LAINE CRISTINA FERNANDES
MORFOLOGIA E EFEITOS DO ATAQUE DE
Metaxyonycha angusta (PERTY) (COL.: CHRYSOMELIDAE)
SOBRE CRESCIMENTO E PRODUÇÃO DE EUCALIPTO
Tese apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Entomologia, para obtenção do título de
Doctor Scientiae.
VIÇOSA
Ficha catalográfica preparada pela Seção de Catalogação e Classificação da Biblioteca Central da UFV
T
Fernandes, Laine Cristina , 1980-
F363m Morfologia e efeitos do ataque de Metaxyonycha angusta (Perty) (Col.: Chrysomelidae) sobre crescimento e produção de eucalipto/ Laine Cristina Fernandes. – Viçosa, MG, 2009.
xi, 86f. : il (algumas coloridas); 29cm.
Inclui anexos.
Orientador: Norivaldo dos Anjos Silva.
Tese (doutorado) - Universidade Federal de Viçosa.
1. Inseto florestal. 2.Besouro. 3. Eucalipto – Crescimento.I. Universidade Federal de Viçosa. II.Título.
LAINE CRISTINA FERNANDES
MORFOLOGIA E EFEITOS DO ATAQUE DE
Metaxyonycha angusta (PERTY) (COL.: CHRYSOMELIDAE)
SOBRE O CRESCIMENTO E PRODUÇÃO DE EUCALIPTO
APROVADA: 31 de julho de 2009.
_________________________________ _________________________________
Prof. Hélio Garcia Leite Prof. Acácio Geraldo de Carvalho (Co-Orientador) (Co-Orientador)
_________________________________ _________________________________
Prof. Rodrigo Diniz Silveira Dra. Dalva Luiz Queiroz
_________________________________ Prof. Norivaldo dos Anjos Silva
(Orientador)
ii
A DEUS que nos deu a vida e a ciência do espírito.
Agradeço
Aos meus pais José Reis Fernandes e Vilma de Oliveira Fernandes Com todo meu amor e carinho,
Dedico
Ao meu esposo Glêison pelo amor, carinho, compreensão e constante incentivo,
iii
AGRADECIMENTOS
Agradeço a Deus, em primeiro lugar, por Seu amor infinito e pela sabedoria, que me permitiram alcançar esta vitória.
Ao meu esposo Glêison, pelo estímulo, pela dedicação, pelo carinho, amizade, pela confiança em minhas atitudes e ações, e por todos os momentos agradáveis ao seu lado, e pela força em diversos momentos deste trabalho.
Ao meu filho Glêison Augusto, meu “companheirinho”, ainda tão pequeno, mas a alegria do meu viver...
Minha família maior (pais, irmãos, sobrinho, tios, primos, sogra, sogro e cunhados) pelo amor, os quais mesmo estando longe, sempre estão no meu coração e na minha lembrança.
À Universidade Federal de Viçosa, através da Coordenação de Pós-Graduação em Entomologia e aos professores, por todos os ensinamentos e pela oportunidade de desenvolvimento dos trabalhos.
À Fundação de Amparo a Pesquisa de Minas Gerais (FAPEMIG), que forneceu a bolsa de mestrado durante todo o período de realização desse curso.
iv
Aos co-orientadores Hélio Garcia Leite e Geraldo Acácio de Carvalho pela dedicação e sugestões. Especialmente ao Hélio pela ajuda e ensinamentos para a análise dos dados.
Aos membros da banca, professor Rodrigo Diniz Silveira e Dalva Luiz de Queiroz, pelas sugestões para melhoria deste trabalho. Sobretudo a Dalva pela ajuda na elaboração dos desenhos esquemáticos.
Aos suplentes João Alfredo Marinho Ferreira e Carolina Rocha da Silva, por terem se colocado a disposição para fazer parte da banca.
Ao Dr. Roger Wills Flowers da Florida Agricultural and Mechanical University, USA por ter identificado o inseto estudado.
A todos os professores que, com serenidade, contribuíram para a minha formação neste curso.
À AGROTORA Reflorestamento e Pecuária, pela oportunidade de desenvolver as pesquisas em parceria, pelo apoio pessoal e material, especialmente aos engenheiros André Rocha, Nardoto e aos funcionários Custódio, João Anselmo e à secretária Graça.
Aos colegas da Casa dos Cupins (CDC), Gláucia, Carolina, Janaína, Dinarte, Carlos, Rodolfo, Pedro, Alexandre e Wagner, pela agradável convivência.
Aos funcionários da Casa dos Cupins e do Insetário, Sr. Antonio Cypriano, Antônio Fundão, Camilo, Sr. Manuel e Sr. Zé Cláudio, pelo apoio e colaboração para a realização deste trabalho.
A todos que me ajudaram na coleta de dados no campo, Gláucia, Alesandre, Janaína, prof. Norivaldo e João Anselmo.
A todos os professores do curso de Entomologia, pelos ensinamentos. Aos colegas do programa de Pós-graduação em Entomologia, pelo pela agradável convivência e troca de experiências.
Aos funcionários da pós-graduação em entomologia, em especial à Mª Paula e Míriam (Secretárias do Curso) pela competência, dedicação ao trabalho e amabilidade no trato com os estudantes.
A Dra. Lisiane Dilli Wendt pela ajuda na comparação direta com os exemplares depositados na “Coleção de Entomologia Pe. J. S. Moure” (DZUP),
v
A todos os companheiros do curso em Entomologia pela oportunidade de trabalho conjunto e troca de experiências durante as pesquisas realizadas.
Às amigas da “diretoria”, Glaucita, Camileixon e Carolleixon, por todo carinho, ajuda e, principalmente, por sempre dividirem minhas alegrias e tristezas.
Finalmente, meus sinceros agradecimentos a todas as pessoas e instituições que direta ou indiretamente, colaboraram na execução deste trabalho, em especial aos colegas de trabalho.
vi
BIOGRAFIA
LAINE CRISTINA FERNANDES, filha de José dos Reis Fernandes e Vilma de Oliveira Fernandes, nasceu em 14 de Janeiro de 1980 em Caeté, MG. Concluiu o ensino fundamental ao médio no Colégio Prof. Servelino Ribeiro (1991 a 1997), em Bocaiúva, MG.
Em março de 1998 ingressou no curso de Ciência
Biológicas/Bacharelado na Universidade Estadual de Montes Claros-MG, diplomando-se em dezembro de 2001.
Em março de 2002, matriculou-se como estudante especial no programa de pós-graduação em Entomologia na Universidade Federal de Viçosa-MG. Em agosto, desse mesmo ano, ingressou no Curso de Mestrado em Entomologia na área de Concentração: Manejo de Pragas Florestais, na mesma instituição, submetendo-se à defesa de tese em julho de 2004.
vii
ÍNDICE
RESUMO ... IX ABSTRACT ... XI
INTRODUÇÃO GERAL ... 1
REFERÊNCIAS BIBLIOGRÁFICAS ... 3
CAPÍTULO 1: MORFOLOGIA DO ADULTO DE METAXYONYCHA ANGUSTA (PERTY) (COL.: CHRYSOMELIDAE) ... 5
RESUMO ... 5
ABSTRACT ... 6
1. INTRODUÇÃO ... 7
2. MATERIAL E MÉTODOS ... 9
3.RESULTADOS E DISCUSSÃO ... 11
3.1MORFOLOGIADOADULTO ... 11
3.1.1CABEÇA ... 11
3.1.2TÓRAX... 13
3.1.3ABDOME ... 15
4. CONCLUSÃO ... 17
5. FIGURAS ... 18
6. REFERÊNCIAS BIBLIOGRÁFICAS... 22
CAPÍTULO 2: EFEITOS DO ATAQUE DE METAXYONYCHA ANGUSTA (PERTY) (COLEOPTERA: CHRYSOMELIDAE) SOBRE O CRESCIMENTO E PRODUÇÃO DE CLONES DE EUCALIPTO ... 25
RESUMO ... 25
viii
2. REVISÃO DE LITERATURA ... 30
3. MATERIAL E MÉTODOS ... 34
4. RESULTADOS E DISCUSSÃO ... 37
5. CONCLUSÃO ... 57
6. REFERÊNCIAS BIBLIOGRÁFICAS... 58
ix
RESUMO
FERNANDES, Laine Cristina, D.Sc., Universidade Federal de Viçosa, julho de 2009. Morfologia e efeitos do ataque de Metaxyonycha angusta (Perty) (Col.: Chrysomelidae) sobre o crescimento e produção de eucalipto.
Orientador: Norivaldo dos Anjos Silva. Co-orientadores: Hélio Garcia Leite e Acácio Geraldo de Carvalho.
Este trabalho teve como objetivo o de ampliar as informações sobre a morfologia de adultos em Metaxyonycha angusta, e avaliar efeitos decorrentes
do seu ataque sobre o crescimento das árvores em clones de eucalipto. Os estudos foram realizados num plantio clonal de eucaliptos no município de Andrelândia, Minas Gerais, nos laboratórios de Entomologia da Universidade Federal de Viçosa e na Empresa Brasileira de Pesquisa Agropecuária/Centro Nacional de Pesquisas de Florestas (Embrapa-Florestas). No campo, árvores de 12 clones de Eucalyptus spp. foram avaliadas quanto a intensidade de
injúrias na copa. O delineamento experimental foi o de blocos casualizados com três repetições e 25 árvores por parcela, avaliando-se o crescimento em altura e diâmetro dos 13 aos 70 meses de idade da cultura. Como resultado obteve-se que M. angusta tem o tegumento recoberto por punctuações; o élitro
x
em outras as respostas no crescimento apresentaram como alterações compensatórias ou até mesmo iguais às das árvores não atacadas ou pouco-atacadas. Foram encontradas reduções de até 40,2 % na produção final de madeira, aos 70 meses após plantio, o que equivale a, aproximadamente, 59 m3 de madeira por hectare. Com base no valor de mercado desta madeira
xi
ABSTRACT
FERNANDES, Laine Cristina, D.Sc., Universidade Federal de Viçosa, july, 2009. Morphology and damage effects of Metaxyonycha angusta (Perty) in eucalypt plantations. Adviser: Norivaldo dos Anjos Silva. Co-advisers: Hélio Garcia Leite and Acácio Geraldo de Carvalho.
This study deals to the morphology of the “four-spotted beetle”, Metaxyonycha angusta, and to evaluate the effect of tree damage by beetle cloned eucalypt
tree. Lab works were developed at the Federal university of Viçosa and Embrapa-Forests . Activities were carried out in a eucalypt plantation of cloned in the Andrelândia country, Minas Gerais State, Brazil. In the last case, A randomized experimental 3-blocks desing was used; each experimental unit had 25 trees randomly marked. The diameter and height growth were evaluated up to the 70 th month after trees were planted. As result, we found adult M.
angusta has the elytra convex, yellowish, and with four blue-greenish spots.
