevivência de seus filhos e de gerar outros (Alcock, 2001).
Os primatas apresentam um longo período de imaturidade, implicando em grande investimento parental (Webb, Houston, McNamara & Székely, 1999). Na maioria dos primatas, a mãe é o principal responsável pelo cuidado no princípio de vida dos filhotes. Posteriormente, há grande variabilidade no que diz respeito à participação dos membros do grupo social nessa atividade.
Em primatas, a evolução de grupos multi-machos e multi-fêmeas foi associada à evolução de altos níveis de tolerância social (Blanckenhorn, 1990). Os grupos sociais mostram coesão duradoura, coordenação das atividades entre os membros do grupo e separação espacial em relação a indivíduos extra-grupo (Fedigan, 1992). Dentro das sociedades dos primatas, alguns indivíduos, especialmente os machos adolescentes deixam seu grupo para juntar-se a outros. Muitos indivíduos são membros permanentes do grupo e mostram hostilidade a estranhos. Algumas espécies formam associações temporárias e viajam em pequenos bandos. O que refletem suas características comportamentais e ecológicas, tais como, sistema de acasalamento, diferença entre sexos, habitat e alimentação (Fedigan, 1992).
Dentre as diversas famílias da Ordem Primata, a Callitrichidae é atualmente
composta pelos gêneros Cebuella, Callithrix, Saguinus, Leontopithecus, Callimico e
Mico (Rylands et al., 2000). Estes apresentam massa corporal variando de 119 a
750g (Leutenegger, 1973; Yamamoto, 1991; Araújo et al, 2000), cauda longa e não
Algumas características morfológicas e comportamentais desses primatas são
a geração de gêmeos, exceto em Callimico, ausência do terceiro molar, órbita ocular
presa no crânio, modificação das unhas, manutenção da forma preênsil do hálux e a redução no tamanho do corpo (Stevenson & Rylands, 1988), que se apresentam como resultado de um processo de especialização ecológica.
Podemos encontrar Callithrix ocupando diferentes habitats, como a costa
Atlântida do Brasil, a Caatinga do nordeste do Brasil e o Cerrado central do Brasil (Stevenson & Rylands, 1988; Coimbra-Filho, 1984). Colonizar um novo habitat depende da distribuição e abundância espacial e temporal dos recursos alimentares, bem como da disponibilidade de uma vegetação que forneça abrigo contra predadores e diminua a competição, especialmente com outros primatas.
As espécies de Callitrichidae se alimentam de três itens básicos - frutos, insetos e goma, podendo incluir na dieta flores, brotos, ovos de aves e pequenos vertebrados (Sussman & Kinzey, 1984; Rylands & Faria, 1993). Cada um desses itens alimentares requer diferentes estratégias de forrageio e técnicas de exploração.
Callithrix, Mico e Saguinus ocupam preferencialmente nichos insetívoros e
frugívoros em manchas de matas secundárias ou fragmentadas; Leontopithecus
ocorre em florestas primárias, alimentando-se de presas e frutos; Cebuella ocupa
nichos insetívoros e gomívoros de florestas inundadas (Rylands, 1996). Callimico
são comumente encontrados em florestas secundárias, explorando os níveis baixos da mata próximo ao solo, alimentando-se basicamente de invertebrados e frutos (Fleagle, 1988).
Os gêneros Callithrix e Cebuella desenvolveram dentição que lhes permite
fazer furos em árvores para extração de exsudado. (Rylands & Faria, 1993). Por
outro lado, Saguinus, Leontopithecus e Mico apresentam caninos inferiores maiores e
1.1 - Padrão de atividade
O padrão de atividade representa a descrição quantitativa das respostas a uma variedade de fatores que influenciam a sobrevivência e o sucesso reprodutivo (Defler, 1995). As características individuais que podem influenciar no padrão de atividade incluem sexo, idade, peso e estado fisiológico (Fragaszy, 1990, Miller & Dietz, 2005). As características do grupo incluem o tamanho e a composição e as características ambientais incluem o risco de predação e disponibilidade de recursos (Fragaszy, 1990).
Os primatas vivem em ambientes complexos e variáveis. O tamanho da área de uso entre populações da mesma espécie e diferentes espécies pode ser compreendido com base na distribuição dos recursos alimentares, ciclos sazonais e áreas de dormir. Assim muitos primatas restringem suas atividades a ambientes particulares, que freqüentemente são englobadas em quatro categorias principais: descaso, deslocamento, alimentação e atividade social (Dunbar, 1988).
Hawkins (1992), estudando o padrão diário de um grupo de macaco de cheiro
Saimiri sciureus, observou que o deslocamento dos animais apresenta uma relação
com as condições climáticas. Nos dias de sol, os animais se deslocavam mais que nos dias chuvosos e o aumento da temperatura também aumentava a freqüência de
deslocamento. O estudo com um grupo de bugios pretos Allouata caraya, mostrou
resultados semelhantes, quanto maior a temperatura média diária, maior a percentagem de tempo dedicado à alimentação e maior a locomoção (Bicca-Marques,
1993). Wallace (2005), observando o padrão sazonal de Ateles chamek, verificou que
em geral os indivíduos gastam 75-90% do seu tempo forrageando e alimentando-se de frutas e o restante do dia passam realizando as demais atividades comportamentais.
A distribuição das atividades de Callithrix jacchus, ao longo do dia segue um
(Alonso & Langguth, 1989). O tempo gasto alimentando-se de frutos, flores ou exsudados gradualmente diminui de manhã e aumenta durante à tarde. O padrão de atividades para a espécie não é rígido e pode sofrer alterações em função das condições bióticas e abióticas do habitat no qual determinada população está inserida, assim como da dinâmica social.
Digby & Barreto (1996) estudaram três grupos de C. jacchus, um que habitava
uma área de plantação experimental com características de pomar e dois que habitavam uma área de mata atlântica secundária. O padrão de atividades foi equivalente nos três grupos, não tendo sido encontradas diferenças entre os registros de descanso, locomoção e comportamentos sociais entre os grupos. Apenas, foram registrados níveis reduzidos de forrageio e alimentação na área de plantação experimental com relação aos demais grupos. Nascimento (1997), estudando um
grupo de C. jacchus, que habitava uma área de mata atlântica comparou o padrão
geral de atividade do grupo após o nascimento de três proles consecutivas. Pode-se verificar que as atividades que mais se destacaram ao longo dos três acompanhamentos foram o parado e o forrageio.
Observando o comportamento em grupos selvagens de C. jacchus, Cavalcanti
(2002) verificou que os perfis de atividades diárias foram semelhantes ao longo de todo o estudo. A autora destaca que a atividade que ocupou a maior parte do tempo dos indivíduos desses grupos foi o forrageio (com valores médios em torno de 50%), ingestão (15%), embora tenha variado bastante em função do mês e do grupo estudado. A seguir, destacou-se a atividade social, com valores médios de 13,6%. A variação de tempo gasto em cada atividade ocorreu em função da estrutura e composição ambiental, que mostrou uma variação no padrão geral para os três grupos estudados, o qual influenciou a atividade social, sendo mais elevada quando o forrageio e a ingestão de alimentos foi reduzida.
