PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS (ÁREA: MICROBIOLOGIA APLICADA)
IFELOJU DAYO-OWOYEMI
TAXONOMIC ASSESSEMENT AND BIOTECHNOLOGICAL POTENTIAL OF YEASTS HOLD AT THE UNESP - CENTRAL FOR MICROBIAL RESOURCES
TAXONOMIC ASSESSEMENT AND BIOTECHNOLOGICAL POTENTIAL OF YEASTS HOLD AT THE UNESP –CENTRAL FOR MICROBIAL RESOURCES
IFELOJU DAYO-OWOYEMI
Thesis presented to the Institute of Biosciences, Universidade Estadual Paulista ´´Julio de Mesquita Filho``- Rio Claro, in fulfilment of requirements for the award of Doctor of Philosophy in Biological Sciences (Applied Microbiology)
Supervisor: Prof. Dr. Fernando Carlos Pagnocca Co-supervisor: Prof. Dr. André Rodrigues
DEDICATION
´If I have seen further it is by standing on the shoulders of
giants``
Acknowledgement
Completing my PhD was a long and challenging task. Many people supported and encouraged me in so many different ways during the process; it is therefore my pleasure to thank those who helped to see my dream come true.
First, I thank the Almighty God, the true source of wisdom and knowledge, for his immense love and infinite mercy towards me. ´´A man has gotten nothing except he be given from above``; I appreciate the rare door of opportunity He opened for me and also for the strength and inspiration given to me for the successful completion of this work.
I thank my supervisor Prof. Dr. Fernando Carlos Pagnocca for the wonderful opportunity he gave me in his laboratory. I am grateful to him for believing in me. I appreciate the fatherly role he also played during my stay and studies. I am also indebted to Dr Andre Rodrigues, who apart from being my co-supervisor also played the role of a colleague and brother. ``If I have seen further it is by standing on the shoulders of giants``, together, I thank him and my supervisor very much; for investing their time, resources and wealth of knowledge in me. Most importantly, I appreciate them for contributing to securing a promising future for me.
I thank the Brazilian government through the National Council of Technological and Scientific Development (CNPq) as well as the Third World Academy of Science (TWAS) for the PhD fellowship awarded me and for the sponsorship of my education. I also
aknowledge Universidade Estadual Paulista “Julio de Mesquita Filho” for the quality
education I received and for all the material and infrastructural resources that were made available during the course of my work. I wish to equally use this opportunity to thank the good people of Brazil whose hard earned resources were used to give me an opportunity of quality education.
Several other lecturers and non lectures as well as colleagues also contributed directly or indirectly to the success of my career and their contributions are well appreciated. They include Dra. Sandra Mara Martins Franchetti, Dr. Lara Durães Sette, Prof. Dr. Vanderlei Martins, Prof. Dr. Jonas Conteiro, Necis Lima, Rosemary D. Oliveira S. Cardoso, Josiele Fernanda Magri, Dr. Mauricio Bacci Jr., Joaquim Martim Jr., Mileni Ferro, Cynara, Lusiana, Alexandre, Alex and Paulo, Dr. Bolatito Boboye, Dr. Victor Oyetayo, Engineer and Mrs Taiwo Mogaji, Dr Mathias C. Ahii, Dr. Adebayo Adeyemo and Dr Olubunmi Adebayo.
This note of acknowledgement would be incomplete if I fail to appreciate Dra. Derlene Attili de Angelis and her wonderful family. I am immensely grateful and specially thank them for being a family away from home. I appreciate Dra. Dejanira de Franceschi de Angelis, my Brazilian grandmother, for her constant love and concern for me.
A special appreciation goes to my husband Olusegun Folaring Jonah. I would not have successfully completed this work without his support and encouragement. I greatly appreciate his unwavering love, trust, patience and understanding. I am grateful to him for standing solidly by me throughout the course of this work.
RESUMO
Atualmente, existe um crescente interesse em explorar diversos habitats, a fim de revelar a biodiversidade microbiana, incluindo as leveduras. Tal diversidade ainda não acessada guarda a descoberta de novas espécies para ciência, provavelmente muitas das quais com potencial para aproveitamento em processos biotecnológicos. Com o objetivo de explorar e conservar a diversidade de fungos, o Central de Recursos Microbianos da UNESP (CRM –
UNESP) mantém em seu acervo várias estirpes de leveduras isoladas de ecossistemas diversos, sendo alguns deles pouco explorados. No início deste trabalho sabíamos que muitas das leveduras depositadas no acervo do CRM – UNESP não estavam totalmente caracterizadas tanto em nível taxonômico, quanto em relação ao potencial biotecnológico que poderiam apresentar. Portanto, o presente estudo foi desenhado para caracterizar e identificar taxonomicamente leveduras depositadas no CRM – UNESP, bem como selecionar estirpes que produzem enzimas extracelulares degradadoras de polissacarídeos como amilase, celulase, xilanase, pectinase e ligninase. Usando uma abordagem polifásica, um total de 340 isolados de leveduras foi identificado, sendo que 71,2% compreendem 43 taxa de ascomicetos e os restantes 28,8% foram classificados em 27 taxa de basidiomicetos. O estudo também levou à descoberta de 8 prováveis novas espécies. Baseado nesta constatação, a classificação taxonômica e análise filogenética foi realizada para duas espécies anamórficas de ascomicetos e uma espécie teleomórfica de basidiomiceto. A descrição destas três espécies é apresentada neste estudo. Os resultados demonstraram que Wickerhamiella kiyanii FB1-1DASPT e W. pindamonhangabaensis H10YT pertencem à clade Wickerhamiella da ordem Saccharomycetales (Ascomycota: Saccharomycetes), enquanto que a espécie Bulleromyces texanaensis ATT 064T pertence à clade Bulleromyces / Papiliotrema / Auriculibuller da ordem Tremellales (Basidiomycota: Agaricomycotina). Num outro estudo, demonstramos a variabilidade intraespecífica em onze (11) isolados de Hannaella kunmingensis (incluindo a type strain CBS 8960T). Essas onze estirpes foram obtidas de substratos e locais diferentes e analisamos sua variabilidade fisiológica e genética. Ainda, usando uma combinação de análise filogenética e de rede parcimônia, demonstramos o grau da divergência genética (região espaçadora intergênica (ITS) e o gene do citocromo b) dentro desta espécie. Os nossos resultados revelaram variabilidade elevada
desta estirpe. Este trabalho mostrou características que não foram previamente descritos em H. kunmingensis, e assim pudemos contribuir para a emenda referente à descrição desta espécie, procedimento necessário para acomodar as novas descobertas. Além disso, a partir da triagem das enzimas extracelulares de 312 estirpes, foi detectada a atividade de amilase em 28 estirpes (8,95% do total), celulase em 64 estirpes (20,51%), xilanase em 87 estirpes (27,88%), poligalacturonase em 45 estirpes (14,42%), pectina liase em 59 estirpes (18,91%) e ligninolítica em 2 estirpes (0,64%). As enzimas celulase, amilase, xilanase foram as mais encontradas entre as leveduras basidiomicetas; enquanto que as leveduras ascomicetas foram maiores produtores de pectinases. A determinação da produção de endoxilanase e β -xilosidase de 73 estirpes degradadoras de xilana levou à descoberta de três estirpes que demonstraram elevada produção de amilase, celulase, xilanase e pectinase na presença de bagaço de cana como substrato, indicando que elas são boas candidatas para as pesquisas envolvendo a produção de enzimas úteis na conversão de biomassa vegetal em bioetanol. No geral, este estudo revelou que o CRM – UNESP abriga um acervo de leveduras diversas, com capacidade de produzir várias enzimas industrialmente úteis. Estas leveduras poderiam ser aproveitadas para futuras aplicações biotecnológicas. Além disso, o acervo do CRM –
UNESP também provou ser uma fonte de conservação de várias espécies novas para ciência, o que reflete a importância desse tipo de conservação ex-situ para o estudo da biodiversidade microbiana.