The head is wider than long. Pronotum is convex and more long than wide with scattered punctuation. Abdome with five visible segments both the sexs. The legs are free and develop without modifications in the three parts of the legs. In some variable the growth was presented with harmful alteration while in others the alterations had been compensatory or exactly equal to the one of the not damage trees. Wood losses were then evaluated up to the different defoliation levels caused by leaf-beetles in the first year, and, the highest value was 40.2%, compared to the control production 59.47 m3 /ha of wood was not
produced due to the most severe defoliation level. Since this lost wood may be more valuable than the IPM techniques usable against leaf beetles, we consider the M. angusta as a new and additional pest to the Brazilian eucalypt
1
INTRODUÇÃO GERAL
No Brasil, o segmento de culturas florestais e seus produtos têm uma crescente participação na economia devido ao aumento e ao melhoramento dessas plantações. A madeira produzida está entre as de mais baixo custo de produção e de melhores qualidades entre as encontradas no mundo (FOELKEL, 2007). Isto se deve a técnicas como a clonagem que permiti a produção uniforme e homogênea da cultura florestal (ZOBEL, 1982 e BORÉM, 2007).
O setor florestal produtivo brasileiro é constituído primordialmente por plantios de espécies do gênero Eucalyptus que são cultivados em varias
regiões devido a sua capacidade de adaptação a vários tipos de habitats (GOMIDE & COLODETTE, 2007). Entretanto, por serem implantados em maciços homogêneos, favorecem o surgimento de insetos que podem tornar-se pragas dessas culturas, o que é propiciado pelo aumento da oferta de alimento e redução na diversidade e abundância de inimigos naturais (ALMEIDA et al.,
1987; PANIZZI & PARRA, 1991), provocado pelo desequilíbrio ecológico que passa a existir nesses ambientes.
2
(OGLOBIN, 1935; MARICONI, 1956; OHMART & EDWARDS, 1991; ANJOS & MAJER, 2003).
O ataque de insetos desfolhadores prejudica o desenvolvimento das árvores, causando danos como à redução na produção de madeira, devido à mudança nas tendências de crescimento até mesmo podendo ocasionar a morte de árvores (CANDY, 1992; ABBOTT et al., 1993, MENDES 1999).
A importância dos besouros da família Chrysomelidae na silvicultura brasileira vem crescendo nos últimos anos em função da ocorrência de grandes surtos em culturas florestais (OHMART & EDWARDS, 1991; MENDES, 1999; ANJOS & MAJER, 2003). Entre estes besouros de interesse florestal no Brasil, destaca-se a espécie Metaxyonycha angusta (PERTY,
3
REFERÊNCIAS BIBLIOGRÁFICAS
ABBOTT, I.; HEURCK, P. V.; BURBIDGE, T. Impact of frequency and intensity of defoliation on growth of Jarrah (Eucalyptus marginata) an experimental study
with saplings. Forest Ecology and Management, v. 56, p. 170-183, 1993. ALMEIDA, A. F.; LARANJEIRO, A. J.; LEITE, J. E. O melhoramento ambiental no manejo integrado de pragas: um exemplo na Aracruz Florestal. Silvicultura, v.39, p.21-25, 1987.
ANJOS, N.; MAJER, J.D. Leaf-eating beetles in Brazilian eucalypt plantations.
School of Enviromental Biology Bulletin, Australia, v.1, n. 23, p. 8-9. 2003. BORÉM , A. Biotecnologia Florestal. Viçosa: Editora UFV, 387p. 2007.
CANDY, S.G.; ELLIOTT, H.T.; BASHFORD, R.J.; GREENER, A. Modelling the impact of defoliation by the leaf beetle, Chrysophtharta bimaculata (Coleoptera:
Chrysomelidae), on height growth of Eucalyptus regnans. Forest Ecology Management, v.54, p.69-87, 1992.
FERNANDES, L. C.; ANJOS, N. Ocorrência do besouro desfolhador
Metaxyonycha angusta (Perty,1832) em reflorestamentos de eucaliptos, em
Minas Gerais. In: Congresso Brasileiro de Entomologia, 20.,2004, Gramado.
Anais...Gramado: Sociedade Entomológica do Brasil, 2004
FERNANDES, L. C. Biologia de Metaxyonycha angusta (Perty) (Coleoptera: Chrysomelidae) e efeito do seu ataque em eucaliptos, num sistema agroflorestal. 2004. 73f. Dissertação (Mestrado em Entomologia) –
Universidade Federal de Viçosa, 2004.
4
GOMIDE, J.L., COLODETTE, J.L. Qualidade da Madeira. In: BORÉM, A. (Ed.). Biotecnologia Florestal. Viçosa: Viçosa: Editora UFV, 387p. 2007.
MARICONI, F. A. M. Alguns besouros depredadores de eucaliptos na região de Piracicaba. O Biológico, v.22, n.11, p.1-141, 1956.
MENDES, J. E. P. Nível de dano e impacto do desfolhamento por Costalimaita ferruginea (Fabr.) (Coleoptera: Chrysomelidae) em Eucalyptus grandis Hill ex Maiden. 1999. 99f. Dissertação (Mestrado em
Entomologia)- Universidade Federal de Viçosa., Viçosa, 1999.
OGLOBIN, A. A. Nueva plaga de los eucaliptos (Colaspoides vulgata Léf.) em
Missiones. Almanaque del Ministerio de Agricultura, v.10, p.255-256, 1935. OHMART, C. P.; EDWARDS, P. B. Insect herbivory on eucalyptus. Annual Review of Entomology, v. 36, p. 637-657, 1991.
PANIZZI, A. R.; PARRA, J. R. P. A ecologia nutricional e o manejo integrado de pragas. In____. PANIZZI, A. R., PARRA, J. R. P. Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo: Manole, 1991. p.313-329.
ZOBEL, B.J. Clonal forestry in the eucalypts. In: Ahuja, M.R. (Eds). Clonal
5
CAPÍTULO 1: Morfologia do adulto de Metaxyonycha angusta (Perty) (Col.: Chrysomelidae)
RESUMO
O objetivo deste trabalho foi o de caracterizar morfologicamente adultos de
Metaxyonycha angusta permitindo uma fácil identificação no campo. Os
adultos foram coletados no município de Andrêlandia, MG, em clones de
Eucalyptus spp. em dezembro de 2005. Os desenhos foram realizados com o
auxilio de estereomicroscópio e de microscópio, ambos acoplados à câmara clara. O adulto apresenta tegumento recoberto por punctuações; élitros convexos, amarelos com quatro manchas azul-esverdeadas, metálica, brilhante. Cabeça dorsalmente mais larga que longa não ultrapassando a maior largura do pronoto. Pronoto convexo, mais comprido que largo, com punctuações esparsas. Escutelo triangular. Pernas livres e desenvolvidas. Fêmur, tíbia e tarsômeros sem modificações nos três pares de pernas. Fêmur e tíbia subiguais no comprimento. Abdome ventralmente amarelo-vermelho, com cinco segmentos visíveis em ambos os sexos. É a primeira vez que a espécie
M. angusta está sendo redescrita de maneira detalhada gerando informações
6
ABSTRACT
The objective of this study was to morphologically characterize adult
Metaxyonycha angusta, allowing an easy identification in the field. Adult beetles
were collected in Andrelândia, a municipality of the State of Minas Gerais, Brazil, from Eucalyptus spp. clones in December 2005. The drawings were
performed with the aid of a microscope and a stereomicroscope, both coupled to a camera lucida. Metaxyonycha angusta presents the following
characteristics: integument covered with punctulae; elytra convex, yellow with four greenish blue spots, metallic, shiny; head dorsally wider than long, but never more than the largest width of the pronotum; pronotum convex, longer than wide, with sparse punctulae; triangular scutellum; legs free and developed; femora, tibiae and tarsomeres with no modifications in the three leg pairs; ventral abdomen yellow-red with five visible segments in both sexes; fifth uroesternite with a slight cut in the posterior margin in males and with a deeper cut in females. This is the first time M. angusta is redescribed with more details,
7
1. INTRODUÇÃO
O gênero Metaxyonycha CHEVROLAT (1847) inclui mais de 50 espécies
de eumolpineos neotropicais, sendo a maioria das espécies compostas de besouros grandes e atrativos (FLOWERS, 1996). A descrição da morfologia externa de Metaxyonycha angusta (PERTY, 1832) foi realizada primeiramente
por PERTY (1832), que a denominou cientificamente como sendo Galleruca
angusta. CHEVROLAT (1847) descreveu uma nova espécie que foi
denominada como sendo Metaxyonycha bouvouloiri. Cerca de 40 anos depois
LEFÈVRE (1875) concluiu que M. bouvouloiri era o mesmo besouro descrito
por PERTY (1832). BECHYNÉ (1953) transferiu esta espécie para o gênero
Colaspis passando a ser denominada de Colaspis angusta. Entretanto,
segundo BROWN (1961), BECHYNÉ (1953) sinonimizou erradamente o gênero
Metaxyonycha com Colaspis. Assim sendo, todos os insetos do gênero Colaspis relatados por BECHYNÉ (1953) são, na verdade, pertencentes ao
gênero Metaxyonycha. Portanto, a espécie em questão deve ser mencionada
como sendo Metaxyonycha angusta (PERTY, 1832).
Esta é uma espécie conhecida popularmente no Brasil como “B esouro-de-quatro-pintas”, cujos adultos se alimentam das folhas de eucaliptos, ocasionando perdas na produção de madeira. Após sua descrição inicial informações sobre a morfologia externa de M. angusta só foram acrescentadas
8
praga dos eucaliptais e por FERNANDES et al. (2005) que observaram
dimorfismo sexual nesta espécie. Os trabalhos existentes sobre esta espécie no que diz respeito à morfologia são escassos, existindo, ainda, a possibilidade de este inseto vir a se tornar praga freqüente na eucaliptocultura brasileira.
Assim sendo, é necessário e importante caracterizar morfologicamente os adultos de Metaxyonycha angusta (Perty, 1832), possibilitando a correta
9
2. MATERIAL E MÉTODOS
Adultos de M. angusta foram coletados em dezembro de 2005 numa
cultura clonal de eucaliptos, no município de Andrelândia (21º 44’ 20” S e 44º 18’ 54” W),na mesoregião Sul/Sudoeste, Estado de Minas Gerais. OS estudos
foram realizados nos Laboratórios de Entomologia da Universidade Federal de Viçosa e na Empresa Brasileira de Pesquisa Agropecuária/Centro Nacional de Pesquisas de Florestas (Embrapa-Florestas). A identificação da espécie foi realizada pelo Dr. R. Wills. Flowers1 e os exemplares foram depositados no
Museu de Entomologia da Universidade Federal de Viçosa (UFVb), em Viçosa, MG, Brasil.
