UNIVERSIDADEFEDERALDO RIO GRANDE DO NORTE
UNIVERSIDADEFEDERAL DORIOGRANDE DO NORTE CENTRO DETECNOLOGIA
PROGRAMA DEPÓS-GRADUAÇÃO EMENGENHARIAELÉTRICA
Estudo da topologia de redes de conexão
funcional no córtex sensorial primário e
hipocampo durante o sono de ondas lentas
Edson Anibal de Macedo Reis Batista
Orientador: Prof. Dr. José Alfredo Ferreira Costa
Dissertação de Mestrado apresentada ao
Programa de Pós-Graduação em Engenha-ria Elétrica da UFRN (área de concentração: Engenharia de Computação) como parte dos requisitos para obtenção do título de Mestre em Ciências.
Estudo da topologia de redes de conexão funcional no córtex sensorial primário e hipocampo durante o sono de ondas lentas. / Edson Anibal de Macedo Reis Batista. – Natal, RN, 2013.
92 f.: il.
Orientador: Prof. Dr. José Alfredo Ferreira Costa.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Tecnologia. Programa de Pós-Graduação em Engenharia Elétrica.
1. Redes cerebrais – Engenharia Elétrica - Dissertação. 2. Análise topológica de redes - Dissertação. 3. Mineração de grafos variantes no tempo (TVG) - Dissertação. 4. Sono de ondas lentas (SWS) – Dissertação. 5. Tolerância à falha – Dissertação. I. Costa, José Alfredo Ferreira. II. Universidade Federal do Rio Grande do Norte. III. Título.
Estudo da topologia de r edes de conexão
funcional no cór tex sensor ial pr imár io e
hipocampo dur ante o sono de ondas lentas
Edson Anibal de Macedo Reis Batista
Agradecimentos
Ao meu orientador, professor José Alfredo Ferreira Costa, sou grato pela orientação e entusiasmo pelo qual abraçou este trabalho.
Ao professor Nivaldo Antonio Portela de Vasconcelos sinceros agradecimentos pela opor-tunidade a mim confiada, pela orientação, incentivo e principalmente por sua amizade.
Ao professor Sidarta Ribeiro pela confiança em ceder os dados que analisamos.
Aos colegas docontainerdo Instituto do Cérebro, pelas críticas e sugestões.
Aos demais colegas de pós-graduação, pelo apoio durante as disciplinas.
À minha família pelo apoio durante esta jornada.
A análise da topologia de redes é uma poderosa ferramenta no estudo de sistemas complexos tal como as redes cerebrais. Este trabalho procura descrever as mudanças na topologia de redes de conexão funcional em neurônios do córtex sensorial e do hipocampo durante o sono de ondas lentas (SWS) em animais expostos à novidade. O sono de on-das lentas é um importante estado do sono onde há reverberação de padrões de atividade elétrica ocorridos na vigília, tendo com isso papel fundamental na consolidação de me-mória. Apesar de sua importância ainda não há estudos que caracterizam a dinâmica da topologia de redes de conexão funcional durante este estado. Tampouco há estudos que descrevem as modificações topológicas que a exposição à novidade traz a essas redes. Observamos que várias propriedades topológicas são modificadas após a exposição à no-vidade e que tais modificações se mantêm por um longo período de tempo. A maior parte das propriedades modificadas pela exposição à novidade está relacionada à tolerância à falha.
Abstract
Complex network analysis is a powerful tool into research of complex systems like brain networks. This work aims to describe the topological changes in neural functional connectivity networks of neocortex and hippocampus during slow-wave sleep (SWS) in animals submited to a novel experience exposure. Slow-wave sleep is an important sleep stage where occurs reverberations of electrical activities patterns of wakeness, playing a fundamental role in memory consolidation. Although its importance there’s a lack of studies that characterize the topological dynamical of functional connectivity networks during that sleep stage. There’s no studies that describe the topological modifications that novel exposure leads to this networks. We have observed that several topological properties have been modified after novel exposure and this modification remains for a long time. Major part of this changes in topological properties by novel exposure are related to fault tolerance.
