1
DIMITRI EMILIANO PACHECO
Nitroforinas salivares de
(Hemiptera: Reduviidae):
Avaliação do RNAi parental e do papel da albumina do hospedeiro na
sua atividade biológica
UNIVERSIDADE FEDERAL DE MINAS GERAIS
BELO HORIZONTE
2
DIMITRI EMILIANO PACHECO
Nitroforinas salivares de
(Hemiptera: Reduviidae):
Avaliação do RNAi parental e do papel da albumina do hospedeiro na
sua atividade biológica
Orientador: Prof. Dr. Marcos Horácio Pereira
UNIVERSIDADE FEDERAL DE MINAS GERAIS
BELO HORIZONTE
2014
Dissertação apresentada ao Programa de Pós graduação em Parasitologia do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais como requisito parcial para obtenção do grau de Mestre em Parasitologia
3
4
Agradecimentos
Agradeço primeiramente pelo apoio incondicional dos membros da minha família e amigos íntimos.
Agradeço a meu orientador Prof. Marcos Pereira e aos professores Nelder, Maurício e Ricardo pela disponibilidade, pelos ensinamentos e discussões relevantes durante a elaboração do trabalho.
Agradeço a Rafaela, Adriana, Bruno, Layana, Felipe, colaboradores do trabalho realizado durante o mestrado pelo auxílio e atenção dispensados.
Agradeço a todos os meus colegas de laboratório pela amizade e convivência agradável durante o tempo em que trabalhamos juntos. Em especial ao César, um grande exemplo pra mim.
Agradeço a Mariana, Carol, Eliseu, Breno, Luísa, Larissa, Adalberto, Fernando, Fernanda, Matheus, Cleyd e Ingrid pelos ótimos momentos que passamos juntos durante o mestrado.
5 Sumário
Lista de abreviaturas e siglas ...8
Lista de figuras ...11
Lista de tabelas ...14
Resumo ...15
Abstract...16
Introdução ...17
1.1 – Biologia e importância dos triatomíneos ... 17
1.2 – Hematofagia e capacidade vetorial dos triatomíneos... 18
1.3 – Ação de biomoléculas salivares no processo alimentar dos triatomíneos ... 19
1.4 – Histórico das nitroforinas salivares ... 20
1.5 – RNA interferente ... 25
1.6 – Efeito sistêmico e longa duração do silenciamento por RNAi ... 26
1.7 – Silenciamento das nitroforinas salivares de por RNAi ... 28
2 – Justificativa ...30
3 – Objetivos...32
3.1 – Objetivo geral ... 32
3.2 – Objetivos específicos ... 32
4 – Materiais e métodos ...33
4.1 – Manutenção dos insetos ... 33
4.2 – Obtenção das amostras de glândulas salivares ... 33
6
4.4 – Genes das nitroforinas salivares (NPs) 1 a 4 ... 34
4.5 – Extração de RNA, síntese de cDNA e PCR ... 34
4.6 – Silenciamento por RNAi – síntese e introdução do RNA de fita dupla (dsRNA) nos insetos ... 36
4.7 – Estimativa de proteína total e hemeproteínas no extrato de glândula salivar .... 38
4.8 – Avaliação do RNAi parental ... 40
4.9 – Preparação e monitoramento do vaso dorsal dos insetos ... 42
4.10 – Atividade biológica do EGS e a influência da soro albumina ... 43
4.11 – Análise de imagem ... 45
4.12 – Análise estatística ... 46
5 – Resultados...47
5.1 – Avaliação da quantidade de proteínas salivares nos insetos ... 47
5.1.1 – Avaliação da quantidade de proteína total e hemeproteínas ... 47
5.1.2 – Avaliação do fenótipo e produção de ovos nos insetos injetados ... 49
5.1.3 – Avaliação da transmissão do silenciamento para a prole dos insetos injetados ... 51
5.2 – Ensaios em vaso dorsal de adulto de ... 59
5.2.1 – Ensaios de aplicação de EGS em soro humano inativado ... 59
5.2.2 – Ensaios de aplicação de EGS em solução de BSA a 20 mg/mL ... 61
5.2.3 – Ensaios com EGS de insetos silenciados para as nitroforinas ... 67
6 – Discussão ...64
7
8
Lista de abreviaturas e siglas
BSA – soro albumina bovina
cDNA – DNA complementar
cGMP – monofosfato cíclico de guanosina
dNTP – desoxirribonucleotídeo trifosfato dp – desvio padrão
dsRNA – RNA de fita dupla
dsRNA NPs – RNA de dupla fita com homologia aos genes das nitroforinas 1 a 4 dsRNA QR – RNA de fita dupla homólogo ao gene codificante da queratina
EDRF – fator relaxante derivado de endotélio EGS – extrato de glândula salivar
FAD – flavina adeninda dinucleotídeo
FPS – frames por segundo g – aceleração da gravidade
Kd – constante de dissociação µg – micrograma
µL – microlitro
µM – micromolar mg – miligrama
9 mL – mililitro
mM – milimolar
mRNA – RNA mensageiro n – número amostral
NADPH – nicotinamida adenina dinucleotídeo fosfato reduzida nm – nanômetros
NO – óxido nítrico
NPs – nitroforinas ns – não significativo
PAC – potencial de ação composto pb – pares de base
PBS – “phosphate buffered saline”
PCR – reação em cadeia da polimerase pH – potencial hidrogeniônico
RdRP – RNA polimerase RNA dependente
RISC – complexo de silenciamento induzido por RNA RNAi – interferência por RNA
RNAse – ribonuclease s – segundo
10 siRNA – “small interfering RNA”
Taq –
U – unidade
UR – umidade relativa
11
Lista de figuras
Fig. 1. Ciclo biológico de desenvolvimento de ... 18
Fig. 2. Aspecto das glândulas salivares de no tórax do inseto. ... 33
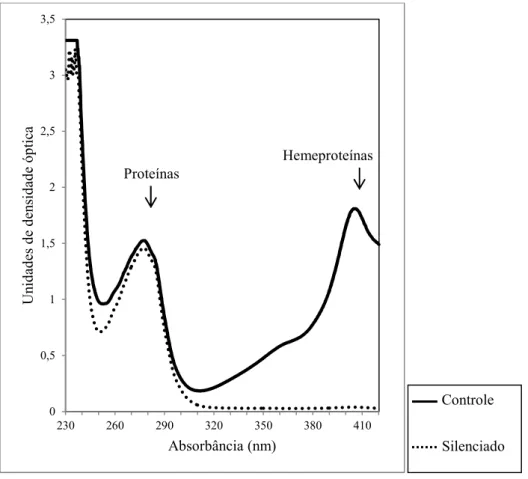
Fig. 3. Estimativa de hemeproteínas e proteína total na glândula salivar de adultos de injetados com dsRNA para NPs 1 4 ou queratina durante o quarto estádio por espectrofotometria. ... 39
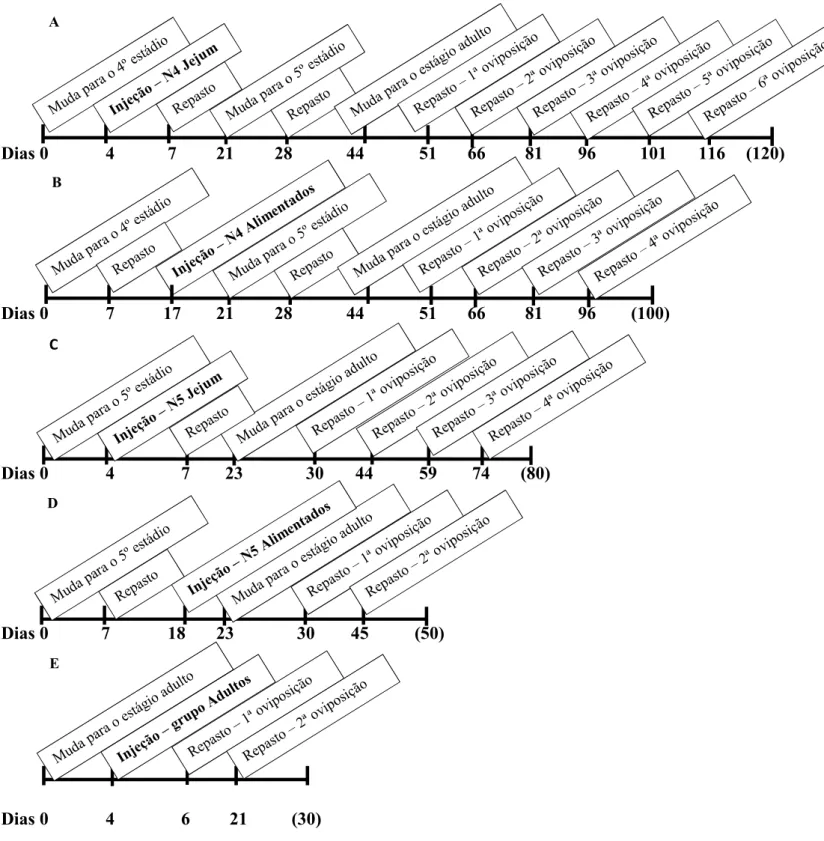
Fig. 4. Representação temporal do tratamento dos grupos de insetos injetados.. ... 41
Fig. 5. Montagem do aparato para experimentos no vaso dorsal de adulto de
... 43
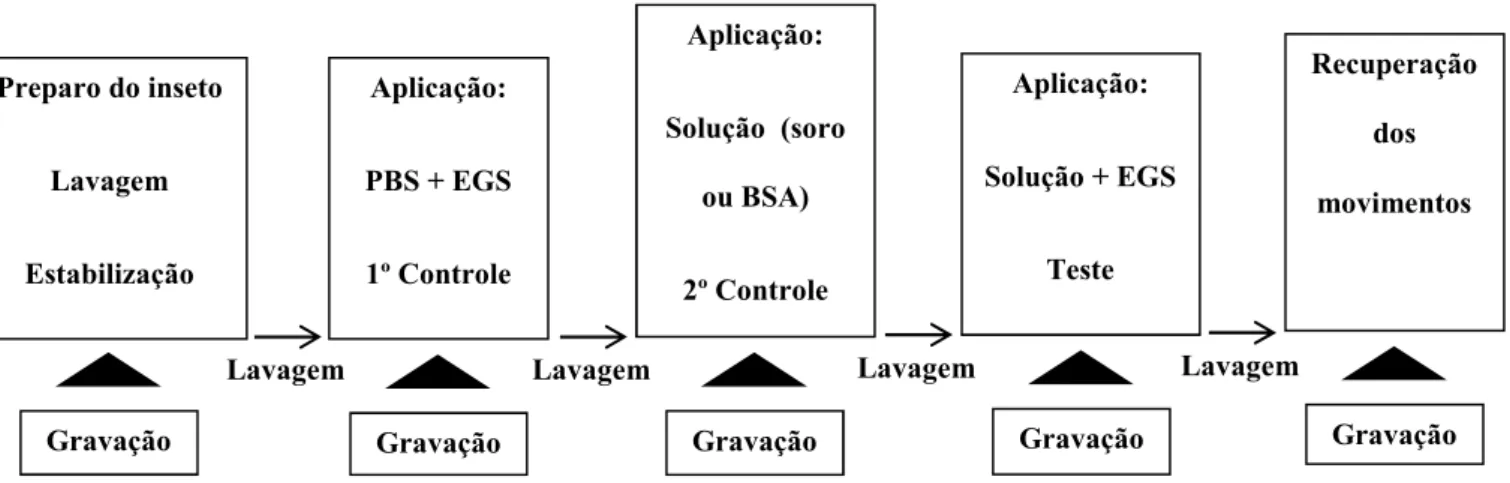
Fig. 6. Representação gráfica mostrando a sequência das amostras aplicadas nos ensaios de vaso dorsal de adultos de ... 44
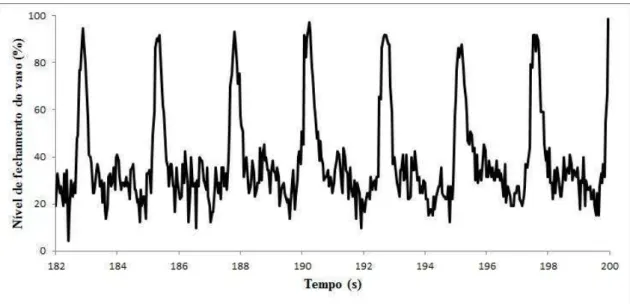
Fig. 7. Análise em vídeo das imagens obtidas a partir das gravações dos experimentos com vaso dorsal isolado ... 45
Fig. 8. Perfil de contrações do vaso dorsal isolado de adulto de a partir da análise das gravações em vídeo analisadas no ImageJ. ... 46
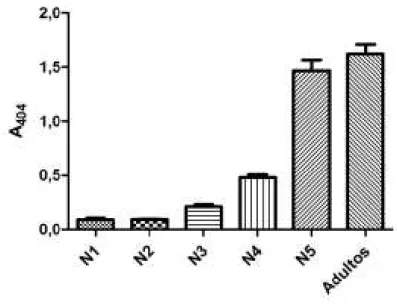
Fig. 9. Valores (expressos em média ± desvio padrão) da absorbância a 404 nm em espectrofotômetro do extrato de glândula salivar para os estágios de desenvolvimento pós embrionário de obtidos a partir de pools de 3 pares de glândulas. ... 47
