FACULDADE DE FILOSOFIAD CIÊNCIAS E LETRAS DE RIBEIRÃO PRETO
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
MISAEL FERNANDO GARCÍA CARRILLO
Alterações da Morfologia Dendrítica e Epilepsia: Uma aproximação neurocomputacional
MISAEL FERNANDO GARCÍA CARRILLO
Alterações da Morfologia Dendrítica e Epilepsia: Uma aproximação neurocomputacional
Tese apresentada à Faculdade de FilosofiaD Ciências e Letras de Ribeirão Preto – USPD como parte das
exigências para obtenção do título de Mestre.
Área de concentração: Psicobiologia Orientador: Prof. Dr. Carlos Antônio
Roque da Silva Filho
AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE TRABALHOD POR QUALQUER MEIO CONVENCIONAL OU ELETRÔNICOD PARA FINS DE ESTUDO E PESQUISAD DESDE QUE CITADA A FONTE.
Ficha Catalográfica García CarrilloD Misael Fernando
Alterações da Morfologia Dendrítica e Epilepsia: Uma aproximação
Neurocomputacional / Misael Fernando García Carrillo; Orientador Prof. Dr. Carlos Antônio Roque da Silva Filho – Ribeirão PretoD 2012.
49 f.
Tese (Mestrado – Programa Pós-graduação em Ciências. Área de concentração de Psicobiologia) Faculdade de FilosofiaD Ciências e Letras de Ribeirão Preto –
Universidade de São Paulo.
FOLHA DE APROVAÇÃO
Misael Fernando García Carrillo
Alterações da Morfologia Dendrítica e Epilepsia: Uma aproximação neurocomputacional
Tese apresentada à Faculdade de FilosofiaD Ciências e Letras de Ribeirão Preto – USPD como parte das
exigências para obtenção do título de Mestre.
Área de concentração: Psicobiologia
Aprovado em:
Banca Examinadora
DEDICATÓRIA
AGRADECIMENTOS
Ao Prof. Carlos Antônio Roque da Silva FilhoD pela sua orientaçãoD as suas ensinançasD e ter me aceito como seu aluno.
À Universidade de São PauloD pela oportunidade de fazer o meu mestrado.
A CAPESD pelo apoio financeiro.
RESUMO
GARCÍAD M.F. Alterações da Morfologia Dendrítica e Epilepsia: Uma aproximação neurocomputacional. 2012. 49f. Tese (Mestrado) – Faculdade de FilosofiaD Ciências e Letras de Ribeirão PretoD Universidade de São PauloD Ribeirão PretoD 2012.
Pesquisas in vivo e in vitroD têm estabelecido uma correlaçãoentre alterações na
morfologia dendrítica e a epilepsia. No entantoD ainda não se conhecem em detalhe as consequências dessas modificaçõesD sobre a eletrofisiologia e o padrão de disparo. Também existe um fenômeno que não tem sido completamente explicadoD conhecido como o paradoxo do dendrito epilépticoD no qual neurônios piramidaisD mesmo com a diminuição dramática do principal lugar de inervação glutamatérgica (como consequência deD por exemploD uma redução do diâmetro e comprimento das árvores dendríticas)D inesperadamente apresentam um estado de hiperexcitabilidade crônica.
Nesta pesquisa foram aproveitadas as vantagens de uma aproximação neurocomputacionalD para induzir sistematicamente alterações na arquitetura dendrítica do mesmo tipo às observadas na epilepsiaD e avaliar os seus efeitos sobre a eletrofisiologia e o padrão de disparo. Para isso foi construído um modelo computacional biologicamente realistaD de um neurônio piramidal do neocórtex. O código-fonte do modelo está na linguagem do NEUROND e foi baseado em dados eletrofisiológicos (i.e. propriedades da membrana e condutâncias iônicas) e morfométricosD obtidos in vitro previamente por outros pesquisadores. A análise foi feita
AVSTRACT
GARCÍAD M.F. Dendritic Morphology Alterations and Epilepsy: A Neurocomputational Approach. 2012. 49f. Tese (Mestrado) – Faculdade de FilosofiaD Ciências e Letras de Ribeirão PretoD Universidade de São PauloD Ribeirão PretoD 2012.
Invivo and in vitro studiesD had found a correlation between dendritic morphology alterations and epilepsy. NeverthelessD it has not been established in detail the consequences of those modificationsD over the electrophysiology and firing pattern. There is also a phenomenon still not completely understoodD known as “the epileptic dendrite paradox”D in which pyramidal neurons with a dramatic reduction in the principal
place of glutamatergic innervation (due toD for exampleD a loss in dendritic trees' diameter and length)D unexpectedly present a chronic hyperexcitable state.
SUMÁRIO
1 INTRODUÇÃO 12
1.1 Árvores Dendríticas: Processamento de Informação e Eletrofisiologia 12
1.2 Epilepsia e Alterações da Morfologia Dendrítica 14
1.3 O paradoxo do dendrito epiléptico 18
2 OVJETIVOS 19
3 MATERIAL E MÉTODO 20
3.1 Sujeito 20
3.2 Equipamentos 21
3.3 Procedimento 22
3.4 Análise de dados 25
4 RESULTADOS E DISCUSSÃO 27
4.1 Correlação de Pearson e Spearman entre variáveis morfológicas e parâmetros do padrão de disparo 27
4.2 Redução do Comprimento Árvore Apical 28
4.4 Redução do Diâmetro Árvore Basilar 36 4.5 Redução do Comprimento Árvore Basilar 37 4.6 Interação entre variáveis morfológicas e efeito combinado
sobre padrão de disparo 38
6 CONCLUSÕES 45
1 INTRODUÇÃO
1.1 Árvores Dendríticas: Processamento de Informação e Eletrofisiologia
Pesquisas acerca da função dos dendritosD iniciadas com os trabalhos pioneiros de Rall (1995)D têm tornado possível estabelecer a importância dessas estruturas para a integração dos sinais sinápticas eD portantoD para o processamento de informação no cérebro. De fatoD estudos recentes têm demonstrado o papel dos dendritos em importantes processos comoD por exemploD a integração e o processamento da informação sináptica (YUSTE e TANKD 1996; SINGLE et al.D 1997; SINGLE e BORSTD 1998; MAGEED 2000; LONDON e HÄUSSERD 2005; BROWN et al.D 2008; STUART et al.D 2008;)D a transmissão glutamatérgica (ENGEL e PEDLEYD 2008)D a potenciação a longo prazo (HÄUSSERD 2004; MEHTAD 2004; SEGALD 2005; STUART et al.D 2008) e outras mudanças plásticas que subjazem a processos de aprendizagem e memória (RALLD 1995; SCHWARTZKROIND 2009).