Antes das observações dos caracteres externos, os adultos foram dissecados e suas partes preparadas em lâminas, conforme os procedimentos recomendados por SILVA et al. (2005). Primeiro, os exemplares foram
previamente aquecidos em água com algumas gotas de detergente por cerca de dois minutos para a remoção de gorduras; em seguida, foram novamente aquecidos em solução de KOH a 10%, por cerca de cinco minutos, e lavados em água destilada. Com o auxílio de pinças e estiletes, foram dissecações em placas de Petri com água. Para a confecção das ilustrações as diferentes partes do exoesqueleto foram colocadas em placas de Petri com água. As
1
10
peças bucais e asas foram fixadas entre lâminas com glicerina e lamínulas. A genitália do macho foi retirada do adulto, introduzindo uma pinça entre o pigídio e o quinto uroesternito visível. A da fêmea foi retirada através de um corte nos pleuritos do abdômen. Em seguida, as estruturas da genitália foram aquecidas em solução de KOH a 10%, por cerca de dois minutos, sendo lavadas em água destilada, posteriormente. Estudos de morfologia externa e da genitália foram realizados com o auxílio de estereomicroscópio Zeis Stemi SV6; para o desenho das demais estruturas utilizou-se o microscópio Standart 20, ambos acoplados a câmara clara, conforme sugerido por LIMA (1992), o qual considerou que, desta maneira, os caracteres podem ser mais facilmente observados e registrados em estudos comparativos. Para a confecção dos desenhos, o material foi fixado entre lâmina e lamínula. Os desenhos foram escaneados e editados com o programa Corel Photo-Paint 11.
11
3. RESULTADOS E DISCUSSÃO
3.1 MORFOLOGIA DO ADULTO
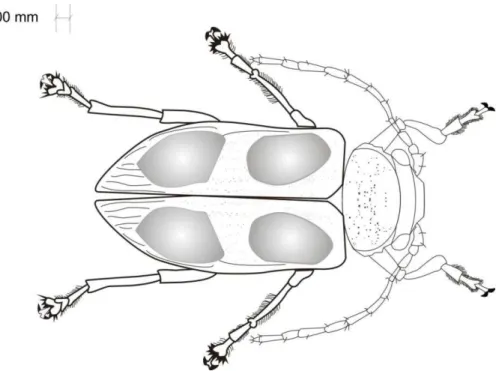
O tegumento é coberto de punctuações; os élitros são amarelos e sobre estes existem quatro manchas da tonalidade azul-esverdeada, metálica e brilhante (4.6 BG conforme MUNSELL, 1952) (Figura 1e 2). PERTY (1832) e LEFÈVRE (1875) atribuíram manchas azuis, verdes ou violáceos brilhantes, nos élitros. Cabeça prognata, pronoto, pró, meso e metasterno, pernas e esternitos abdominais visíveis, com coloração amarelada e brilhante. O abdome, em vista ventral, tem coloração amarelo-vermelha, com cerdas bem finas, em sentido longitudinal no centro e nas margens de cada segmento abdominal.
3.1.1 CABEÇA
12
dos olhos, em M. angusta é a mesma relatada por CABRERA et al. (2005) para
Maecolaspis monrosi (Bechyné). Olhos emarginados pelas antenas também
foram constatados no gênero Cryocolaspis (FLOWERS, 2004a), Apterodina
bucki (Bechyné) e Brachypterodina morae Flowers (FLOWERS, 2004c) e Prionodera adiastola Flowers, Prionodera arimanes Flowers, Prionodera esmeralda Flowers e Prionodera furcada Flowers (FLOWERS, 2004d), todas
estas espécies pertencentes a sub-família Eumolpinae.
Segundo GUÉRIN (1953), no gênero Metaxyonycha as antenas
apresentam-se em formato filiforme sendo um pouco mais comprida do que a metade do corpo, como observado nos exemplares estudados. As antenas são inseridas dorsalmente; com 11 artículos ultrapassam o comprimento do pronoto; todos os artículos com microcerdas (Figura 3). PERTY (1832) e LEFÈVRE (1875) atribuíram a M. angusta a presença de fina pilosidade e de
coloração escura nas antenas como também observado neste trabalho. O diâmetro basal dos artículos diminui em direção ao ápice da antena, sendo o último artículo de formato cônico e curto na extremidade. O pedicelo globoso e de comprimento igual à metade do comprimento do escapo. Escapo subcilíndrico com o maior diâmetro da antena. Escapo e pedicelo, amarelos com pubescência esparsa, aumenta gradativamente em quantidade nos artículo.
Numerosos órgãos sensoriais ou sensilas em forma de pêlos são encontrados nas antenas dos insetos e funcionam como quimiorreceptores mecanorreceptores ou termorreceptores (GULLAN & CRANSTON, 2004) e a morfologia dessas estruturas podem fornecer informações sobre o comportamento (PLOOMI et al., 2003). Pêlos encontrados nos antenômeros
de M. angusta podem ser estruturas que atuam no comportamento destes
besouros, pois conforme FERNANDES (2004), quando se sentem ameaçados os besouros voam pousando nos galhos mais baixos, no solo ou em árvores próximas.
Modificações na forma da antena podem permitir a diferenciação entre machos e fêmeas como observado por BARTLET et al. (1999) em adultos de Psylliodes chrysocephala (Linnaeus). Entretanto, em M. angusta, nenhuma
13
Mandíbulas triangulares em vista dorsal, distintamente curvas, fortemente quitinizadas; aproximadamente duas vezes mais longas do que a largura da base; margem interna com pilosidade esparsa e curta nas laterais; ápice projetado, com uma cerda curta (Figura 4).
Labro sub-retangular, mais largo que comprido mais estreito que a largura do clípeo, pouco visível dorsalmente, com punctuações esparsas e cerdas curtas (Figura 5).
Maxilas com gálea e lacínia distintas, externamente providas de cerdas. Os palpos maxilares são constituídos de quatro segmentos em formato cônico com último artículo afilado, com ápice cortado obliquamente, com sensilas (Figura 6). A presença dos quatro artículos do palpo maxilar que segundo MATSUDA (1965) é típico de formas menos especializadas da ordem Coleoptera. Lábio estreito e alongado, mento com cerdas longas, palpos labiais pequenos, com três artículos, o apical truncado (Figura 5).
3.1.2 TÓRAX
Tórax rígido. Pronoto dorsalmente mais comprido que largo; convexo; possui punctuações finas, esparsas. Borda lisa, arqueada para fora, formando
uma linha basal, visível dorsalmente; margem posterior mais estreita que os élitros; superfície coberta por punctuações com microcerdas.
Prosterno alargado no ápice; maior largura subigual ao diâmetro da cavidade da coxa e com a margem posterior sub-reta. Mesosterno tão largo quanto a cavidade da mesocoxa, com poucas e delicadas cerdas. Metasterno aplainadas com punctuações rasas e esparsas.
Escutelo triangular, conforme descrição por LEFÈVRE (1875), em forma de “V”; base subigual no comprimento, superfície lisa, poucas punctuações, cor amarela (Figura 1 e 2). A coloração corrobora as observações de PERTY (1832).
A forma do pronoto neste estudo, bem como as punctuações parecem ser características comuns entre os insetos da subfamília Eumolpinae, conforme os registros de FLOWERS (2004d) ao descreverem o gênero
Prionodera e de CABRERA et al. (2000) ao descrever Maecolaspis monrosi
14
descrito por FLOWERS (2004a) em Cryocolaspis crinita Flowers e por
FLOWERS (2003) ao descrever Thysanomeros ulateae Flowers. Contudo,
estes caracteres parecem ser semelhantes a vários grupos e por isto não devem ser utilizadas para identificação de M. angusta.
Pernas livres, desenvolvidas, coloração amarela; tarsos negros (Figura 7, 8 e 9). Fêmur, tíbia e tarsômeros sem modificações nos três pares de pernas. Trocânteres expostos, subtriangulares, margens arredondadas, cerdas curtas na margem inferior. Fêmur e tíbia subiguais no comprimento. Fêmures subcilíndricos, ligeiramente engrossados na região mediana. Tíbias tão longas quanto os fêmures, duas vezes mais estreita que o fêmur, alargadas na extremidade, carenadas no bordo externo, cerdas curtas dispersas em toda extensão.
Tarsômero I subtriangular, 1/3 mais comprido que o II; III bilobado, IV ligeiramente arqueado com um par de garras apendiculares, afilada e recurvada (Figura 10 e 11).
Segundo GUÉRIN (1953), no gênero Metaxyonycha as pernas são
longas e fracas como constatado em M. angusta. Em ambos os sexos as
pernas apresentam-se semelhantes, aparentemente sem caracteres de importância taxonômica. Contrariamente, SILVEIRA et al. (2000) observaram
que há diferença entre os sexos mediante a presença de um esporão no ápice da mesotíbia de machos tanto em pupas quanto no adulto do crisomelídeo
Coelomera lanio (Dalman) o mesmo também foi observado por WEBER (1976)
em Pyrrhalta luteola (Muller).
Élitros (Figura 1), convexos; três vezes mais longo que largo, estreitando gradativamente na parte posterior, margem posterior apical bilobada, punctuações moderadamente ordenadas em fileiras duplas sob estrias longitudinais; largura humeral um pouco maior que a largura do pronoto.
A presença de estrias e punctuações nos élitros são características descritas por PERTY (1832) e LEFÈVRE (1875) e comuns da família Chrysomelidae, como registrado por PRATHAPAN & KONSTANTINOV (2006) para Orisaltata azurea, Prathapan & Konstantinov por MOSEYKO (2006) ao
15
(Baly), Metaxyonycha chloroptera (Germar) e Metaxyonycha ocanana
(Lefèvre).
Asa membranosa (Figura 12) pelo menos três vezes mais longa que larga. Costa (C) reduzida, restrita a região basal; subcostal (Sc) alcança a metade basal da asa; célula radial (Rc) fechada com aspecto triangular; R-s evidente; árculo (arc) presente; S-m vestigial; sistema cúbito-anal típico. A venação presente nas asas de M. angusta, como constatada deste estudo, é
típica da família Eumolpinae, conforme JOLIVET (1957). Contudo são poucos os trabalhos de descrição que se referem às asas, talvez por serem estruturas difíceis de serem estudadas.
3.1.3 ABDOME
Abdome com cinco segmentos visíveis, em ambos os sexos. Cerdas curtas e uniformes. O abdome ventralmente de coloração amarela-vermelha (7.10 YR), conforme MUNSELL (1952).
Os seis últimos esternitos abdominais, tanto em machos quanto nas fêmeas, não são visíveis por se apresentarem retraídos por baixo do quinto urômero. Estes segmentos se apresentam estreitos e membranosos. Este tipo de estrutura abdominal pode ser visualizada em Demotina medvedeviana
Moseyko (MOSEYKO, 2006), Maecolaspis monrosi Bechyné (CABRERA et al.,
2000).