Fig. 10. Valores (expressos em média ± desvio padrão) da concentração de proteína total presentes no extrato salivar para os estágios de desenvolvimento pós embrionário de obtidos a partir de pools de 3 pares de glândulas. ... 48
12
Fig. 12. Aspecto típico apresentado pelas glândulas salivares de adultos de
em microscópio estereoscópico. ... 50
Fig. 13. Curva espectral típica (240 420 nm) utilizada na estimativa de hemeproteínas e proteína total no EGS de ninfas de primeiro ínstar de por espectrofotometria. ... 52
Fig. 14. Razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quarto estádio em jejum. ... 55
Fig. 15. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quarto estádio após a alimentação.. ... 56
Fig. 16. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quinto estádio em jejum. ... 57
Fig. 17. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quinto estádio após a alimentação. ... 58
Fig. 18. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o estágio adulto em jejum. ... 58
Fig. 19. Frequência de contrações do vaso dorsal de adulto de após aplicação de soro humano inativado para o complemento e EGS (teste), individualmente e na presença de soro. ... 59
13
Fig. 21. Reestabelecimento do ritmo de contrações do vaso dorsal de exemplares adultos de após aplicação de amostra de extrato de glândula salivar + soro e subsequente lavagem com solução salina 0,9%. ... 60
Fig. 22. Frequência de contrações do vaso dorsal de adulto de após aplicação de BSA e EGS (teste), individualmente e em conjunto com BSA. ... 61
Fig. 23. Frequência de contrações do vaso dorsal de adulto de após aplicação de BSA e EGS de inseto silenciado para as NPs, individualmente e em conjunto com BSA. ... 62
Fig. 24. Frequência de contrações do vaso dorsal de adulto de após aplicação de BSA e EGS de insetos injetados com dsRNA para queratina, individualmente e em conjunto com BSA. ... 63
14
Lista de tabelas
Tabela 1 Sequencias dos iniciadores utilizados nos experimentos de PCR ... 37
Tabela 2 Número de casais, ciclos de postura e ovos avaliados em cada grupo experimental. ... 51
15
Resumo
Os triatomíneos são vetores do , causador da doença de Chagas, em várias regiões das Américas. A capacidade vetorial desses insetos está diretamente relacionada a sua performance alimentar, que por sua vez, é afetada pela fisiologia do hospedeiro vertebrado e por características do aparato alimentar do inseto. Além disto, os triatomíneos possuem moléculas bioativas salivares e intestinais que interferem na hemostasia e nas respostas infamatória e imune do hospedeiro, auxiliando no repasto sanguíneo. A técnica de RNA interferente (RNAi) tem possibilitado a realização de estudos funcionais de genes em organismos não modelo, incluindo os insetos vetores de doenças. Muitos dos genes alvo são de importância na alimentação dos insetos ou sua interação com os hospedeiros. Recentemente, o nosso grupo demonstrou a existência do RNAi parental em
16
Abstract
A variety of triatominae species are vectors of , causative agent of Chagas disease endemic in many regions throughout the Americas. The vectorial capacity of these insects is directly related to their feeding efficiency, which in turn depends on factors related to the host physiology and characteristics of the insect feeding apparatus. Furthermore, triatomine bugs possess bioactive salivary and intestinal compounds which interfere with the host inflamatory and immune responses, aiding the insects during their blood meal. The RNA interference technique has made possible the studies of gene funtion in non model organisms, such as disease vector insects. Many of the target genes in these studies are related to the feeding process in hematophagous insects. Recently, our research group has shown the occurrence of parental RNAi in Thus, trying to optimize RNAi application in triatomine gene function studies, the present work aims to standardising the parental RNAi by using salivary nitrophorins 1 4 (NPs 1 4) as markers. To this end, couples obtained from insects injected during different post embrionic developmental stages with double strand RNA specific to the nitrophorins genes (dsRNA NPs) or keratin (dsRNA QR, inespecific control) were monitored and their offspring phenotype (1st instar – N1) evaluated by estimation of salivary hemeprotein contents. A single dose injection of 10 µg dsRNA NPs in adults or fifth instar nymphs (before or after a blood meal) resulted in N1 nymphs presenting the knockdown phenotype from the first oviposition. However, fourth instar nymphs under the same treatment during fasting or after a blood meal only begun the nitrophorin knockdown from the fourth and third oviposition, respectively. Control group insects injected with dsRNA QR did not produce N1 with reduced salivary haemprotein (knockdown) during the evaluated period. Our second goal was to assess the possibility of serum albumin (BSA) stabilizing and aiding the nitric oxide activity, compound carried and released by nitrophorins. Salivary glands extract (EGS) from normal or knockdown insects homogenised in PBS (pH 7,4) with or without BSA (28 nM or 90% inactivated serum) were used in assays dorsal vessel. Only EGS of normal insects with BSA were able to stop the dorsal vessel contraction. Based on these results, we propose a model that demonstrates BSA and NPs interactions modulating the biological activities of
17
Introdução
1.1 – Biologia e importância dos triatomíneos
Triatomíneos são insetos da ordem Hemiptera, família Reduviidae, subfamília Triatominae e atualmente seu grupo tem reconhecidas 142 espécies agrupadas em 15 gêneros (Schofield & Galvão, 2009). Os triatomíneos são insetos hematófagos obrigatórios em todos seus estádios evolutivos pós embrionários e necessitam 20 a 40 minutos para completar seu repasto sanguíneo em vertebrados, principalmente aves e mamíferos. Seu contato com os hospedeiros ocorre apenas durante a hematofagia.