pode ser entre 10 a 20 vezes maior que a do corpo celular em neurônios motores e piramidais (AITKEN 1955; SHADÉ e BAXTERD 1960; SHOLLD 1955; YOUNGD 1958) eD em alguns casosD até cem vezes maior como nas células de Purkinje (FOX e BARNARDD 1957).
As características dos dendritos mais estudadas em termos de seu efeito sobre o processamento neuronal têm sido (1) a classe morfológica geral à qual o neurônio pertenceD i.e. piramidalD estreladoD etc (McCORMICK et al.D 1985; BASTIAN e NGUYENKIMD 2000; VETTER et al.D 2001)D e (2) os tiposD as distribuições e as densidades das condutâncias dos canais iônicos presentes na árvore dendrítica (TRAUBD R.D.D 1979; KIM e CONNORSD 1993; LERCHE et al.D 2001; DOIRON et al.D 2002; MIZIELINSKAD S.M.D 2007).
A extraordinária diversidade de morfologiasD composição e plasticidade dos dendritosD além das suas especializações sinápticasD sugerem que as contribuições funcionais destas estruturas à mente e o cérebro são enormemente variadas (STUART et al.D 2008). No entantoD ainda pouco se conhece acerca de como mudanças na estrutura dendríticaD se associam com propriedades funcionais dos neurôniosD tais como a integração sinápticaD a excitabilidadeD e os padrões de disparo (STUART et al.D 2008).
1.2 Epilepsia e Alterações d a Morfologia Dendrítica
A epilepsia é uma condição devastadoraD e uma das patologias neurológicas com maior prevalênciaD afetando aproximadamente 1% da população (HAUSER et al.D 1991; ENGEL e PEDLEYD 2008; SCHWARTZKROIND 2009).
A epilepsia envolve dois fenômenos muito interessantes: (1) A hiperexcitabilidade na qualD através de dinâmicas que têm a ver com a despolarização e repolarização da membrana (ENGEL e PEDLEYD 2008)D e que ainda não se conhecem completamenteD o neurônio pós-sináptico fica muito mais sensível à estimulação elétrica de um neurônio pré-sinápticoD gerando respostas com uma intensidade e frequência muito mais fortes que as de um neurônio saudável. (2) Na hipersincronicidadeD um grupo de neurônios exibem um padrão de disparo altamente sincrônico entre elesD levandoD entre outras coisasD à propagação da atividade epileptiforme a outras áreas do cérebro (ENGEL e PEDLEYD 2008). Acredita-se que a hipersincronicidade tem a ver com um defeito nos mecanismos de integração neuralD como consequência de alterações na eletrofisiologia dos neurônios.
Estes dois fenômenos são fundamentais para o entendimento da Epilepsia eD portantoD a Liga Internacional contra a Epilepsia (International Leaghe against Epilepsy
– ILAE)D define este doença como a ocorrência transiente de signos ou sintomasD como consequência de atividade elétrica e sincrônica excessiva no cérebro (ENGEL e PEDLEYD 2008).
diâmetro e comprimento dos segmentosD e uma perda na complexidade das ramificações) com a epilepsia (WESTRUM et al.D 1964; SCHEIBEL et al.D 1974; MULTANI et al.D 1994; COLLING et al.D 1996; JIANG et al. 1998; SWANN et al.D 2000; WASHINGTON et alD 2000; SCHWARTZKROIND 2009). Por exemploD o Jiang et al. (1998) no modelo da epilepsia pela neurotoxina tetânicaD encontraram que neurônios piramidais apresentam uma redução promédio de 24% no comprimento da árvore dendrítica basilar. Também uma diminuição de 48% no diâmetro máximo da árvore apical e 33% da árvore basilarD junto com a redução de 55% e 35% no diâmetro mínimo da árvore apical e basilarD respectivamente. Ainda não se conhece em detalhe as consequências eletrofisiológicas desse tipo de modificaçõesD portantoD estas pesquisas oferecem importantes subsídios biológicos a partir dos quaisD é possível avaliar sistematicamente as consequências dessas anormalidadesD através de uma modelação computacional realista.
Não obstante várias pesquisas tenham correlacionado modificações na morfologia dendrítica com a epilepsiaD ainda não se sabe com precisão se essas correlações implicariam uma relação causalD nem a direção desta possível causalidadeD isto éD se as alterações observadas seriam consequência da atividade patológicaD ou se estas alterações poderiam levar à geração de atividade epileptiforme.