No quinto uroesternito do macho, a margem posterior apresenta um leve recorte (FERNANDES et al. 2005). Genitália do macho (Figura 13), lobo médio
alongado, esclerotinizado, encurvado lateralmente; extremidade apical estreita (Figura 14), ponta revolvida. No quinto uroesternito da fêmea, a margem posterior apresenta um entalhe mais profundo (FERNANDES et al. 2005).
Ovipositor alongado. Báculo indistinto, mais curto que a gonocoxa. Gonocoxa curta, robusta, com cerdas longas na base; coxitos pequenos, cerdas apicais longas (Figura 15). Espermateca (Figura 16) recurvada, ducto espermatecal fino, esclerotinizado.
A forma do quinto uroesternito abdominal também é diferenciada entre os sexos de Coelomera lanio (Dalman) e Leptinotarsa decemlineata (Say)
16
importante caráter taxonômico para distinguir as subfamílias e gêneros de Chrysomelidae (VARMA, 1955; SUZUKI, 1988). A forma da cápsula da espermateca dos Eumolpinae lembra um gancho (VARMA, 1955), ferradura (SUZUKI, 1988) ou dedo fortemente recurvado (FERRONATTO, 2000). Todas essas designações lembram um mesmo padrão, que também se confirma em
17
4. CONCLUSÃO
M. angusta foi redescrito de forma detalhada evidenciando as
características morfológicas para sua identificação.
18
5. FIGURAS
Figura 1. Desenho esquemático de Metaxyonycha angusta (Perty, 1832), vista
dorsal.
19
20
Figuras 7-11. Metaxyonycha angusta (Perty, 1832). Perna anterior (7); perna
média (8); perna posterior (9); tarso (10) e garra tarsal (11). (Barra: 1,00mm).
21
22
6. REFERÊNCIAS BIBLIOGRÁFICAS
BARTLET, E. ; ROMANI, R.; WILLIAMS, I.; ISIDORO, N.Functional anatomy of sensory structures on the antennae of Psylliodes chrysocephala L. (Coleoptera;
Chrysomelidae). International Jornal of Insect Morphology and
Embryology, Amsterdam, v. 28, n. 4, p. 291-300, 1999.
BECHYNÉ, J. Katalog der neotropischen Eumolpiden (Col. Phytoph.
Chrysomeloidea). Entomologische Arbeiten aus dem Museum George Frey,
Munchen. v.4, p.26-303, 1953.
BORROR, D. J., DELONG, D.M. Introdução ao estudo dos insetos. Tradução de Diva Diniz Correa, Cláudio Gilberto Froehlich, Sérgio de Almeida Rodrigues, Érika Schlenz, Edith Fanta. São Paulo: Edgard Blücher, 1988. 653 p. Tradução de: An introduction to the study of insects.
BROWN, W.J. Notes on North American Chrysomelidae (Coleoptera). The Canadian Entomologist, v. 93, p.967-977, 1961.
CABRERA, N.; LÁZARO, H.; NASCA, A. Caracterización morfológica y presencia de Maecolaspis monrosi Bechyné (Coleoptera: Chrysomelidae: Eumolpinae) en el cultivo de soja en Tucumán. Anais da Sociedade Entomológica do Brasil. v. 9, n.2, p. 277-284, 2000.
CHEVROLAT, L. A.A. Catalogue des Coléoptères de la Collection de M. le Comte. Dejean, Troisiense edition. liv.5, p.385-503, 1837.
23
FERNANDES, L.C., Anjos, N.; Silveira, R. D. Dimorfismo sexual em
Metaxyonycha angusta (Perty, 1832) (Coleoptera: Chrysomelidae). Acta Scientiarum. Biological Sciences, v. 27, n. 2, p. 125-127, 2005.
FERRONATO, E. M. A Cápsula da Espermateca dos Eumolpinae (Coleoptera: Chrysomelidae) Associados a Theobroma cacao L. (Sterculiaceae) do Sul da
Bahia. Anais da Sociedade Entomológica do Brasil v.29, n.1, p 57-63 , 2000. FLOWERS, R.W. La subfamilia eumolpinae (Coleoptera: Chrysomelidae) en America Central. Biologia Tropical, n.2, 60p. 1996. Special publication.
FLOWERS, R.W. A new genus and a new Costa Rican species of
Endocephalites (Chrysomelidae: Eumolpinae: Eumolpini). Zootaxa . v.221, p.
1-11, 2003.
FLOWERS, R.W. Cryocolaspis, a new genus and species of eumolpinae
(Chrysomelidae) from Costa Rica. The Coleopterists Bulletin, v.58, n.1, p.97–
101, 2004a.
FLOWERS, W. Colaspis… Maecolaspis… Metaxyonycha. Whatever an old solution to a persistent problema. Chrysomela n.44,p.13, 2004b.
FLOWERS, R.W. New flightless Eumolpinae of the genera Apterodina Bechyné
and Brachypterodina n. gen. (Coleoptera: Chrysomelidae) from the Neotropics. Zootaxa v. 549, p. 1-18, 2004c.
FLOWERS, R.W. A review of the Neotropical genus Prionodera Chevrolat
(Coleoptera, Chrysomelidae, Eumolpinae) with description of a new genus.
Zootaxa. v.631, p. 1–54, 2004d.
GULLAN, P.J. & CRANSTON, P.S. 2004. The Insects. An Outline of Entomology, 3rd ed. Blackwell Publishing,Oxford.
GUERIN, J. Coleópteros do Brasil. 1953. p. 122-124.
JOLIVET, P. Recherches sur I aile des Chrysomeloidea (Coleoptera). 2eme serie. Mem. Inst. Rech. Sci. Nat. v.58, p. 1-193, 1957.
KUKALOVÁ-PECK, J.; LAWRENCE, J. F. 1993. The evolution of the hind wing in Coleoptera. The Canadian Entomologist v.125, p. 181-258.
LEFÈVRE, E. Descriptions d’Eumolpides nouveaux ou peu connus. Revue et Magasin de Zoologie, v.3, n.3, p.65-74, 1875.
24
MATSUDA, R. Morphology of mouthparts in different orders: the Coleoptera. In: MATSUDA, R. Morphology and evolution of the insect head. Michigan: The American Entomological Institute, 1965. p. 64-68, 1975-184. (Memoirs of the A.E.I., 4).
MOSEYKO, A.G. A new species of the leaf-beetle genus Demotina Baly
Coleoptera: Chrysomelidae: Eumolpinae) from Vietnam. Proceedings of the Russian Entomological Society. v. 77, p. 241–244. 2006
MUNSELL color company. Munsell color charts for plant tissue. Baltimore. 1952. 15 p.
PELLETIER, Y. A method for sex determination of the colorado potato beetle pupa, Leptinotarsa decemlineata. Entomological News, v. 104, n. 1, p.
140-142, 1993.
PERTY, J.A.M. Insecta Brasiliencia. In: Delectus animalium articulatorium, quae itineri per Brasília annis MDCCCXVII – MDCCCXX jusso et auspicis Maximilinai Josiphi I. Bavariae Regis Augustissimi Peracto..., Fasc. 2. p. 108, 1832.
PLOOMI A. ; MERIVEE, E.; RAHI, M.; BRESCIANI, J. ; RAVN , H. P.; LUIK A.; SAMMELSEG V. Antennal sensilla in ground beetles (Coleoptera: Carabidae).
Agronomy Research v.1, n.2, p. 221-228, 2003
PRATHAPAN, K. D. & KONSTANTINOV A. S. Orisaltata, a new genus of flea
beetles from the Oriental Region (Coleoptera: Chrysomelidae). Zootaxa
v.1109,p. 39–47, 2006.
SILVA, A.S., ALMEIDA, L.M. & A.C. BUSOLI. Morfologia dos imaturos e do adulto de Coccidophilus citricola Brèthes (Coleoptera, Coccinellidae,
Sticholotidinae), predador de cochonilhas-de-carapaça (Hemiptera,
Diaspididae) de citros. Revista Brasileira de Entomologia v.49, n.1, p. 29-35, 2005.
SILVEIRA, R.D.; SILVA, N.A.; DELLA LUCIA, T.M.C. Dimorfismo sexual em
Coelomera lanio (Dalman) (Coleoptera: Chrysomelidae). Revista Agro-Ciência, v. 16, n. 2, p. 265-268, 2000.
SUZUKI, K. 1988. Comparative morphology of the internal reproductive system of the Chrysomelidae (Coleoptera). p. 317- 355. In: P. Jolivet , E. Petitpierri & T. H. Hsiao. (eds.), Biology of Chrysomelidae. Amsterdam, Kluwer Academic. VARMA, B.K. Taxonomic value of spermathecal capsules as subfamily characters among the Chrysomelidae (Coleoptera). Indian Journal of Entomology,v. 17, p.189-192, 1955.
WEBER, R.G. Sexing the elm leaf beetle, Pyrralta luteola (Coleoptera:
Chrysomelidae). Annals of the Entomological Society of America, v. 69, n.
25
CAPÍTULO 2: Efeitos do ataque de Metaxyonycha angusta (Perty) (Coleoptera: Chrysomelidae) sobre o crescimento e produção de clones
de eucalipto
RESUMO
Este trabalho teve o objetivo de avaliar os efeitos do ataque de M. angusta
sobre a produção de povoamentos de clones de eucaliptos com um ano de idade. Os estudos foram realizados em plantações de clones de eucaliptos (Eucalyptus urophylla, E. saligna) e híbridos de clones de (E. saligna vs urophylla), no município de Andrelândia, MG. Utilizou-se delineamento em
blocos casualizados, com três repetições de grupos de 25 árvores. Os
tratamentos consistiram em “Árvores sem nenhuma injúria”, “Árvores com injúrias limitadas a poucas folhas”, “Árvores com vários ramos com uma ou
26
estes resultados pode se considerar o besouro M. angusta como praga de
27
ABSTRACT
The damage caused by leaf-beetles may affect tree growth and wood quality. Studies on defoliation effects are not well known for the four-spotted beetle (Metaxyonycha angusta), in eucalypt plantations. This work deals with the
evaluation of the damage effects of different defoliation levels caused by such leaf beetle on cloned eucalypt trees in a 1-year-old plantation of Eucalyptus
urophylla. The diameter and height growth were evaluated up to the 70th month
after the trees were planted. Wood losses were then evaluated according to the different defoliation levels caused by four-spotted beetles in the first year, and the highest value was 40.2%, compared to the control production 59.47 m3/ha
28
1. INTRODUÇÃO
No Brasil, a eucaliptocultura é baseada em culturas clonais, que permite a manutenção plena das características da planta-mãe, de modo a obter a uniformidade e homogeneidade da matéria prima e conseqüentemente, obter ganhos máximos na produtividade volumétrica, chegando a atingir valores da ordem de 60m3. ha-1.ano-1 (MORA & GARCIA, 2000; BORÉM, 2007;
ALFENAS et al., 2004).