ABSTRACT
In recent time, there has been an increasing interest in exploring diverse ecological habitats in order to reveal the yeast biodiversity. The increased awareness in the biotechnological potentials of yeasts has also spurred attempts to search for new species with novel biotechnological capabilities. Aiming to explore and conserve the fungal diversity from various ecosystems, the UNESP – Central for Microbial Resources (UNESP – CMR) harbors various strains of ecologically diverse yeasts isolates, some of which were yet to be identified. Therefore, this study was designed to identify and characterize some yeasts from the UNESP – MRC and to select strains possessing extracellular plant polysaccharide degrading enzymes namely amylase, cellulase, xylanase, pectinase and ligninase. Using a polyphasic approach, a total of 340 strains were identified. Taxonomic classification grouped 71.2% of these isolates into 43 ascomycetous taxa while the remaining 28.8% were classified in 27 basidiomycetous taxa. The study also led to the discovery of 8 putative new species. As a result, we classified two anamorphic species in the Ascomycota and one teleomorphic species in the Basidiomycota. In this study we provide the description of both species. Our results demonstrated that the two ascomycetous species proposed as Wickerhamiella kiyanii FB1-1DASPT and W. pindamonhangabaensis H10YT belong to the Wickerhamiella clade of the Saccharomycetales (Saccharomycetes) while the basidiomycetous species proposed as Bulleromyces texanaensis ATT064T belong to the Bulleromyces / Papiliotrema / Auriculibuller clade of the Tremellales (Agaricomycotina). In order to show the significance of intraspecific diversity in yeasts, in one of our studies, we subjected 11 strains, (including the type strain CBS 8960T) of Hannaella kunmingensis, obtained from different substrates and geographic locations, to detailed physiological and genetic characterization. Using a combination of phylogenetic and parsimony network analysis, we demonstrated the extent of genetic (internal transcribed spacer region (ITS), D1/D2 domains of the large subunit rDNA (LSU), and cytochrome b gene) divergence within this species. Our findings revealed the high variability of morphological and biochemical characteristic as well as the existence of 3 genetic haplotypes in H. kunmingensis. One of the strains (CBS 8356T) exhibited a 27.3 % divergence from the other strains in the cytochrome b gene; hence, we concluded the possibility of
speciation of this strain. This work led to the discovery of additional strains and characteristics not previously reported in H. kunmingensis, therefore, the emendation of H. Kunmingensis was done to
accommodate the new discoveries. Furthermore, from the screening of 312 yeast strains for secreting
(27.88%), polygalacturonase activity in 45 strains (14.42%), pectin lyase activity in 59 strains (18.91%) and lignolytic activity in 2 strains (0.64%). This study further revealed that amylase, cellulase and xylanase are the major enzymes found among the basidiomycetous yeasts while ascomycetous yeasts are producers of pectinases. Determination of extracellular
endoxylanase and β-xylosidase activities in culture supernatants of 73 xylanase positive strains led to the discovery of three strains which demonstrated high amylase (endo- and exomylase), cellulase, xylanase and pectinase activities in presence of sugar cane bagasse; therefore are good candidates for research involving production of enzymes useful in biomass conversion. Overall, this study revealed that UNESP – MRC possess metabolically diverse yeasts with ability to produce various industrially useful enzymes. Such strains could be harnessed for future biotechnological applications. In addition, the UNESP – MRC proved to harbors new species to science that are now preserved ex-situ of long-term maintenance.
TABLE OF CONTENTS
INTRODUCTION 19
Aims and objectives 21
STUDY OUTLINE 23
CHAPTER 1 25
BACKGROUND OF STUDIES AND LITERATURE REVIEW 1 BACKGROUND OF STUDIES AND LITERATURE REVIEW 26 1.1 UNESP – Central for Microbial Resource (UNESP-CMR) 26 1.2 Importance of culture collections to microbiology and biotechnology 30 1.3 Yeast: introduction and definition 32 1.4 Evolution of yeast identification methods 32 1.5 Yeast classification 36
1.6 Yeast ecology and diversity 42 1.7 Biotechnological importance of yeasts 47
1.8 Biodegradation of starch, lignocelluloses and pectin 47
REFERENCES 57
CHAPTER 2 69
TAXONOMIC STUDIES OF YEASTS HOLD AT UNESP – CENTRAL FOR MICROBIAL RESOURCES 2.1 Abstract 70
2.2 Introduction 71
2.3 Material and methods 72
2.3.1 Cultural Characterization 72
2.3.2 Molecular Identification 74 2.4 Results 76
2.5 Discussion 79
REFERENCES 85
SCREENING FOR AMYLOLYTIC, LIGNOCELLOLYTIC AND PECTINOLYTIC YEASTS
3.1 Abstract 89
3.2 Introduction 90
3.3 Materials and methods 91
3.3.1 Screening procedures for extracellular enzymatic activities 91
3.3.2 Amylase activity 93 3.3.3 Cellulases 93
3.3.4 Xylanases 93
3.3.5 Pectinases 94
3.3.6 Ligninase 94
3.3.7 Xylanase enzymes assays 94 3.3.8 Fermentation of sugarcane bagasse 96
3.3.9 Enzyme assays 96
3.3.10 Statistical analysis 98 3.4 Results 98
3.4.1 Screening for enzymatic activities 98
3.4.2 Xylanase (endoxylanase and β-xylosidase) assays 106
3.4.3 Extracellular enzyme production from sugar cane bagasse fermentation 110
3.5 Discussion 112
3.5.1 Enzymatic activity profile 112
3.5.2 Enzyme production from sugar cane baggase 113
REFERENCES 115
CHAPTER 4 118
WICKERHAMIELLA KIYANII SP. NOV. AND W. PINDAMONHANGABAENSIS SP. NOV., TWO ANAMORPHIC YEASTS ISOLATED FROM NATIVE PLANTS OF THE SOUTH EASTERN ATLANTIC RAINFOREST OF BRAZIL 4.1 Abstract 119
4.2 Introduction 120
4.3 Material and methods 121
4.4 Results and discussion 123
4.6 Description of Candida pindamonhangabaensis Pagnocca, Rosa,
Dayo-Owoyemi and Rodrigues sp. nov. 129
REFERENCES 131
CHAPTER 5 134
DESCRIPTION OF BULLEROMYCES TEXANAENSIS SP. NOV., ISOLATED FROM FUNGUS GARDEN OF THE LEAFCUTTER ANT ATTA TEXANA AND LEAVES OF BROMELIAD NEOREGELIA CRUENTA (BROMELIACEAE) 5.1 Abstract 135
5.2 Introduction 136
5.3 Materials and methods 137
5.3.1 Strain information 137
5.3.2. Morphological and phenotypic characterization 138 5.3.3 DNA extraction, Sequence and phylogenetic analyses 138 5.4 RESULTS AND DISCUSSION 139
5.4.1 DNA sequence and phylogenetic analysis 139
5.5 Description of Bulleromyces texanaensis Dayo-Owoyemi, Rodrigues, Garcia, Hagler and Pagnocca sp. nov. 144
5.5.1 Growth on YM broth 144
5.5.2 Growth on YM agar 144
5.5.4 Dalmau plate culture on corn meal agar 144 5.5.5 Formation of ballistoconidia 144
5.5.6 Sexual reproduction 144
5.6 PHENOTYPIC DESCRIPTION 147
5.7 Origin of the strains studied 147 5.8 Systematics and Ecology of Bulleromyces texanaensis 147
References 151 CHAPTER 6 154
ABBREVIATIONS °C – Degree centigrade
CBS – Centraalbureau voor Schimmelcultures
CMCase – Carboxymethyl cellulase
DBB – Diazonium blue B
DNA – Deoxyribo nucleic acid
g. – Gravitational force
g.L – Gram per Liter
gm – gram
ITS – internal transcribed spacers
LSU – Large subunit
MCase- Microcrystalline cellulase
M – Molar
min – minutes
mM – Milimolar
MSP-PCR – Microsatellite Primed Polymerase chain reaction.