Algumas pesquisasD por exemploD sugerem que as mudanças dendríticas observadas seriam uma consequênciaD devido a que a atividade elétrica epileptiforme levaria a uma alteração de mecanismos intrínsecosD que têm a ver com a regulação e manutenção do citoesqueleto dos dendritos (KURZ et al. 2008; SWANN et alD 2000; ZENG et al. 2007). Sabe se tambémD que a plasticidade da árvore é altamente dependente do ambiente local (STUART et al.D 2008). Por exemploD entre os fatores de maior influencia sobre a morfologia dendrítica está o fornecimento de novas conexõesD pela entrada de sinais excitatórios pré-sinápticosD tal como tem sido estabelecido por estudos comoD por exemploD os desenvolvidos pelo Globus (1975). Segundo outras teoriasD existe a possibilidade de que as mudanças encontradas na morfologia dendríticaD além de ser uma consequência da epilepsiaD possam agir como algum tipo de mecanismo compensatório ou homeostáticoD com o fim de reduzir a hiperexcitabilidade dos neurônios epileptogênicos (STUART et al.D 2008; SWANND 2008).
crônico de hiperexcitabilidade. De fato para alguns pesquisadoresD as alterações na árvore dendrítica podem estar acompanhadas por mudanças em vários níveisD comoD por exemploD na expressão molecularD e no lugar de implantação de canais iônicos nos dendritos eD por consequênciaD contribuir na hiperexcitabilidade dos neurônios (SWANN et al.D 2000). Além dissoD um estudo feito por WASHINGTON et al. (2000)D estabelece que um menor tamanho da árvore dendrítica leva a uma maior excitabilidade. Isso faria possível pensar na existência de uma relação causalD entre as alterações dendríticas observadas e a epilepsiaD na qual essas mudanças poderiam levar a um aumento da excitabilidade eD portantoD à geração de atividade elétrica patológica.
1.
3 O paradoxo do dendrito epiléptico
Por que neurônios piramidais com menor área dendrítica total (i.e. perda de diâmetroD comprimentoD espinhas e complexidade das ramificações) eD por consequênciaD menor entrada de sinais excitatórias do tipo glutamatérgicoD inesperadamente apresentam maior excitabilidade? Este fenômeno se conhece como o paradoxo do dendrito epiléptico (the epileptic dendrite paradox) (SCHWARTZKROIND
2 OVJETIVOS
1) Induzir e avaliar sistematicamente a partir de uma aproximação neurocomputacionalD as consequências na eletrofisiologia e padrão de disparo de alterações morfológicas nas árvores dendríticasD do mesmo tipo às observadas in vivo e in vitro em neurônios
piramidais epileptogênicos.
2) Indagar teórica e computacionalmenteD na possibilidade de uma relação causal nas correlações encontradasD junto com a possível direção na qual se daria.
3 MATERIAL E MÉTODO
3.1 Sujeito
Modelo computacional realista biofisicamente detalhado de um neurônio piramidal (Figura 1)D o qual foi representado computacionalmente como na Figura 2. As árvores dendríticas apical e basilar foram modeladas segundo a lei da potência dos diâmetros do Rall (1995)D fazendo possível colapsar todas as ramificações num só compartimento para cada árvore (Figura 3). O modelo computacional está composto por quatro compartimentosD que representam a árvore basilarD o axônioD o soma e a árvore apical (Figura 3). Este modelo realista possui as propriedades biofísicas descritas na Tabela 1.
Figura 2. Representação do neurônio piramidal com as árvores dendríticas sem serem colapsadas. Na esquerda localiza-se a árvore dendrítica apicalD no centro o somaD e na direita a árvore dendrítica basilar.
Figura 3. Estrutura das árvores dendríticas apical e basilarD colapsadas cada uma segundo a lei do Rall (1995) num só compartimento. Na esquerda localiza-se a árvore dendrítica apicalD no centro o somaD e na direita a árvore dendrítica basilar.
3.2 Equipamentos
Cluster de dez nós processadoresD do Laboratório de Sistemas Neurais (SisNe)D coordenado pelo orientador do projeto.
3.3 Procedimento
A partir do software NEUROND sua linguagem (hoc) e baseado na programação orientada ao objeto (“object oriented”)D foi construído um modelo computacional realista de um neurônio piramidal (Figuras 1D 2D 3). Os dendritos das árvores apical e basilarD foram colapsados em um só compartimento cilíndrico para cada árvoreD segundo a lei do Rall (1995) (Figura 3). Neste modelo foram usadas propriedades passivas para as árvores dendríticas (Tabela 1). No caso do soma e do axônio foram usadas propriedades ativasD com base no modelo do Hodgkin e HuxleyD para geração e propagação ativa dos potenciais de ação (HODGKIN e HUXLEYD 1952) (Tabela 1).
COMPARTIMENTO COMPRIMENTO (um) DIÂMETRO (um) PARÂMETROS VIOFÍSICOS
Soma 18.8 18.8
Propriedade Ativas: Hodgkin e Huxley gnabar_hh = 0.120 S/cm2
gkbar_hh = 0.036 S/cm2 gl_hh = 0.0003 S/cm2 ena = 50 mV
ek = -77 mV el_hh = -54.3 mv
Árvore Apical 710.9 3.18
Propiedades passivas com Ra = 123 ohm-cmD
E = -65mV
Árvore Basal 549.1 2 Mesmas propriedades passivas árvore apical
Axônio 1000 1 Mesmas propriedades ativas do soma
Todos os
compartimentos N/A N/A Cm = 1uF/cm2; RC= 100 Ohms*cm; T = 6.3C
O protocolo de estimulação foi um IClamp de corrente contínua despolarizanteD cujos parâmetros são descritos na tabela 2. Esse protocolo de estimulação foi usado para permitir a geração continua de potenciais de ação durante tudo o tempo de estimulação.
PARÂMETROS DO ICLAMP
Amplitude da estimulação 0.1nA (nanoamperes) Tempo inicio 100 ms (milissegundos) Duração estimulação 900 (milissegundos) Localização eletrodo 50% do comprimento total árvore apical
Tabela 2. Parâmetros da estimulação com eletrodo.
O modelo foi construído de modo que alguns dos parâmetrosD que caracterizam a morfologia das árvores dendríticas apical e basilarD além dos parâmetros que têm a ver com o tipo de estimulaçãoD pudessem ser modificados sistematicamente para a realização dos estudos propostos. Os parâmetros morfológicos que foram avaliadosD com base no tipo de alterações observadas na epilepsiaD foram o diâmetro e comprimento das ramificações da árvore apical e da árvore basilar. Todos os mecanismos e parâmetros biofísicos relacionados com as dinâmicas dos canais iônicosD foram mantidos constantes para todas as morfologias neurais.
alta complexidade na qual ao correr o código principalD só é necessário introduzir no prompt do MaltabD os vetores com as magnitudes porcentuais (em números decimais)D da redução exercida sobre os valores originais dos parâmetros morfológicos avaliados (i.e. diâmetros e comprimentos árvore apical e basilar). Tudo isso foi feito trabalhando diretamente com a linguagem do NEURON e do MATLABD sem fazer uso da interface gráficaD o qual permitiu induzir sistematicamente varias mudanças morfológicasD fazer combinatórias entre as distintas variáveisD e ter maior versatilidade para uma avaliação e análise muito mais abrangente.