Cabe ressaltar que monoculturas de eucalipto, como quaisquer outras, podem favorecer a ocorrência de pragas devido à grande disponibilidade de alimento para os insetos e ao desequilíbrio na estrutura vegetal natural, acarretando uma simplificação do ecossistema (ALMEIDA et al., 1987; PANIZZI
& PARRA, 1991; RISCH,1981; SCHOWALTER et al., 1986).
29
EDWARDS (1991), ignorar a ocorrência das pragas locais pode inviabilizar o empreendimento florestal com espécies exóticas.
Os besouros desfolhadores, entre eles os da família Chrysomelidae, são os mais importantes em termos de expressão econômica para a cultura de
Eucalyptus spp. (OHMART & EDWARDS, 1991 e ANJOS, 1992). Dentre os
insetos desta família, o besouro desfolhador Metaxyonycha angusta (Perty,
1832), popularmente conhecido como “Besouro-de-quatro-pintas”, acarreta grande preocupação pelo hábito de furar as folhas, deixando-as rendilhadas ou, ainda, de devorá-las por inteiro, deixando apenas a nervura principal (FERNANDES, 2004). O ataque nas árvores de eucalipto é realizado, preferencialmente nas partes apicais mais tenras, nas brotações como folhas jovens com até um ano de idade, (FERNANDES, 2004 e ANJOS & MAJER, 2003). Segundo ANJOS (1992), esse hábito alimentar pode provocar alterações na tendência normal de crescimento apical das árvores, prejudicando o seu desenvolvimento.
A resposta ao desfolhamento é de grande interesse principalmente quando tem um alto valor econômico, como o é nos plantios de eucalipto (STRAUSS & AGRAWAL, 1999). Diversos estudos foram feitos a respeito de métodos de avaliação de infestação de besouros desfolhadores em plantações de eucaliptos (KULMAN, 1971), dos danos causados às árvores (COOPER et
al., 1987; PEDIGO, 1989; ABBOTT et al., 1993; MENDES, 2004) e de como
controlar as populações dessas importantes pragas florestais (ELEK et al.,
2000). No Brasil, são poucos os trabalhos relacionados ao impacto do ataque de desfolhadores a árvores de eucalipto (FREITAS, 1988; FREITAS & BERTI FILHO, 1994a; OLIVEIRA, 1996; SILVA et al., 1997; CRUZ, 1997; MENDES,
2004), assim o presente estudo teve como objetivo o de avaliar os efeitos do ataque do Besouro-de-quatro-pintas (M. angusta) sobre o crescimento de
30
2. REVISÃO DE LITERATURA
Como a produção de madeira depende da fotossíntese gerada pelas folhas, qualquer fator que altere a área foliar pode, também, afetar a produção de material lenhoso (CEULEMANS & SAUGIER, 1991). Nestes fatores se inclui o ataque de insetos que, por atuarem diretamente na diminuição da área foliar, acarretam reduções na taxa fotossintética (SPHEPHERD, 1994). Portanto, estes insetos que atacam essências florestais podem representar um fator limitante na produção primária e até mesmo na sobrevivência das árvores cultivadas (KULMAN, 1971; MATTSON & ADDY, 1975; COULSON & WITTER, 1984).
As árvores desfolhadas respondem com a diminuição de folhagem nova e com o crescimento de ramos laterais (DICKSON, 1991). Outros reflexos do desfolhamento mencionados pelo mesmo autor são o aumento da fotossíntese tanto na folhagem residual como nas novas folhas produzidas, que são de tamanho reduzido em relação às anteriores.
As conseqüências no crescimento de árvores desfolhadas por insetos envolvem perdas diretas, como a morte da árvore (MATTSON & ADDY, 1975 e MENDES, 2004) e a redução do crescimento (KRAMER & KOZLOWSKI, 1972; CRUZ, 1997; MENDES, 2004), e indiretas como a alteração da qualidade da madeira (VITAL 1990; MENDES, 2004). Para repor as partes vegetais perdidas
pelo desfolhamento, o uso “desordenado” dos fotoassimilados pode acarretar
31
(crescimento em diâmetro), por ser mais dependente da fotossíntese corrente do que das reservas acumuladas na árvore; já o crescimento em altura é mais dependente das reservas da árvore (KRAMER & KOZLOWSKI, 1972). Com o aumento da capacidade fotossintética da folhagem residual de árvores que passaram por desfolhamento, ocorre uma maior conicidade no fuste de árvores desfolhadas (DICKSON, 1991; MENDES, 2004), pois KRAMER & KOZLOWSKI (1972) constataram que as folhas remanescentes na retenção de galhos na base do fuste das árvores podem alterar a sua forma (MENDES, 1999; ELEK et
al., 2000; De NADAI, 2008).
Vários estudos têm sido realizados sobre os efeitos dos desfolhamentos causados por insetos em eucaliptos (READSHAW & MAZANEC, 1969; ODA & BERTI FILHO, 1978; SCHOWALTER et al., 1986;
ELLIOTT et al., 1992; ANJOS, 1992; SHEPHERD, 1994; RAYMOND, 1995;
MENDES, 2004; De NADAI, 2008; CORDEIRO, 2008) e através de desfolhamentos artificiais, em eucaliptos (CARNE et al., 1974; FREITAS, 1988;
CANDY et al., 1992; FREITAS & BERTI FILHO, 1994a e 1994b; OLIVEIRA,
1996; CRUZ, 1997; SILVA et al., 1997; MATRANGOLO, 1998; SILVA, 1998).
Esses últimos autores, por sua vez, estudaram os efeitos do desfolhamento em árvores de eucaliptos através da simulação do ataque de insetos a elas e, posteriormente, efetuaram as medições do diâmetro a 20 cm de altura (d0,20),
ou à altura do peito (dap) ou, ainda, a circunferência nessa mesma altura (cap) e da altura total de árvores com diferentes intensidades de desfolhamentos.
Para NAUTIYAL & WATERS (1975), as técnicas baseadas na simulação de injúrias são adequadas para avaliação dos impactos causados por insetos desfolhadores quando se conhece a relação entre o hospedeiro e a praga. Entretanto, HEICHEL & TURNER (1976) afirmaram que as plantas desfolhadas por insetos sofrem alterações fisiológicas gradativas, com conseqüente aumento na taxa de transpiração e na produção de enzimas, em relação àquelas que são desfolhadas artificialmente, podendo enfraquecê-las ainda mais do que a súbita remoção de todas as folhas, como ocorre nos desfolhamentos simulados. O mesmo foi mencionado por KRAMER & KOZLOWSKI (1972) e CARNE et al. (1974) sobre as possíveis diferenças entre
32
causados por besouros desfolhadores que persistem nas árvores por algum tempo.
No Brasil, estudos sobre impactos dos desfolhamentos causados por insetos foram feitos por ODA & BERTI FILHO (1978), ANJOS et al. (1987),
ANJOS (1992), MENDES (1999 e 2004), De NADAI (2008) e por CORDEIRO (2008) os quais constataram efeitos na quantidade e qualidade de madeira produzida. Esses estudos buscaram conhecer reações de diferentes espécies, procedências ou clones ao ataque de insetos, sendo importante para a aplicação correta do manejo integrado de pragas (GRAHAM, 1963).
Para ilustrar a importância dos besouros crisomelídeos, ANJOS (1992) constatou perdas de até 33% de madeira em árvores desfolhadas por
Costalimaita ferruginea (Fab., 1801), através de medições realizadas um ano
após um intenso ataque do besouro. Segundo o autor, o crescimento diamétrico foi reduzido em 14,06%, enquanto a redução em altura foi de 7,69%, com conseqüente redução de 33,02% no volume da madeira. Avaliações realizadas em outras condições por MENDES (2004), o qual estudou o comportamento daninho de C. ferruginea em plantios de Eucalyptus grandis,
evidenciaram que o impacto do ataque sobre o crescimento e a produção das árvores aumentava proporcionalmente com as diferentes intensidades de desfolhamento das árvores e com o passar do tempo. Este autor constatou que o ataque de C. ferruginea pode causar perdas de até 43,61% na produção
volumétrica de madeira, aos 84 meses de idade das árvores.
FERNANDES (2004) discorreu sobre o potencial daninho do “Besouro -de-quatro-pintas” em cultura de eucaliptos, porém não mediu o impacto de
33
34
3. MATERIAL E MÉTODOS
O trabalho foi realizado em uma área experimental (teste clonal) de eucalipto plantado no espaçamento inicial 10 x 4 m. O teste clonal foi instalado em 2001 com 13 clones e 3 repetições, em blocos ao acaso. Cada parcela experimental foi constituída de 25 árvores úteis. No primeiro ano, o eucalipto foi consorciado com a cultura de arroz (Oryza sativa: Graminae) e no segundo
com soja (Glicine max: Leguminosae). Esse experimento localizado no município de Andrelândia (“21º 44’ 20” S e 44º 18’ 54” W), na região do Campo
das Vertentes, Estado de Minas Gerais. Ele foi escolhido para esta pesquisa devido à ocorrência natural de um surto de Besouro-de-quatro-pintas,
Metaxyonycha angusta (Perty, 1832), constatado em novembro de 2002
quando a idade das árvores era de 12 meses. Antes e após este surto não foi constatado nenhum outro surto de quaisquer insetos desfolhadores no local de estudos.
Em dezembro de 2002, logo após a ocorrência do surto, as árvores foram avaliadas segundo a intensidade de injúrias na copa das mesmas (Desfolhamento mais o consumo de ponteiros tenros), sendo agrupadas através de notas atribuídas visualmente por três avaliadores. Cada avaliador
atribuiu nota “0” (sem nenhuma injúria), ”1” (injúrias limitadas a poucas folhas), ”2” (vários ramos com uma ou duas folhas atacadas) ou “3” (com a maioria dos
35
tratamentos, foi obtida através de uma média entre as notas atribuídas pelos três avaliadores. A quantidade de árvores agrupadas por nota variou de 3 a 48, por clone, entretanto, alguns clones não apresentaram árvores em todas as notas. Todas as árvores que obtiveram nota média compreendida no intervalo
de “0” até “0,5” correspondem ao tratamento 0 (T0), árvores com notas médias superiores a “0,5” e até “1,5, Tratamento 1 (T1); árvores com notas superiores
a “1,55” e até “2,5”, Tratamento 2 (T2); e as demais correspondem ao
Tratamento 3 (T3). Tais árvores foram então usadas para a avaliação do efeito destas intensidades de injúrias causadas pelo Besouro-de-quatro-pintas sobre o crescimento em altura e diâmetro das tais árvores. Esta técnicas de avaliar visualmente as injúrias é, segundo COOOPER et al., (1987), a melhor técnica
para medir a intensidade de desfolhamento na copa das árvores, em culturas florestais e já foi utilizada por FERNANDES (2004).