1.2 - Comportamento alimentar
animais deverão estar envolvidos na maior parte do seu período de atividade na procura e ingestão do alimento.
O tipo de dieta determina quanto o animal deverá investir na procura e aquisição do alimento a fim de equilibrar os custos e benefícios relacionados com o forrageio. Também está relacionado com outros aspectos do padrão de atividade diária, entre eles, o tamanho da área de uso, área diária percorrida e tamanho do grupo (Fleagle, 1988).
Investigação sobre os compostos químicos presentes na dieta dos primatas tem promovido um entendimento único nas estratégias de forrageio, facilitando a
avaliação das explicações socioecológicas da organização social (Chapman et al,
2003). Esses autores investigando a variação nos valores nutricionais dos alimentos dos primatas sugerem que há considerável diversidade dos nutrientes ingeridos por
Colobus guerezai entre as árvores e área visitada.
Todas as espécies de primatas diurnos são forrageadores sociais e viajam na busca de recursos como membros de um grupo social, no qual os indivíduos empregam diferentes estratégias para localizar áreas de alimentação (Bicca-Marques & Garber, 2005). Os diferentes tipos de alimento ingeridos pelos primatas indicam as estratégias de forrageamento. Assim dentro de uma categoria alimentar, tais como frugivoria, diferentes espécies podem apresentar diversas estratégias de forrageamento dentro de um mesmo tipo de habitat (Garber, 1987). Estas diferenças nas áreas de alimentação demonstram as ricas adaptações dos primatas.
A utilização dos recursos está relacionada em parte ao tamanho corporal e das necessidades metabólicas e nutricionais (Strier, 1999). Pequenos animais apresentam altas taxas metabólicas, necessitando de uma dieta rica em nutrientes e altamente energética por unidade de peso (Garber, 1987).
primatas em decorrência dos custos associados com a reprodução, gestação interna e a lactação (Strier, 1999). Dessa forma é importante um balanço adequado dos nutrientes/energia, cuidado aos filhotes e estratégias eficientes de ingestão alimentar.
O acesso ao alimento pelos calitriquideos aponta para uma diferença intersexos, apresentando a fêmea uma tendência ao acesso preferencial aos itens
alimentares (Maier et al, 1982; Box, 1997), tendo em vista que a reprodução em
calitriquídeos é bastante custosa para fêmeas reprodutoras. As fêmeas reprodutoras do Callithrix jacchus, em ambiente natural são agressivas com os demais do grupo,
no que se refere a manter a prioridade de acesso alimentar (Araújo, 1996). Isto pode ser visto como uma estratégia para manter seu desempenho reprodutivo, uma vez que a disponibilidade de recursos exerce uma influência significativa nas variações de fecundidade das fêmeas.
Cavalcanti (2002), analisando o comportamento alimentar de três grupos de C.
jacchus, concluiu que essa espécie apresenta perfil exsudatívoro-frugívoro em
ambientes que quase não sofreram ação antrópica, em função de sua flexibilidade de dieta. O comportamento de modo geral, torna-se frugívoro-insetívoro em ambientes degradados, podendo apresentar variação nas estratégias de busca do alimento de acordo com mudanças no tipo de recurso.
1.3 - Interações sociais
O estudo do comportamento afiliativo, como a catação, tem sido descrito em algumas espécies. Para o indivíduo, a catação mantém e estabelece a relação social,
reduz a tensão e previne a agressão (Lazaro-Perea et al, 2000).
Algumas características da organização social dos primatas são refletidas no
padrão de catação. Ahumada (1992), estudando um grupo de Ateles geoffroyi em
os juvenis apresentam um longo período de dependência, os machos são filopátricos e a dispersão é feita palas fêmeas.
O padrão de catação pode variar dependendo não somente do sistema social, mas também das condições ecológicas que podem afetar a defesa dos recursos em diferentes espécies. O estudo de duas populações de babuínos com diferentes padrões de relações de dominância mostra que o padrão de catação muda dentro das espécies
quando o balanço entre subordinados e dominantes variam (Barrett et al, 1999).
Nas espécies de primatas em que as fêmeas migram do grupo natal, a teoria da seleção de parentesco sugere que os machos podem associar-se preferencialmente com outros, sendo mais afiliativos e cooperativos do que com as fêmeas (Fiore & Fleischer, 2005).
Nos grupos sociais, a freqüência e a duração da catação variam de acordo com o sexo, idade, condições reprodutivas, posição hierárquica e grau de parentesco entre os animais. Em grupos de chimpanzés, a catação concentra-se em regiões corporais que são inacessíveis ao próprio animal. Essa atividade promove um benefício biológico essencial que compreende a remoção de ectoparasitas, mas também requer pouco custo energético. Entre os chimpanzés essa atividade atua como uma reciprocidade para futuras cópulas, acesso aos infantes e suporte para o combate (Nishida, 1988). As fêmeas tendem a se acasalar com machos que têm mostrado alta freqüência de afiliação, como a catação. A catação é uma atividade social também
importante em Callithrix jacchus, sendo realizado predominantemente pelo casal
reprodutor (Woodcock, 1978; Albuquerque, 1998). Os animais dominantes são apontados como os principais parceiros dos animais não reprodutivos na catação. Os quais preferem interações sociais com os dominantes como forma de criar laços sociais e permanecer no grupo (Digby, 1995a).
Em grupos familiares de Callithrix jacchus o pai faz mais catação nos filhotes
do que a mãe e os machos tendem a fazer mais catação do que as fêmeas (Silva & Sousa, 1997; Yamamoto, 1991), sendo estas mais freqüentemente receptoras da
catação (Azevedo et al, 2001). As interações de catação da fêmea reprodutora
alteram-se de acordo com seu estado reprodutivo. Durante a gravidez, as freqüências
o estro pós-parto (Woodcock, 1978; Dixson & Lunn, 1987; Barreto, 1996, Sousa, 1999; Busch, 2000).
Em relação ao padrão de catação entre fêmeas de sagüi no mesmo grupo
familiar em ambiente natural, Lazaro-Perea et al (2004) investigaram a relação entre
a estrutura social e a catação entre fêmeas. Constataram que as fêmeas reprodutoras cataram mais as não-reprodutoras e houve baixos índices de agonismo. Os autores sugerem que em grupos com sistemas de reprodução cooperativa, as fêmeas reprodutoras podem usar a catação como um incentivo para as ajudantes ficarem no grupo.
1.4 - O Bioma Caatinga
Ecorregião semi-árida exclusivamente brasileira compreende cerca de 12% do território nacional incluindo as áreas de transição para outros biomas (IBAMA, SEMACE, IDEMA, SUDEMA, IMA, ADEMA & CRA, 2000). A caatinga tem uma fisionomia de deserto, com índices pluviométricos muito baixos, durante o ano. A região da caatinga está submetida a ventos fortes e secos, que contribuem para a aridez da paisagem nos meses de seca.
A vegetação da caatinga é constituída de espécies lenhosas e herbáceas de pequeno porte, geralmente dotadas de espinhos e caducifólias que perde suas folhas no início da estação seca para reduzir a perda da água por transpiração (Sampaio &
Rodal, 2000) e de cactáceas e bromeliáceas (Kiill et al, 2000). A diversidade,
freqüência e dominância das espécies são determinadas pelas variações topográficas, tipo de solo e pluviosidade.