Já tendo a representação do neurônio piramidal no código hoc, além de toda a
plataformaD foram induzidas sistematicamente alterações no diâmetro e comprimento da árvore dendrítica e da árvore basilar. Essas modificações morfológicas foram do mesmo tipo observado na epilepsia. Com esse fimD em cada iteração foi reduzido num 2.5% a magnitude de cada uma das variáveis morfológicas separadamente (84 iterações/novas morfologias neurais)D ou fazendo combinatórias entre elas (1764 iterações/novas morfologias neurais)D para simular uma situaçãoD por exemploD na qual as duas árvores perdem diâmetro ou se recolhem simultaneamenteD tal como é observado em neurônios epileptogênicos. A simulação foi feita até atingir uma redução de 50% nas variáveis morfológicas de interesse.
inter-disparoD baseado no promédio do tempo entre os picos de cada potencial de ação gerado (máximos locais). (3) Comprimento promédio do potencial de açãoD definido como o promédio da duração entre o ponto no qual a voltagem de membrana aumenta positivamenteD até atingir o patamar para a geração do potencial (-55mv)D e o tempo no qual a voltagem de membrana retorna ao potencial de repouso (-65mv) (4) Amplitude promédio da despolarização (promédio valor máximos locais da voltagem). (5) Amplitude promédio da hiperpolarização depois da geração do disparo (promédio valor mínimos locais da voltagem). (6) Taxa de disparos durante a janela de análiseD isto éD o tempo desde o início da estimulação e durante um intervalo de tempo de 800ms (100ms antes de finalizar a simulação).
3.4 Análise de dados
Os primeiros 100 milissegundos da simulação não foram levados em conta na análiseD para evitar artifícios e permitir que se estabilizasse o potencial de membrana. Foi criada uma janela de análise de 800ms entre t = 100ms e t = 900msD dos 1000ms totais da simulação.
4 RESULTADOS E DISCUSSÃO
4.1 Correlação de Pearson e Spearman entre variáveis morfológicas e parâmetros do padrão de disparo.
Na análise inicial dos resultados obtidosD foi possível estabelecer uma correlação muito forte para cada uma das interações testadasD entre as variáveis morfológicas e os parâmetros usados para quantificar diferenças no padrão de disparo (Tabela 3 e 5). No caso do coeficiente de Spearman (Tabela 4)D as correlações apresentaram uma magnitude maior para todas as associações avaliadasD o que permite inferir que o efeito das diferentes morfologias sobre o padrão de disparoD é melhor explicado em termos não lineares.
Tabela 3. Correlação de Pearson para a interação entre variáveis morfológicas (dAA = diâmetro árvore apical; dAB = diâmetro árvore basilar; CAA = comprimento árvore apical; CAB = comprimento árvore basilar) e os parâmetros do padrão de disparo (LGPD = latência para a geração do primeiro disparo; PIID = promédio do intervalo inter-disparo; CPPA = comprimento promédio do potencial de ação; APD = amplitude promédio da despolarização; APR= Amplitude promédio da repolarização e Taxa de disparos)
Correlação (Pearson) Variáveis Morfológicas e Parâmetros Padrão de Disparo
LGPD PIID CPPA APD APR Taxa Disparos
dAA 0.9494 0.9849 0.9878 -0.9970 0.9995 -0.9838
dAV 0.9947 0.9957 0.9903 -0.9969 0.9986 -0.9826
CAA 0.9986 0.9983 0.9690 -0.9988 0.8442 -0.9934
Tabela 4. Correlação de Spearman para a interação entre variáveis morfológicas (dAA = diâmetro árvore apical; dAB = diâmetro árvore basilar; CAA = comprimento árvore apical; CAB = comprimento árvore basilar) e os parâmetros do padrão de disparo (LGPD = latência para a geração do primeiro disparo; PIID = promédio do intervalo inter-disparo; CPPA = comprimento promédio do potencial de ação; APD = amplitude promédio da despolarização; APR= Amplitude promédio da repolarização e Taxa de disparos)
Tabela 5. Correlações de Pearson e SpearmanD para a interação entre o comprimento promédio do potencial de ação (CPPA)D e a amplitude promédio da despolarização (APD).
Já tendo estabelecidas tais correlaçõesD uma análise detalhada das consequências que exercem cada uma das variações feitas na morfologia dendríticaD permite observar alguns padrões interessantes:
4.2
R edução c omprimento á rvore a pical
De todas as variáveis morfológicas avaliadasD a redução do comprimento da árvore apical foi a variável morfológica cuja modificaçãoD teve o efeito mais significativo sobre a maioria dos parâmetros do padrão de disparo. Por exemploD uma redução de 10% no comprimento desta árvoreD levou à diminuição de mais de 8% na latência para a geração do primeiro disparo (Tabela 6D Figuras 5)D e de quase 5% no intervalo
inter-LGPD PIID CPPA APD APR Taxa Disparos
dAA 0.9854 1.0000 1.0000 -1.0000 1.0000 -0.9889
dAV 1.0000 1.0000 0.9997 -1.0000 1.0000 -0.9840
CAA 1.0000 1.0000 0.9987 -1.0000 0.7766 -0.9935
CAV 0.9793 1.0000 0.9987 -1.0000 0.8636 -0.9415
Correlação (Spearman) Variáveis Morfológicas e Parâmetros Padrão de Disparo
Correlação CPPA e APD Pearson Spearman
CAA -0.9714 -0.9987
CAV -0.9798 -0.9987
dAA -0.9969 -1.0000
Tabela 6. Variação (em porcentagens) dos parâmetros do padrão de disparoD com base numa redução sistemática de até 50% nas diferentes variáveis morfológicas da arquitetura dendrítica: Diâmetro Árvore Apical (d AA)D Diâmetro Árvore Basilar (dAB)D Comprimento Árvore Apical (CAA)D Comprimento Árvore Basilar (CAB). Nas diferentes colunas encontra-se o valor porcentual da magnitude da variação (aumento o diminuição) para cada um dos parâmetros avaliados i.e. 1) LGDP: Latência geração primeiro disparo; 2) PIID: Promédio do intervalo inter-disparo; 3) PCPA: Promédio do comprimento dos potenciais de ação; 4) APD: Amplitude promédio da despolarização; 5) APR: Amplitude promédio da repolarização; 6) TPA: Taxa potenciais de ação.