A altura total de todas as árvores (Ht) foi obtida com hipsômetro tipo “Haga”. O diâmetro a 1,30m de altura (dap) e a altura da base (d0,20) foram
obtidos a partir da leitura da circunferência, medida a 1,30 m e a 0,20 m de altura ambos com auxílio de uma fita métrica. A partir dos valores de diâmetro e da altura total, foi calculada a produção volumétrica de madeira em pé com casca, por hectare, utilizando o fator de forma médio correspondente aos já utilizados para cada clone nas operações de inventário florestal nas plantações locais. As avaliações do crescimento em altura e diâmetro na altura da base iniciaram logo após o termino do surto, quando as árvores tinham 13 meses de idade (Dezembro/2002). As avaliações do dap tiveram início aos 21 e as
medições seguintes forma aos 27, 46, 58 e 70 meses de idade. A produção por unidade de área (ha) foi obtida com base no nº de árvores e na área útil
por árvore assumida como constante e igual a 40m2.
A partir das análises preliminares, o modelo Richards (RICHARDS, 1959) foi selecionado e ajustado para todas as variáveis (Ht, dap, d0,20 e V),
sendo y(1e( *i))1/ , em que x é a idade em meses, Y é a
36
As curvas de crescimento em altura, dap e d0,20 e as curvas de produção
(m3 .ha-1) foram comparadas empregando testes de identidade de modelos, em
nível de 5% de probabilidade. A estatística F QMRH*QM Res1, com g1,
g2 e g.l., foi utilizada para cada clone, sendo:
comparação na envolvidos tratamento de número NT , NT
g1 4*
resíduo l g NT n
g2 4 4 .
; 4 4 NT SQPR SQPC QMRH completo, modelo o para parâmetros a devido quadrados de Soma SQPC i. trat. e j clone o para s observaçõe de número reduzido modelo o para parâmetros a devido quadrados de Soma
, s SQ SQPRn
)
y
y
(
ji jinji i ji 2 3 0 1 1 Re
O modelo completo, para comparações de curvas estimadas para quatro tratamentos, é definido por:
37
4. RESULTADOS E DISCUSSÃO
Todos os níveis de injúrias causadas pelos besouros M. angusta
às árvores de eucalipto resultaram em alterações significativas no crescimento das mesmas e na produção de madeira da cultura. Evidenciou-se diferença entre as respostas entre os clones, sendo que em algumas variáveis o crescimento apresentou-se com alterações prejudiciais enquanto em outros as alterações foram compensatórias ou o crescimento foi igual ao das árvores não atacadas.
38
Tabela 1- Modelos de regressão, coeficientes ajustados, “S” (desvio padrão da
regressão) e “r” (coeficiente de correlação simples) dos modelos
ajustados para variáveis relacionadas ao crescimento e produção das árvores de clones de eucalipto em função de diferentes intensidades de injúrias causadas por Metaxyonycha angusta e da
idade do povoamento. Andrelândia (MG). 2002 a 2007.
Clone Variável T
Coeficientes da regressão
a b c d S r
01
Altura 1-2-3 20,113466 0,375481 0,054331 0.158193 1,48 0,97
D0,20
1 22,65886 4,707612 0,113902 2,279273 1,19 0,99 2 22,551495 3,206071 0,091991 1,618016 1,37 0,98 3 25,036098 0,241868 0,054649 0,333530 1,54 0,96 DAP 2-3 1 21,299305 21,225754 2,574238 0,767049 0,076748 0,055791 0,990107 0,117742 1,30 1,54 0,98 0,96
Volume
1 160,10218 12,952052 0,238024 3,681237 10,36 0,99 2 201,52981 0,981056 0,048989 0,052802 19,87 0,95 3 209,94358 2,124238 0,047331 0,018532 19,45 0,95
02
Altura
1 24,053768 0,375252 0,049502 0,156636 1,19 0,98 2 22,872457 0,101747 0,057235 0,1911733 1,00 0,99 3 21,018769 1,701932 0,077322 0,5872546 1,22 0,98
D0,20
1 26,687476 1,881483 0,073506 0,785811 1,71 0,96 2 25,239478 2,569180 0,081984 1,227829 1,42 0,99 3 23,851475 3,0757517 0,093013 1,450057 1,32 0,99
DAP
1 23,697348 2,398572 0,082179 0,988746 1,68 0,97 2 23,708152 0,057934 0,082179 0,222197 1,23 0,98 3 22,237204 0,036174 0,065690 0,210242 1,63 0,97 Volume 1-2 3 255,19441 203,35667 1,091702 0,712909 0,050225 0,069247 0,046720 0,170988 15,75 20,10 0,98 0,96
04
Altura
0-1 21,574766 0,6729228 0,068071 0,328045 0,93 0,99 2 22,38239 3,8120939 0,109200 1,432163 1,34 0,98 3 22,95213 2,4125348 0,0824227 0,868733 1,18 0,98
D0,20
0 25,130737 5,431007 0,143781 2,364310 1,33 0,99 1 25,31909 6,918994 0,179465 3,293106 2,26 0,95 2 26,55701 9,6371527 0,235049 4,736540 2,08 0,97 3 27,002547 12,084766 0,288176 5,961867 1,63 0,98 DAP 0-1 2-3 22,298449 22,075469 3,6928343 1,509004 0,079405 0,117048 0,550789 1,341584 1,57 1,29 0,96 0,97
Volume 0-1 2-3 242,37326 224,85181 1,555736 2,337339 0,0928025 0,057557 0,3820882 0,025709 16,00 18,89 0,98 0,97
06
Altura
0 9,98894 0,271221 0,058160 0,159691 0,71 0,99 1 21,940857 0,84488 0,045233 0,115570 1,38 0,97 2 21,369042 0,259485 0,0561283 0,191566 1,83 0,96 3 23,192265 0,306559 0,055333 0,306217 1,95 0,95
D0,20
0 29,67400 1,381808 0,052549 0,072651 1,35 0,98 1 26,019506 2,1461761 0,079656 0,936015 1,98 0,97 2 28,231849 1,576027 0,070690 0,803719 2,71 0,94 3 31,070474 2,693931 0,076881 1,430584 2,74 0,94
DAP
0 26,347738 1,979920 0,040659 0,048886 0,97 0,98 1 23,734988 1,504396 0,048598 0,067004 1,68 0,96 2 24,469071 0,318193 0,053635 0,216362 2,23 0,92 3 23,198617 1,263706 0,080034 0,552477 2,24 0,93
Volume
0-1 282,70115 1,240516 0,037484 0,049579 18,76 0,96 2 252,37263 0,648597 0,049088 0,082018 32,85 0,90 3 275,95663 0,53976 0,053968 0,208394 29,30 0,96
07 Altura
0 21,436285 0,004929 0,048886 0,226735 1,10 0,98 1 24,277219 0,092931 0,042813 0,236576 0,91 0,99 2 25,755111 1,240807 0,040449 0,083746 0,78 0,99 3 24,516982 0,060423 0,050731 0,247278 0,83 0,99
39
...Continuação
Clone Variável T
Coeficientes da regressão
a b c d S r
07
D0,20
0 30,354172 1,886903 0,074114 0,724344 2,05 0,98 1 29,676194 2,5935191 0,083700 1,175139 0,97 0,99 2 29,189928 3,3914633 0,094667 1,688092 1,11 0,99 3 30,387703 1,624419 0,070438 0,781677 1,18 0,99 DAP 0 24,284386 0,076249 0,070164 0,157781 2,22 0,96 1-2-3 27,333022 0,710243 0,051209 0,132590 1,00 0,99
Volume
0 250,50094 1,396011 0,047270 0,033074 12,69 0,99 1 364,77059 0,827559 0,035174 0,073766 12,68 0,98 2 312,68426 0,797536 0,044000 0,064482 11,59 0,99 3 316,09526 0,900344 0,044256 0,061759 11,46 0,99
09
Altura
0 21,58958 0,530163 0,056355 0,129452 0,97 0,99 1 24,228727 0,506567 0,046598 0,154532 1,09 0,99 2 21,104103 0,340214 0,049038 0,154812 1,28 0,96 3 18,425599 0,592695 0,054429 0,118299 1,16 0,99
D0,20
0 24,444902 4,194136 0,111844 2,189167 1,87 0,92 1 26,010926 2,399692 0,081757 1,118578 1,55 0,98 2 23,444991 1,94811 0,335656 9,964765 2,20 0,96 3 21,547649 4,576966 0,128873 1,933192 0,94 0,99
DAP
0 20,656182 1,264303 0,082057 0,462939 2,02 0,94 1 23,015419 0,156802 0,057975 0,268287 1,38 0,97 2 19,634529 1,891338 0,066810 0,029232 1,49 0,97 3 18,878026 0,411572 0,069611 0,250958 1,11 0,97
Volume
0 212,57067 0,011686 0,057906 0,113365 18,66 0,96 1 277,21857 0,421301 0,044264 0,095369 14,60 0,98 2 204,59336 1,686988 0,046978 0,025666 15,64 0,97 3 172,77594 0,403839 0,0531714 0,161203 10,89 0,98
27
Altura
0 21,927328 0,180932 0,050296 0,288877 1,70 0,96 1 22,952354 0,150315 0,046388 0,202725 1,71 0,96 2 25,301457 1,541362 0,042297 0,056702 1,08 0,99 D0,20 1-2 0 28,701492 27,237986 0,946309 1,784384 0,058918 0,072848 0,507530 0,759753 1,37 1,32 0,99 0,99
DAP 0-1 27,202269 0,955217 0,042528 0,119984 1,37 0,98 2 28,161595 1,106184 0,041692 0,098654 1,26 0,98 Volume 1-2 0 295,47906 254,5206 0,377793 4,759206 0,044491 0,093064 0,191403 1,213506 19,48 17,26 0,97 0,98
28
Altura 0-3 1 21,767229 26,86079 0,675299 0,908453 0,040832 0,051091 0,143595 0,112331 0,96 2,23 0,99 0,93
D0,20 0-1 3 34,858012 30,587427 0,229560 1,034648 0,049411 0,066864 0,216786 0,495038 1,95 2,55 0,99 0,96
DAP
0 27,556949 1,271334 0,058169 0,065722 0,72 0,99 1 27,707038 1,295619 0,054379 0,072069 2,57 0,93 3 32,084132 0,855118 0,043310 0,130513 1,37 0,98 Volume 0-1 3 295,50023 302,6895 6,715975 0,96976 0,118141 0,04516 0,059076 1,980271 35,71 15,54 0,91 0,99
29
Altura 0-1 21,68906 0,36816 0,051068 0,183523 1,52 0,97 2-3 17,60218 0,53441 0,072004 0,275945 1,88 0,95
D0,20
0 29,56788 1,696823 0,070061 0,772588 1,99 0,97 1 30,76307 2,438866 0,066084 1,237415 2,38 0,95 2 28,71113 0,38872 0,045806 0,191351 1,92 0,97 3 23,30413 1,261016 0,07302 0,539309 1,26 0,98
DAP
0 25,02417 0,91164 0,04955 0,124118 1,47 0,97 1 25,03281 0,44603 0,047112 0,191663 1,72 0,96 2 20,80693 0,127444 0,061857 0,24477 1,48 0,97 3 19,78547 0,249241 0,058892 0,280913 1,17 0,97
Volume
0 265,44636 1,013648 0,044942 0,059849 20,96 0,96 1 266,86588 0,452676 0,048683 0,195266 20,81 0,96 2 179,57365 0,186137 0,054962 0,108524 20,23 0,94 3 164,47323 0,616520 0,0636146 0,173394 11,99 0,98
Continua...