A importância médica dos triatomíneos decorre, principalmente, de serem vetores do , agente etiológico da doença de Chagas nas Américas. No Brasil, considerando se os últimos 50 anos, as espécies que tiveram importância
epidemiológica são Klug, 1834, Burmeister,
1835, Neiva, 1911, Corrêa & Espínola, 1964.
A espécie Stal, 1859 (Fig. 1) além de ser um dos principais vetores de na América do Sul (Venezuela e Colômbia), onde pode ser encontrado na copa de palmeiras ou estabelecido em domicílio (Lent & Wygodzinsky, 1979), é também importante em alguns países da América Central (Honduras, Guatemala, El Salvador), onde apresenta hábitos domiciliares (Schofield, 1994). Além de ser uma espécie amplamente estudada devido à sua importância epidemiológica,
18
Fig. 1. Ciclo biológico de desenvolvimento de
1.2 – Hematofagia e capacidade vetorial dos triatomíneos
De maneira geral, o tamanho da população de triatomíneos dentro do domicílio humano está relacionado com o número de hospedeiros disponíveis. Entretanto, o status nutricional da população depende do número de insetos por hospedeiro. Já foi demonstrado que a quantidade média de sangue ingerido por e
em hospedeiro não anestesiado é inversamente proporcional à densidade populacional do inseto (Schofield, 1980; Piesman et al., 1983). Esta diminuição provavelmente decorre do aumento da percepção dos insetos pelo hospedeiro devido à irritação causada pelas picadas. Schofield et al. (1985), utilizando voluntários humanos, observaram que a picada de ninfas de 2º estádio de é perfeitamente perceptível e esta percepção é maior no momento da penetração das peças bucais do que na fase posterior de ingurgitamento. A redução no tamanho médio do repasto com aumento de densidade é menor em do que em , e esta melhor exploração dos recursos alimentares explicaria o fato da primeira espécie desenvolver maiores densidades dentro das habitações humanas (Pereira et al., 1995; 2006).
Uma outra implicação da melhor exploração do recurso alimentar pelos triatomíneos refere se à dinâmica de dejeções e, consequentemente, a possibilidade de transmissão do para os humanos. Segundo Trumper e Gorla (1991), o momento
19
da dejeção não só depende da espécie de triatomíneo, como também da quantidade de sangue ingerida, pois, os triatomíneos que fazem repastos sanguíneos maiores, tendem a defecar mais rapidamente do que aqueles que fazem repastos menores, favorecendo assim a transmissão do . via dejeções contaminadas. Neste sentido, as duas principais espécies vetoras de . para humanos nas Américas (Schofield 1994), . e . estão entre as que apresentam maiores taxas de ingestão de sangue (Guarneri et al. 2000, 2003; Sant'Anna et al. 2001).
1.3 – Ação de biomoléculas salivares no processo alimentar dos triatomíneos
Os vertebrados possuem sofisticados sistemas hemostáticos que visam minimizar a perda de sangue. Entretanto, os artrópodes hematófagos apresentam um amplo repertório de proteínas com atividades anti hemostáticas como anticoagulantes, inibidores de agregação plaquetária e vasodilatadores, capazes de facilitar a ingestão do sangue de hospedeiros vertebrados (Ribeiro & Francischetti 2003; Ribeiro 1995).
Os triatomíneos são artrópodes solenofágicos, retirando o sangue diretamente dos vasos sanguíneos (Lavoipierre et al., 1959), sendo que a liberação da saliva ocorre durante todo o processo alimentar do inseto na pele do hospedeiro (Soares et al., 2006).
20
destas substâncias que têm sua função dividida entre auxiliar o triatomíneo a localizar um vaso e prevenir interrupções no fluxo sanguíneo, outros compostos com atividade imunossupressora (Kalvachova et al., 1999), anticomplemento (Cavalcante et al., 2003) e outras podem auxiliar no processo de alimentação, reduzindo a resposta inflamatória do hospedeiro.
As espécies de triatomíneos estudadas até o momento apresentaram diferenças quantitativas e qualitativas nas suas biomoléculas salivares. Enquanto as proteínas salivares mais abundantes em são as nitroforinas, em as mesmas não são observadas (Champagne et al., 1995). Além disto, estudos comparativos sobre as atividades anticoagulante, vasodilatadora e da enzima apirase presentes na saliva tem demonstrado diferenças quantitativas e qualitativas entre as espécies de tritomíneos (Pereira et al., 1996; Ribeiro et al., 1998).
1.4 – Histórico das nitroforinas salivares
21
Ribeiro et al. (1990) descreveram uma atividade vasodilatadora em
provavelmente devido a substâncias presentes na saliva destes insetos capazes de liberar o gás óxido nítrico na corrente sanguínea dos seus hospedeiros. Posteriormente foi demonstrado que o óxido nítrico é associado às hemeproteínas salivares (nitroforinas), ligando se reversivelmente a elas dependendo da concentração e do pH. O óxido nítrico é liberado pelas nitroforinas sob diluição do meio e em pH neutro ou alcalino, permanecendo ligado a elas em pH ácido (Ribeiro et al., 1993). A confirmação de que o óxido nítrico era produzido nas glândulas salivares de veio com o encontro de atividade característica da enzima óxido nítrico sintase nas glândulas salivares deste triatomíneo. Nussensveig et al. (1995) concluíram que o óxido nítrico produzido intracelularmente difunde se através de vesículas lipídicas até o lúmen da glândula, onde aí se liga às nitroforinas.
A confirmação de que as nitroforinas eram responsáveis pela atividade anti histamínica observada nos homogenatos salivares de veio com o trabalho realizado por Ribeiro & Walker (1994), onde os autores mostraram que a histamina e outros compostos imidazólicos ligam se fortemente as nitroforinas, sendo que essa ligação desloca o óxido nítrico. A histamina pode ser encontrada no sítio da picada de um inseto hematófago, sendo liberada por mastócitos previamente sensibilizados por antígenos salivares causando rubor, aumento de temperatura e coceira no local da picada. A neutralização da ação da histamina no sítio de alimentação do inseto é de fundamental importância, constituindo em uma vantagem em prol do inseto no sentido de facilitar sua alimentação sangüínea.
22
nitroforinas contém o átomo de ferro em estado de oxidação 3 Fe III onde a ligação ao óxido nítrico torna se reversível (Kd~10 6 M). Nas demais moléculas de globina onde o átomo de ferro é encontrado em estado ferroso (Fe II), a ligação do grupamento heme ao óxido nítrico é irreversível (Kd~1012 M) (Andersen et al., 1998). A origem da reversibilidade da ligação das nitroforinas ao óxido nítrico vem da presença do Fe+3 no grupamento heme que lhes confere resistência à auto redução pela ligação do óxido nítrico (Andersen et al., 1998). Dentre as diversas nitroforinas encontradas na saliva de NP1, NP2, NP3 e NP4, foram nomeadas de acordo com sua abundância na glândula salivar. As propriedades físicas, seqüência de aminoácidos, seqüência amino terminal e de cDNA das 4 moléculas foram determinadas, sendo que NP1 e NP4 possuem 90% de identidade em suas seqüências. Já NP2 e NP3 apresentam entre si 80% de identidade. Por outro lado, o grupo NP1 4 possui apenas 42% de identidade com o grupo NP 2 3 (Champagne et al., 1995).