PORCENTAGENS DO EFEITO DA VARIAÇÃO NA MORFOLOGIA SOVRE PÂRAMETROS DO PADRÃO DE DISPARO % Variação LGPD % Variação PIID % Variação PCPA % Variação APD % Variação APR % Variação TPA
dAA (10%) -6.29 -5.18 -4.90 6.16 0.94 6.25
dAA (20%) -10.86 -9.30 -8.34 11.51 1.82 9.38
dAA (30%) -14.29 -12.51 -11.48 16.24 2.70 12.50
dAA (40%) -15.43 -14.98 -13.81 20.52 3.53 15.63
dAA (50%) -15.43 -16.62 -15.71 24.45 4.30 18.75
dAV (10%) -5.71 -3.57 -2.72 3.40 0.43 3.12
dAV (20%) -10.29 -6.62 -4.96 6.34 0.85 6.25
dAV (30%) -14.29 -9.27 -6.81 8.94 1.25 9.38
dAV (40%) -17.71 -11.61 -8.24 11.28 1.61 12.50
dAV (50%) -20.57 -13.63 -9.63 13.37 1.94 15.63
CAA (10%) -8.57 -4.97 -0.88 2.25 -0.20 3.12
CAA (20%) -16.57 -9.16 -1.25 4.01 -0.23 9.38
CAA (30%) -23.43 -12.90 -1.49 5.55 0.15 15.63
CAA (40%) -30.29 -16.62 -1.96 7.35 0.76 18.75
CAA (50%) -36.57 -20.11 -3.15 9.10 1.98 25.00
CAV (10%) -0.57 -1.14 -0.28 0.88 -0.07 0.00
CAV (20%) -0.57 -2.52 -0.57 1.84 -0.03 3.13
CAV (30%) -1.71 -4.10 -0.85 2.91 0.12 3.13
CAV (40%) -2.86 -5.81 -1.34 4.04 0.54 6.25
Figura 4. Mudança da Latência para a geração do primeiro disparo (LGPD) para cada variação na morfologia. O eixo x mostra os distintos valores porcentuais (em decimal) para cada variável morfológica (CAA: Comprimento Árvore ApicalD CAB: Comprimento Árvore BasilarD dAA: Diâmetro Árvore Apical e dAB: Diâmetro Árvore Basilar)D sendo 1 o 100% do tamanho originalD e 0.5 o 50% do tamanho original. O eixo Y na esquerdaD representa a LGDP (em milissegundos). O eixo Y na direitaD representa a porcentagem de variação da LGPDD em relação ao valor obtido para a morfologia original.
Figura 6. Mudança no promédio do comprimento do potencial de ação (PCPA) para cada variação na morfologia. O eixo x mostra os distintos valores porcentuais (em decimal) para cada variável morfológica (CAA: Comprimento Árvore ApicalD CAB: Comprimento Árvore BasilarD dAA: Diâmetro Árvore Apical e dAB: Diâmetro Árvore Basilar)D sendo 1 o 100% do tamanho originalD e 0.5 o 50% do tamanho original. O eixo Y na esquerdaD representa o PCPA (em milissegundos). O eixo Y na direitaD representa a porcentagem de variação do PCPAD em relação ao valor obtido para a morfologia original.
Figura 8. Mudança na amplitude promédio da repolarização (APR) para cada variação na morfologia. O eixo x mostra os distintos valores porcentuais (em decimal) para cada variável morfológica (CAA: Comprimento Árvore ApicalD CAB: Comprimento Árvore BasilarD dAA: Diâmetro Árvore Apical e dAB: Diâmetro Árvore Basilar)D sendo 1 o 100% do tamanho originalD e 0.5 o 50% do tamanho original. O eixo Y na esquerdaD representa o APR (em milivolts). O eixo Y na direitaD representa a porcentagem de variação do APRD em relação ao valor obtido para a morfologia original.
Um fato interessante é que alterações no comprimento da árvore apicalD não têm um efeito tão forte sobre os parâmetros do padrão de disparo (em relação a uma diminuição dos diâmetros da árvore apical ou basilar)D que têm a ver mais com a forma do potencial de ação (i.e. comprimento do disparoD amplitude da despolarização e da repolarização)D apresentandoD por exemploD uma redução só de 3.1% no comprimento do potencial de ação (Tabela 6D Figura 6)D junto com um aumento de 9.1% e 1.98 na amplitude da despolarização e repolarizaçãoD respetivamente. PortantoD no caso desta variávelD as consequências sobre o aumento de um parâmetro global tal como a taxa de disparoD relacionam-se principalmente com uma diminuição da latência e do intervalo entre disparosD contrário ao que acontece quando são modificados os diâmetros.