40
Clone Variável T
Coeficientes da regressão
a b c d S R
31
Altura 0-1 2 20,452637 17,450507 0,276603 6,566774 0,058249 0,132856 0,253203 2,880372 1,56 0,94 0,97 0,98
D0,20
0 27,814658 3,151408 0,087397 1,4304943 2,27 0,96 1 29,712131 2,976436 0,085493 1,359674 2,60 0,96 2 25,110121 4,680556 0,121346 2,121049 3,14 0,94 DAP 0-2 25,406492 0,166887 0,059363 0,171421 2,74 0,92 1 27,674732 0,183614 0,053883 0,189958 7,84 0,94
Volume
0 228,14533 0,126507 0,056318 0,093301 26,27 0,94 1 228,85556 2,797888 0,078028 0,599602 26,75 0,94 2 186,5228 2,50916 0,075592 0,500577 26,95 0,94
32
Altura 0 1 23,315945 22,070069 1,041574 1,203314 0,072904 0,075043 0,350274 0,390002 1,21 1,28 0,99 0,98 2 21,902687 1,237494 0,074675 0,386871 0,98 0,99
D0,20
0 28,159657 3,779149 0,111996 1,567302 1,72 0,98 1 26,727927 2,867568 0,100648 1,082596 1,63 0,98 2 26,678553 2,476987 0,091216 0,903894 0,94 0,99
DAP
0 25,615799 30,605639 0,63566026 15,482932 1,89 0,97 1 23,750688 0,993555 0,078202 0,365577 1,80 0,96 2 23,76595 0,717668 0,070671 0,315206 0,63 0,99
Volume
0 268,41086 7,098500 0,161935 1,574785 23,08 0,98 1 255,38249 0,228016 0,068810 0,105329 24,89 0,96 2 256,68169 0,103393 0,063514 0,082371 11,42 0,99
33
Altura 0 19,380957 1,819841 0,083119 0,606878 1,38 0,97 1 19,742921 3,607096 0,115476 1,1605 1,06 0,98 D0,20 0 22,354984 5,156323 0,138055 2,488129 1,19 0,98 1 22,80444 4,622447 0,133332 1,918419 1,95 0,96 DAP 0-1 21,74123 1,135861 0,066379 0,066931 2,25 0,93 Volume 0 1 181,5961 196,981 0,56795 0,3775 0,071497 0,076925 0,052165 0,051656 22,47 22,04 0,93 0,96
41
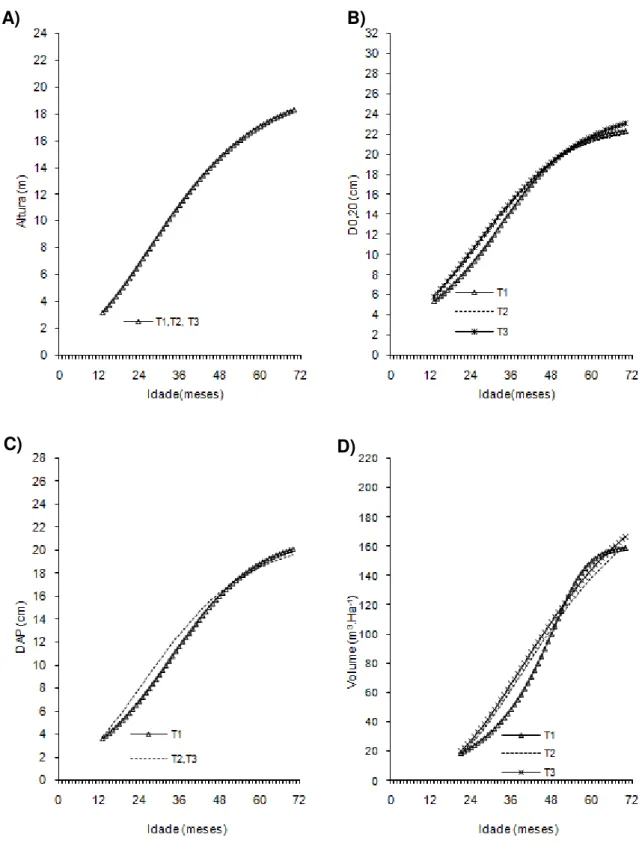
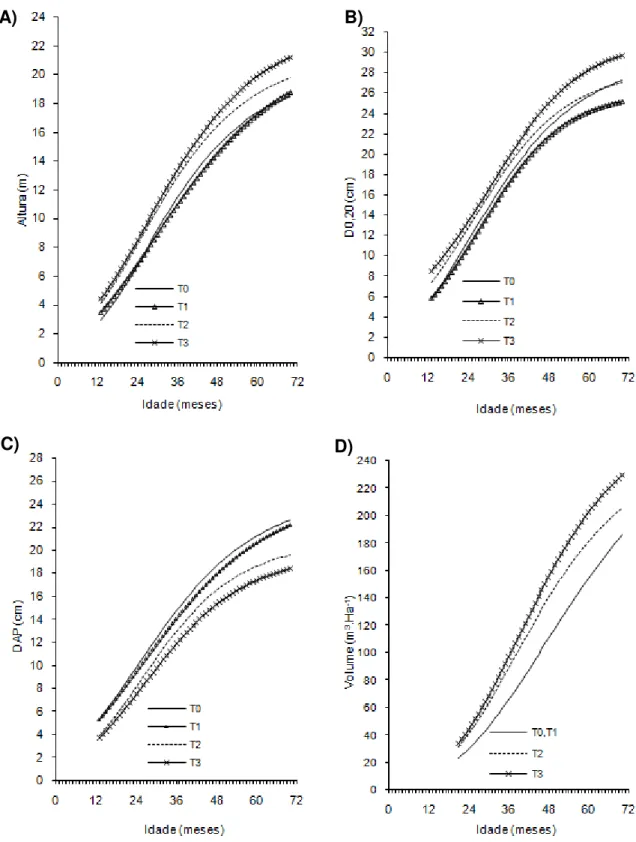
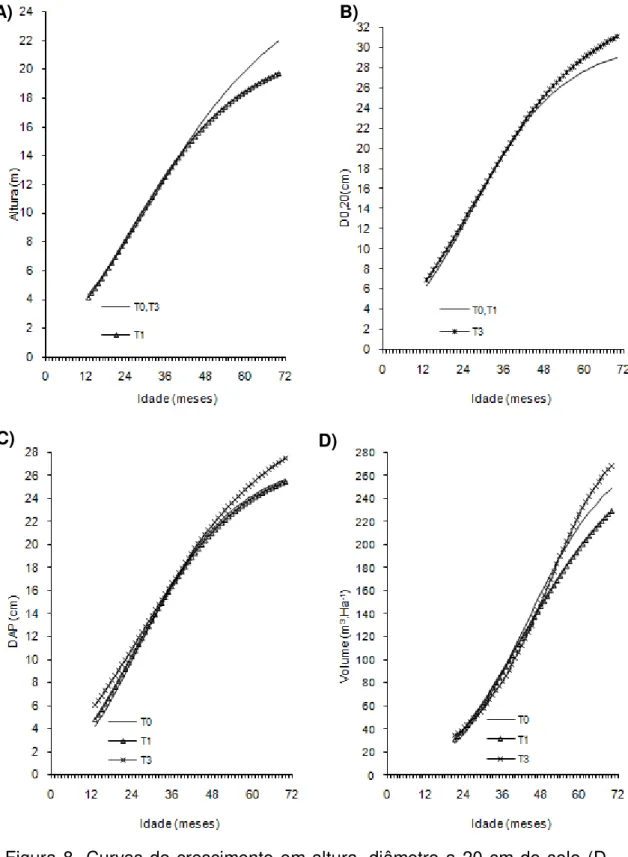
Figura 1- Curvas de crescimento em altura, diâmetro a 20 cm do solo (D0,20),
diâmetro a 1,30cm do solo (Dap) e volume de árvores do clone 01 de eucalipto após diferentes intensidades de injúrias causadas por
Metaxyonycha angusta Perty (1832) (T0= árvores com notas médias compreendidas no intervalo de “0” até “0,5”; T1= árvores com notas médias superiores a “0,5” e até “1,5”; T2= árvores com notas
superiores a “1,55” e até “2,5” e as demais correspondem ao T3).
Andrelândia, MG, 2002 a 2007.