As espécies vegetais da caatinga apresentam três estratos: arbóreo (8 a 12
metros), arbusto (2 a 5 metros) e herbáceo (abaixo de 2 metros) (Kiill et al, 2000). A
flora dos sertões é constituída por espécies de longa história da adaptação ao calor e à seca, é incapaz de reestruturar-se naturalmente se máquinas forem usadas para alterar o solo. A degradação é, portanto, irreversível na caatinga.
seca. Entre estas se destaca o juazeiro, uma das plantas mais típicas desse ecossistema. Ao caírem às primeiras chuvas no fim do ano, a caatinga perde seu aspecto rude e torna-se rapidamente verde e florida. Além das cactáceas, com o
Cereus (mandacaru e facheiro) e Pilocereu (xiquexique), também apresenta muitas
leguminosas (mimosa, acácia e imburana).
Análises da mastofauna da caatinga têm revelado uma baixa incidência de endemismo, com uma mastofauna relativamente restrita a algumas espécies (Willig & Mares, 1989). Os mamíferos são de pequeno porte, sendo os roedores os mais abundantes, e as espécies encontrados em maior número são aquelas que apresentam
comportamento migratório na época da seca (Kiill et al, 2000). Um levantamento
realizado por Oliveira, Gonçalves & Bonvicino (2003) permitiu listar 143 mamíferos
para a região da caatinga, dentre eles espécies de primatas (guariba: Alouatta
belzebue, A. ululata e A. caraya; macaco-prego: Cebus apella; sauá: Callicebus
barbarabrownae; sagüi: Callithrix jacchus e C. penicillata).
Dentre os primatas, C. jacchus pode ser encontrado em vários tipos de habitat,
cujas fisionomias e comunidades florísticas distintas estão intrinsecamente relacionadas com a ecologia comportamental e relações sociais. Há, no entanto, uma lacuna de registro sistemático dessa espécie em vegetação de Caatinga, com relação aos aspectos sócio-ecológicos, tais como o nicho alimentar, reprodução e relações sociais.
2. OBJETIVOS
2.1 - Geral:
- Descrever o padrão de atividades do grupo de sagüi Callithrix jacchus no Bioma
Caatinga.
2.2 - Específicos:
- Registrar as atividades dos indivíduos do grupo da saída da árvore de dormir até o retorno à mesma nas estações seca e chuvosa;
- Registrar as interações sociais intragrupo nas estações seca e chuvosa; - Registrar o comportamento alimentar;
3. MÉTODO
3.1 - Local de Estudo
A Floresta Nacional (FLONA) de Açu (5° 34’ 20” S e 16° 54’ 33” W) é uma Unidade de Conservação Federal (IBAMA), situada no sudoeste do sítio urbano de Açu, na região central do Rio Grande do Norte (figura 01).
A FLONA confronta-se ao norte, leste e oeste com propriedades particulares cujas atividades econômicas são voltadas para a pecuária, especificamente a caprinocultura, e ao sul, com a Avenida Senador João Câmara, localizada no Bairro Alto de São Francisco na cidade do Açu.
A Unidade em questão foi criada na década de 50 através de Decreto Federal Nº 1.175 de 19 de agosto de 1950, instituindo a criação do Horto Florestal de Açu. Seu funcionamento teve início após aprovação das leis municipais de Açu de Nº 04/52 e 07/52 e do Decreto Lei que criou o ex – IBDF em 28 de fevereiro de 1967. Posteriormente foi regulamentado a doação da área de 215,25 hectares da FLONA, com anexação da Lagoa do Piató.
Compreende uma área de caatinga do tipo arbóreo arbustivo, sendo que sua vegetação vem sendo preservada desde 1950 (Ferreira, 1988). Os arbustos presentes são vegetais lenhosos que começam a esgalhar desde a sua base compondo os maciços florestais da zona seca. Alguns levantamentos preliminares foram feitos para execução do plano de manejo da FLONA/IBAMA, dentre eles o da mastofauna (Feitosa, 2004), com registros de 07 espécies e 03 gêneros de mamíferos não-voadores, representadas por espécies noturnas e diurnas, incluindo o sagüi (Martins, 2004), estando algumas espécies de mamíferos ameaçadas de extinção. Levantamento preliminar da herpertofauna da unidade registrou 19 espécies distribuídas em 15 famílias (Rodrigues, 2004). Quanto à composição da ornitofauna, foram catalogadas 59 espécies de aves (Oliveira, 2004), pertencentes a 15 ordens e 27 famílias.
não é inferior a 26°C. A precipitação pluviométrica média anual atinge 704 mm, sendo o clima do tipo BSwh´, segundo a classificação de Köppen (região mais quente com estação chuvosa no outono), ou seja, clima muito quente e semi-árido. O solo predominante é do tipo litólico, com cascalho de quartzo arredondado e areia
quartzosa(IBAMA, 2001).
A área utilizada no estudo está situada ao sul na FLONA (figura 02), próximo à entrada principal, onde há toda a infra-estrutura para as atividades administrativas da instituição, alojamentos, trilhas escolares para fins de educação ambiental e pesquisas científicas.
A reserva tem uma área de 518 hectares. Esta área possui espécies vegetais
como o angico, Anadenathera colubrina, aroeira, Myracrodruon urundeuva,
catingueira, Caesalpinia pyramidales, coqueiros Coccos nucífer, cumaru, Amburana
cearensis, feijão-brabo, Capparis flexuosa, goiabeira, Psidium guajava, juazeiro,
Zizyphus joazeiro, Juca, Caesalpinea ferrea, jurema branca, Mimosa verrucosa,
limão Cirus limon, mangueira, Mangifera indica, pau-d’arco, Tabebuia impetiginosa,
pereiro, Aspidosperma pyrifolium, pinha Anona sp, sabiá, Mimosa caesalpiniaefolia e
timbaúba, Enterolobium contortisiliquum.
A FLONA é considerada uma área muito importante para a conservação da flora e fauna da Caatinga, em especial os primatas, devido à sua extensão de ambientes naturais preservados.
3.2. Grupo de Estudo
O acompanhamento do grupo teve início em novembro de 2004. No entanto, os animais foram capturados e marcados em março de 2005, quando o grupo passou a ser chamado de grupo do Bosque (figura 03) e foi iniciada a coleta de dados. Era composto então por sete indivíduos, o casal reprodutor B2 e B3, um macho adulto B4 e a fêmea adulta B1, o macho infante B5 e o casal de filhotes, com data estimada de nascimento em fevereiro de 2005 (Tabela I). O nascimento da prole que se seguiu, ocorreu em julho de 2005, sendo de filhote único. Uma terceira prole, desta vez de gêmeos, nasceu em dezembro do mesmo ano. O grupo passou a ter, no final do período de estudo 10 indivíduos.
Tabela I – Composição do Grupo em janeiro de 2006.