NCBI – National Center for Biotechnology Information
SNA – Synthetic nutrient agar
TCS – Parsimony network analysis
UNESP – CMR – Universidade Estadual Paulista – Central for Microbial Resources
UV – Ultraviolet
LIST OF FIGURES
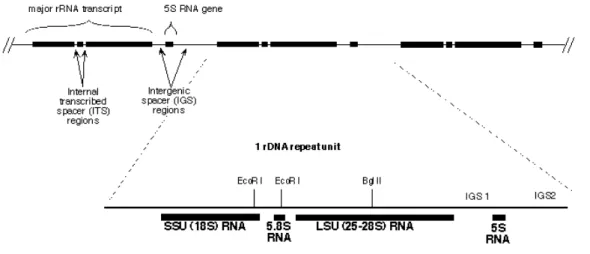
Figure 1.1 Map of fungi rRNA gene showing the internal transcribed spacer (ITS) region, intergenic spacer (IGS) regions,18S
small subunit (SSU) and 25-28S large subunit (LSU) 34
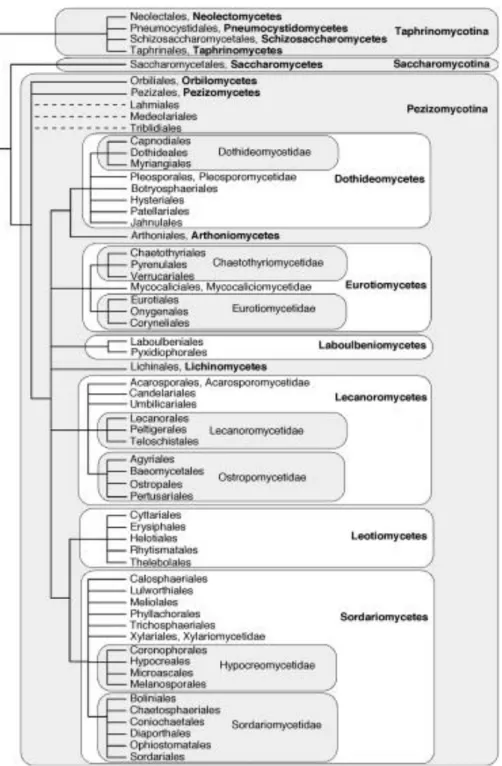
Figure 1.2 Phylogeny of the phylum Ascomyceota showing the
classification of Ascomycetous yeasts . 38
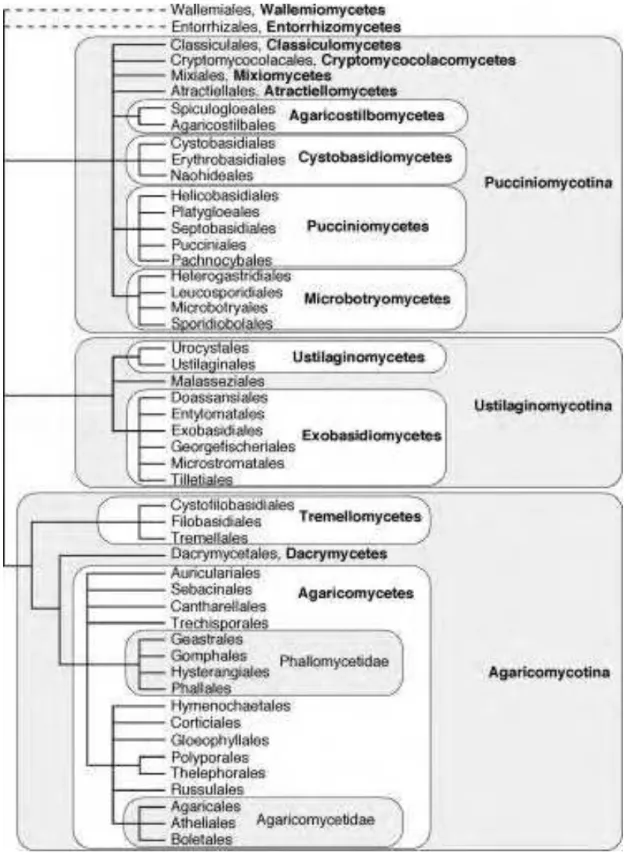
Figure 1.3 Phylogeny of the phylum Basidiomycota showing the classification
of Basidiomycetous yeasts 40
Figure 1.4 Plant cell wall structure. 48
Figure 1.5 Action of major cellulase enzymes 51
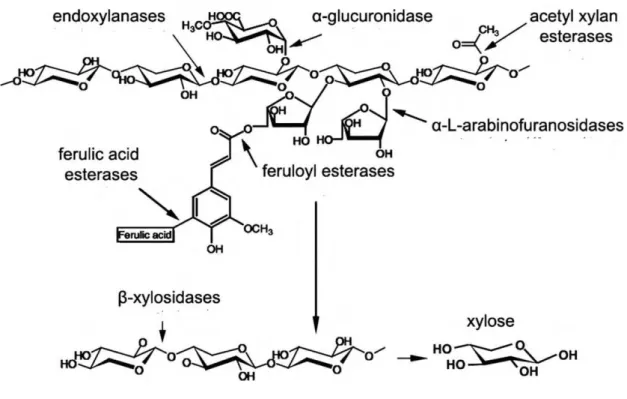
Figure 1.6 Action of major enzymes involved in the depolymerization of
Xylan 52
Figure 1.7 Action of major enzymes involved in the deconstruction of
pectin. 53
Figure 1.8 Schemeshowing the actions of lignin degrading enzymes 55 Figure 1.9 Schemeshowing the actions of starch degrading enzymes 56 Figure 2.1 Growth (assimilation) test on carbon compounds 77 Figure 2.2 PCR fingerprinting patterns of some identified strains 77
Figure 3.1 Degradation halos around yeast strains producing 100 amylase (A), cellulose (B), xylanase (C), polygalacturonase
(D), pectin lyase (E) and ligninase(F).
Figure 3.2 Enzymatic activity profiles of the ascomycetous and basidiomycetous 103
yeasts screened
Figure 3.3 Comparisons ofenzyme activity profiles of ascomycetous
and basidiomycetous yeasts respectively 104
Figure 3.4 Extracellular enzyme production from sugar cane bagasse fermentation by Aureobasidium pullulans strain CG5-5BY,
Aureobasidium pullulans strain PBM1and Pseudozyma
hubeiensis strain MP2-2CB 111
(Saccharomycetes, Saccharomycetales) determined from Neighbor-joining analysis of sequences from LSU rRNA gene.
Bootstrap values are from 1000 replicates. T = type species. 131 Figure 4.2 Candida kiyanii (A) and Candida pindamonhangabaensis
(B and C). Phase contrast micrograph showing budding cells (A and B) with pseudomycelium after 3 days at 25 °C on YM
agar (A) and on corn meal agar 25 °C (C). 126
Figure 4.3 Lipase activity test of Cpindamonhangabaensis
(upper colonies) and C. kiyanii(lower colonies). 128
Figure 5.1 Evolutionary tree showing the relationships of Bulleromyces texanaensis and related species based on combined LSU and
ITS sequences. 141
Figure 5.2 Phylogenetic relationships of strain Bulleromyces texanaensis and other closely related species based on
LSU D1/D2 rRNA gene sequences. 142
Figure 5.3 Phylogenetic relationships of strain Bulleromyces texanaensis and other closely related species based on
ITS sequences. 143
LIST OF TABLES
Table 1.1 Some major international microbial culture collections, their
acronyms and type of culture holding 27
Table 1.2 Quality control procedures recommended for microorganisms
(OECD, 2007) 29
Table 1.3 Industrial applications of some enzymes 49 Table 3.1 Number and origin of yeasts and dimorphic fungi profiled for
enzymatic activity 92
Table 3.2 Ascomycetous yeasts screened for amylolytic and lignocellulolytic
and pectinolytic activities 100
Table 3.3 Basidiomycetous yeasts screened for amylolytic and
lignocellulolytic and pectinolytic activities 102
Table 3.4 Xylanase activities in extracellular and cell wall associated cell
free supernatant of selected strains 107
Table 3.5 Enzyme yield per gram of substrate 112
Table 4.1 Extent of D1/D2 LSU rDNA and ITS sequence divergences of
C. kiyanii and close relatives pairwise some based on alignment. 125
Table 4.2 Physiological characteristics differentiating C. kiyanii from closely
related strains 125
Table 5.1 Phenotypic characteristics of strain Bulleromyces texanaensis
LIST OF APPENDIX
Appendix 1 Identities of yeasts and dimorphic fungi maintained at
UNESP –CMR 166
Appendix 2 Result of extracellular enzyme screening with some yeasts
in the UNESP - Central for Microbial Resources 179
Appendix 3 Statisitical (one-way anova) analysis of reducing sugars (RS) produced from sugar cane bagasse fermentation by
Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium pullulans strain PBM 1 and Aureobasidium pullulans strain
CG5-5BY 192
Appendix 4 Statisitical (one way anova) analysis of activitiy of
endoamylase produced through sugarcane baggase fermentation by Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium pullulans strain PBM 1and Aureobasidium pullulans strain
CG5-5BY (using 0.5% starch) 194
Appendix 5 - Statisitical (one way anova) analysis of activitiy of exoamylase produced through sugarcane baggase fermentation by
Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium pullulans strain PBM 1 and Aureobasidium pullulans strain
CG5-5BY (using 1.0% starch) 196
Appendix 6 Statisitical (one way anova) analysis of activitiy of pectinase produced through sugarcane baggase fermentation by
Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium pullulans strain PBM 1and Aureobasidium pullulans strain
CG5-5BY 198
Appendix 7 Statisitical (one way anova) analysis of activitiy of xylanase produced through sugarcane baggase fermentation by
Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium pullulans strain PBM1and Aureobasidium pullulans strain
CG5-5BY 200
Appendix 8 Statisitical (one way anova) analysis of activitiy of carboxymethyl cellulase (CMCase) produced through sugarcane baggase fermentation by Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium
pullulans strain PBM 1 and Aureobasidium pullulans strain CG5-5BY 202 Appendix 9 Statisitical (one way anova) analysis of activitiy of microcrystaline
cellulase (MCase) produced through sugarcane baggase fermentation by Pseudozyma hubeiensis strain MP2-2CB, Aureobasidium pullulans
INTRODUCTION
Since the last decade, the destruction of the natural ecosystem by human activities and changing global climatic conditions has raised concerns to microbiologists and ecologist as to the endangering of microorganisms. Such events are leading to gradual microbial species extinction, hence reduction of microbial diversity. According to Hibbett et al. (2011) more than 100,000 fungal species have been described to date. Although, the exact number of extant fungal species is not known, several estimatives were raised by mycologists including an an estimate of 712,000 extant species elaborated by Schmit and Mueller (2007). However, recent estimates based on high-throughput sequencing methods suggest that as many as 5.1 million fungal species exist (BLACKWELL, 2011). Authors trying to estimate the number of extant yeast species usually conclude that more than 98% of yeasts are yet undiscovered. Given the average number of fungal species described each year since 1999 to be about 1200, Hibbett et al. (2011) predicted that at the current rate of discovery, it may take up to 4000 years to describe all fungal species. Since, local extinctions (i.e. strong reductions in the abundance of microbial species) occur quite frequently due to clearing of forests, agricultural activity or erupting volcanoes, it was predicted that microbial extinction rate might soon surpass the recovery rate of extant undescribed species.