Sumário i
Lista de Figuras iii
Lista de Tabelas ix
1 Introdução 1
1.1 Apresentação do problema . . . 2
1.2 Objetivos . . . 3
Lista de Símbolos e Abreviaturas 1 2 Fundamentação Teórica 5 2.1 Sistema Sensorial . . . 5
2.1.1 Córtex somatossensorial de ratos . . . 6
2.1.2 Córtex visual . . . 6
2.1.3 Hipocampo . . . 7
2.2 Eletrofisiologia . . . 9
2.2.1 Neurônio . . . 9
2.2.2 Potencial de ação . . . 10
2.2.3 Matriz de micro-eletrodos . . . 10
2.3 Conectividade funcional entre neurônios . . . 12
2.4 Redes complexas . . . 14
2.4.1 Redes de Mundo Pequeno . . . 15
2.4.2 Eficiência . . . 18
2.4.3 Centralidade . . . 18
2.4.4 Escores . . . 19
2.4.5 Buracos estruturais . . . 19
3 Revisão de Literatura 21 4 Materiais e Métodos 29 4.1 Origem dos dados . . . 29
4.2 Estimativa da taxa de disparo . . . 31
4.3 Obtenção dos estados comportamentais do ciclo sono-vigília . . . 32
4.4 Construção dos grafos variantes no tempo . . . 32
4.5 Experimento controle . . . 35
5 Resultados e Discussão 39
5.1 Dinâmica da topologia . . . 39
5.2 Modelos Nulos . . . 49
5.3 Bin Shuffle . . . 54
6 Conclusões 63 6.1 Conclusões . . . 63
6.2 Trabalhos Futuros . . . 64
6.2.1 Conectividade funcional . . . 64
6.2.2 Redes de Mundo Pequeno . . . 64
6.2.3 LFP . . . 64
1.1 Grafos representativos da matriz de conectividade efetiva de uma rede neural de 42 células. O primeiro grafo representa o SWS pré-experimento, o grafo central representa o momento do experimento (RUN) durante a vigília, o terceiro grafo representa o SWS pós-experimento. Os pontos pretos no perímetro do círculo (nós) representam os neurônios. As linhas que interligam os pontos (arestas) representam as correlações positivas entre os pares de neurônios, com a cor refletindo a magnitude dessa cor-relação [vermelho, alta corcor-relação (0.2); azul, baixa corcor-relação (0.002)]. As linhas grossas indicam que os pares de neurônios estiveram correlaci-onados tanto na vigília (RUN) quanto no SWS PRÉ ou no SWS PÓS. É possível observar que a maioria dos pares altamente correlacionados que aparecem durante a vigília (RUN), também aparecem no SWS PÓS, mas
não no SWS PRÉ.Adaptado de [Wilson & McNaughton 1994]. . . 2
2.1 Cérebro de rato: áreas corticais e bulbo olfatório. Adaptado de [Puche
2003]. . . 5 2.2 Sistema somatosensorial do rato. (a) Caminhos neurais das vibrissas ao
córtex; (b) Cada vibrissa está associada à uma área específica no córtex. (adaptado de [Petersen 2007]). . . 6 2.3 Subdiviões do córtex de rato. 17, córtex visual primário. 18a e 18b,
res-pectivamente área lateral e medial do córtex extra-estriado. S1, córtex somatossensorial primário. A1, córtex auditivo primário. RS, córtex
re-trosplenial. Adaptado de [Coogan & Burkhalter 1993]. . . 7
2.4 Localização do hipocampo no cérebro de um rato. Adaptado de [Kandel
et al. 2012]. . . 8 2.5 Predição da localização espacial do animal através da taxa de disparo das
células de lugar.Adaptado de [Kandel et al. 2012]. . . 8
2.6 Partes básicas de um neurônio.Adaptado de [Kandel et al. 2012]. . . 9
2.7 Potencial de Ação de um neurônio piramidal do hipocampo em CA1.
Adaptado de [Bean 2007]. . . 10 2.8 Ilustração de um implante de micro-eletrodo intracelular e extracelular.