COMPOSIÇÃO DO
GRUPO1 SEXO NASCIMENTO CLASSE DE IDADE2
B1 B2 B3 B4 B5 - - - - - F F M M M M F F M F ? ? ? ? ? 02/05 02/05 07/05 12/05 12/05 Adulto Adulto Adulto Adulto Adulto Subadulto Subadulto Juvenil Infante Infante
1B1 fêmea adulta subordinada; B2 fêmea adulta reprodutora; B3 macho adulto reprodutor;
Figura 03: Grupo Bosque: B1, fêmea subordinada forrageando, com a metade da cauda inferior pintada; B2, fêmea reprodutora alimentando-se com inseto, com os tufos tingidos; B3, macho reprodutor com colar de identificação (seta); B4, macho adulto subordinado com colar de identificação (seta); B5, macho juvenil (início do estudo) em repouso, com as patas posteriores e metade da cauda superior tingida.
B2 B1
B3 B4
3.3 - Captura e Marcação dos Animais
A primeira etapa do processo de captura consistiu na habituação dos animais à presença de uma armadilha em sua área de uso, para onde foram atraídos através de frutos da época, utilizados como iscas. As armadilhas continham cinco compartimentos individuais, permitindo a captura de animais separadamente. Capturados, os animais foram levados para o laboratório e, ainda na armadilha, anestesiados, exceto os infantes com até 2 meses de idade, utilizando-se o Ketalar – 50 mg/ml, em doses variando de 0,02 a 0,04 ml de acordo com a idade do animal.
Inicialmente, foi verificado se o animal possuía marcas naturais. Posteriormente, foi realizada na parte interna da coxa uma tatuagem, a seguir, a biometria, a avaliação das condições físicas, e nas fêmeas adultas, em particular, a sua condição reprodutiva.
Ao final do processamento, efetuou-se a marcação na pelagem do animal. Essa marca foi feita através de pinturas com ácido pícrico (que produz uma cor amarelada que contrasta com o pêlo predominantemente cinza da espécie) na pelagem; colar de três contas coloridas (figura 03), sendo a primeira vermelha para identificar os animais do grupo, e as demais de cores sortidas para distinção entre os indivíduos. Realizou-se também a tricotomia do pêlo da cauda dos animais. Essas marcas geralmente desaparecem após três ou quatro meses, já o colar pode ser visualizado com binóculo e raramente é perdido, garantindo a existência de uma marca externa mesmo após o desaparecimento das marcas no pêlo.
3.4. Observação Comportamental
3.4.1 - Técnicas utilizadas
minutos, com registro instantâneo a cada minuto. Uma vez concluído um focal, iniciava-se o seguinte, tendo como base a seqüência do sorteio. Cada animal era amostrado três vezes ao longo do dia. Além deste tipo de amostragem, foram feitos
registros ad libitum.
3.4.2 - Atividades comportamentais registradas
As categorias comportamentais consideradas neste trabalho foram:
1. Catação social: quando um animal realizava limpeza da pelagem de
indivíduos do grupo, utilizando as mãos ou a boca. A catação foi dividida em:
Receber: limpeza da pelagem do corpo de um indivíduo por outro animal;
Fazer: quando um animal realizava limpeza da pelagem de um outro
indivíduo.
2. Contato: quando um animal permanecia parado em contato físico com outro
(s) membro do grupo.
3. Proximidade: quando os animais estavam a uma distância inferior a 2m do
animal focal sem que houvesse contato físico.
4. Forrageio: procura de itens alimentares, no qual o animal se deslocava
lentamente utilizando as mãos para a manipulação de itens ou captura de insetos, bem como a perfuração das árvores para a extração do exsudado.
5. Ingestão de alimentos: quando o animal introduzia algum item alimentar na
boca e o ingeria, lambia ou mordia. Era anotado o tipo de alimento ingerido (fruto, exsudado ou inseto).
6. Locomoção ou deslocamento: quando o animal deslocava-se de um ponto
para outro a uma distância mínima de 2m; diferencia-se do forrageio porque nessa atividade os animais não param para examinar o ambiente em busca de alimento.
7. Descanso ou repouso: quando o animal estava imóvel, às vezes realizando
8. Outras Atividades: atividades que ocasionalmente ocorreram concomitantemente às atividades principais. Incluíram-se neste caso as categorias comportamentais sexuais (Cópula e tentativa de cópula) e a
agonísta (agressão), com registros ad libitum.
3.5 - Registro de Utilização da Área
Para obtenção dos dados referentes à área de uso do grupo, foram identificadas as árvores utilizadas pelos animais ao longo do dia, tendo como referência o percurso diário nas trilhas ali abertas em função do deslocamento dos animais na área (figura 04). O registro foi realizado com o auxílio de um “mapa” esboçado durante o período de observação dos animais.
Apesar de não ter sido realizado um estudo fenológico, era anotado o tipo de recurso utilizado dentro da área, fazendo um levantamento quando possível das espécies vegetais utilizadas como fonte de alimento (fruto e exsudado) e o local de dormida.
3.6 - Análise Estatística dos Dados
Para a análise dos dados das categorias comportamentais (repouso, forrageio, locomoção, ingestão e catação), bem como para o padrão de ingestão (fruto, exsudado, inseto) quanto as estações do ano (chuvosa e seca), utilizou-se o teste U de Mann-Whitney. Esse teste estatístico não-paramétrico compara se uma amostra de dois grupos independentes foi ou não extraída de uma mesma população. Podendo-se identificar assim se houve ou não diferenças estatísticas na freqüência observada nas estações analisadas (chuvosa e seca). A estação chuvosa compreendeu os meses de março a julho de 2005 e a estação seca, os meses de agosto de 2005 a janeiro de 2006.
que algum comportamento foi mais praticado em relação aos demais, considerando o grupo do animal analisado. Nos casos onde foi rejeitada a hipótese de igualdade, usou-se o teste de Wilcoxon para comparar os diversos animais dois a dois. A justificativa para utilização destes testes (Friedman e Wilcoxon) se dá pelo fato dos comportamentos serem relacionados, ou seja, a unidade observacional (sagüi) ser a mesma para cada comportamento estudado.
Figura 04 - Representação da área de uso de Callithrix jacchus, na FLONA-Açu. O Ponto 005
representa a Entrada Principal da FLONA. O Ponto 006 representa o limite direito inferior da área de uso. O Ponto 007 representa o final da trilha 1. O Ponto 008 representa o final da trilha 2. O Ponto
4. RESULTADOS
4.1 - Padrão de Atividade do Grupo
O padrão diário de atividades do grupo de Callithrix jacchus ao longo dos
meses de observação, apresentou variações (figura 05). O forrageio foi, de um modo geral, a atividade predominante com uma porcentagem média de 46,7% do tempo, variando os percentuais entre 31,6% (março) e 59,9% (outubro). Quanto à ingestão 14,1%, os meses com maior número de registros foram junho, julho e agosto, não tendo havido necessariamente coincidência com o perfil do forrageio ao longo do período de estudo.