Effort towards microbial diversity recovery has included sampling of microorganisms from various ecological habitats, especially those whose biodiversity are been endangered and development of culture collections by private laboratories, institutions and universities for ex situ conservation. The urgent need therefore arises for biodiversity and taxonomic studies with emphasis on the discovery, classification and description of novel species.
starch to convert it to linear oligomers and ultimately to glucose units, that are then fermented by yeasts. Xylan is the most abundant hemicellulose and xylanases are one of the
major hemicellulases which hydrolyse the β-1,4 bond in the xylan to short xylooligomers which are further hydrolysed into single xylose units by β-xylosidase. Microbial xylanases are becoming more demanding due to their wide application in various industrial sectors. In the energy sector, one area of considerable importance is the enhanced production of ethanol through the release of substantial amount of fermentable feedstock (PÉREZ et al., 2002; ALMEIDA et al., 2007). In the step involving the conversion of hemicelluloses to fermentable sugars, some of the hemicelluloses, mostly xylan, remain associated with the cellulosic-rich water insoluble fraction (CHANDRA et al., 2007). Because, effective enzymes capable of digesting these woody materials are still lacking, in order to improve cellulose accessibility, hence, enhance substrate digestibility, cellulose enzymes are often supplemented
with xylanases as ‘accessory enzymes’ (KUMAR; WYMAN, 2009). Hence, the demand for
xylanase producing microorganisms has increased.
One of the main areas of research in enzyme biotechnology has been driven by the need to isolate and identify organisms which are good producers of plant polysaccharides (lignocelluloses) degrading enzymes. In contrast to fungi and bacteria, few types of yeast are known to be capable of degrading lignocelluloses. Ability of yeasts to utilize plant polysaccharides is important because such information could be useful for taxonomic classification as well as biotechnological applications. Therefore, more information is needed about yeasts possessing these characteristics. However, it is known that yeasts are highly diverse in terms of nutrition, exploitation of ecological niches and secondary metabolism (this diversity reflects in their wide biotechnological applications in various industrial sectors such as food, beverages, chemicals, industrial enzymes, pharmaceuticals agriculture and environment) and that microbial diversity is the foundation for biotechnology; the basis for the discovery of new products, secondary metabolites and genes. Microbial culture collection serves as a pool where metabolically and genetically diverse yeast strains with unique properties and applications could be discovered.
Aims and objectives
State of São Paulo as well as to predicting biotechnological utility of newly discovered taxa and already existing taxa making up this collection.
The specific objectives of this research therefore were:
(i) to identify and characterize some yeast and yeast-like organisms at the UNESP –
Central for Microbial Resources of the Institute of Biosciences, Rio Claro. (ii) to describe novel species in the culture collection
(iii) to identify yeast and yeast-like organisms with ability to produce lignocellulose degrading enzymes namely: cellulases, hemicellulases and ligninase enzymes.
STUDY OUTLINE
The primary objectives of this study were to provide accurate identification of some yeasts hold at the UNESP – CMR with emphasis on classifying and describing new strains; as well as to provide information about the applicability in enzyme technology of some yeasts in this collection, particularly plant polysaccharide degrading enzymes namely amylases, cellulases, xylanases and pectinases. Here we organized the study into chapters with the first one being a brief background while the subsequent chapters present investigations and findings relating to the theme of the study.
The chapter opener begins with a brief introduction of the UNESP – CMR and how its activities have contributed to broadening the knowledge of fungal diversity. It also present reviews of literature about the definition of yeasts and how advancements in molecular biology have shaped the methods used for yeast identification. Because the yeasts examined in this work were isolated from different habitats during different ecological studies, effort was laid on briefly reviewing some roles played by yeasts in these habitats. Furthermore, this chapter highlights the enzyme systems involved in the degrading of plant polysaccharides and yeasts found from previous studies to possess these enzymes.
Chapter 2 presents taxonomic studies of yeasts hold at UNESP – CMR. Basically, this study involved the identification and classification of 340 yeast isolates obtained from various ecological studies embarked upon by the UNESP – CMR research team using a polyphasic approach. The identification process led to the discovery of several undescribed fungal species including two ascomycetous species whose novel status were proposed in the genus Candida (n = 2) and a basidiomycetous species proposed in the genus Bulleromyces. The findings of this study also revealed that the UNESP – CMR collection holds many metabolically and genetically diverse types of yeasts that could be harnessed for further biotechnological studies.
among other things that xylanase and cellulase activities are characteristics more expressed by basidiomycetous yeasts whereas, pectin degrading activities is more linked to ascomycetous yeasts. In addition, this study revealed three strains capable of producing amylases, celluloses, and pectinases at high levels from sugarcane bagasse fermentation.
Chapter 4 and 5 presents the description of three novel species discovered in this study. Two anamorphic ascomycetous species namely Wickerhamiella kiyanii (strain FB1-1DASPT) and Wickerhamiella pindamonhangabaensis (strains H10YT and H10-10AY) were proposed and their taxonomic descriptions as well as systematic classifications are provided in chapter 4. The presence of lipase enzyme systems was demonstrated in the latter species. These two species were found to phylogenetically belong to the Wickerhamiella clade (Saccharomycetes, Saccharomycetales). On the other hand, chapter 5 presents the description of a teleomorphic species proposed in the genus Bulleromyces. These two chapters contribute to our knowledge of yeast diversity. The description of these species will permit the deposition of their holotypes as well as their nomenclatural information in internationally recognized and publicly accessible culture collections as well as the official publication of their names.
Finally, chapter 6 presents a study on the intraspecific variation and emendation of Hannaella kunmingensis. Our study so far with yeasts has been revealing the existence of intraspecific genetic and phenotypic variations among different strains of yeasts belonging to the same species. In one of such scenarios, three strains of yeasts from the UNESP – CMR were found to differ by 11 nucleotide substitutions in the internal transcribed spacer (ITS) region from H. kunmingensis (CBS 8960T), but based on their conspecificity in the D1D2 domains of the large subunit ribosomal DNA gene, the 3 strains were considered as H. kunmingensis species. The later discovery of sequences of 7 other similar strains deposited in the GenBank offered the opportunity to intensively study the intraspecific variation in genetic as well as phenotypic properties of this species. Using parsimony network analysis, the presence of three genetic haplotypes in H. kunmingensis was demonstrated. The study also revealed variations in morphological characteristics as well as biochemical characteristics among the 11 strains studied. Based on these findings, an emendation of H. kunmingensis species was carried out. Besides contributing to the knowledge of the intraspecific diversity, the study contributed to increasing the number of strains of H. kunmingensis, which was formely
CHAPTER 1
1. BACKGROUND OF STUDIES AND LITERATURE REVIEW
The increasing awareness of the importance of biodiversity and the hidden genetic potential has resulted in a rise in recognition of the value of microbial culture collections. From the early days of biodiversity surveys and fieldtrip collections, microbiologists have been gathering samples and evidences related to their discoveries. In order to identify and perform more advanced investigations and analyses on their collected specimens, they had to be kept alive and maintained in a condition as close as possible to their original states. When properly preserved, microbial strains can maintain the same properties found in nature and can therefore be reused in many different types of studies, such as physiology, genetics or applied biotechnology. This is where the idea of culture collections came into being.
Microbial culture collections are living libraries and reference sources of microorganisms. In the past, due to the lack of adequately functioning and reliable culture banks, many microbial cultures were lost (MAHILUM-TAPAY, 2002). The first scientist to realize the importance of culture collection was Professor Frantisek Kral (1846-1911) who collected cultures and made it available for free to other researchers. His collection was later transferred to Vienna in 1915 (MALIC; CLAUSE, 1987). The Centraalburreau voor Schmmelcultures (CBS) culture collection, Netherlands, is the next oldest collection been founded in 1906. A list of some major culture collections in various countries is given in Table 1.1.