4.3 Redução diâmetro árvore apical
sejam menoresD ao ser comparados com o efeito da modificação do comprimento (Tabela 6D Figura 9). O mais interessanteD é que ao analisar os parâmetros do padrão de disparo cuja modificação se reflete diretamente na taxa (i.e. latência geração primeiro disparoD promédio do intervalo inter-disparoD e comprimento do potencial de ação)D é possível observar que não existe uma diferença muito alta no efeito das mudanças do diâmetro e do comprimento da árvore apicalD sobre o intervalo inter-disparo (Tabela 6)D mas sim sobre a latência do primeiro potencial e o promédio do comprimento do disparo. Por exemploD a redução do 50% nestas duas variáveis morfológicasD leva a uma diferença de 4% na diminuição do promédio do intervalo inter-disparo (-16.62% para o diâmetro e -20.11% para o comprimento)D quando uma redução morfológica da mesma magnitude leva a uma diferença de mais do dobro na diminuição da latência (-15.43% para o diâmetro em comparação com o -36.57 para o comprimento)D e cinco vezes maior no caso do promédio do comprimento do disparo (-3.15% para o comprimentoD e -15.71% para o diâmetro). Isso é bem importanteD pois quer dizer que a taxa de disparos depende da interação dos vários parâmetros descritosD e que o efeito da mudança da morfologia se reflete diferencialmente sobre cada um desses parâmetros.
Segundo o nosso modeloD as diferentes morfologias neurais poderiam ter consequências diretamente sobre o processamento e codificação de informaçãoD pois sabe-se queD por exemploD a codificação neural não depende só da taxa de disparosD mas do tempo no qual eles são transmitidos e processados (“timing”)D o qual estaria
redução de 40% no diâmetro da árvore apicalD ou de 30% no comprimento desta mesma árvoreD exercem um mesmo efeito enquanto a um aumento de 15.63% na taxa de disparo (Tabela 6D Figura 9)D no entantoD esse mesmo efeito está sustentado por diferentes variações ao nível da latência (-23.43% para o comprimentoD -15.43% para o diâmetro) (Tabela 6D Figura 4)D do comprimento do disparo (-1.49% para o comprimentoD -13.81% para o diâmetro) (Tabela 6D Figura 6)D e do promédio do intervalo inter-disparo (-12.90% para o comprimentoD - 14.98% para o diâmetro) (Tabela 6D Figura 5).
4.4 Redução diâmetro árvore basilar
Ao fazer uma análise da correlação entre o comprimento do potencial e a amplitude da despolarizaçãoD inesperadamente foi determinada uma relação inversa entre os dois parâmetros (Tabela 5)D isto éD uma amplitude maior se correlaciona com um comprimento menor. Isso é bem interessanteD pois quer dizer que mesmo tendo uma variação maior na voltagem de membrana durante a despolarizaçãoD isso não levaria a uma maior quantidade de tempo necessário para o disparo ser gerado (i.e. despolarização e repolarização da voltagem de membrana)D o que se refletiria num potencial de ação com um comprimento maior. Mais interessante aindaD é o fato de que variações na morfologiaD sobretudo no que tem a ver com os diâmetrosD estariam mudando o efeito sobre a voltagem de membranaD dos processos biofísicos relacionados principalmente com as dinâmicas dos canais iônicosD mesmo quando os parâmetros que têm a ver com esses processos se mantem constantes.
4.5
Redução c omprimento á rvore b asilar
4.6 Interação entre variáveis morfológicas e efeito combinado sobre padrão de disparo
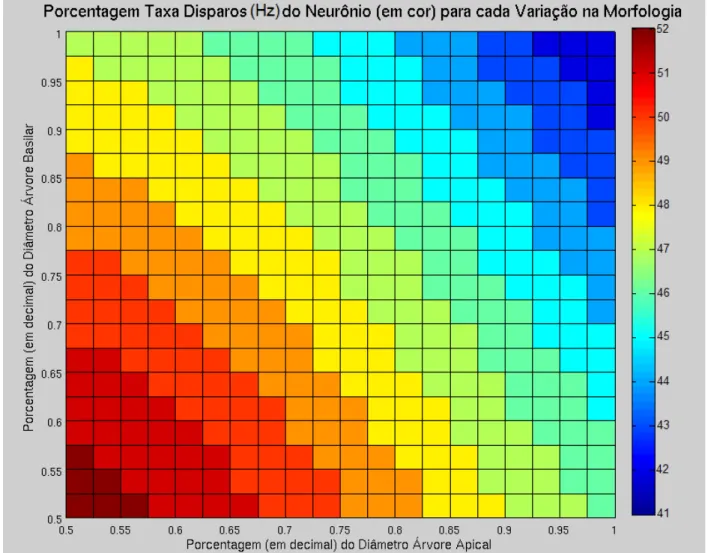
Figura 13. Efeito combinado da diminuição até um 50% (0.5) do diâmetro e comprimento da árvore basilarD sobre a taxa de disparo (codificado em cor) do neurônio piramidal.
variação porcentual (aumento ou redução) do (1) LGDP: Latência para geração do primeiro disparoD (2) PIID: promédio intervalo inter-disparoD (3) PCPA: Promédio comprimento do potencial de açãoD (4) APD: Amplitude promédio da despolarizaçãoD (5) APR: Amplitude promédio da repolarizaçãoD (6) TPA: Taxa de disparos.
A partir dos resultados obtidosD também é possível indagar na interação entre as distintas variáveis morfológicasD e o seu efeito combinado sobre parâmetros do padrão de disparoD tais como a taxa de disparo.
A primeira interação avaliadaD foi a redução simultânea dos diâmetros da árvore apical e basilar. Esse resultados são mostrados na Figura 10D na qual é possível observar alguns padrões interessantes. Por exemploD a primeira linha horizontal (de acima para abaixo)D representa a mudança na voltagem (codificada em cor) quando o diâmetro da árvore apical é mudada sistematicamente um 2.5% até atingir um 50% do tamanho original (de direita para esquerda)D ao mesmo tempo que se mantém constante o diâmetro da árvore basilar num 100% do seu tamanho original. Daí descendo linha por linhaD é possível observar como há um aumento do efeito sobre a taxa de disparoD quando a diminuição do diâmetro da árvore apicalD interage com a redução do diâmetro da árvore basilar.