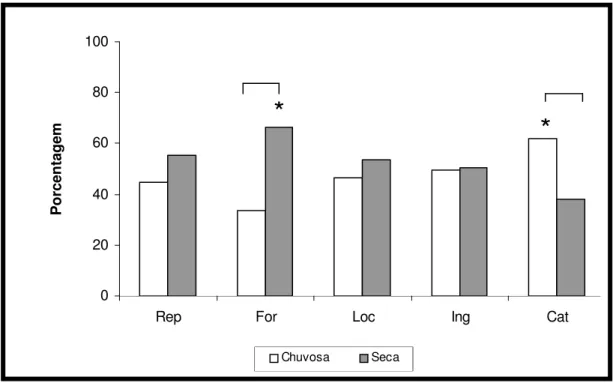
A segunda atividade mais freqüente foi o repouso, com um percentual médio em torno de 22,4%. O mês com maior expressão deste comportamento foi dezembro (46%), tendo havido em contrapartida uma redução nos níveis de locomoção, ingestão e catação (figura 05).
Na catação, foram registradas variações ao longo do estudo. O mês de maior índice correspondeu a maio com 28%. A partir de agosto, foi observada uma tendência à redução na expressão desse comportamento atingindo um menor percentual em janeiro. Mesmo assim, destacamos níveis elevados em novembro (20,1%), mês que antecedeu o nascimento de filhotes.
A atividade de locomoção foi menos freqüente em relação às demais, apresentando um percentual médio em torno de 3,9% ao longo de todo período de estudo.
Figura 05 – Padrão de atividade de Callithrix jacchus em ambiente natural (Bioma
Caatinga) no período de mar/2005 a jan/2006.
Figura 06 - Padrão de atividade de Callithrix jacchus em ambiente natural (Bioma
Caatinga), durante a estação chuvosa (mar-jul/2005) e seca (ago/2005-jan/2006). Rep: repouso; For: forrageio; Loc: locomoção; Ing: ingestão; Cat: catação. As linhas contínuas acima das barras indicam diferenças significativas na variável entre as estações chuvosa e seca. (*) indica níveis significativamente mais elevados.
0 20 40 60 80 100
Rep For Loc Ing Cat
P o rc e n ta g e m Chuvosa Seca
*
*
0% 20% 40% 60% 80% 100%mar abr mai jun jul ago set out nov dez jan
P o rc e n ta g e m
4.2 – Padrão de ingestão do grupo
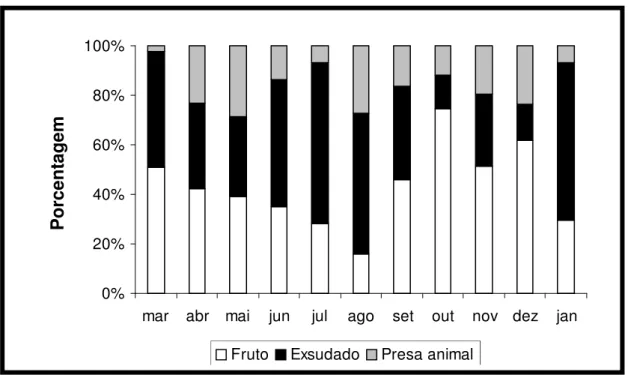
Durante o período de estudo (figura 07), o grupo ingeriu diferentes itens alimentares, os quais foram agrupados em três tipos: fruto, exsudado e inseto. O alimento mais ingerido por todo o grupo foi o exsudado (média percentual de 45,4%), seguido do fruto (39,2%). Os recursos de origem animal (15,4%) foram ingeridos em menor proporção.
A ingestão do exsudado apresentou dois picos, sendo o primeiro nos meses de junho (51,1%), julho (65,4%) e agosto (56,9%), coincidindo com níveis mais reduzidos na ingestão do fruto. O segundo pico ocorreu nos meses de janeiro (63,35%) e março (46,8%), quando foram observados os menores índices de consumo do inseto (figura 07).
O consumo de frutos a partir de março apresentou uma tendência a reduzir, atingindo o menor consumo em agosto (16%) (figura 07). A partir de setembro, houve um aumento no consumo desse item, tendo sido registrados os níveis mais elevados em outubro (74%) e dezembro (61%).
Nesse grupo, o recurso de origem animal mais consumido foram os insetos, os quais estiveram presentes nos onze meses de estudo, sendo seu consumo mais expressivo no mês de abril, maio, agosto e dezembro. Entretanto nos meses de março e abril foi observada a ingestão de ovos de pássaros e em dezembro a ingestão de lagartixa.
Figura 07 - Perfil alimentar de Callithrix jacchus em ambiente natural (Bioma Caatinga), no
período de mar/2005 a jan/2006.
Figura 08 - Perfil alimentar de Callithrix jacchus em ambiente natural (Bioma Caatinga),
durante a estação chuvosa (mar-jul/2005) e seca (ago/2005-jan/2006).
0% 20% 40% 60% 80% 100%
mar abr mai jun jul ago set out nov dez jan
P
o
rc
e
n
ta
g
e
m
Fruto Exsudado Presa animal
0 20 40 60 80 100
Fruto Exsudado Presa animal
P
o
rc
e
n
ta
g
e
m
O limão Cirus limon (Rutáceas) foi à fonte mais procurada pelo grupo, com
uma porcentagem de 56% (figura 09). Em seguida, tivemos a jurema branca Mimosa
verrucosa (Leguminosae-Mimosoideae) (16%). Com níveis ainda mais reduzidos de
procura, identificamos o angico Anadenathera colubrina
(Leguminosae-Mimosoideae) (8%), caju Anacardium occidentale (Anacardiaceae) (5%), cumaru
Amburana cearensis (Leguminosae-Papilionoideae) e a catingueira Caesalpinia
pyramidales, (Leguminosae) ambas com 4%, e outras espécies com uso ocasional
(7%).
Na análise ao longo dos meses de estudo (figura 10), verificamos que a jurema
branca Mimosa verrucosa (Leguminosae-Mimosoideae) foi ingerida em todos os
meses, porém o maior consumo foi registrado nos meses de maio (48,1%), outubro
(38,4%), e dezembro, quando atingiu o pico máximo (100%). O cumaru, Amburana
cearensis (Leguminosae-Papilionoideae) foi ingerido principalmente em junho (11%)
e novembro (15%). Os animais ingeriram o exsudado da catingueira Caesalpinia
pyramidales (Leguminosae) basicamente entre maio e setembro, sendo maio, o mês
de maior exploração, com 33,3%. O limãoCirus limon (Rutáceas) foi consumido em
quase todos os meses do estudo, em geral, com valores acima de 50%, exceto em junho, outubro, novembro e janeiro, quando outros itens foram mais explorados. O
consumo do angico Anadenathera colubrina (Leguminosae-Mimosoideae) esteve
concentrado apenas em junho (59,8%) e a exploração realizada em uma única árvore.
O exsudado do caju Anacardium occidentale (Anacardiaceae) foi consumido em
cinco meses, atingindo valores mínimos em janeiro (1,2%) e máximos em novembro (35%), sendo esse consumo concentrado em uma única árvore. Outras espécies vegetais nativas da caatinga foram consumidas em menor quantidade ao longo de todos os meses de estudo, as quais somadas variaram entre 1% e 31%.