Adaptado de [Silver 2010] . . . 11 2.9 Matriz de micro-eletrodos. (a) Implante de uma matriz de micro-eletrodos
no cérebro de um rato. (b) Ilustração do implante de duas matrizes de micro-eletrodos em regiões distintas do cérebro. (c) Imagem real de uma matriz de micro-eletrodos. . . 11
2.10 Comparativo entre correlação e métodos baseados em modelos na infe-rência da conexão funcional entre neurônios. (a) e (d) conexão real entre os neurônios. (b) e (e) estimativa das conexões usando correlograma cru-zado. (c) e (f) estimativa das conexões usando método baseado em mo-delo de máxima verossimilhança. Em (b), (e), (c) e (f) fixando-se o trem de disparos do neurônio da coluna, mede-se a influência do neurônio da
linha variando-se o tempo. Adaptado de [Stevenson et al. 2008]. . . 13
2.11 Modelos Lineares Generalizados para estimar os disparos de um determi-nado neurônio. O modelo proposto leva em consideração tanto a atividade recente do neurônio a ser estimado, quanto dos demais neurônios e
tam-bém as variáveis externas (movimento do braço). Adaptado de [Stevenson
et al. 2008]. . . 14
2.12 Redes de mundo pequeno estão entre a ordem e a aleatoriedade. (A) Par-tindo de uma rede perfeitamente regular, onde todos os nós tem o mesmo
número de arestas, e aumentando a probabilidade, p, de uma determinada
aresta ser reconectada a dois nós quaisquer, haverá um intervalo de
valo-res de ponde o fenômeno mundo pequeno acontece. (B) Este fenômeno
é caracterizado por um rápido decréscimo no comprimento do caminho
característicoL(P), por isso a escala logarítmica é usada no eixo y,
cau-sado pela introdução de conexões de longa distância (atalhos), enquanto
o coeficiente de agrupamento C(p) mantém-se elevado para valores de
p<<1. Adaptado de [Watts & Strogatz 1998]. . . 16
2.13 Representação de uma rede complexa usando grafos. Os nós são repre-sentados por padrões circulares e as arestas por linhas que interligam os nós. (A) O comprimento do caminho entre dois nós, em amarelo, é de-finido como o número mínimo de arestas percorridas de um nó ao outro, ou seja, a distância em arestas, entre dois nós. O comprimento do
cami-nho característico L(p) é a média das distâncias entre todos os pares de
nós. (B) O coeficiente de agrupamentoC(p)é a fração dos vizinhos de
um nó que também são vizinhos entre si (em amarelo). (C) O grau kde
um nó é o número de nós conectados a ele. Um hub é um nó cujo grau é muito maior que o grau médio. O grau médio da rede é 3.3, portanto os nós com grau 6 são os nós hub (em amarelo). Adaptado de [Bassett & Bullmore 2006]. . . 17
indivíduos, as conexões que interligam os buracos estruturais garantem eficiência na transmissão de informação ao passo que a não redundância minimiza o custo em manter tais conexões (A). Uma maior densidade de conexão implica numa maior eficiência na troca de informação, ao passo que a redundância nas interligações entre buracos estruturais não aumenta a eficiência do fluxo de informação (B) e (C). As ligações não redundantes
entre buracos estruturais são mais econômicas. Adaptado de [Burt 1995]. 22
3.3 Topologia das redes cerebrais minimiza o custo metabólico de fiação en-quanto maximiza a integração entre áreas. Redes construídas visando apenas a minimização do custo de fiação terão uma topologia em grade pois os vizinhos topológicos também são os vizinhos espaciais (imagem esquerda). Essa topologia não facilita a integração entre diferentes áreas. Uma topologia aleatória irá maximizar a integração entre áreas, porém a um alto custo de fiação (imagem direita). A topologia de mundo pequeno das redes cerebrais (imagem central) facilita a integração entre áreas ao mesmo tempo que minimiza o custo de fiação devido a existência de al-gumas poucas conexões de longo alcançe. Adaptado de [Bullmore & Sporns 2012]. . . 23 3.4 Construção de redes de conectividade cerebral. As redes de conectividade
estrutural, em geral, advém de técnicas de histologia (traçador de vias), ou neuroimagem (MRI de difusão). As redes de conectividade funcional, em geral, advém de técnicas de neuroimagem (fMRI) ou neurofisiologia (EEG, MEG). Essas redes são representadas por matrizes de adjacências. Com o objetivo de simplificar a análise as redes são binarizadas, ou seja, os pesos de cada aresta serão desconsiderados, a rede é então transfor-mada numa rede não ponderada e simétrica. Adaptado de [Rubinov & Sporns 2010]. . . 25 3.5 Dados de fMRI foram particionados em N=112 diferentes estruturas