Descendo por esta diagonalD é possível observar o efeito combinado da redução nestas duas variáveis sobre a taxa de disparoD levando a um aumento maior na taxa do disparo que o efeito individual da diminuição em só um dos diâmetros. Isso acontece em todas as interações morfológicas avaliadas (Figura 10D 12D 13)D exceto ao avaliar o efeito combinado de uma redução simultânea no diâmetro e no comprimento da árvore basilar (Figura 13)D na qual o efeito destas duas variáveis sobre o padrão de disparoD pode ser explicado principalmente pelo efeito da redução só do diâmetro (primeira linha horizontal de acima para abaixo)D em comparação há uma diminuição só do comprimento (primeira linha vertical de direita a esquerda). De fatoD o efeito observado da combinação na redução simultânea destas duas variáveis num 50%D é basicamente o mesmo efeito de uma redução só do diâmetro na mesma magnitudeD mesmo quando existe um efeito significativo sobre a taxa de disparo quando se reduz só comprimento.
Um outro fato interessante é que mesmo quando há um aumento significativo sobre a taxa de disparoD pelo efeito combinado da redução em duas variáveis (Figura 10D 12D 13)D este aumento é muito menor que a soma do efeito pela modificação de cada variável independentemente (Figuras 10D 11D 12D 13)D o qual reafirma a não linearidade não só do efeito da morfologia sobre o padrão de disparoD mas do efeito combinado e da interação entre variáveis morfológicas.
redução no diâmetro e no comprimento da árvore dendrítica apicalD teve um efeito muito maior sobre a taxa de disparo (Figura 12 e 15)D em comparação ao efeito combinado da redução do diâmetro e comprimento da árvore basilar (Figura 13 e 15).
Esse mesmo efeito diferencial da árvore apical e basilar sobre o aumento da taxa de disparoD foi observado em todos os parâmetros avaliados (Figura 14). De fatoD o efeito da combinatória da redução do diâmetro e comprimento da árvore apicalD foi maior em todos os parâmetros em comparação com o efeito da mesma combinatória mas na árvore basilar (Figura 14).
A magnitude do efeito observado sobre o padrão de disparo das diferentes combinatórias foiD de maior a menor em: (1) Redução da latência para a geração do primeiro disparo; (2) Aumento da taxa de disparo; (3) Diminuição no promédio do intervalo inter-disparo; (4) Aumento na amplitude da despolarização; (5) Redução do comprimento do potencial de ação; e (6) Aumento na amplitude da repolarização.
mecanismos através dos quais as variáveis morfológicas interagem entre elasD e exercem o seu efeito sobre as diferentes características do padrão de disparoD possuem uma natureza altamente não linear.
6 CONCLUSÕES
O nosso modelo neural predizD que mesmo mantendo constantes todos os mecanismos biofísicos envolvidos nas dinâmicas dos canais iônicosD pequenas mudanças na morfologia dendríticaD são suficientes para exercer efeitos significativos sobre a geração de potenciais de açãoD e sobre o padrão no qual eles são gerados.
Ainda maisD segundo os nossos resultadosD é possível sugerir que alterações na morfologia dendrítica do mesmo tipo às observadas na epilepsiaD se refletiriam num aumento importante do comportamento elétrico dos neurônios piramidaisD e na sua geração de potenciais de ação. Isso é muito interessanteD pois permite indagar na possibilidade de algum tipo de relação causal nas correlações encontradas in vivo e in vitro, entre modificações na morfologia dendrítica e a epilepsia.
7 REFERÊNCIAS
AITKEND J.T. Observations on the larger anterior horn cells in the lumbar region of the cat’s spinal cord. J. Anat.D v. 89D p. 571D 1955.
BASTIAND J.; NGUYENKIMD J. Dendritic Modulation of burst-like firing in sensory neurons. J. Neurophysiol., v. 85D p. 10-22D 2000.
BERNARD et al. Acquired dendritic channelopathy in temporal lobe epilepsy. Science v. 305D p. 532-535D 2004.
BROWND K.M.; GILLETTED T.A.; ASCOLID G.A. Quantifying neuronal size: Summing up trees and splitting the branch difference. Seminars in Cell and Developmental Viology, v. 19D p. 485-493D 2008.
CHKLOVSKIID D.B. Synaptic connectivity and neuronal morphology: two sides of the same coin. Neuron, v. 43D p. 609–17D 2004.
COLLINGD S.B. et al. Dendritic shrinkage and dye-coupling between rat hippocampal CA1 pyramidal cells in the tetanus toxin model of epilepsy. Vrain Research, v. 741D p. 38-43D 1996.
ENGELD J.Jr.; PEDLEYD T.A. Epilepsy: a comprehensive textbookD 2nd Ed. Lippincott Williams & Wilkins. Philadelphia. 2008.
DOIROND B.; LAINGD C.; LONGTIND A.; MALERD L. Ghostbursting: a novel neuronal burst mechanism. J. Comput. Neurosci. v. 12D p. 5–25D 2002.
DONOHUED D.E.; ASCOLID G.A. A comparative computer simulation of dendritic morphology. Plos, Computational Viology, v. 4(6): e1000089. Doi: 10.1371D 2008. FOXD C.A.; BARNARDD J.W. A quantitative study of the Purkinje cell dendritic branchlets and their relationship to afferent fibres. J. AnatD v.91D p. 299-313D 1957.
GLOBUSD A. Brain morphology as a function of presynaptic morphology and activity. In Developmental neurophysiology of sensory deprivation. Academic Press. New YorkD 1975.
GRAVESD T.D. Ion Channels and Epilepsy. Q.J. Med.D v. 99D p. 201-217D 2006.
HAUSERD W.A.; ANNEGERSD J.F.; KURLANDD L.T. Prevalence of epilepsy in RochesterD Minnesota: 1940–1980. Epilepsia v. 32D p. 429–445D 1991.
HODGKIND A.; HUXLEY A. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol.D v. 117D p. 500–544D 1952.
JIANGD M.D et al. Spine loss and other persistent alterations of hippocampal pyramidal cell dendrites in a model of early-onset epilepsy. The Journal of Neuroscience v. 18(20)D p. 8356-8368D 1998.