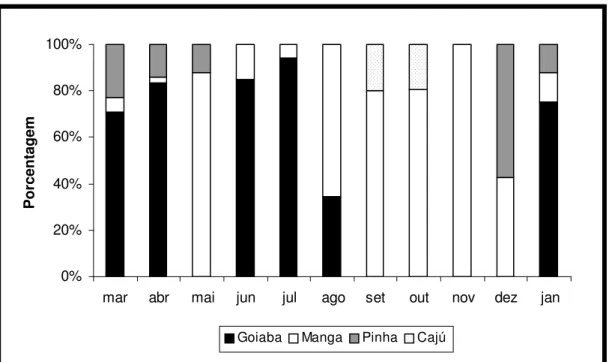
Quanto à ingestão de frutos, observou-se que a goiaba Psidium guajava
(Myrtaceae) (48%) e a manga Mangifera indica (Anacardiaceae) (38%) foram os
mais consumidos (figura 09). Com níveis ainda mais reduzidos de ingestão,
identificamos a pinha Anona sp. (Anonaceae) (9%) e o caju Anacardium occidentale
Ao longo dos meses (figura 11) verificou-se que a goiaba Psidium guajava,
quando presente, foi, de um modo geral, o fruto mais procurado pelos animais, exceto em agosto, quando começou a frutificação da manga. Por outro lado, a manga
Mangifera indica foi ingerida em todos os meses, porém em maior quantidade nos
meses entre agosto e dezembro, atingindo a ingestão máxima em novembro (100%).
O caju Anacardium occidentale foi ingerido nos meses de setembro e outubro em
proporções equivalentes. Quanto à pinha Anona sp, foi observado consumo máximo
em dezembro (57%).
Figura 09 – Árvores de exsudado e fruto utilizados na dieta de Callithrix jacchus em
ambiente natural (Bioma Caatinga), no período de mar/2005 a jan/2006.
Figura 10 - Espécies vegetais utilizadas para a gomivoria de Callithrix jacchus em
ambiente natural (Bioma Caatinga), no período de março/05 a janeiro/06.
0% 20% 40% 60% 80% 100%
mar abr mai jun jul ago s et out nov dez jan
P o rc e n ta g e m
Limão Jurema Catingueira Cumarú A ngic o Caju Outros
Exsudado 56% 16% 4% 4% 8% 5% 7%
Limão Jurema Catingueira Cumarú Angico Caju Outros
Fruto
48%
38% 9%
5%
Figura 11 - Árvores de frutos utilizados na dieta de Callithrix jacchus em ambiente natural
(Bioma Caatinga), no período de mar/2005 a jan/2006.
4.3 – Utilização da área
A área domiciliar apresentou diferenças quanto à composição florística e o tipo de recurso utilizado (figura 04).
A área de caatinga utilizada pelo grupo era caracterizada por árvores de
tamanhos bastante variados, entre elas, algumas de grande porte como a catingueira, timbaúba e cumaru. Essas foram utilizadas principalmente como árvores de dormir. Plantas arbustivas, geralmente com troncos finos e pouca folhagem quando ramificadas, foram utilizadas principalmente para a gomivoria. Herbáceas, as quais se ramificavam desde a base, foram utilizadas principalmente durante o forrageio e a procura de insetos.
Uma segunda área utilizada foi a de pomar, situada atrás das dependências físicas da FLONA, onde eram encontradas árvores frutíferas, exploradas diariamente pelo grupo, sendo o local onde os animais passavam a maior parte do seu tempo.
Devido a problemas quanto à marcação de trilhas, não foi possível fazer uma estimativa precisa dos locais que foram mais utilizados durante as estações chuvosa e seca.
0% 20% 40% 60% 80% 100%
mar abr mai jun jul ago set out nov dez jan
P
o
rc
e
n
ta
g
e
m
4.4 – Interação Social
4.4.1 – Catação
Os resultados mostraram que todos os indivíduos se envolveram nesta atividade, entretanto os maiores índices de catação ocorreram entre os reprodutores (B2-fêmea e B3-macho) (Tabela II). Estes animais foram considerados os reprodutores do grupo com base no registro de cópulas, que só foi registrada entre eles, em março de 2005 (pariu em julho) e em janeiro de 2006 (pariu em junho). O macho reprodutor foi o indivíduo que mais catou a fêmea reprodutora (Teste de
Wilcoxon: p= 0,0000) e por ela foi catado (Teste de Wilcoxon: p= 0,0000).
Considerando as interações macho-macho, o dominante (B3) interagiu significativamente mais com o adulto subordinado (B4), tanto fazendo quanto
recebendo catação (Teste de Wilcoxon: p= 0,0000 e p= 0,0001, respectivamente).
Quanto à catação entre a fêmea reprodutora (B2) e a não-reprodutora (B1) foi bem menos freqüente, não sendo observada diferença significativa na interação entre elas.
Tabela II: Matriz de interação do comportamento de catação feita e recebida pelos
animais adultos do grupo.
Recebe B1 B2 B3 B4 B5 TOTAL
Faz
B1 - 8 6 4 16 34 B2 10 - 206 6 5 227 B3 41 194 - 128 25 388 B4 1 21 95 - 10 127 B5 32 11 30 23 - 96
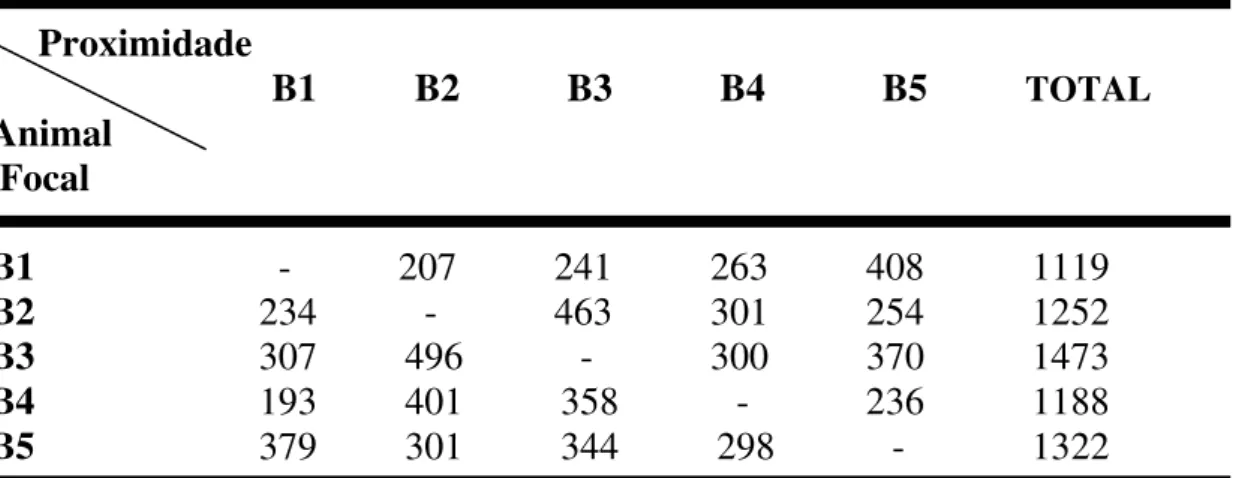
4.3.2 - Proximidade
Com base na tabela III, que apresenta a matriz de proximidade, verificamos que o casal reprodutor (B3-macho e B2-fêmea) teve mais vezes um ao outro como o
animal mais próximo (Teste de Wilcoxon: p= 0,0011).