O encontro de uma nitroforina semelhante à de em
foi pioneiramente descrito por Valenzuela et al. (1995), uma vez que as glândulas salivares destes cimicídeos possuíam a tonalidade laranja avermelhada, sugerindo a presença de hemeproteínas. Foi verificada a presença de uma nitrosil hemeproteína que também possui Fe+3 ligado ao heme e que também libera óxido nítrico dependendo da concentração e do pH do meio. No entanto, suas propriedades espectroscópicas e o seu tamanho (32 kDa comparada com as proteínas de 19 20 kDa de ) sugerem que a nitroforina de possui uma origem diferente das nitroforinas de
(Valenzuela & Ribeiro, 1998).
23
triatomíneo. A purificação do anticoagulante salivar de . revelou ser esta molécula uma nitroforina (NP2). A atividade antifator VIII realizada por essa molécula pode estar localizada em uma pequena porção da molécula não relacionada ao sítio de ligação ao óxido nítrico, uma vez que as outras 3 nitroforinas não apresentam atividade anticoagulante, apesar de apresentarem a mesma função de carreamento de óxido nítrico. A existência de proteínas com múltiplas funções provavelmente foi uma característica selecionada durante a evolução destes insetos (Ribeiro et al., 1995).
Yuda et al. (1997) produziram uma prolixina S (NP2) recombinante, realizando algumas caracterizações da mesma como carreadora de óxido nítrico. Esta nitroforina mostrou forte atividade vasodilatadora dose dependente em bio ensaios utilizando se musculatura lisa aórtica. O óxido nítrico é gradualmente liberado por esta nitrosil hemeproteína em pH 7,0 e 37oC (situação fisiológica encontrada no hospedeiro vertebrado). Entretanto, à temperatura de 20oC e pH 6 (situação encontrada na glândula salivar do inseto), a sua liberação torna se muito lenta. Esta característica possibilita à NP2 agir como reservatório de óxido nítrico na glândula salivar, liberando o prontamente quando esta proteína carreadora entra em contato com a corrente sangüínea.
24
Zhang et al. (1998), trabalhando com a NP2 isolada de glândulas salivares de . mostraram que esta proteína atua como um inibidor específico da ativação do complexo Fator Xase da via intrínseca da coagulação. NP2 foi capaz de inibir a ativação do fator Xase na presença de fosfolípides, de fator VIII, mas não na ausência destes dois fatores. Esses resultados sugerem que a NP2 interage mais favoravelmente ao fator IXa quando o mesmo se encontra ligado ao complexo fator VIIIa/superfície de membrana/fatorX. Posteriormente, estudos realizados por Isawa et al. (2000) demonstraram que a NP2 liga se diretamente aos fatores IX e IXa na presença de íons Ca2+, inibindo tanto a ação catalítica do fator XIa sobre o fator XI quanto a ativação do complexo Xase da via intrínseca pela interação do fator IXa com o fator VIIIa e os fosfolípedes de membrana. Além disso, a NP2 também inibiu a ativação do fator IX em IXa pela inibição da ligação do fator IX com o fator VIIa complexado com fator tissular da coagulação (fator III), demonstrando que essa proteína também interfere na via extrínseca da coagulação.
25
pequenos metabólitos (Flower, 2000). Várias lipocalinas com atividade anti hemostática já foram descritas na saliva de artrópodes hematófagos (Montfort et al., 2000, Ribeiro et al., 2004; Santos et al., 2007; Konnai et al., 2011).
O estudo da estrutura cristalina da NP2 (Andersen & Montfort, 2000) veio confirmar que as nitroforinas possuem semelhança estrutural com as lipocalinas, caracterizada por essas 8 folhas pregueadas β antiparalelas que envolvem uma ampla cavidade onde se situa o único grupamento heme que se liga à proteína através do contato com dez resíduos de aminoácidos hidrofóbicos, além da coordenação da cadeia lateral da histidina proximal (His 59) com o átomo de ferro. Além disso, esses autores demonstraram que apenas a NP2 recombinante apresenta atividade anticoagulante.
As nitroforinas apresentam diferenças na quantidade e momento de sua expressão nas glândulas salivares de (Moreira et al., 2003). As NPs 5 e 6, mais recentemente descritas, aparentam ser encontradas mais abundantemente nos estádios iniciais de desenvolvimento do inseto, enquanto poucas quantidades de NP7 só foram encontradas durante os estádios finais de desenvolvimento (Andersen et al., 2004). As nitroforinas 5 7 são consideravelmente menos abundantes e menos estudadas que as NPs 1 4. Sabe se que NP 7 é capaz de se ligar à membrana negativamente carregada de plaquetas, o que as NPs 1 4 não são capazes de fazer (Andersen et al., 2005), agindo assim como um anticoagulante através de atividade inibidora de prototrombinase.
1.5 – RNA interferente
O termo RNA interferente (RNAi) foi primeiramente usado por Fire et al. (1998) na descrição de um mecanismo de inativação gênica em células de
26
alvo e facilidade de aplicação utilizando se organismos inteiros, ou usando se tecidos em cultura.
O silenciamento gênico ocorre em resposta à introdução de RNA dupla fita (dsRNA) homólogo ao gene alvo nas células, onde o dsRNA é clivado em fragmentos menores (21 25 nucletídeos) pela enzima . Os fragmentos gerados,
! (siRNAs), se associam a proteínas celulares formando um complexo protéico multimérico denominado RISC, que contém dentre outras proteínas uma helicase, responsável pela separação das fitas dos fragmentos de dsRNA. Uma das fitas é descartada, permanecendo a fita antisenso associada ao complexo RISC, onde servirá como molde para o alvo de mRNA. Enzimas da família Argonauta presentes no complexo RISC desempenham os papéis de se ligar à fita molde e clivar o mRNA alvo.
A técnica de silenciamento gênico pela tecnologia de RNA interferente (RNAi) já teve seu uso adaptado para diversas espécies de insetos pertencentes a diferentes ordens (Belles, 2010), incluindo (St Johnston, 2002), ! (Bladin et al., 2002), " (Sant'Anna et al., 2008), ! (Drake et al., 2012).
1.6 – Efeito sistêmico e longa duração do silenciamento por RNAi
27
introduzido. As propriedades sitêmicas também evidenciadas nesse nematódeo dizem respeito à expansão célula a célula e tecido a tecido dos efeitos de RNAi no organismo inteiro. A ocorrência desse processo em depende do gene Sid 1, que codifica uma proteína com domínios trans membrana previstos expressa em células sensíveis ao RNAi e parece atuar como um canal para o dsRNA (Winston et al., 2002). O RNAi parental é uma extrapolação das propriedades sistêmicas do fenômeno, que ocorre quando o dsRNA ou seus subprodutos pocessados alcançam as células da linhagem germinativa e os efeitos de # # $ são passados para os embriões/indivíduos da geração seguinte.
Embora o mecanismo central do RNAi aparente ser similar e bem conservado em diversos organismos, algumas proteínas e mecanismos relacionados podem diferir ou estar ausentes em certas espécies (Terenius et al., 2010). Apesar da aparente ausência de homólogos dessas duas moléculas (Sid 1, RdRP) em insetos, um intenso e duradouro efeito de silenciamento já foi verificado em várias espécies, dentre elas
(mais de dois meses de persistência dos efeitos de RNAi), !
(mais de 21 dias) (Amdam et al., 2003), no gafanhoto % (10 e 14 dias) (Dong & Friedrich, 2005) e na traça (ao menos um mês) (Uryu et al., 2013). Sugere se que alguma amplificação e/ou manutenção do mecanismo ocorra em insetos (Tomoyasu & Dennel, 2004).
A eficiência do mecanismo de RNAi em insetos varia grandemente, não apenas entre diferentes espécies evolutivamente próximas, mas também entre diferentes tecidos de um mesmo inseto (Ren et al., 2014). A ocorrência de RNAi parental em
28
estar relacionadas à essa diferente susceptibilidade à entrada de dsRNA e manutenção dos níveis de silenciamento nos tecidos que posteriormente formam os ovos.
1.7 – Silenciamento das nitroforinas salivares de por RNAi
A utilização do RNAi em triatomíneos foi iniciada pelo nosso grupo de pesquisa com objetivo de avaliar a influência de genes salivares e intestinais no processo alimentar destes insetos (Araújo et al., 2006; 2007; 2009; Paim et al., 2011; 2012).