1.1 UNESP Central for Microbial Resources (UNESP- CMR),
In year 2006, the Microbiology laboratory (LAM) of the Institute of Biosciences, Rio Claro, developed a private collection of microorganisms, mainly yeasts, filamentous fungi and actinomycetes formerly known as Center for the Study of Social Insects (CEIS) UNESP campus of Rio Claro. Recent institutionalization of this culture collection led to its renaming
Table 1.1 - Some major international microbial culture collections, their acronyms and type of culture holding
Culture collection (acronym) Patent institute Type of culture Location Web address Agricultural Research Service Culture
Collection (NRRL)
National Center
for Agricultural Utilization Research Actinomycetes, Bacteria, Fungi Peoria, Illinois USA http://nrrl.ncaur.usda.gov
All-Russian Culture Collection (VKM) Russian Academy of Sciences Bacteria, Archaea, Fungi Moscow Region,
Pushchino, Russia http://www.vkm.ru
American Type Culture Collection (ATCC) American Type Culture Collection Bacteria, Archaea, Fungi, viruses Manassas, Virginia USA http://www.atcc.org
Brazilian Collection of Environmental and
Industrial Microorganisms University of Campinas, São Paulo
Bacteria, Filamentous fungi and
Yeast Campinas, Brazil
http://webdrm.cpqba.unicamp.br /cbmai/
Belgian Coordinated Collections of Micro-Organisms (BCCM)
Belgian Coordinated Collections of
Micro-Organisms Bacteria and Fungi Various cities, Belgium http://bccm.belspo.be/index/php Centraalbureau voor Schimmelcultures
(CBS)
Institute of Royal Academy, Art and
Science Fungi
Utrecht,
The Netherlands http://www.cbs.knaw.nl German Collection of Microorganisms and
Cell Culute (DSMZ) Leibniz-Institute Bacteria, Fungi and Plant Viruses Braunschweig, Germany http://www.dsmz.de/home.html
Japan Collection of Microorganisms (JCM) RIKEN BioResource Center Bacteria, Archaea, Fungi Saitama, Japan http://www.jcm.riken.jp
National Collection of Yeast Cultures (NCYC)
Food Research Institute, Colney Lane, Norwich NR47UA, Norfolk, UK
Yeasts and other known pathogens
Norwich,
United Kingdom http://www.NCYC.co.uk National Institute of Technology Evaluation
_ Biological Resource Center (NBRC)
National Institute of Technology and Evaluation
Actinomycetes, archaea and
Fungi Chiba, Japan http://www.nbrc.nite.go.jp Phaff Yeast Culture Collection, University
of California, University of Carlifonia, Davis Yeasts Davis, California, USA http://www.phaffcollection.org Portuguese Yeast Culture Collection
(PYCC)
Faculdade de Ciências e Tecnologia Universidade Nova de Lisboa
Yeast
Caparica, Portugal http://[email protected]
environments impacted with petroleum and its products, 1,000 fungi associated with marine environments (coastal Antarctica and Brazil), and about 100 bacteria used as reference strains; and the number of cultures is rapidly growing due to the many biodiversity studies embarked upon by researchers affiliated with this culture collection.
The collection was formed from research projects or collaborative research coordinated with the participation of researechers of UNESP and has been used for diversity studies and prospecting compounds of biotechnological interest, such as, enzymes, antibiotics, biofuels and bioremediation agents. UNESP - CMR is divided into two main parts namely (i) the main collection and (ii) the research collection. The main collection comprises isolates with biotechnological potential or are representatives of the biodiversity of a given environment and the reference strains and which have been correctly identified based on molecular taxonomy (ribosomal DNA sequencing, genetic fingerprinting and phylogenetic analysis) and conventional taxonomy (morphological and biochemical) while the strains contained in the research collection comprises isolates recovered from different environmental samples still under study and are potential candidates to be incorporated into the main UNESP- CMR collection after being subjected to necessary preliminary bioprospective studies as well taxonomic characterization. All the microorganisms in the UNESP - CMR are stocked and preserved according to the recommended guidelines of the Organization for Economic co-operation and Development OECD Best Practice Guidelines for microbial recource centers (Table 1.2) for microorganisms.
heliconiae, Candida picinguabensis and Candida saopaulonensis from flowers of Heliconia (RUIVO et al. 2006) and Trichosporon chiarellii from a nest of the fungus-growing ant Myrmicocrypta (PAGNOCCA et al. 2010). In addition, the culture collection contributed to the description of a new yeast genus, Bandoniozyma and seven new species inside (VALENTE et al., 2012).
Due to the knowledge accumulated over the years, from various physiological and taxonomic studies, about the vast metabolic activities of the microorganisms making up the UNESP – CMR, bioprospecting of the collection has also been extended to screening of yeasts for biotechnologically important enzymes. Furthermore, currently, a research involving the bioprospection of the yeasts from this collection for the conversion of lignocellulosic materials (sugarcane bagasse) to ethanol is being implemented.
Hence, UNESP – CMR, not only serve as a biotechnology-based bank of valuable microorganisms that could be harnessed for biotechnological applications, teaching and research and other purposes, but also as microbial repository for the preservation of Brazilian microbial biodiversity which constitute the heritage of the country.
1.3 Importance of culture collections to microbiology and biotechnology
i. Depository of Microbial Diversity
been isolated since then. Furthermore, many recent species descriptions and publications are based on materials collected in the past but preserved in culture collections with the hope that new technologies that could facilitate their description would later be developed (LACHANCE, 2006).
ii. Screening and exploitation of microbial diversity
Culture collections play important roles in providing lage microbial resources for screening for important biotechnological products. Introducing screening programs to collections allow biodiversity rich countries to benefit from exploitation of the microbial diversity they have (SMITH, 2003). For scientists who are interested in screening for novel products, culture collections provide an opportunity of access to large numbers of authenticated microorganisms. For example, in a search for L-arabinose fermenting yeasts for the bioconversion of biomass to ethanol, Dien et al. (1996) carried out an extensive screening of 116 yeast strains, from the ARS Culture Collection (National Center for Agricultural Utilization Research, Peoria, Illinois) in which four species namely Candida auringiensis, Candida succiphila, Ambrosiozyma monospora, and Candida sp. (YB-2248) were found to be able to ferment the sugar. In the same vein, Hou and Johnston (1992) screened 1229 selected microbial cultures (including 508 bacteria, 479 yeasts, 230 actinomycetes and 12 fungi) obtained from the ARS Culture Collection for lipase activities in which 25 % were lipase positive.
ii. Development and training of new preservation methods and skills
group of microorganisms. They also provide specialized training in culture handling and taxonomic studies. In addition, the identification of organisms is generally a specialized activity; hence, many collections provide identification services for people who have do not have the required skills or facilities required.
1.3 Yeast: introduction and definition
Yeasts are microscopic fungi, which reproduce asexually by budding or fission and by the production of forcefully ejected ballistoconidia on stalks termed sterigmata; resulting in growth made up of single cells and whose sexual states are not formed within or upon a fruiting body (SUH et al. 2006; KURTZMAN; FELL; BOEKHOUT, 2011a). These characteristics differentiate them from other filamentous fungi and mushrooms that are predominantly multicellular, and whose sexual structures are enclosed within complex fruiting bodies (KURTZMAN; FELL; BOEKHOUT, 2011a). In contrast to the yeasts, filamentous and dimorphic fungi grow by means of hyphae that extend at their apices while branching sub-apically, thereby resulting in an interconnected network of hypha known as mycelium. Several yeasts however exhibit pseudohyphae made up of chains of elongated buds that remain attached to the parent cell after formation. In addition to ascomycete and basidiomycete yeasts, some fungi are dimorphic and exhibit a yeast stage that shifts to mycelial growth under certain cultural conditions. The term
‘‘yeast-like’’ has also been used to represent the cellular phase of dimorphic members of the
zygomycete genus Mucor (FLEGEL, 1977), the black yeasts (HOOG, 1999), which comprise diverse pigmented ascomycete genera such as Aureobasidium, Fonsecaea and Phaeococcomyces as well as certain achlorophyllous algae in the genus Prototheca (KURTZMAN; FELL; BOEKHOUT, 2011a).
1.4 Evolution of yeast identification methods
of hyphae and pseudohyphae, fermentation of simple sugars and growth on various carbon and nitrogen compounds (KURTZMAN, 2011). Chemotaxonomic characteristics such as cell wall carbohydrate composition and coenzyme Q are also sometimes used in yeast taxonomy (PRILLINGER et al. 2011). The need for more accurate diagnostic tools for yeast species identification soon became apparent after various works began to reveal variations in phenotypic characteristics displayed by yeast strains of the same species (KURTZMAN; FELL, 2006; KURTZMAN, 2011). The shift from phenotypic identification of yeasts to molecular identification began with DNA reassociation techniques. In this technique, single-stranded DNAs of two isolates are mixed and allowed to repair as a double strand. According to Price, Fuson and Phaff (1978), strains that showed 80% or greater nuclear DNA relatedness on the basis of shared phenotype, as measured by reassociation, are members of the same species. Although, DNA reassociation technique provided the first opportunity of genetic based yeast species delimitation, it is limited by the fact that only closely related species can be resolved using the technique.