Quanto aos animais subordinados a fêmea B1 ficou significativamente mais
próxima de B5 (Teste de Wilcoxon: p= 0,0008). O macho subordinado B4 esteve
sempre significativamente próximo da fêmea reprodutora (B2) (Teste de Wilcoxon:
p= 0,0033).
Tabela III: Matriz da interação do comportamento de proximidade (distância
máxima de 2 metros) entre os animais adultos do grupo.
Proximidade
B1 B2 B3 B4 B5 TOTAL Animal
Focal
B1 - 207 241 263 408 1119 B2 234 - 463 301 254 1252 B3 307 496 - 300 370 1473 B4 193 401 358 - 236 1188 B5 379 301 344 298 - 1322
5. DISCUSSÃO
Os resultados aqui encontrados nos permitem indicar que o sagüi C. jacchus
habitando a caatinga nordestina, desenvolve um padrão de atividades semelhante daqueles que habitam regiões de mata atlântica. O forrageio ocupa a maior parte do tempo destes, seguido pelo repouso, ingestão, catação e locomoção. A comparação com os dados citados na literatura mostra que as adaptações dos sagüis ao ambiente refletem-se claramente no ciclo de suas atividades diárias. Estas por sua vez, podem trazer contribuições para melhor compreensão da flexibilidade do comportamento desta espécie.
Verificamos que o forrageio efetuado pelos sagüis ocupou cerca de 47% do tempo de atividade dos animais independente da área, sendo intensificada significativamente no período seco. Assim como em nosso estudo, Cassimiro (2003)
e Cavalcanti (2002) verificaram que os grupos de C. jacchus em área de mata
atlântica investiram mais tempo no forrageio, com a ingestão dos alimentos conseguidos nessa procura. Alonso & Langguth (1989) e Digby & Barreto (1996) descrevem o mesmo padrão de atividade, tendo o forrageio e a alimentação como atividades principais dos animais adultos, em sítios diferentes de mata atlântica. O fato dos animais ocuparem a maior parte do seu tempo forrageando está relacionada com a obtenção dos recursos necessários para sua sobrevivência (Giraldeau & Caraco, 2000).
frouxas, permanecendo parados na maior parte do tempo, olhando em volta e saltando sobre as presas quando eram avistadas.
Esse padrão também foi observado por Cavalcanti (2002) ao estudar C.
jacchus em ambiente alterado de mata atlântica. Isso reflete mais uma vez a
flexibilidade destes animais, uma vez que os ambientes são muito diferentes, principalmente quanto à composição vegetal. Araújo (1996), estudando um grupo que vivia em plantação experimental, encontrou que os animais utilizaram freqüentemente, o extrato médio (2-10 metros de altura) no forrageio. Já o extrato inferior (0-2 metros) foi utilizado com menor freqüência. Em contraste, o grupo que vivia no fragmento de mata atlântica forrageou, preferencialmente, entre 0-6 metros de altura. Segundo o autor, a altura do forrageio depende das características do ambiente.
Porter (2001) estudando as técnicas utilizadas por Callimico goeldii no
forrageio de insetos, observou que, ao contrário de Saguinus fuscicollis, eles não
forrageiam nos buracos e cavidades das árvores, e utilizam as garras como técnica de captura, ao explorar diversos extratos arbóreos na procura de insetos. Bicca-Marques
(1999) e Peres (1993) ao estudar Saguinus fuscicollis, verificaram que a maneira de
forragear presas nesta espécie era manipulativa e ocorria em substratos altos e médios da floresta, com exploração de microhabitats de floresta. Esses primatas demonstraram uma procura visual das presas móveis expostas nas folhagens, com um
padrão semelhante ao observado para Callithrix jacchus quando forrageava nos
troncos das árvores, uma vez que nesta espécie a atividade manipulativa é exploratória (Varella & Yamamoto, 1991).
Nossos resultados demonstraram que o repouso foi a segunda atividade realizada pelos animais. Isso também é relatado com relação a dois (grupo PB e ES)
dos três grupos de C. jacchus estudados por Cassimiro (2003). Por outro lado, o
Segundo Alonso & Langguth (1989), o repouso é mais acentuado no período de meio-dia (12:00h às 14:00h), as horas mais quentes do dia, favorecendo os
comportamentos de socialização, como a catação. Em Ateles chamek selvagem, foi
observado padrão semelhante, com os animais permanecendo em repouso mais intensamente por volta das 12:00-14:00 horas. Esse padrão deveu-se principalmente às condições ambientais, ou seja, elevada temperatura e alta umidade (Wallace, 2001). Em nossa pesquisa, após este intervalo, registrava-se um novo aumento de forrageio, exploração de exsudados vegetais e frutos até o deslocamento para a árvore de dormir, quando se encerrava a atividade diária. Os locais utilizados para o descanso diurno estiveram sempre associados às áreas onde os animais previamente tinham comido. As árvores escolhidas para o descanso eram sempre árvores de frutos
(mangueira Mangifera indica) e de goma (sabiá Mimosa caesalpiniaefolia).
O padrão de repouso do Callithrix geoffroyi habitando a floresta Atlântica
pode estar relacionado ao grande consumo do exsudado, uma vez que os maiores índices de repouso compreenderam o período chuvoso, quando havia maior disponibilidade desse recurso. Outro fator que pode ter contribuído para esse padrão foi à concentração de árvores próximas umas das outras (Passamani, 1998).
Em nosso trabalho, a locomoção apresentou níveis mais reduzidos quando
comparada às demais atividades. Esteve relacionada principalmente com o forrageio e com a dispersão dos animais dentro da área domiciliar, a qual apresentava uma distribuição homogênea em sua vegetação. Analisando o padrão de atividade
realizada por grupos de C. jacchus em mata atlântica, Cassimiro (2003) observou que
o comportamento de locomoção mostrou índices variados para os diferentes grupos. Sugeriu que a expressão da atividade sofre influências particulares de cada área, tendo como base a relação entre seu tamanho e a forma como estão disponíveis sazonalmente os recursos dentro da mesma. Em estudo sobre a influência da presença de filhotes e do tamanho do grupo nas atividades diárias de um grupo silvestre de sagüis, Nascimento (1997) registrou que no período com filhotes dependentes, o deslocamento tendeu a ser menor.
Em Saguinus mystax e S. fuscicollis no Peru, Garber (1993) verificou que não
seca e chuvosa, assim como na nossa pesquisa. Também, Passamani (1998)
demonstra que em Callithrix geoffroyi, o tempo gasto na locomoção não apresentou
diferença significativa entre as estações. O principal pico dessa atividade seria no final da tarde, quando o grupo dirigia-se para sua árvore de dormir.
Os nossos resultados nos permitem destacar que os sagüis ingeriram frutos, exsudados e insetos diariamente, sendo fácil observar os itens ingeridos, exceto pequenos insetos na copa das árvores e nos arbustos próximos ao chão. Em estudo
realizado por Ferrari (1991) com Callithrix aurita, C. geoffroyi e C. flaviceps
habitando na floresta Atlântica, e por Rylands (1981) com C. humeralifer na
Amazônia, foi observado um aumento no consumo de goma na estação seca. Já na estação chuvosa ocorreu uma maior ingestão de frutos, mais abundantes nessa estação, sendo a goma incluída na dieta em menor quantidade. Essa variação teve uma estreita relação com o padrão de sazonalidade do ambiente.