cor-ticais que constituem os nós da rede. A correlação de pearson entre a atividade média em cada região nos dá as arestas da rede. Adaptado de [Bassett et al. 2011]. . . 26 3.6 Flexibilidade e aprendizagem. O experimento consistiu em três sessões de
uma tarefa de aprendizagem motora. (A) Há um aumento da flexibilidade nas primeiras sessões, seguido por um decréscimo nas sessões finais. (B) Predição da aprendizagem a partir da correlação entre a flexibilidade me-dida na sessão 1 e a aprendizagem da sessão 2, e entre a flexibilidade na sessão 2 e a aprendizagem na sessão 3. (C) Regiões cerebrais onde a fle-xibilidade da sessão 1 predisse a aprendizagem da sessão 2. (D) Regiões cerebrais onde a flexibilidade da sessão 2 predisse a aprendizagem da ses-são 3. Nas figuras (C) e (D) as cores indicam o coeficiente de correlação
de postos deSpearman, r, entre flexibilidade e aprendizagem. Adaptado
4.1 Visão geral do experimento. O animal implantado é colocado em uma caixa inicialmente vazia durante o período de pré-exposição (a). Após 2h inicia-se o período de exposição (b) onde 4 objetos completamente novos para o animal são colocados na caixa por 20min e o animal pode os explorar livremente. Em seguida, no período de pós-exposição (c), os objetos são retirados e o animal continua na caixa por um período de 3h. Adaptado de [Ribeiro et al. 2007]. . . 30
4.2 Sistema de seleção e registro: (a) Animal implantado ligado ao sistema. (b) Visão geral do sistema. (c) Identificação dos disparos de cada neurônio baseada na forma de onda do sinal. . . 31
4.3 Trem de disparos: cada linha horizontal representa um neurônio, cada barra vertical representa um disparo neuronal ao longo do tempo. As taxas de disparo de cada neurônio de (a) e (b) são iguais, porém seu ISI é
diferente. Adaptado de [Dayan & Abbott 2005] . . . 32
4.4 Quantização da força de conexão. (a) Inicialmente a força de conexão é
dada pelo coeficiente de correlação de Pearsonentre cada par de
neurô-nios. (b) A força de conexão é então substituída pelo seu módulo, afim de obter apenas valores positivos. (c) Finalmente é feita uma quantização dos valores de força de conexão, usando o algoritmo k-means, com k=3. O grupo dos menores valores tem suas arestas excluídas do grafo. O grupo dos maiores valores tem a força de conexão de suas arestas substituída por 2, significando forte conexão. Já o grupo dos valores intermediários tem a força de conexão de suas arestas substituída por 1, significando conexão fraca. Dessa forma temos um grafo quantizado no qual os limiares que de-finem as forças de conexão se modificam dinamicamente de acordo com a atividade instantânea da rede. Esta quantização só é considerada válida
caso o teste deMann-Whitneyindique que há diferença significativa entre
os três grupos (α=0.02). . . 34
4.5 Obtenção das propriedades topológicas da rede. (a) A partir do trem de disparo de cada neurônio move-se sem sobreposição uma janela de 1s. (b) A conexão funcional é então estimada como explicado na figura 4.4. (c) A base de dados a ser utilizada neste trabalho é então construída através da extração de várias propriedades da topologia de tais redes. . . 35
4.6 Bin Shuffle: A ordem dos bins, de cada trem de disparo individualmente, é trocada aleatoriamente para cada janela de 1s. Isso destrói a ordem tem-poral da atividade neural, dessincronizando os neurônios, porém
man-tém a taxa de disparo global de cada neurônio. Adaptado de [Louie &
(b). As propriedades da topologia dessas dez redes foram então extraídas e a mediana de cada um dos dez valores foi calculada (c). De forma que a mediana desses dez valores, para cada uma das propriedades topológicas, representa o comportamento daquela propriedade para o grafo aleatório associado ao grafo real. . . 37
5.1 Taxa de disparo por área. Pontos são a mediana da taxa(bins de 20ms)em 10 minutos
de sws. Barras verticais são o desvio padrão. A linha vertical vermelha separa os momentos de pré e pós exposição. Linha horizontal azul evidencia a diferença
(Mann-Whitney,α≤0.05)entre pré e pós. . . 41
5.2 Curvas representativas para a propriedadenúmero de comunidades. Cada
ponto representa a mediana da propriedade em um intervalo de 10 minu-tos durante o sono SWS. A linha vertical vermelha separa os momenminu-tos de PRÉ e PÓS exposição à novidade. Caso exista diferença significativa entre estes momentos uma linha horizontal azul é exibida na mediana dos pontos. . . 42