29
30
2 – Justificativa
A maior parte da população brasileira vive sob risco de transmissão de doenças parasitárias de alta prevalência, como a doença de Chagas, a malária e as leishmanioses. Os agentes etiológicos dessas doenças são transmitidos aos seres humanos através de insetos hematófagos, como os triatomíneos, mosquitos e flebotomíneos.
Com o advento da genômica, a entomologia molecular tem passado por uma grande mudança, o que tem conduzido a uma reestruturação dessa área de estudo. Atualmente, o número de sequências gênicas de insetos de importância médica depositado nos bancos de dados vem aumentando consideravelmente. Com relação aos triatomíneos já estão disponíveis os sialomas (transcriptomas e proteomas de glândulas salivares, obtidos a partir do sequenciamento em larga escala de bibliotecas de cDNA e proteínas) de diversas espécies. No caso de , o projeto genoma já encontra se em fase de anotação, com quase sete milhões de sequências WGS (Whole Genome Shotgun), disponibilizadas no NCBI e em http://rprolixus.vectorbase.org/
31
hematofagia. Entretanto, o curto período de meia vida dessa molécula, uma vez liberada nos tecidos, sugere a participação de outras proteínas em seus efeitos biológicos. Nesse contexto, buscamos avaliar a interação do óxido nítrico liberado por hemeproteínas salivares de com proteínas abundantes do hospedeiro presentes no local da picada.
32
3 – Objetivos
3.1 – Objetivo geral
Avaliar a transmissão vertical do silenciamento induzido por RNA interferente (RNAi parental) de utilizando as nitroforinas salivares como marcadores e estudar a possível interação destas hemeproteínas com a soro albumina do hospedeiro na persistência das atividades biológicas induzidas pelo NO no sítio de alimentação do inseto.
3.2 – Objetivos específicos
a) Avaliar o fenótipo de ninfas de 1º estádio provenientes de adultos injetados com dsRNA para nitroforinas (1 4) salivares em diferentes fases de desenvolvimento pós embrionário;
33
4 – Materiais e métodos
4.1 – Manutenção dos insetos
Os insetos da espécie (Honduras) utilizados neste trabalho foram mantidos sob condições semicontroladas de temperatura e umidade (28 ± 2°C e 65 ± 10% UR), com fotoperíodo de 12 horas claro/escuro, no insetário do Laboratório de Fisiologia de Insetos Hematófagos do Departamento de Parasitologia (ICB/UFMG). A alimentação dos insetos foi realizada semanalmente em ratos, hamsters ou camundongos previamente anestesiados.
4.2 – Obtenção das amostras de glândulas salivares
Para a coleta das glândulas salivares, os triatomíneos foram dissecados na presença de solução salina 0,9% em lupa estereoscópica, com o auxílio de pinças e micro tesoura (Fig. 2). Após a coleta, as glândulas salivares foram processadas para a obtenção de amostras de extratos de glândula salivar (EGS) apropriadas, conforme o especificado na descrição dos diferentes ensaios realizados no presente trabalho.
Fig. 2. Aspecto das glândulas salivares de no tórax do inseto (aumentado 20x).
34
4.3 – Obtenção e conservação do soro e inativação do complemento
O sangue de pelo menos seis voluntários saudáveis foi coletado através de punção venosa para a obtenção de soro. Após a coleta de 10 mL de sangue de cada voluntário, os tubos de ensaio contendo o material foram deixados descansando a temperatura ambiente por um período de três horas. Após esse tempo, o sangue já estava totalmente coagulado e os tubos foram centrifugados a 500 g por dez minutos. O soro coletado de cada tubo de ensaio foi misturado com os demais para formar um pool de soros humanos. Esse material foi aliquotado em frações de 300 µL e congelado a – 80ºC em freezer até o momento do uso.
Para inativação do sistema do complemento, os tubos contendo soro retirados do freezer foram mantidos em banho maria a 56°C durante 30 minutos antes de ser utilizados nos ensaios (Mendes Souza, 2013).
4.4 – Genes das nitroforinas salivares (NPs) 1 a 4
Neste trabalho foram utilizados como alvos do RNAi os genes codificantes das nitroforinas NP1 (GenBank: L39654.1), NP2 (GenBank: U70582.1), NP3 (GenBank: U61143.1), NP4 (GenBank: U70584.1), moléculas multifuncionais abundantes nas glândulas salivares de (Champagne et al. 1995).
4.5 – Extração de RNA, síntese de cDNA e PCR
35
36
4.6 – Silenciamento por RNAi – síntese e introdução do RNA de fita dupla
(dsRNA) nos insetos
As moléculas de dsRNA utilizadas neste trabalho (nitroforinas 1 4) foram sintetizadas separadamente utilizando o Kit MegaScript T7 (Ambion) de acordo com as recomendações do fabricante. Os produtos de PCR continham o promotor T7 (5’ taatacgactcactatagggaga 3’) em ambas as extremidades (Tabela 1). A síntese do dsRNA ocorreu com incubação de 37°C por 14 horas. Após esse período, o dsRNA foi tratado com 1 µL de DNAse RQ1 (livre de RNAse; Ambion) por reação (a cada 20 µL de dsRNA). O volume do RNA dupla fita recém sintetizado e tratado foi então triplicado com água milliQ livre de RNAse, precipitado com isopropanol 100% e depois lavado com etanol 70%. O dsRNA foi ressuspendido com água milliQ e quantificado em triplicata, por espectrofotometria no comprimento de onda de 260 nm. Para a quantificação, foi utilizado 0,5 µL do dsRNA, diluído 1:200 em água no volume final de 100 µL. A qualidade das fitas foi avaliada por eletroforese em gel de agarose a 1,5%, onde foi utilizado 0,5 µL do dsRNA, agora diluído 1:40 em água. Após a quantificação o dsRNA foi secado em centrífuga evaporadora e os quatro dsRNAs redissolvidos em conjunto em solução salina 0,9% na concentração de 10 µg/µL (2,5 µg/µL para cada dsRNA NP).
37
experimentais foram separados em grupos controle (injetados com dsRNA queratina) e teste (silenciados para os genes alvo) com média de idade e peso semelhantes entre os grupos.
Para os experimentos de RNAi parental, utilizou se ninfas injetadas durante o quarto e quinto estádios ninfais e estágio adulto. Já para os experimentos de atividade biológica das nitroforinas sobre o vaso dorsal isolado, utilizou se insetos injetados durante o terceiro estádio ninfal.
A avalição do fenótipo de silenciamento das nitroforinas salivares foi realizada em insetos injetados com dsRNA específico (NP 1a 4) após atingirem o quinto estádio ninfal ou o estágio adulto, pela observação visual das glândulas salivares. Já a caracterização da prole dos casais experimentais foi realizada por espectrofotometria (item 4.7) a partir das glândulas salivares de ninfas de primeiro estádio.
Tabela 1. Sequencias dos iniciadores utilizados nos experimentos de PCR (molde para a síntese de dsRNA).
Gene alvo Iniciador : 5’→3’ Iniciador : 5’→3’
Amplicon (pares de base – pb)
NP1 T7 + tttgctgcagtgggtgtaag T7 + agttgcccgacgttacatct 334 pb
38
4.7 – Estimativa de proteína total e hemeproteínas no extrato de glândula salivar
Para a estimativa de hemeproteínas e da quantidade total de proteínas, as amostras foram formadas por conjuntos (pools) de três pares de glândula salivar cada em tubos de microcentrífuga contendo solução salina (NaCl 0,9%). Logo após a coleta das glândulas, o material foi sonicado (42 KHz, – Bransonic® 1510R MT, Branson) por 1 minuto e centrifugado a 12000g a 4°C. O sobrenadante obtido foi transferido para um tubo e armazenado a 20°C até a realização dos ensaios.
39
Fig. 3. Estimativa de hemeproteínas e proteína total na glândula salivar de adultos de injetados com dsRNA para NPs 1 4 ou queratina durante o quarto estádio por espectrofotometria. As setas indicam os picos característicos relativos à proteínas (280 nm) e hemeproteínas (404 nm).
0 0,5 1 1,5 2 2,5 3 3,5
230 260 290 320 350 380 410
40
4.8 – Avaliação do RNAi parental
A fim de averiguar qual é o melhor momento de apresentação do dsRNA para a obtenção do fenótipo de silenciamento na prole, foram formados cinco grupos experimentais (cada grupo contendo subgrupos controle e teste) com insetos injetados em diferentes idades e estados fisiológicos (Fig. 4).
As ninfas e adultos injetados com aproximadamente 4 dias de jejum foram alimentados três dias após a inoculação do dsRNA. Já as ninfas injetadas após o repasto, somente foram alimentadas aproximadamente 7 dias após mudarem para o próximo estádio/estágio.