Semelhante ao observado com as espécies de Callithrix citadas acima, Castro
et al (2000) mostraram que o consumo de goma em C. jacchus diminuiu em função
do aumento da disponibilidade de frutos, na estação chuvosa. Na estação seca, os frutos, exsudados e insetos foram consumidos igualmente. Esse mesmo padrão foi
verificado para C. jacchus na Floresta Atlântica costeira (Scanlon et al, 1991) e para
C.penicillata em áreas do cerrado no Distrito Federal (Vilela & Faria, 2002).
O padrão de ingestão do grupo em ambiente de caatinga não foi semelhante ao dessas espécies, ou seja, durante o período seco os animais ingeriram mais o fruto e o inseto. É importante destacarmos a presença de um bosque no limite da área de caatinga, onde o grupo passava grande parte do período de atividades, no qual eram encontradas árvores frutíferas exóticas, irrigadas diariamente, algumas delas com maior disponibilidade de frutos na estação seca e bastante exploradas pelos sagüis. Já no período chuvoso, a ingestão mais intensa foi do exsudado, provavelmente devido ao aumento da água presente no solo o que intensifica a troca de substâncias, conseqüentemente o fluxo de exsudado.
fonte de energia e de sais minerais para os organismos capazes de metabolizá-los
(Melo et al, 1997). Algumas espécies vegetais foram consumidas em meses bastante
específicos, como por exemplo, o Angico Anadenathera colubrina
(Leguminosae-Mimosoideae), na qual os animais exploraram seu exsudado somente em junho.
Em algumas espécies do gênero Callithrix, como C. penicillata, C. kuhli, C.
humeralifer, C. flaviceps, C. geoffroy, C. aurita, C. melanura e C. jacchus, foi
observada a exploração de goma em espécies vegetais das famílias Anacardiaceae, Mimosaceae, Meliaceae, Combrataceae, Leguminosae e Vochysiaceae (Stevenson & Rylands, 1988; Alonso & Langguth, 1989; Araújo, 1996). Cavalcanti (2002) ao
analisar o comportamento alimentar do C. jacchus em mata Atlântica secundária e
numa área de plantação, verificou que as duas únicas espécies vegetais incluídas na dieta dos animais para o consumo de exsudado pertencem à família Anacardiaceae
estando uma espécie situada na área de mata (Tapirira guianensis) utilizada pelos
três grupos e outra na área de plantação (Anacardium occidentale) utilizada por dois
grupos. Ainda nesse trabalho, a pesquisadora constatou que houve uma concentração no consumo do exsudado de um cajueiro localizado na parte central de sua área, no período em que os frutos estavam escassos. Alonso & Languth (1989), Araújo (1996)
e Castro et al (2000) relatam a ingestão de exsudados nas espécies vegetais
anacardiáceas citadas acima.
No estudo realizado por Garber (1984), os exsudados das espécies
Anacardium excelsum e Spondias mombim (Anacardiaceae) foram avaliados. A
análise química dos exsudados do Anacardium excelsum mostrou a presença de
diversos minerais, em particular o cálcio, com uma relação de cácio: fósforo de 31:1.
Saguinus fuscicollis e S. labiatus, em particular, consomem exsudados de poucas
espécies vegetais (Garber, 1993) particularmente durante a estação seca (Lopes & Ferrari, 1994).
No presente estudo, mesmo quando havia uma concentração de exemplares de uma mesma espécie vegetal numa determinada área, ocorria uma seletividade com relação às árvores utilizadas pelos animais para a gomivoria. Se os sagüis selecionam árvores exsudativas, é de se esperar que eles as escolham de acordo com a sua
C. aurita, em Monte Belo (Minas Gerais) explora o exsudado principalmente da
Acácia paniculata o que indica o uso do recurso alimentar que se encontra em
abundância (Martins & Setz, 2000). Miranda & Faria (2001) estudando os aspectos
ecológicos do C. penicillata no Cerradão do Brasil Central, analisou as espécies
arbóreas e arbustivas utilizadas como alimento. De acordo com os autores, o uso de exsudado de Vochysiaceae e Araliaceae na estação chuvosa pode ser devido à alta densidade de indivíduos destas espécies nas áreas de vida dos grupos e, conseqüentemente, a maior facilidade de acesso e uso das mesmas. A maior facilidade de acesso foi também observada para a espécie em estudo, pois apesar de
existirem apenas dois exemplares do limão Cirus limon na área de pomar era de fácil
acesso. A clara preferência pelo exsudado do limão Cirus limon (Rutáceas) e a
jurema branca Mimosa verrucosa (Mimosoideae) em relação aos exsudados das
demais árvores pode ser devido a uma maior palatabilidade, conteúdo nutricional ou pela facilidade de roer e exsudação.
Scanlon et al (1991) relata que os saguis da Floresta Atlântica costeira
utilizam árvores de todos os tamanhos. A porcentagem de árvores maiores com lesões produzidas pelos sagüis foi maior que a de árvores menores. As maiores também foram visitadas mais freqüentemente. Os nossos dados não evidenciaram o
mesmo padrão para C. jacchus na caatinga. Apesar de não ter conduzido um
levantamento da distribuição das árvores na área de estudo, bem como da medição do diâmetro, foi possível verificar que as duas espécies vegetais mais utilizadas para a gomivoria (limão e jurema branca) eram arbustos, apesar de na mesma área haver árvores de grande porte com troncos largos.
O consumo freqüente de frutos pelos animais como fonte de carboidratos não estruturais de diversas espécies pode ser explicado pela necessidade de uma dieta
nutricional balanceada (Aguiar et al, 2003). Apesar de ser um dos principais itens na
dieta de Callithrix, o consumo de frutos não variou significativamente entre as
da frutificação em algumas espécies coincidiu com o início desse evento biológico em outras. Isso permitiu o grupo consumir frutos em ambas estações.
Com relação ao efeito da sazonalidade na produção de frutos e sobre o consumo realizado pelos animais Stevenson & Rylands (1988) verificaram que
Callithrix humeralifer apresenta um padrão marcadamente sazonal. De forma
semelhante, Ferrari & Lopes-Ferrari (1989) evidenciaram que a sazonalidade de recursos ocorre de modo bastante marcante. A sazonalidade na produção de frutos é maior em áreas de mata secundária ou que sofreram ação antrópica, onde normalmente a diversidade de espécies vegetais é menor que em áreas de mata primária.
As frutas são componentes importantes na dieta de Saguinus fuscicollis, S.
labiatus e Callimico goeldii (Porter, 2001). Este autor discute que durante o início da
estação seca, ambos primatas consomem frutas de espécies vegetais variadas. Peres
(1993) relata que Saguinus fuscicollis, S. mystax dedicaram a maior parte do tempo
de alimentação ao fruto maduro, sendo esses sempre maduros, pequenos, amarelos ou vermelhos e espacialmente agrupados.
No nosso estudo, o grupo apresentou um elevado índice no consumo de frutos
da goiaba Psidium guajava. Este fato deve-se provavelmente a alta disponibilidade