5.3 Curvas representativas para a propriedadecentralidade de intermediação
de nó. Cada ponto representa a mediana da propriedade em um intervalo de 10 minutos durante o sono SWS. A linha vertical vermelha separa os momentos de PRÉ e PÓS exposição à novidade. Caso exista diferença significativa entre estes momentos uma linha horizontal azul é exibida na mediana dos pontos. . . 43
5.4 Curvas representativas para a propriedade restrição. Cada ponto
repre-senta a mediana da propriedade em um intervalo de 10 minutos durante o sono SWS. A linha vertical vermelha separa os momentos de PRÉ e PÓS exposição à novidade. Caso exista diferença significativa entre estes momentos uma linha horizontal azul é exibida na mediana dos pontos. . . 44
5.5 Curvas representativas para a propriedadenúmero de arestas. Cada ponto
Lista de Tabelas
3.1 Parâmetros topológicos, em redes de conectividade anatômica, que defi-nem as redes de pequeno mundo para dados de conectividade anatômica do córtex visual e do córtex inteiro de macaco, como também do córtex de
gato.L,C,λeγmostram que essas redes são de mundo pequeno [Hilgetag
et al. 2000]. Eglobal e Elocal medidos para os mesmos dados em outro
estudo [Latora & Marchiori 2003], corroborando a característica mundo pequeno de tais redes. Adaptado de [Bullmore & Sporns 2012] . . . 24
5.1 Quantidade de neurônios por área. O número de neurônios não é uniformemente distribuído por área. A maior parte dos animais possui uma quantidade conside-ravelmente maior de neurônios localizados no hipocampo, HP, (GE9-GE17). Já uma pequena parcela possui a maior parte dos neurônios está localizada ou no córtex somatossensorial primário, S1, ou no córtex visual primário, V1, (GE1-GE5). . . 39
5.2 Diferença nas propriedades topológicas entre pré-exposição (PRÉ) e
pós-exposição à novidade (PÓS), nos dados reais. A letra M significa que
houve diferença significa na mediana dos valores (Mann-Whitney, α≤
0.05). A letra D significa que a diferença ocorreu no desvio padrão. A
seta apontando para cima,↑, significa que houve um aumento de PRÉ para
PÓS. A seta apontando para baixo,↓, significa que houve uma diminuição
de PRÉ para PÓS. . . 47
5.3 Continuação da tabela 5.2 para as demais propriedades topológicas. A
letra M significa que houve diferença significa na mediana dos valores
(Mann-Whitney, α≤0.05). A letra Dsignifica que a diferença ocorreu
no desvio padrão. A seta apontando para cima,↑, significa que houve um
aumento de PRÉ para PÓS. A seta apontando para baixo,↓, significa que
houve uma diminuição de PRÉ para PÓS. . . 48
5.4 Diferença nas propriedades topológicas entre pré-exposição (PRÉ) e
pós-exposição à novidade (PÓS), nos modelos nulos. A letraMsignifica que
houve diferença significa na mediana dos valores (Mann-Whitney, α≤
0.05). A letra D significa que a diferença ocorreu no desvio padrão. A
seta apontando para cima,↑, significa que houve um aumento de PRÉ para
PÓS. A seta apontando para baixo,↓, significa que houve uma diminuição
de PRÉ para PÓS. . . 50
nifica que a diferença ocorreu no desvio padrão. A seta apontando para
cima,↑, significa que houve um aumento de PRÉ para PÓS. A seta
apon-tando para baixo, ↓, significa que houve uma diminuição de PRÉ para
PÓS. . . 51
5.6 Dados Reais ∩ Modelos Nulos. O símbolo ✕ marca as propriedades
que apresentaram diferença significa (Mann-Whitney,α≤0.05) tanto nos
dados reais quanto nos modelos nulos. . . 52
5.7 Continuação da tabela 5.6. O símbolo ✕marca as propriedades que
apre-sentaram diferença significa (Mann-Whitney, α≤0.05) tanto nos dados
reais quanto nos modelos nulos. . . 53 5.8 Diferença nas propriedades topológicas entre pré-exposição (PRÉ) e
pós-exposição à novidade (PÓS), nos dados de Bin Shuffle. A letraMsignifica
que houve diferença significa na mediana dos valores (Mann-Whitney,
α≤0.05). A letraDsignifica que a diferença ocorreu no desvio padrão. A
seta apontando para cima,↑, significa que houve um aumento de PRÉ para
PÓS. A seta apontando para baixo,↓, significa que houve uma diminuição
de PRÉ para PÓS. . . 55
5.9 Continuação da tabela 5.8. A letra M significa que houve diferença
sig-nifica na mediana dos valores (Mann-Whitney,α≤0.05). A letraD
sig-nifica que a diferença ocorreu no desvio padrão. A seta apontando para