The use of DNA sequence comparisons for yeast identification soon became widely embraced after Peterson and Kurtzman (1991) and Kurtzman and Robnett (1998), studied the variable domains (D1 and D2) of the large subunit (LSU) rRNA gene of ascomycetous yeasts (Figure 1.1) and revealed that these regions offer the opportunity to resolve most closely and distantly related species. Kurtzman and Robnett (1998) predicted that strains showing six or more nucleotides differences (1% substitution) in the D1/D2 nucleotide domains of the ribosomal DNA (Figure 1.1) represent different species. A complementary D1/D2 database was created for basidiomycetous yeasts by Fell et al. (2000). Molecular analysis of the small and large subunit of the rRNA gene led to significant progress in systematic of basidiomycetous yeasts while various works such as Sugita et al. (1999, 2000) and Scorzetti et al. (2002) contributed to the development of databases for the internal transcribed spacers (ITS) 1 and 2 regions of the rDNA (figure 1) for basidiomycetous yeasts. The ITS region is highly substituted and provides a higher resolution for some closely related species that could not be separated by D1/D2 region (SCORZETTI et al. 2002). Several other genes have been successfully used for yeast taxonomic classification and they include the intergenic spacer (IGS) region, translation elongation factor-1α
Figure 1.1 Map of fungi rRNA gene showing the internal transcribed spacer (ITS) region, intergenic spacer (IGS) regions, 18S small subunit (SSU) and 25-28S large subunit (LSU).
Source: http://biology.duke.edu/fungi/mycolab/primers.htm
DNA include restriction-enzyme fragment length polymorphism (RFLP), random amplified polymorphic DNA (RAPD), amplified polymorphic length polymorphism (AFLP) and microsatellites (OLIVE; BEAN, 1999).
offers the opportunity of detecting nucleotide substitution rates and basal lineages are generally well supported (KURTZMAN; ROBNETT, 2003; KURTZMAN; FELL, 2006). Scorzetti et al. (2000) proposed the analysis of combined sequences of the D1/D2 and ITS regions for species identification after examining the resolution provided by these rRNA regions for the identification of basidiomycetous yeasts in the Pucciniomycotina and Agaricomycotina. Using a multigene approach of combined sequences of SSU, LSU, ITS, mitochondrial small subunit rDNAs with elongation factor 1-α and cytochrome oxidase II, species of the Saccharomyces complex were phylogentically resolved into 14 clades including 4 (Saccharomyces sensu stricto, Zygosaccharomyces sensu stricto, Torulaspora and Eremothecium) well-supported monophyletic clades (KURTZMAN; ROBNETT, 2003). By analyzing the combined sequences of SSU, LSU, ITS region and mitochondrial cytochrome b gene, Wang and Bai (2008) clearly separated three monophylectic clades namely, Dioszegia clade, Derxomyces mrakii and Hannaella sinensis from Cryptococcus luteolus lineage of the Tremellales. A similar approach by Kurtzman, Robnett and Basehoar-Powers (2008) found species formerly assigned to Issatchenkia to belong to the Pichia membranifaciens clade after the analysis of concatenated gene sequences from EF-1α and the LSU and SSU rRNA genes, hence, were consequently transferred to the genus Pichia.
Phylogenetic tree constructing methods do not accurately measure gene genealogies of haplotypes resulting from intraspecific polymorphisms (CLEMENT; POSADA; CRANDALL, 2000). The extent of intraspecific genetic and phenotypic (morphological and physiological) variations among yeast strains that share a common gene pool have been continuously revealed by several studies (LACHANCE et al., 2010, 2011; DAYO-OWOYEMI et al., 2012) and this phenomenon is more pronounced in basidiomycetous yeasts as was exemplified in the work of Scorzetti et al. (2002).
especially where such taxa were adequately sampled. The inter-specific sequence variation and high discriminative power of the ITS region have assisted in solving many taxonomic and systematic problems relating to separation of species, hence it has served the role of a DNA barcode marker for some fungi identification (BEGEROW, 2010). However, the ITS region is often highly variable within species (BEGEROW, 2010), consequently, members of the same species may be treated as being separate when pairwise ITS sequence divergence is used as a means of species delimitation. Nevertheless when combined with the D1/D2 region for parsimony network analysis, intraspecific discontinuities can be identified (LACHANCE et al. 2010, 2011).
1.5 Yeast classification
Yeasts are classified under two broad taxonomic groups, i.e. ascomycetes and basidiomycetes; each comprising anamorphic and teleomorphic states. These two groups of yeast differ in their cell wall composition and molecular structure and also in their mode of bud formation (i.e. asexual reproduction) and spore formation (sexual reproduction). Yeast cellular compositions vary with their phylogenetic diversity, as observed by the variety in the biochemical composition of the cell walls, ultrastructural organization and morphology of the septa (VAN DER KLEI et. al. 2011). However, cell walls of ascomycetous yeasts consist of two layers: an inner skeletal layer consisting of load-bearing polysaccharides and an outer layer consisting of glycoproteins that are covalently linked to the inner layer (YAMAGUCHI et al. 2002; SUH et al. 2006, VAN DER KLEI et al. 2011). In basidiomycetous yeasts, the walls are often, but not always, multilayered with alternating regions of dense and electron-translucent material. While the cell walls of ascomycetous and basidiomycetous fungi contain a
similar β-1,3-glucan, cell wall polysaccharide composition is dominated by chitin in the basidiomycetes (SUH, 2006, VAN DER KLEI et. al., 2011).
In the ascomycetous yeasts, budding (vegetative reproduction) is holoblastic, in which budding results from the stretching out of the entire cell wall of the mother cell; the bud separates from the narrow base leaving a scar through which no further budding occurs.
to recurrent formation and separation of a succession of buds (KURTZMAN; FELL; BOEKHOUT, 2011a). In ascomycetous yeasts, sexual reproduction occurs through spore formation whereas; sexual states of basidiomycetous yeasts are characterized by formation of septate dikaryotic hyphae with clamp connections. Basidiomycetous yeasts can also be differentiated from ascomycetous yeasts based on Diazonium Blue B (DBB) test. A dark red reaction is observed on the former when a buffered solution of Diazonium Blue B (DBB) is applied to cultures left overnight at 60 °C (BOEKHOUT et al., 2011).
i. Phylogenetic classification of ascomycetous yeasts
The relationships among the ascomycetes are becoming clearer since the introduction of multigene sequence analysis for the estimation of phylogeny; and consequently have resulted in changes in the classification of the ascomycetous yeasts. For instance, the reassignment of some formerly Pichia species (e.g P. anomala) to a new genus Wickerhamomyces after the polyphylectic nature of Pichia species was further confirmed by phylogenetic analyses (KURTZMAN; ROBNETT; BASEHOAR-POWERS, 2008; KURTZMAN, 2011). Another important change is the re-assignment of the saturn spored genus Williopsis into four distinct genera namely Barnettozyma, Lindnera, Ogatae and Wickerhamomyces.
RPB2) (JAMES et al., 2006) confirmed that ascomycetous yeasts belong to a single lineage, Saccharomycotina.
Yeasts in the subphylum Taphrinomycotina are classified in the order Schizosaccharomycetales (Schizosaccharomycetes) while the subphylum Pezizomycotina consists majorly of filamentous fungi. The teleomorphic genus Schizosaccharomyces Linder is the only genus of yeasts currently known in the Schizosaccharomycetales and currently includes 3 species namely Schiz japonicas, Schiz. pombe and Schiz.octosporus. Some distinct characteristics of this genus is their cylindrical shape and asexual form of reproduction which is by fission (VAUGHAN-MARTINI; MARTINI, 2011).
ii. Phylogenetic classification of basidiomycetous yeasts
Basidiomycetous yeasts including sexual and asexual forms are currently classified into three subphyla namely (i) Subphylum Pucciniomycotina, (formerly class Urediniomycetes), (ii) Subphylum Ustilaginomycotina (formerly class Ustilaginomycetes) and (iii) Subphylum Agaricomycotina (formerly Hymenomycetes) (Figure 1.3) (HIBBETT et al. 2007). The presence of three main lineages within the Basidiomycota was earlier demonstrated by many phylogenetic studies including sequence analyses small-subunit SSU (FELL et al. 2000), LSU and ITS (SCORZETTI et al. 2002).