Após a aplicação do dsRNA (NPs 1 4 ou queratina), os insetos foram separados individualmente e alimentados periodicamente até atingir o estágio adulto, quando então foi realizada a formação dos casais experimentais em potes separados e devidamente identificados. Os casais foram alimentados a cada 15 dias e os ovos produzidos por casal após cada repasto foram recolhidos e transferidos para um novo pote.
41
Dias 0 4 7 21 28 44 51 66 81 96 101 116 (120)
Dias 0 7 17 21 28 44 51 66 81 96 (100)
Dias 0 4 7 23 30 44 59 74 (80)
Dias 0 7 18 23 30 45 (50)
Dias 0 4 6 21 (30)
A
B
C
D
E
42
4.9 – Preparação e monitoramento do vaso dorsal dos insetos
Para realização dos experimentos da atividade biológica dos extratos de glândula salivar (EGS) sobre o vaso dorsal de utilizou se metodologia semelhante à descrita por Lee & Lange (2011). Em resumo, a cutícula abdominal ventral (do terceiro ao sexto segmento) e os sistemas digestivos e reprodutores dos insetos adultos foram removidos sob microscópio estereoscópico. O vaso dorsal e diafragma dorsal foram mantidos intactos ligados à cutícula e lavados repetidamente com salina 0,9 %. Cada inseto foi preso a uma placa de Petri com fita adesiva dupla face e 40 µL de salina foram adicionados sobre o vaso.
43
Fig. 5. Montagem do aparato para experimentos no vaso dorsal de adulto de A seta indica o vaso dorsal observado ao microscópio em aumento de 20x.
4.10 – Atividade biológica do EGS e a influência da soro albumina
Para avaliar a atividade biológica do EGS de , bem como, do possível sinergismo desempenhado pela soro albumina bovina (BSA), escolhemos utilizar a preparação de vaso dorsal de inseto. Já que, o nosso grupo demonstrou que a mesma fração semi purificada da saliva de apresenta tanto uma ação paralisante sobre o vaso dorsal de inseto como inibidora do potencial de ação composto (PAC) em nervo ciático isolado de rato (Alves, 2011; Alves et al., 2011). Além disto, boa parte das atividades presentes na saliva de é devida a liberação de óxido nítrico no seu sítio de alimentação pelas nitroforinas salivares (Moncada et al., 1991; Ribeiro et al., 1993; Andersen et al., 2005) e tem sido demonstrado que moléculas doadoras de óxido nítrico apresentam atividade sobre o PAC de terminações nervosas (Renganathan, 2002), bem como sobre vaso dorsal de inseto (Silva et al., 2012).
44
ninfas de quinto estádio injetadas durante o terceiro estádio com dsRNA das nitroforinas (silenciadas), dsRNA da queratina ou não injetadas.
As soluções utilizadas nos ensaios foram tampão fosfato salino (PBS – NaCl 137 mM, KCl 2,7 mM, Na2HPO4.2H2O 9,1 mM, KH2PO4 1,8 mM) a pH 7,4, soro humano inativado (item 4.3) diluído (9:1 v/v) em PBS a pH 7,4 e albumina sérica bovina (BSA) a 20 mg/ mL em PBS a pH 7,4. O soro foi inativado para o complemento antes dos experimentos a fim de descartar a participação do dano causado por este mecanismo na interrupção das contrações do vaso.
Antes da aplicação das amostras, o funcionamento do vaso dorsal era monitorado durante três minutos. O volume total de líquido que banhava a preparação era mantido em 40 µL, assim, antes da aplicação dos 10 µL de amostra, 10 µL de salina era retirado. Após a aplicação de cada amostra, o funcionamento do vaso era registrado também durante três minutos. Entre as aplicações das amostras, a preparação foi lavada pelo menos três vezes com salina 0,9%. Nos ensaios em que houve a paralização total da atividade do vaso dorsal, um período adicional de observação de três minutos foi realizado antes da lavagem. A sequência de aplicação das diferentes amostras realizada nas preparações de vaso dorsal encontra se na fig. 6.
Preparo do inseto Lavagem Estabilização
Aplicação: PBS + EGS 1º Controle
Aplicação: Solução (soro
ou BSA) 2º Controle
Aplicação: Solução + EGS
Teste
Recuperação dos movimentos
Gravação Gravação Gravação Gravação Gravação
Lavagem Lavagem Lavagem Lavagem
45
4.11 – Análise de imagem
As imagens dos vídeos (25 FPS) produzidos durante os ensaios de vaso dorsal isolado foram analisados utilizando o software ImageJ (versão 1.46 http://rsb.info.nih.gov/ij/) adaptando metodologia utilizada por Alves et al. (2011).
Através de ferramentas do programa selecionou se em cada quadro do vídeo uma fração da área da luz do vaso dorsal do inseto. Os pixels dentro da área selecionada de cada são convertidos em imagens preto e branco, onde o preto representa as paredes do vaso dorsal, visíveis dentro da área delimitada apenas durante cada contração, e o branco representa a luz do vaso sem aproximação entre as paredes laterais (Fig. 7). O programa calcula, para cada , a proporção de pixels pretos/total de pixels e os valores dessa proporção são utilizados na construção de um gráfico do perfil de contrações do vaso ao longo do tempo (Fig. 8).
A B
46
Fig. 8. Perfil de contrações do vaso dorsal isolado de adulto de a partir da análise das gravações em vídeo analisadas no ImageJ. Cada pico da imagem representa o fechamento das paredes do vaso na área da luz do vaso selecionada.
4.12 – Análise estatística
47
5 – Resultados
5.1 – Avaliação da quantidade de proteínas salivares nos insetos
5.1.1 – Avaliação da quantidade de proteína total e hemeproteínas
O conteúdo salivar de insetos normais (não injetados) pertencentes a todos os estágios de desenvolvimento pós embrionário presentes na colônia foi avaliado quanto aos valores de absorbância no espectrofotômetro a 404 nm e à concentração total de proteínas (Figs. 9, 10).
48
Fig. 10. Valores (expressos em média ± desvio padrão) da concentração de proteína total presentes no extrato salivar para os estágios de desenvolvimento pós embrionário de obtidos a partir de pools de 3 pares de glândulas.
49
Fig. 11. Correlação entre a média estimada dos valores de absorbância a 404 nm e concentração de proteína total da saliva para os estágios de desenvolvimento pós
embrionário de .
5.1.2 – Avaliação do fenótipo e produção de ovos nos insetos injetados
50
Os dados referentes ao número de casais, número de posturas e quantidade de ovos produzidos por cada casal dos diferentes grupos experimentais avaliados encontram se resumidos na Tabela 2. A diferença no número de casais avaliados (teste e controle) dos grupos deveu se, basicamente, à morte dos insetos durante o desenvolvimento até o estágio adulto e ao número desproporcional de machos e fêmeas obtidos. Entretanto, a média de ovos por postura produzidos pelos grupos teste (18,8 0,7) foi semelhante ao obtido pelo grupo controle (17,5 1,3) (teste , ns).
Fig. 12. Aspecto típico apresentado pelas glândulas salivares de adultos de
em microscópio estereoscópico (aumento de 20x). (A) Glândulas salivares de um inseto silenciado para as NPs 1 4. (B) Glândulas salivares de um inseto injetado com dsRNA queratina (controle inespecífico). Neste exemplo o dsRNA foi administrado via injeção única durante o quarto estádio ninfal em jejum.
A
51
Tabela 2. Número de casais, ciclos de postura e ovos avaliados em cada grupo experimental.
Grupo insetos N° de casais avaliados
Ciclos de postura avaliados
N° de casais que ovipuseram em
cada ciclo de postura
Nº total de ovos por ciclo de postura (média de
ovos por postura)
N4 jejum Teste Controle
4 2
6 4
4 4 3 4 2 1 2 2 1 2 0 0
64 81 56 74 32 14 (17,8) 40 13 29 36 0 0 (17,1) N4 alimentados Teste Controle 1 1 4 4
2 2 2 1 1 1 1 1
33 44 39 16 (18,9) 13 17 25 14 (18,7)
N5 jejum Teste Controle
4 2
4 4
4 4 3 2 2 1 1 1
94 67 42 34 (18,2) 16 17 39 21 (18,6)
N5 alimentados Teste Controle 2 1 2 2 2 1 1 1
43 16 (19,7) 16 12 18 16 (15,5) Adultos Teste
Controle 3 2 2 2 3 3 2 2
57 58 (19,2) 35 35 (17,5)
5.1.3 – Avaliação da transmissão do silenciamento para a prole dos insetos
injetados
52
médios de absorbância a 404 nm no extrato de glândula salivar de ninfas descendentes em todos os insetos silenciados na fase adulta e nos dois grupos silenciados durante o quinto estádio quando comparadas às ninfas descendentes de insetos injetados com dsRNA QR nos mesmos períodos (Tabela 3). Tal diminuição só foi observada a partir do quarto ciclo de postura para as ninfas descendentes do grupo de insetos injetado durante o quarto estádio ninfal em jejum. Na prole dos insetos injetados durante o quarto estádio após alimentação, houve diferença na expressão de hemeproteínas salivares entre os grupos controle e teste a partir da segunda oviposição realizada. Entretanto, a concentração média estimada de proteína total presente no EGS de N1 foi semelhante entre os teste e respectivos controles em cada um dos grupos experimentais (Tabela 3).