cima,↑, significa que houve um aumento de PRÉ para PÓS. A seta
apon-tando para baixo, ↓, significa que houve uma diminuição de PRÉ para
PÓS. . . 56
5.10 Dados Reais ∩ Bin Shuffle. O símbolo ✕ marca as propriedades que
apresentaram diferença significa entre os momentos PRÉ e PÓS
(Mann-Whitney,α≤0.05) tanto nos dados reais quanto nos dados de Bin Shuffle. 57
5.11 Continuação da tabela 5.10. O símbolo ✕ marca as propriedades que
apresentaram diferença significa entre os momentos PRÉ e PÓS
(Mann-Whitney,α≤0.05) tanto nos dados reais quanto nos dados de Bin Shuffle. 58
5.12 Dados Reais \ Bin Shuffle. O símbolo ✕ marca as propriedades que
apresentaram diferença significa entre os momentos PRÉ e PÓS
(Mann-Whitney,α≤0.05) nos dados reais mas não nos dados de Bin Shuffle. . . 59
5.13 Continuação da tabela 5.12. O símbolo ✕ marca as propriedades que
apresentaram diferença significa entre os momentos PRÉ e PÓS
Capítulo 1
Introdução
Assim como ocorre em diversas áreas do conhecimento [Foster 2001], os estudos em neurociência envolvem a análise de bases de dados de volume e complexidade cada vez maiores [Stevenson & Körding 2010]. Em neurociência uma importante fonte de grande volume dos dados e da complexidade de questões e análises advém do registro da ativi-dade eletrofisiológica de populações de neurônios [Hirase et al. 2001, Kim et al. 2007, Logothetis 1995, Nicolelis 2008, Nicolelis & Chapin 1994, Nicolelis et al. 1995, Nicole-lis et al. 2003, Wilson & McNaughton 1994]. Esta técnica, de uma maneira geral, utiliza implantes de matrizes de microeletrodos para monitorar a atividade elétrica extracelular de uma população de neurônios, tendo como principal dado resultante uma sequência de instantes de tempo nos quais cada neurônio efetuou um potencial de ação (disparo). Um
trem de disparos de um neurônionié o conjunto de disparos coletados num determinado
intervalo de tempo onde ocorreramkpotenciais de ação,ni= [t1, ...,tk]. Após o registro da atividade elétrica deN neurônios tem-se uma lista de trens de disparos,Pj= [n1, ...,nN]. Uma outra informação importante que pode ser registrada através de matrizes de mi-croeletrodos são os potenciais de campo local (LFP) [Buzsáki & Draguhn 2004], que
correspondem ao sinal do potencial elétricosk(t)captado no eletrodokno tempot.
Uma questão central na neurociência é como o comportamento do indivíduo emerge das interações entre neurônios [Strogatz 2001]. Em todos os vertebrados superiores, in-cluindo os humanos, sucessões espontâneas de estados cerebrais globais alternam-se entre
os estados de sono e vigília, o que define ociclo sono-vigília. Apesar da onipresença deste
fenômeno em todos os vertebrados superiores, os fatores associados às transições entre estes estados cerebrais continuam pouco entendidos [Gervasoni et al. 2004]. Em 2004,
1.1
Apresentação do problema
O sono desempenha papel fundamental na consolidação de novas memórias, que é o processo que transforma memórias ainda instáveis em representações de longa duração mais estáveis [Ribeiro et al. 2007, Diekelmann & Born 2010]. Esse processo envolve a constante reativação (Figura 1.1), no sono de ondas lentas (SWS), dos circuitos neu-rais utilizados durante a aquisição destas memórias [Wilson & McNaughton 1994, Lee & Wilson 2002, Ribeiro et al. 2007, Diekelmann & Born 2010], e posterior migração da in-formação mnemônica do hipocampo para o neocórtex [Pavlides & Winson 1989, Ribeiro et al. 2007].
PRÉ RUN PÓS
Figura 1.1: Grafos representativos da matriz de conectividade efetiva de uma rede neural de 42 células. O primeiro grafo representa o SWS pré-experimento, o grafo central re-presenta o momento do experimento (RUN) durante a vigília, o terceiro grafo rere-presenta o SWS pós-experimento. Os pontos pretos no perímetro do círculo (nós) representam os neurônios. As linhas que interligam os pontos (arestas) representam as correlações positivas entre os pares de neurônios, com a cor refletindo a magnitude dessa correlação [vermelho, alta correlação (0.2); azul, baixa correlação (0.002)]. As linhas grossas indi-cam que os pares de neurônios estiveram correlacionados tanto na vigília (RUN) quanto no SWS PRÉ ou no SWS PÓS. É possível observar que a maioria dos pares altamente correlacionados que aparecem durante a vigília (RUN), também aparecem no SWS PÓS,
mas não no SWS PRÉ.Adaptado de [Wilson & McNaughton 1994].