A) B)
42
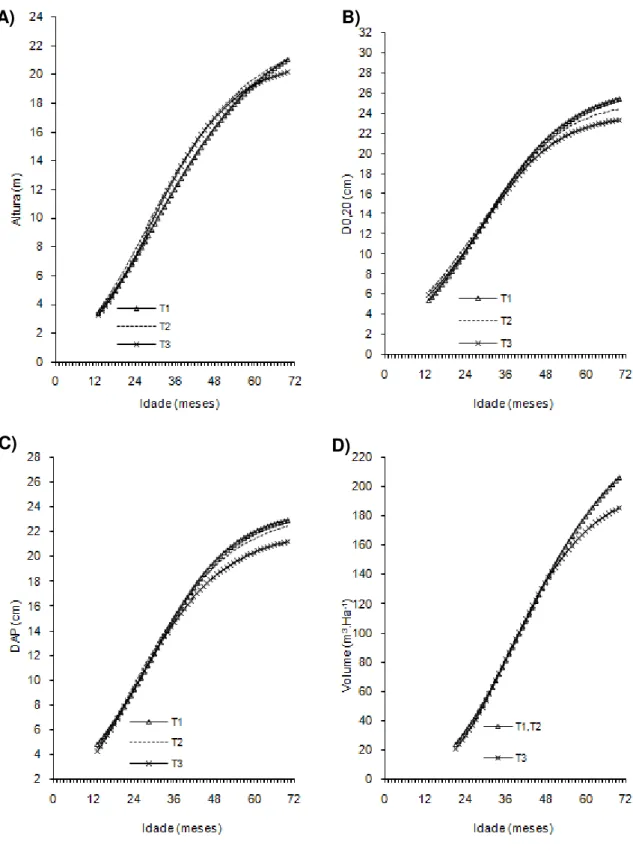
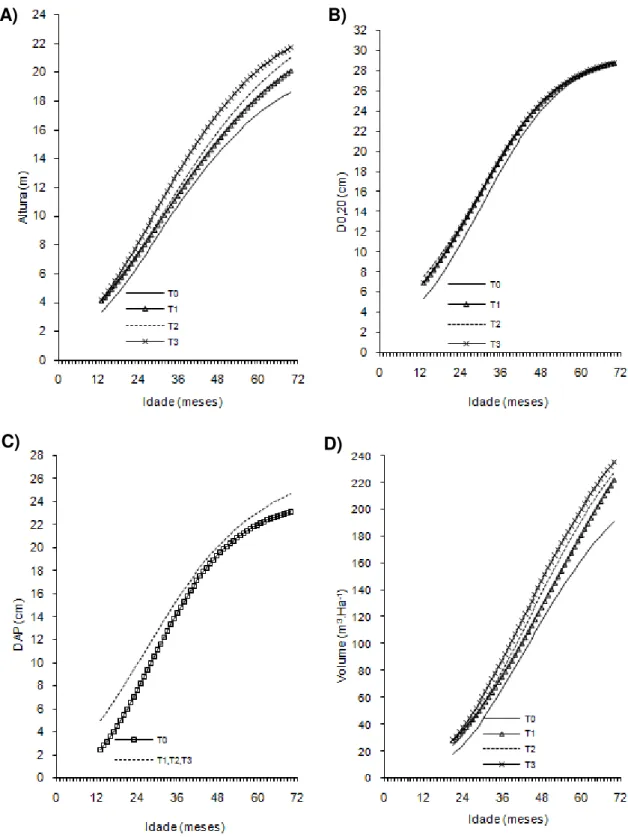
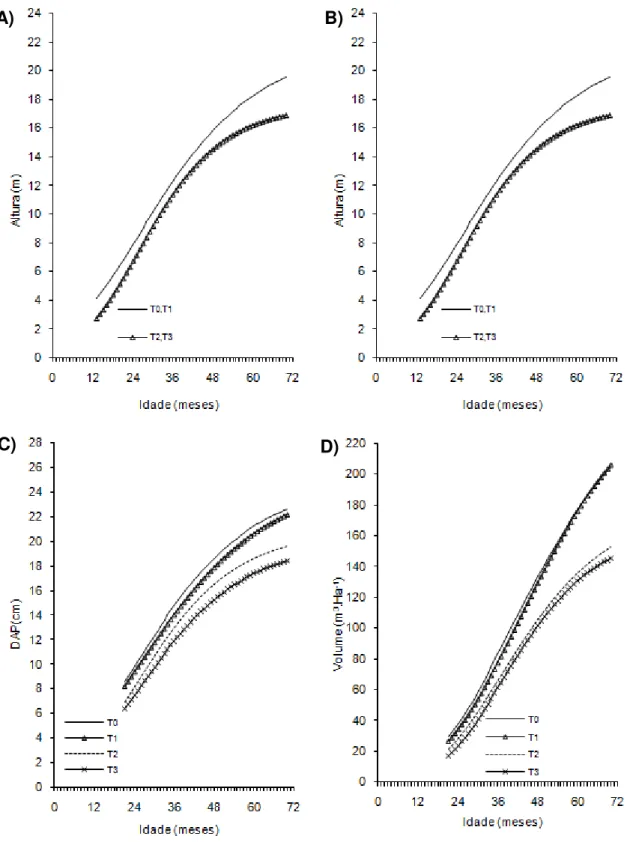
Figura 2- Curvas de crescimento em altura, diâmetro a 20 cm do solo (D0,20),
diâmetro a 1,30cm do solo (Dap) e volume de árvores do clone 02 de eucalipto após diferentes intensidades de injúrias causadas por
Metaxyonycha angusta Perty (1832) (T0= árvores com notas médias compreendidas no intervalo de “0” até “0,5”; T1= árvores com notas médias superiores a “0,5” e até “1,5”; T2= árvores com notas
superiores a “1,55” e até “2,5” e as demais correspondem ao T3).
Andrelândia, MG, 2002 a 2007.
A) B)
43
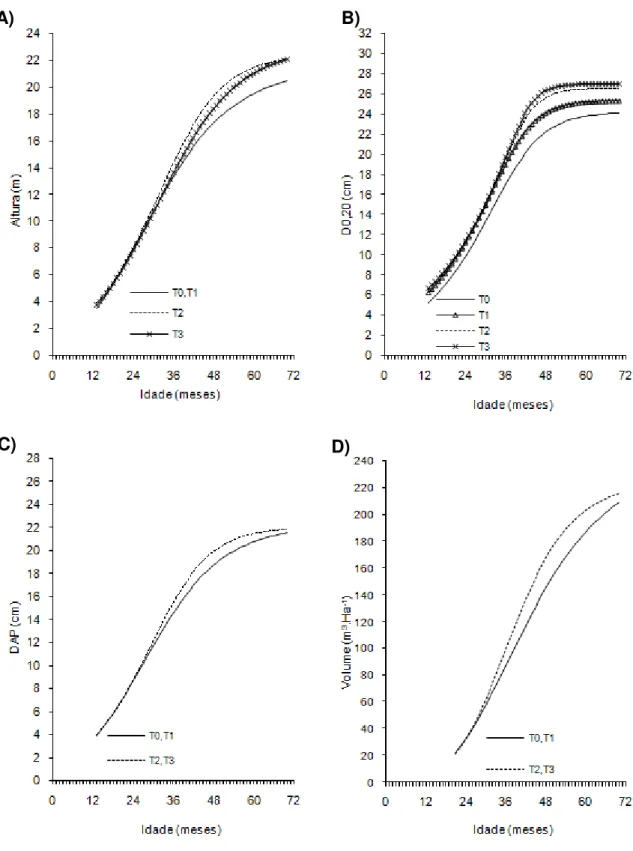
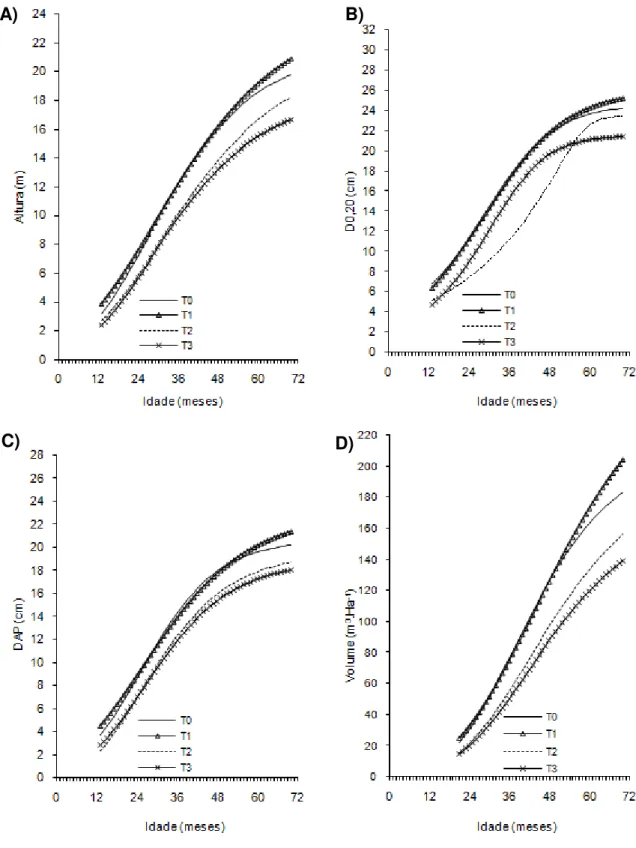
Figura 3- Curvas de crescimento em altura, diâmetro a 20 cm do solo (D0,20),
diâmetro a 1,30cm do solo (Dap) e volume de árvores do clone 04 de eucalipto após diferentes intensidades de injúrias causadas por
Metaxyonycha angusta Perty (1832) (T0= árvores com notas médias compreendidas no intervalo de “0” até “0,5”; T1= árvores com notas médias superiores a “0,5” e até “1,5”; T2= árvores com notas
superiores a “1,55” e até “2,5” e as demais correspondem ao T3).
Andrelândia, MG, 2002 a 2007.
A) B)
44
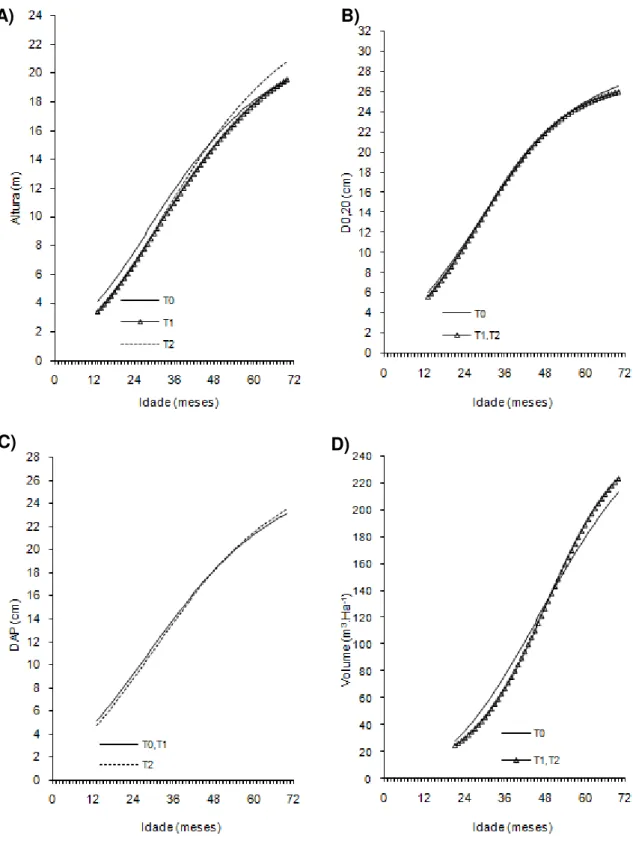
Figura 4- Curvas de crescimento em altura, diâmetro a 20 cm do solo (D0,20),
diâmetro a 1,30cm do solo (Dap) e volume de árvores do clone 06 de eucalipto após diferentes intensidades de injúrias causadas por
Metaxyonycha angusta Perty (1832) (T0= árvores com notas médias compreendidas no intervalo de “0” até “0,5”; T1= árvores com notas médias superiores a “0,5” e até “1,5”; T2= árvores com notas
superiores a “1,55” e até “2,5” e as demais correspondem ao T3).
Andrelândia, MG, 2002 a 2007.
A) B)