Ustilaginomycetes and class Exobasidiomycetes. The Exobasidiomycetes include anamorphic species such as Tilletiopsis and its teleomorph Tilletiaria, Acaromyces, Meira, Malassezia and Sympodiomycopsis while only two species namely Farysizyma and Pseudozyma are currently classified in the Ustilaginomycetes.
The subphylum Agaricomycotina formerly known as Hymenomycetes comprises three classes, namely Agaricomycetes, Dacrymycetes and Tremellomycetes (HIBBETT et al. 2007). Yeast states are found only in the Tremellomycetes and are classified in four distinct orders namely Tremellales, Trichosporonales, Filobasidiales and Cystofilobasidiales (SCORZETTI et al. 2002). While ballistoconidia may be produced by members of the Tremellales (e.g., Bullera, Dioszegia, Kockovaella) and the Cystofilobasidiales (e.g., Udeniomyces), distinct morphological trait of the Trichosporonales (e.g Trichosporon chiarellii, T. ashaii, T. insectorum) is the formation of arthroconidia. Some sexual states of the Tremellales such as Auricullibuller fuscus, Bulleribasidium and Papiliotrema are characterized by the formation of conspicuous basidiocarps and tremelloid haustoria branches that are adapted to their mycoparasitic mode of life (BOEKHOUT; FONSECA; BATENBURG-VAN DER VEGTE, 1991; SAMPAIO et al. 2002). The teleomorphic genus Tremella and the anamorphic genera Bullera and Cryptococcus are polyphyletic and latter is distributed in all the four lineages of the Tremellales (SCORZETTI et al. 2002). Three teleomorphic genera are dinstincted in the Cystofilobasidiales namely Mrakia, Cystofilobasidium and Xanthophyllomyces. The first two genera are characterized by the formation of teleospores. Examples of species found in this lineage include species of the cold tolerant genus Mrakia (e.g M. frigida and M. gelida) and the carotenoid pigmented species of Cystofilobasidium e. g Cystofilobasidium infirmominiatum.
1.6 Yeast ecology and diversity
Yeasts are typically known as decomposers of dead organic matter in which they bring about nutrient transformation. They can engage in intimate relationships with other organisms as mutualists, competitors, parasites, or pathogens (STARMER; LACHANCE, 2011). Yeasts are widely distributed in almost all biomes of the world and their ubiquity is complemented by their diversity (KURTZMAN; FELL; BOEKHOUT, 2011c). They grow well in moist environments where there is availability of simple sugars, amino acids. Some are however able to degrade complex polysaccharide such as starch, cellulose, hemicellulose and pectin (ALONSO et al. 2010; BIELY; KREMNICKÝ, 1998). The following discussions of diversity of yeasts in various habitats relate to some of the isolation sources of the yeasts used in this study.
i. Yeasts associated with plants (flowers and fruits)
Due to their rich content of easily utilizable carbon, fruits serve as natural habitats for a variety of yeasts. Yeasts particularly ascomycetes are distributed on surfaces of fruits, exudates of leaves, flowers and tree trunks. Prada and Pagnocca (1997) reported the isolation of two hundred and two strains of yeasts and yeast-like fungi from naturally occuring fruits in the tropical rain forest of Juréia-Itatins Ecological Reserve, with 38 species constituting 74 % of the total isolates been ascomycetous. The finding that naturally occurring apple yeasts can protect fruit against postharvest diseases spurred interest in the isolation of yeasts from various fruits with the aim of discovering new yeast antagonists against postharvest diseases (JANISIEWICZ, 1987). Certain yeasts associated with fruits can produce sugar-derived prebiotics such as fructooligosaccharides. According to Maugeri and Hernalsteens (2007) apud Johnson and Echavarri-Erasun (2011), yeasts from fruits and flowers in Brazilian tropical forests including Candida,Rhodotorula, and Cryptococcus produced substantial quantities of fructooligosaccharides.
ii. Yeasts associated with leaf phylloplane
FONSECA; INÁCIO, 2006). Basidiomycetous yeasts particularly species of the genera Sporobolomyces, Rhodotorula, Cryptococcus, Bensingtonia, Cystofilobasidium, Leucosporidium and Pseudozyma are dominant on leaf phylloplanes (MAKSIMOVA; CHERNOV, 2004; STAMER; LACHANCE, 2011). Species of Bullera, Sporobolomyces and Tilletiopsis are particularly adapted to this environment due to the production of forcibly ejected ballistospores (FONSECA; INÁCIO, 2006). A survey on the phylloplane yeasts from Mediterranean plants (Acer monspessulanum, Quercus faginea, Cistus albidus and Pistacia lentiscus) collected at the
‘Arrabida Natural Park,’ Portugal revealed the high frequency of occurrence of species of
Taphrina and Lalaria including five previously unknown species of the latter (INÁCIO, 2004). In an investigation studying the yeasts community colonizing the leaf surfaces of various fruit trees in southwest Slovakia, Slávicová, Vadkertiová and Vránová (2009) isolated 150 strains belonging to seventeen yeast species out of which Aureobasidium pullulans, Cryptococcus laurentii, and Metschnikowia pulcherrima were the most abundant species isolated.
Yeasts are also found living in fruiting bodies of mushrooms, which has also been the source of many ascomycetous and basidiomycetous yeast species (NAKASE et al. 1999; BABJEVA et al., 2000; MIDDELHOVEN, 2004). Yurkov et al. (2012) isolated various yeasts species of Rhodotorula, Rhodosporidium, Mastigobasidium, Cryptococcus,
Cystofilobasidium, Holtermanniella, Trichosporon and the ascomycetous Kluyveromyces from Boletales fruiting bodies truffle ascocarps. Nakase et al. (1999) isolated three new species of yeasts namely Candida fungicola, C. sagamina and C. fukazawae from fruiting bodies of unidentified mushrooms collected from Tanzawa Mountains, Kanagawa Prefeitura.
iii. Yeasts associated with insects
Yeasts have been isolated from insects in many different families. While such insects serve the primary role of dispersing yeasts to new habitats, many of these yeasts have also been shown to improve insect nutrition and to detoxify plant chemicals to which insects are exposed (SUH et al., 2005; LACHNACE et al., 2001, ROSA et al., 2003).
Acromyrmex) exclusively cut fresh leaves and plant material to cultivate their mutualistic fungus (WEBER, 1972; MUELLER; REHNER; SCHULTZ, 1998). Breakdown and transformation of the protein and starch rich plant materials by the cultivated fungus make nutrients available within the fungus garden matrix and this is a major target for exploitation by other microorganisms including yeasts (CARREIRO et al., 1997; RODRIGUES et al., 2009; SCOTT et al., 2010).
Yeasts associated with fungus-growing ant nests contribute to the stability of the complex microbiota found in the leaf-cutting ant nests through nutrients generation and removal of potentially toxic compounds (MENDES et al., 2012). Carreiro (1997) identified yeasts species of the genera Candida, Pichia, Cryptococcus, Rhodotorula, Sporobolomyces, Tremella and Trichosporon from the nests of the leaf-cutting ant Atta sexdens. Rodrigues et al. (2009) investigated the diversity of yeasts in Atta texana gardens and isolated ascomycetous yeasts belonging to the genera Aureobasidium, Candida, Kodamaea, Saccharomyces and basidiomycetous yeasts of the genera Bullera, Bulleromyces, Cryptococcus, Pseudozyma, Rhodosporidium, Rhodotorula, Sporidiobolus and Trichosporon.
The dispersal of yeasts by leaf-cutting ants was studied by Pagnocca et al. (2008) and it was revealed that these insects harbor various species of yeasts in their body including several opportunistic human pathogens e.g. Candida parapsilosis and C. metapsilosis, hence, may serve as vectors of these pathogens. Another groups of yeasts commonly found associated with fungus-growing ants are black yeasts in the genus Phialophora. Black yeast is the term used to refer to groups of yeasts characterized by melanized cell wall (STERFLINGER, 2006). These yeasts grown on the cuticle of the ants and are considered symbionts that play antagonistic roles in the fungus-growing ant mutualism (LITTLE; CURRIE, 2008).
Cryptococcus liquefaciens and Rhodotorula mucilaginosa being most frequently encountered (SANCHEZ DE SOUSA, 2011).