53
Tabela 3. Estimativa média (± dp) da quantidade de hemeproteínas e de concentração de proteína total no extrato de glândula salivar de ninfas de primeiro estádio da prole dos diferentes grupos experimentais. Teste (*) ≤0,05; (**) ≤0,01; (***) ≤0,001.
Grupo insetos Ciclo de
postura Hemeproteína1 Absorbânc ia 404 (Teste/ Controle)
Proteína total (µg/µL)2
Proteína
total
(Teste/
Controle) Teste (n) Controle (n) Teste (n) Controle (n)
N4 jejum 1ª 2ª 3ª 4ª 5ª 6ª
0,077 ± 0,034 (10) 0,058 ± 0,03 (8) 0,079 ± 0,04 (8) 0,040 ± 0,01 (10)
0,045 ± 0,01 (6) 0,0243 (2)
0,089 ± 0,03 (6) 0,096 ± 0,03 (5) 0,090 ± 0,02 (5) 0,074 ± 0,04 (6) 0,085 ± 0,07 (22) 0,085 ± 0,07 (22)
0,87 0,60 0,89 0,54** 0,53***
0,075 ± 0,037 (10) 0,085 ± 0,028 (8) 0,096 ± 0,055 (8) 0,081 ± 0,031 (10)
0,090 ± 0,028 (6) 0,0803 (2)
0,078 ± 0,026 (6) 0,090 ± 0,035 (5) 0,101 ± 0,041 (5) 0,088 ± 0,025 (6) 0,090 ± 0,046 (22) 0,090 ± 0,046 (22)
0,96 0,91 0,95 0,92 1,00 N4 alimentados 1ª 2ª 3ª 4ª
0,066 ± 0,01 (4) 0,039 ± 0,01 (5) 0,036 ± 0,02 (5)
0,0443 (2)
0,097 ± 0,03 (5) 0,090 ± 0,02 (5) 0,087 ± 0,01 (6) 0,105 ± 0,04 (5)
0,68 0,43** 0,41**
0,054 ± 0,014 (4) 0,070 ± 0,055 (5) 0,041 ± 0,015 (5)
0,0463 (2)
0,081 ± 0,035 (5) 0,065 ± 0,031 (5) 0,058 ± 0,018 (6) 0,070 ± 0,030 (5)
0,67 1,08 0,71 N5 jejum 1ª
2ª 3ª 4ª
0,045 ± 0,02 (11) 0,053 ± 0,02 (11) 0,055 ± 0,03 (7) 0,038 ± 0,02 (6)
0,095 ± 0,02 (5) 0,089 ± 0,02 (6) 0,087 ± 0,01 (5) 0,081 ± 0,04 (3)
0,47** 0,60* 0,62* 0,46*
0,098 ± 0,028 (11) 0,106 ± 0,048 (11) 0,105 ± 0,025 (7) 0,101 ± 0,036 (6)
0,049 ± 0,026 (5) 0,070 ± 0,025 (6) 0,081 ± 0,031 (5) 0,085 ± 0,017 (3)
2,00 1,51 1,23 1,19 N5 alimentados 1ª 2ª
0,046 ± 0,01 (4) 0,0823 (2)
0,090 ± 0,03 (4) 0,088 ± 0,04 (5)
0,51* 0,056 ± 0,018 (4) 0,1333 (2)
0,069 ± 0,029 (4) 0,078 ± 0,033 (5)
0,81 Adultos 1ª
2ª
0,027 ± 0,01 (14) 0,033 ± 0,01 (12)
0,090 ± 0,03 (4) 0,086 ± 0,03 (5)
0,30*** 0,38***
0,074 ± 0,047 (14) 0,080 ± 0,032 (12)
0,055 ± 0,038 (4) 0,072 ± 0,024 (5)
54
Mesmo utilizando se amostras de EGS constituídas por grupos de 3 insetos, observou se uma alta dispersão nos valores de concentração de proteína total nos diferentes grupos avaliados (dp superior a 40% da média). Tal fato provavelmente explica a ausência de diferença estatística nas comparações realizadas. Nesse contexto, a fim de minimizar a possível influência da variação na quantidade de proteína total entre as amostras sobre a estimativa de hemeproteínas salivares, optamos por normalizar a absorbância a 404 nm obtida pela concentraçao de proteína estimada.
55
Fig. 14. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quarto estádio em jejum. Teste : (**) ≤0,01; (***) ≤0,001. (A e B) como não houve a 5ª e 6ª posturas do grupo controle, utilizamos todos os valores obtidos nas posturas anteriores.
***
***
56
Fig. 15. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quarto estádio após a alimentação. Teste : (*) ≤0,05.
57
Fig. 16. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quinto estádio em jejum. Teste (**) ≤0,01; (***) ≤0,001.
*** ***
58
Fig. 17. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o quinto estádio após a alimentação. Teste (*) ≤0,05.
Fig. 18. Valores da razão entre absorbância a 404 nm e concentração total de proteínas do conteúdo salivar para ninfas descendentes de insetos injetados durante o estágio adulto em jejum. Teste (***) ≤0,001.
*
59
5.2 – Ensaios em vaso dorsal de adulto de
5.2.1 – Ensaios de aplicação de EGS em soro humano inativado (n=5)
A aplicação de 10 µL das amostras de conteúdo salivar de ninfas de quinto estádio de diluído em soro humano a 90% em PBS pH 7,4 inativado para o complemento apresentou efeito paralisante (de 30” a 3’) sobre o vaso dorsal de insetos adultos em 100% dos experimentos, resultando em redução de sua frequência de contrações (Fig. 19). O efeito paralisante ocorreu logo após a aplicação da amostra (Fig. 20) e os vasos dorsais paralisados retomaram a atividade de contração após a lavagem com solução salina (Fig. 21). Contudo, as aplicações de 10 µL dos controles contendo apenas soro ou extrato de glândula salivar rompida em PBS não resultaram em parada ou alteração significativa no ritmo de contrações do vaso.
Fig. 19. Valores da frequência de contrações do vaso dorsal de adulto de
após aplicação de soro humano inativado para o complemento e EGS (teste), individualmente e na presença de soro. Valores comparados à frequência após período de estabilização dos insetos. Tukey: (*) ≤0,05; (**) ≤0,01; (***) ≤0,001.
*** **
60
Fig. 20. Perfil de contrações do vaso dorsal de exemplares adultos de nos momentos antes e logo após aplicação de amostra de extrato de glândula salivar + soro. A seta indica o momento da aplicação.
61
5.2.2 – Ensaios de aplicação de EGS em solução de BSA a 20 mg/mL (n=5)
A aplicação de 10 µL das amostras de conteúdo salivar de ninfas de quinto estádio de diluído em albumina a 20 mg/mL em PBS pH 7,4 apresentou efeito paralisante (de 35” a 3’) sobre o vaso dorsal de insetos adultos em 100% dos experimentos, resultando em redução de sua frequência de contrações (Fig. 22). Similar ao ocorrido nos experimentos utilizando de soro humano inativado, o efeito paralisante ocorreu logo após aplicação da amostra e com exceção de uma ocorrência, os vasos dorsais paralisados retomaram a atividade de contração após lavagem com solução salina. A aplicação dos controles contendo apenas solução de albumina ou extrato de glândula salivar rompida em PBS não resultaram em parada ou alteração significativa no ritmo de contrações do vaso.
Fig. 22.Valores da frequência de contrações do vaso dorsal de adulto de
após aplicação de BSA e EGS (teste), individualmente e em conjunto com BSA. Valores comparados à frequência após período de estabilização dos insetos. Tukey: (**)
≤0,01; (***) ≤0,001.
***
***
62
5.2.3 – Ensaios com EGS de insetos silenciados para as nitroforinas (n=5 dsRNAM
NPs; n=5 dsRNAMQR)
A aplicação de EGS em solução de BSA das glândulas de ninfas de quinto estádio de silenciadas para as NPs 1 4 não resultou em alteração significativa do ritmo de contrações do vaso dorsal dos insetos adultos (Fig. 23). A aplicação do extrato das glândulas de insetos injetados com dsRNA QR em solução de BSA resultou em paralisia (de 1’ a 3’) e consequente diminuição do ritmo de contrações (Fig. 24). A aplicação dos controles contendo apenas EGS dos dois grupos de insetos ou solução de BSA não resultou em paralisia e redução da frequencia de contrações dos vasos. Não houve retomada do funcionamento do vaso dorsal em dois dos insetos que receberam aplicação contendo saliva de ninfas injetadas com dsRNA queratina.