Devido à natureza do sinal do LFP, que não é formado pela atividade de um único neurônio mas sim pela soma das atividades elétricas de várias unidades neuronais, sua análise isolada não permite entender como as redes complexas de interação neural se rela-cionam com o sono de ondas lentas (SWS). Para tanto é necessária a análise em conjunto do registro das atividades eletrofisiológicas extracelulares de populações de neurônios através do implante de matrizes de microeletrodos.
1.2. OBJETIVOS 3
mineração de grafos ou análise de redes complexas [Washio & Motoda 2003, Chakrabarti & Faloutsos 2006]. O conceito de grafo foi proposto a mais de 300 anos pelo matemático
Leonhard Euler. Porém apenas recentemente foram realizados estudos de grafos onde a dimensão temporal das arestas é explorada [Tang et al. 2010]. A análise de redes comple-xas tem sido utilizada em vários estudos de diferentes áreas, tais como classificação de psicoses [Mota et al. 2012], predição de complexos proteicos [Li et al. 2005], análise de rede de consumidores [Domingos & Richardson 2001], e mineração de fragmentos mo-leculares [Nijssen & Kok 2004], entre outros. Estes estudos mostram que, em suas redes correspondentes, existe informação relevante sobre os respectivos processos subjacentes. Este trabalho descreve as mudanças na topologia de redes de conectividade funcional durante o sono de ondas lentas (SWS) em ratos que foram expostos à novidade. Tais mudanças podem estar relacionadas à consolidação da informação mnemônica referente à novidade a qual os animais foram expostos.
1.2
Objetivos
O objetivo geral deste trabalho é verificar se a topologia das redes de conectividade funcional do neocórtex e hipocampo, durante o sono de ondas lentas (SWS), é alterada devido à exposição à novidade ocorrida na vigília, e se essa alteração mantem-se estável durante sucessivas entradas no SWS pós-exposição.
Capítulo 2
Fundamentação Teórica
2.1
Sistema Sensorial
A adaptação dos animais, de uma forma geral, requer o processamento constante de informações recebidas do meio. A detecção das alterações do meio através do sistema sensorial permite a regulação e o controle das atividades do indivíduo. Dentro da evo-lução geral dos sistemas sensoriais, os mamíferos adquiriram a capacidade de qualificar e quantificar os parâmetros do estímulo em termos de qual é a natureza do estímulo, o quanto ele atua e onde ele está sendo aplicado [Oliveira et al. 2002]. O sistema sensorial codifica um determinado estímulo sob quatro aspectos: modalidade, intensidade, locali-zação e duração. A modalidade sensorial é o tipo do fenômeno físico que pode ser sentido (pressão, temperatura, tato, som, etc). Já a intensidade indica o quanto o estímulo atua. A localização indica onde o estímulo atua, e a duração indica por quanto tempo o estímulo atua [Oliveira et al. 2002].
O córtex cerebral é um elemento anatômico e funcional onde acontece a maior parte da interpretação, organização e armazenamento das informações sensoriais. Para cada modalidade sensorial tem-se uma área do córtex especializada em seu processamento [Kandel et al. 2012].
Figura 2.1: Cérebro de rato: áreas corticais e bulbo olfatório.Adaptado de [Puche 2003].
Na seção 2.1.1 o córtex somatossensorial será apresentado. A seção 2.1.2 apresentará o córtex visual. Já o hipocampo será introduzido na seção 2.1.3.
2.1.1
Córtex somatossensorial de ratos
O rato é um animal cujo tato é bastante desenvolvido. Acostumado a viver em um am-biente de pouca luminosidade e espaço físico limitado, ele conta principalmente com um conjunto de vibrissas, localizado próximo ao focinho (Figuras 2.2.a e 2.2.b), para navegar em busca de comida e proteção. Através das vibrissas o animal recebe a informação táctil de maneira bastante precisa, podendo fazer a discriminação fina de textura e localização de objetos [Petersen 2007].
