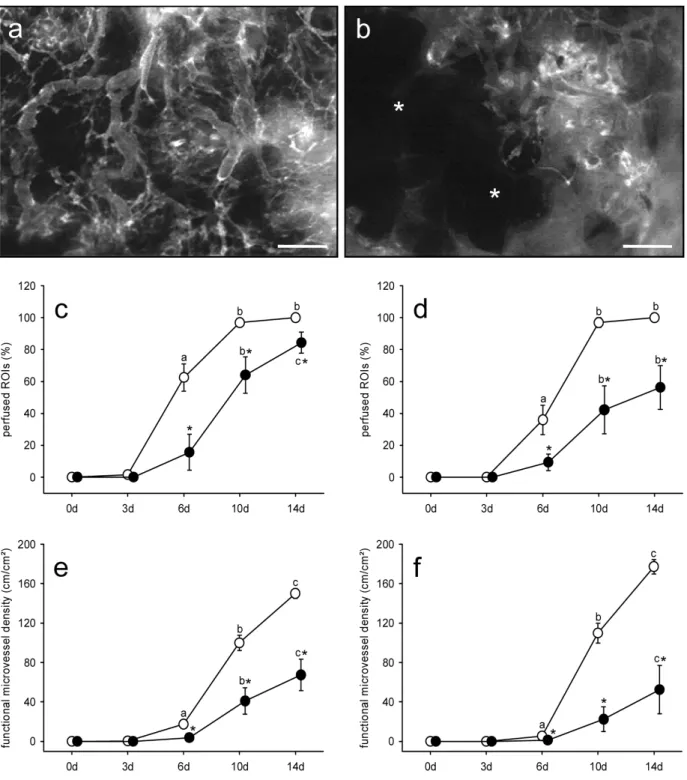
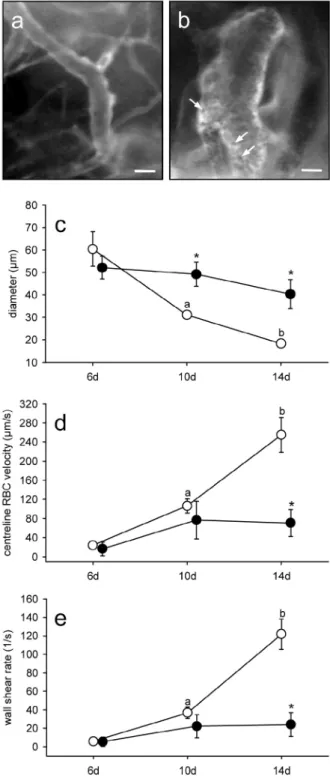
Adipose tissue-derived microvascular fragments from aged donors exhibit an impaired vascularisation capacity
Texto
Imagem
Documentos relacionados
Aos 7, 14, 21 e 28 dias após a aplicação (DAA), foi avaliada a porcentagem de controle das seguintes espécies de plantas daninhas: Ipomoea grandifolia, Brachiaria decumbens,
Also we have confirmed that, for fixed p, the growing of the internal structure of the defect (connected with larger values of q for the one-field model and smaller values of s in
CANTOS PARA A EUCARISTIA DOS 75 ANOS_cifras - Do Jaguaré padroeiro, aclamado com fé, Vosso povo inteiro, vem a vós,
O Sítio Arqueológico Sampaio, Alto Vale do Araçuaí, Felício Dos Santos, Minas Gerais: Paisagem, Cronologia e Repertório Cultural para Compreensão das Ocupações
7º - VIGÊNCIA: O prazo máximo de validade do registro de preços será de 12 (doze) meses, contados a partir da data da publicação da respectiva Ata.. 8º - A Ata de Registro
Este estudo tem como objetivo contribuir para a escrita da História da Paraíba durante o período colonial através da personagem Quitéria Bandeira de Mello, acusada por
Projetos Pedagó- gicos: Curso Técnico Subsequente em Plástico – Campus Caçador; Curso Técnico Concomi- tante em Vestuário – Campus Caçador; Curso Técnico Integrado
As condições previstas neste documento se aplicam apenas às pessoas físicas ou jurídicas não assinantes da Viasat Brasil, que contratarem o Serviço da Viasat Brasil