Ascomycetous yeasts associated with flowers and exudates are usually found in specialized niches involving interactions with insects or other invertebrate animals that they rely upon for dispersal (MORAIS et al., 1992; SUH et al., 2005; LACHNACE et al., 2001; ROSA et al., 2003). Lachance et al. (2001) reported the affiliation of yeasts in the Metschnikowia, Kodamaea, Wickerhamiella, and Starmerella clades with these floricolous insects that visit Hibiscus flowers and some flowers in the families Convolvulaceae and Cactaceae. Beetles may have a yet unclear strong symbiotic relationship with yeasts; possibly, the yeasts may carry out the transformation of scarce and poorly digestible components of flowers into a richer diet for the insects (STARMER; LACHANCE, 2011).
iv. Yeasts associated with honey
Ability of some yeast to tolerate low temperature, low oxygen concentration, high acidic conditions, high osmotic pressure or high salinity are important adaptive properties that determine their ability to survive under restricted habitats. Zygosaccharomyces rouxii and Z. bailii are often implicated in the spoilage of honey and jam because of their unique abilities to tolerate the high osmotic stresses and low water activity (MARTORELL et al. 2007). Other yeasts such as Z. bisporus, Z. mellis, Schizosaccharomyces pombe, Torulaspora delbrueckii, Debaryomyces hansenii, and various Candida and Moniliella species are commonly associated with foods containing high concentrations (40-70%) of sugar such as honey (FLEET, 2011). v. Yeasts found in soil
materials and carbon cycling, whereby they serve as mutrient sources for bacteria and other predators in the soil (FITTER et al. 2005; BOTHA, 2006; 2011).
Ecological soil surveys revealed the most abundant yeast soils to include Cryptococcus albidus, Cr. curvatus, Cr. gastricus, Cr. gilvescens, Cr. humicolus, Cr. laurentii, Cr. podzolicus. Cr. erreus, Filobasidium uniguttulatum, Cystofilobasidium capitatum, Leucosporidius scottii,
Mrakia frigida, Rhodotorula aurantiaca, R. glutinis, R. mucilaginosa, Sporobolomyces roseus, Trichosporon cutaneum and Schizoblastosporion starkeyi-henricii (BOTHA, 2006). Recently, Yurkov; Schäfer and Begerow (2012b) investigated the diversity of cultivable yeasts in soils under different land use and isolated 40 yeast species, 11 of which had earlier been reported from soil, i.e. the basidiomycetous Cryptococcus aerius, Cr. laurentii, Cr. terreus, Cr. terricola, Cr. podzolicus, Geotrichum pullulans and the ascomycetous Barnettozyma californica, B. pratensis, Schwanniomyces (Debaryomyces) occidentalis, Lindnera (Williopsis) saturnus and Schizoblastosporion starkeyi-henricii.
vi. Yeasts in Antarctic environments
Many species of yeasts have been found to successfully colonize the Antarctic continent. While some of these yeasts are Antarctic indigenes and obligate psychrophiles that do not survive when subjected to temperatures different from those obtainable in their natural habitat (i.e., > 20 °C), several could be considered as non-indigenes that were brought in by wind and ocean currents as well as by birds, humans and other animals who occasionally visit this habitat, and became adapted to Antarctic habitat (VISHNIAC, 1996; 2006b). The latter groups are mostly psychroptrophs and mesophiles, which equally grow
and multiply at room temperature (25 ± 2 °C), although may remain dormant for a long time at low temperatures. The ability to tolerate low temperature, high salinity, high radiation and other extreme conditions are fundamental adaptations of yeasts found in Antarctic environments (RAY et al., 1989; ROBINSON, 2001; SHIVAJI; PRASAD, 2009). Yeasts such as Candida psychrophila, Leucosporidium antarcticum, Cr. vishniacii, Mrakia frigida, Mrakia robertii and Mrakia blollopis are obligate psychrophiles are not able to grow at temperatures above 20 °C (VISHNIAC, 2006b; THOMAS-HALL et al., 2010).
Leucosporidium scottii (VISHNIAC, 2006b; VAZ et al., 2011). Some of them are mesophilic yeasts that became adapted to Antarctic habitat. The extreme environmental conditions obtainable in the Antartica means that microorganisms found there would have evolved unique characteristics for survival that could be exploited for biotechnological applications. Hence, yeast biodiversityof the Antarctica has raised interest for bioprospection for novel enzymes and biomolecules (SHIVAJI; PRASAD, 2009; VAZ et al., 2011).
1.7 Biotechnological importance of yeasts
The biotechnological potentials of yeasts have been exploited by man in many industrial processes ranging from food industries to the biofuel industries where yeast is used for the production of bioethanol. Yeasts, especially Saccharomyces cerevisiae are used for making various fermented products such as beer, wine, bakery products, cheese etc. They are also used in the production of enzymes, biocatalysts, pigment, flavours, and pharmaceutical prouducts as well as as biocontrol agents (JOHNSON; ECHAVARRI-ERASUN, 2011). Meyerozyma (Pichia) guilliermondii is known as a hyper producer of riboflavin (SIBIRNY; BORETSKY, 2009). Due to the ease of growth and genetic manipulation, yeasts such as S. cerevisiaeSchizosaccharomyces pombe are used as model organisms for genetic studies (TAKEGAWA, et al., 2009).
Other biotechnological potentials been investigated in yeasts include phenol and alkane degradation by Candida maltosa and C. tropica, production of biosurfactants by Pseudozyma spp., production of heterologous protein by Schizosaccharomyces pombe, fermentation of xylose to ethanol by Scheffersomyces stipitis and production of lipids and single cell oil by Yarrowia lipolytica (SATYANARAYANA; KUNZE, 2009).
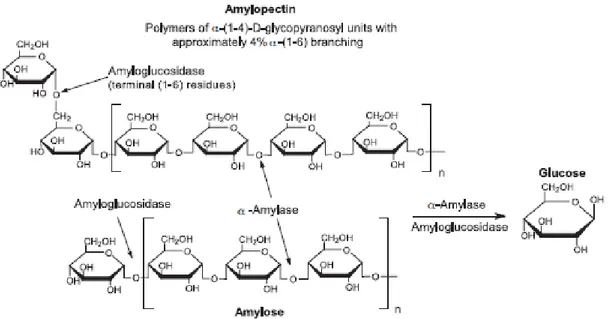
1.8 Biodegradation of starch, lignocelluloses and pectin
Plant cell walls are the planet`s dominant form of lignocellulose biomass. The main
Figure 1.4. Plant cell wall structure. Source: Lodish et al. (2000)
ZIPURSKY et al., 2000; PÉREZ et al., 2002; HOWARD et al., 2003). Hemicelluloses are polysaccharides consisting of xylan, glucuronoxylan, arabinoxylan, glucomannan, and xyloglucan present along with cellulose in plant cell walls (LODISH et al. 2000; SCHELLER; ULVSKOV, 2010).
i. Cellulose degradation
Cellulose is the most abundant renewable organic polymer in the biosphere and is highly crystalline, water insoluble and relatively resistance to depolymerization. The degradation of cellulose to glucose requires the synergistic action of three distinct classes of enzymes namely, (i)
endoglucanases, (ii) exoglucanases and (iii) β-glucosidases (cellobiases) (EC 3.2.1.21) (KARMAKAR; RAY, 2011) (i) The ´´endo-1,4-β-glucanase`` or 1,4-β-D-glucan 4-glucanohydrolases (EC 3.2.1.4), breaks internal bonds of cellulose i.e., β(1→ 4) linkages, to
disrupt the crystalline structure and expose individual cellulose polysaccharide chains. (ii) The ´´exo-1,4- β-D-glucanases`` include both the 1,4- β-D-glucan glucohydrolases (EC 3.2.1.74) which liberate D- glucose from 1,4-β-D-glucans and 1,4- β-D-glucan cellobiohydrolase (EC 3.2.1.91) which liberates D-cellobiose from 1,4- β-glucans; resulting in tetrasaccharides or disaccharides, such as cellobiose. (iii) The ´´β-D-glucosidases`` or β-D-glucosidase glucohydrolases (EC 3.2.1.21) hydrolyses the exoglucanase product into individual monosaccharides i.e. release of D-glucose units from cellobiose (Figure 1.5) (KARMAKAR; RAY, 2011).
Cellulose activities have been found in yeasts such as Candida glabrata, C. stellata, C. sheatae, Kloeckera apiculata (STRAUSS et al., 2001) Aureobasidium pullulans (KUDANGA;
MWENJE, 2005; THONGEKKAEW; KHUMSAP; CHATSA-NGA, 2012). Trichosporon
cutaneum, T. pullulans (STEVENS; PAYNE, 1977) and Gueomyces pullulans (SONG et al. 2010).
Figure 1.5. Action of major cellulase enzymes. Endoglucanases cleave internal β-1,4-linkages; exoglucanases cleaves two to four units from the ends of the exposed chains produced by endoglucanase
while β-glucosidases cleave cellobiose to glucose units (KARMAKAR; RAY, 2011).
ii. Xylan degradation
The major enzymes responsible for the complete depolymerization of xylan are
collectively known as xylanases and are composed of various hydrolases namely β
-1,4-endoxylanase, β-xylosidase, α-L-arabinofuranosidase, α-glucuronidase, acetyl xylan esterase, and phenolic acid (ferulic and p-coumaric acid) esterase. Endoxylanases (EC3.2.1.8) act on the back