Fig. 23. Valores da frequência de contrações do vaso dorsal de adulto de
63
Fig. 24. Valores da frequência de contrações do vaso dorsal de adulto de
após aplicação de BSA e EGS de insetos injetados com dsRNA para queratina, individualmente e em conjunto com BSA. Valores comparados à frequência após período de estabilização dos insetos. Tuley: (**) ≤0,01.
Os valores das frequencias de contração e tempo de interrupção do funcionamento dos vasos dorsais de cada grupo de insetos tratados com as diferentes soluções encontram se resumidos no Anexo I.
64
6 – Discussão
6.1 – Transmissão vertical do silenciamento
Apesar de todos os artrópodes hematófagos enfrentarem problemas semelhantes durante a aquisição de sangue, tal capacidade evoluiu independentemente nos vários grupos. Existe, assim, uma grande variedade de moléculas salivares bioativas distintas mas com atividades biológicas convergentes (anti hemostáticas, anti inflamatória, imussupressora) facilitando o repasto sanguíneo nos hospedeiros vertebrados.
Os resultados dos sialomas publicados principalmente nos últimos 10 anos tem mostrado uma diversidade de moléculas de triatomíneos bem maior do que esperado para insetos pertencentes a mesma subfamília. Diferentemente do ocorrido no gênero , as moléculas encontradas em maior quantidade na saliva de
são nitroforinas (NPs), hemeproteínas responsáveis pela coloração avermelhada da glândula salivar. Sete delas (NP 1 7) já foram descritas na saliva deste inseto e compreendem cerca de 50% do conteúdo protéico salivar (Champagne et al., 1995; Moreira et al., 2003; Knipp et al., 2007).
A escolha das NPs como genes alvo para avaliação do RNAi parental deveu se ao fato do silenciamento destas hemeproteínas já ter sido padronizado em trabalhos anteriores do grupo, por serem expressas apenas na glândula salivar e por não afetar a integridade física dos insetos silenciados. A coloração clara do conteúdo salivar dos insetos silenciados para as NPs, fenótipo indicativo da redução na quantidade de nitroforinas, responsáveis pela coloração cereja das glândulas salivares (Araujo et al., 2006), foi outro aspecto importante na escolha das Nps para a padronização do RNAi
parental de .
65
silenciamento (glândulas salivares claras), evidenciando a persistência do efeito do RNAi por até 100 dias (fig. 4A). Tal achado corrobora o estudo recente de Paim et al. (2013) que publicaram o primeiro trabalho mostrando persistência prolongada dos efeitos do RNAi em podendo o silenciamento perdurar por mais de 7 meses após a introdução do dsRNA. Em e algumas plantas, o efeito prolongado do silenciamento é associado ao mecanismo de amplificação do RNAi mediado pela enzima RdRP (Price & Gatehouse, 2008). Entretanto, apesar da ausência de uma proteína homóloga ao RdRP no genoma de e nos demais insetos avaliados, os resultados apresentados acima sugerem fortemente a existência de algum mecanismo de amplificação de dsRNA/siRNA também em insetos.
Dois trabalhos recentes demonstraram a existência de RNAi parental em triatomíneos (Lavore et al., 2012 e Paim et al., 2013). Lavore et al. (2012) injetaram fêmeas adultas com dsRNA para o gene (Rp gt), envolvido na embriogênese, e avaliaram o fenótipo induzido pelo RNAi nos embriões. Paim et al. (2013), observaram a transmissão parental dos efeitos do RNAi para ninfas (segundo estádio) da geração F1 de adultos oriundos de ninfas de quinto estádio inoculadas com dsRNA (dos genes das NPs 1 4 ou para o gene da rhodinina), entretanto esta transmissão do silencamento não foi observada quando a injeção foi realizada em ninfas de terceiro estádio. Estes achados sugerem que a transmissão parental do silenciamento ocorra somente a partir de determinada fase de desenvolvimento gonadal do inseto, observação que nos levou a realizar os experimentos de avaliação do RNAi parental em diferentes fases de desenvolvimento.
66
(2008) sugeriu que Sid 1 não é essencial para os efeitos sistêmicos de RNAi em algumas espécies de inseto e que um mecanismo alternativo de introdução do dsRNA nas células provavelmente ocorra.
Em , os ovários contém sete ovaríolos do tipo telotrófico. A maior parte dos eventos de diferenciação das células dos ovaríolos e desenvolvimento do órgão reprodutor como um todo ocorre durante o quinto estádio ninfal após a alimentação (Lutz & Huebner, 1980). Os ovaríolos antes do repasto encontram se num estado de repouso, contendo células germinativas cercando uma região nutridora central e uma região basal de tecido pré folicular. Nos primeiros oito dias após o repasto, cada ovaríolo passa por uma fase proliferativa em que aumenta de tamanho através de sucessivas mitoses de suas células. Ainda não se observa diferenciação morfológica no tecido germinativo durante essa fase.
O surgimento de especialização regional no trofário marca a passagem para o estágio inicial de diferenciação dos ovaríolos, entre o 9º e 15º dias após a alimentação. O citoplasma das células da região basal do trofário se torna menos intensamente corado por azul de toluidina do que na região apical, além disso os núcleos das células basais não contém o nucléolo visível característico das células germinativas da região apical. Durante essa fase, a diferenciação das células também é visível por alterações na morfologia do núcleo, tornando as distinguiveis em três regiões distintas.
67
fase, vários oócitos dão início a um processo de crescimento pré vitelogênico preparando se para o término de seu desenvolvimento durante o estágio de vida adulto do inseto.
O fato da maturação das gônadas femininas de começar no quinto estádio após a alimentação poderia explicar os resultados obtidos por Lavore et al. (2012) e Paim et al. (2013), onde o RNAi parental só aconteceu quando a introdução do dsRNA foi realizada em ninfas de quinto estádio ou em adultos. Ou seja, quando os insetos fossem injetados no quinto estádio alimentado ou na fase adulta, o dsRNA penetraria diretamente nas células germinativas, explicando assim a transmissão do silenciamento para prole.
Entretanto, os nossos resultados demonstram que o fenótipo de silenciamento da prole também pode ser observado, a partir do terceiro ou quarto ciclo de postura, em insetos injetados com dsRNA na fase de ninfa de quarto estádio. Se consideramos que existe um mecanismo de amplificação do efeito do RNAi em triatomíneos, uma explicação plausível para este resultado seria que pelo fato da gônada na ninfa de quarto estádio ainda não ter iniciado o processo de maturação, a penetração da partícula indutora do RNAi (provavelmente siRNA) nas células germinativas deve ocorrer apenas após a muda para o quinto estádio.
68
Interessante ressaltar que, até o terceiro ciclo de oviposição dos insetos injetados no quarto estádio em jejum, não há diferença significativa entre os valores estimados de concentração de hemeproteínas entre a prole de insetos teste e controle (Fig. 14). Contudo, existe uma elevada dispersão desses valores nos grupos teste, o que sugere que alguns indivíduos da prole possam apresentar redução na expressão de proteínas antes de outros.
69
Acerca do impacto que o uso do RNAi parental poderá trazer nos estudos de genoma funcional dos triatomíneos pode se considerar, sobretudo, dois pontos principais: a) Inalteração do estado fisiológico do insetos, pois atualmente o dsRNA é injetado diretamente na hemolinfa destes, podendo alterar o seu comportamento em decorrência da lesão causada; b) Menor custo do silenciamento, já que uma única dose de dsRNA poderá silenciar, hipoteticamente, não apenas um mas até dezenas de insetos, sem considerar a economia de tempo dedicado à produzir um grupo de insetos com genes silenciados.
70
6.2 – Interação entre NO e proteínas do hospedeiro
Nossa proposta inicial era de que a soro albumina, que circula em abundância no soro de mamíferos, poderia interagir com o NO liberado pelas nitroforinas salivares de no sítio da picada. Tal interação ocorreria entre o grupo tiol do resíduo de cisteína livre da albumina, formando S nitrosoalbumina, que aumentaria o tempo de meia vida do NO através de sua liberação mais lenta e constante nos tecidos e microvasculatura do hospedeiro. Seguindo tal prerrogativa, a formação de S nitrosoalbumina aparenta ser um possível mecanismo responsável pela ocorrência e longa duração da vasodilatação e do extravasamento de plasma no local da picada por