KIMD H.G.; CONNORSD B.W. Apical dendrites of the neocortex: Correlation between Sodium and Calcium-dependent Spiking and Pyramidal Cell Morphology. The Journal of NeuroscienceD v. 13 (12)D p; 5301-5311D 1993.
KRICHMARD J. L. et al. Effects of dendritic morphology on CA3 pyramidal cell electrophysiology: a simulation study. Vrain ResearchD v. 941D p. 11-28D 2002.
KURZ et al. A cellular mechanism for dendritic spine loss in the pilocarmine model of status epilepticus. EpilepsiaD v. 49(10)D pD 1696-1710D 2008.
LERCHED H.; JURKAT-ROTTD K.; LEHMANN-HORND F. Ion Channels and Epilepsy. American Journal of Medical GeneticsD v. 106D p. 146-159D 2001.
LONDOND M.; HÄUSSERD M. Dendritic computation. Annual Review Neuroscience v. 28D p. 503-532D 2005.
MAGEED J.C. Dendritic integration of excitatory synaptic input. Nature ReviewsD v. 1D p. 181-190D 2000.
MAINEND Z.F.D et al. A model of spike initiation in neocortical pyramidal neurons. NeuronD v. 15D p. 1427-1439D 1995.
MAINEND Z.F.; SEJNOWSKID T.J. Influence of dendritic structure on firing pattern in model neocortical neurons. NatureD v. 382D p. 363-366D 1996.
MAINEND Z.F.; SEJNOWSKID T.J. Modeling active dendritic processes in pyramidal neurons. In C. Koch e L. Segev (Eds.)D Methods in Neuronal ModelingD Cambridge: MIT PressD 1998.
MASOND A.; LARKMAND A. Correlations between morphology and electrophysiology of pyramidal neurons in slices of rat visual cortex. II. Electrophysiology. The Journal of NeuroscienceD v. 10(5)D p. 1415-1428. 1990.
McCORMICKD D.A. et al. Comparative electrophysiology of pyramidal and sparsely spiny stellate neurons in the neocortex. J. NeurophysiologyD v. 54(4)D p. 782-806D 1985.
MEHTAD M.R. Cooperative LTP can map memory sequences on dendritic branches. Trends in NeurosciencesD v. 27(2)D p. 69-71D 2004.
35D p. 1077-1079D 2007.
MULTANID P. et al. Neocortical dendritic pathology in human partial epilepsy: a quantitative Golgi study. Epilepsia v. 35(4)D p. 728-736D 1994.
RALLD W. The theoretical foundations of dendritic functionD CambridgeD Massachusetts. The MIT PressD 1995.
SEGALD M. Dendritic spines and long-term plasticity. Nat Rev Neurosci.D v. 6(4)D pD 277-284D 2005.
SHAEFERD A.T. et al. Coincidence detection in pyramidal neurons in tuned by their dendritic branching pattern. J. NeurophysiologyD v. 89D p. 3143-3154D 2003.
SCHEIBELD M.E.; CRANDALLD P.H.; SCHEIBELD A.B. The hippocampal-dentate complex in temporal lobe epilepsy. A Golgi study. EpilepsiaD v. 15(1)D p. 55-80D 1974.
SCHWARTZKROIND P. Encyclopedia of basic epilepsy. Elsevier. USAD 2009.
SHADÉD J.P.; BAXTERD C.F. Changes during growth in the volume and surface area of cortical neurons in the rabbit. Exptl. Neurol.D v. 2D p. 158-178D 1960.
SHOLLD D.A. The surface area of cortical neurons. J. Anat.D v. 89D p. 571-572D 1995. SINGLED S.; HAGGD J.; BORSTD A. Dendritic computation of direction selectivity and gain control in visual interneurons. The journal of NeuroscienceD v. 17(16)D p. 6023-6030D 1997.
SINGLED S.; BORSTD A. Dendritic integration and its role in computing image velocity. ScienceD v. 281D p. 1848-1850D 1998.
STEINLEIND O.K. Genetic mechanisms that underlie epilepsy. Nature ReviewsD v. 5D p. 400-408D 2004.
STUARTD G.D SPRUSTOND N.D HÄUSSERD M. Dendrites (Second Edition). Oxford University Press. New YorkD 2008.
SWANND J.W. The spine loss paradox: Clues to mechanisms and meaning. Epilepsy CurrentsD v. 8(6)D p. 168-169D 2008.
SWANND J.W. et al. Spine loss and other dendritic abnormalities in epilepsy. HIPPOCAMPUSD v. 10D p. 617-625D 2000.
VAN ELBURGD R.A.J.; VAN OOYEND A. Impact of dendritic size and dendritic topology on burst firing in pyramidal cells. PLoS Comput Viol. v. 6(5): e1000781 doi: 10.1371/journal.pcbi.1000781D 2010.
VETTERD P.D ROTHD A.D HÄUSSERD M. Propagation of action potentials in dendrites depends on dendritic morphology. J. NeurophysiologyD v. 85D p. 926-937D 2001.
YOUNGD J.Z. Anatomical considerations. EEG Clin. Neurophysiol. v. 10D p. 9-11D 1958. YU F.H. et al. Reduced Sodium Current in GABAergic Interneurons in a Mouse Model of Severe Myoclonic Epilepsy in Infancy. Nature NeuroscienceD p. 1142-1149D 2006. YUSTED R.D TANKD D.W. Dendritic integration in mammalian neuronsD a century after Cajal. NeuronD v. 16D p. 701-716D 1996.
WASHINGTOND S.D.; ASCOLID G.A.; KRICHMARD J.L. A statistical analysis of dendritic morphology’s on neuron electrophysiology of CA3 pyramidal cells. NeurocomputingD v. 32-33D p. 261-269D 2000.
WESTRUMD L.E.; WHITED L.E.; WARDD A.A. Morphology of the experimental epileptic focus. J NeurosurgD v. 21D p. 1033-46D 1964.
ZENG et al. Kainate seizures cause acute dendritic injury and actin depolymerization in vivo. The Journal of NeuroscienceD v. 27(43)D p. 11604-11613D 2007.