Figura 2.2: Sistema somatosensorial do rato. (a) Caminhos neurais das vibrissas ao
córtex; (b) Cada vibrissa está associada à uma área específica no córtex. (adaptado de
[Petersen 2007]).
Durante o processo de exploração do ambiente, há um intenso processamento senso-rial táctil, feito pelo córtex somatossensosenso-rial, e o processamento motor, responsável pela movimentação das vibrissas. Na figura 2.2.a pode ser observado o caminho que a infor-mação sensorial percorre das vibrissas até o córtex somatosensorial, bem como o caminho que a informação motora percorre do córtex motor até as vibrissas. A figura 2.2.b mostra que cada vibrissa está associada individualmente à uma região anatômica específica do córtex somatossensorial do rato denominada barril [Woolsey & Van der Loos 1970]. Já a figura 2.2.c é uma fotografia do córtex somatossensorial detalhando os barris.
2.1.2
Córtex visual
O córtex visual é uma área específica do cérebro de vários animais especializada no processamento da visão. A percepção visual emerge da codificação de várias proprie-dades óticas (cor, contraste, orientação, forma, movimento) e posterior integração dessa informação em diferentes áreas do córtex visual hierarquicamente organizadas [Coogan & Burkhalter 1993]. Dentre as diferentes áreas especializadas do córtex visual, encontra-se o córtex visual primário (V1) e as demais áreas V2, V3, V4 e V5. O córtex visual
primário é também chamado de córtex estriado e corresponde à área 17 de Broadmann
2.1. SISTEMA SENSORIAL 7
chega e é então enviada para diferentes áreas do córtex visual [Malach 1989, Coogan & Burkhalter 1993].
Figura 2.3: Subdiviões do córtex de rato. 17, córtex visual primário. 18a e 18b, res-pectivamente área lateral e medial do córtex extra-estriado. S1, córtex somatossensorial
primário. A1, córtex auditivo primário. RS, córtex retrosplenial. Adaptado de [Coogan
& Burkhalter 1993].
2.1.3
Hipocampo
Figura 2.4: Localização do hipocampo no cérebro de um rato. Adaptado de [Kandel et al. 2012].
Nos roedores (Figura 2.4), o hipocampo tem sido extensivamente estudado como parte do sistema cerebral responsável pela memória espacial e navegação. Muitos neurônios do hipocampo de ratos e camundongos respondem de forma característica quando o animal
passa por uma região específica do ambiente. Tais células são chamadas de células de
lugar (place cells). Uma vez que seja encontrada uma célula de lugar, é possível saber qual a região do espaço o animal se encontra observando a taxa de disparo de tal célula (Figura 2.5).
Figura 2.5: Predição da localização espacial do animal através da taxa de disparo das
2.2. ELETROFISIOLOGIA 9
2.2
Eletrofisiologia
No sistema nervoso a informação é transportada no interior e entre os neurônios por meio de sinais elétricos e químicos. Os sinais elétricos são particularmente importantes na transmissão da informação cobrindo rapidamente longas distâncias [Kandel et al. 2012]. Na seção 2.2.1 será descrito o neurônio, a principal célula do cérebro. Já na seção 2.2.2 os sinais elétricos associados à transmissão da informação no cérebro serão melhor descritos. Na seção 2.2.3 será detalhada a abordagem utilizada para efetuar o registro da atividade elétrica extracelular de populações de neurônios.
2.2.1
Neurônio
O neurônio é a célula do sistema nervoso responsável pelo processamento de infor-mação. É considerada a unidade básica da estrutura do cérebro e do sistema nervoso. Em termos gerais, o neurônio é constituído pelas seguintes partes: corpo celular, dendritos e o axônio (Figura 2.6).
Dendritos (corpo celular)Soma
Núcleo
Dendritos
Axônio
Nodos de Ranvier Bainha de Mielina
Terminais Axonais
Figura 2.6: Partes básicas de um neurônio.Adaptado de [Kandel et al. 2012].
A membrana exterior de um neurônio forma vários ramos chamados de dendritos, que são os responsáveis por coletar os sinais elétricos vindos de outras células neuronais, e uma estrutura chamada de axônio que envia os sinais elétricos, resultado do processa-mento feito pelo neurônio, às demais células interligadas a ele. O espaço entre o dendrito de um neurônio e o axônio de outro é chamado de sinapse. O impulso nervoso é trans-portado através das sinapses por uma variedade de substâncias químicas chamadas de neurotransmissores.
2.2.2
Potencial de ação
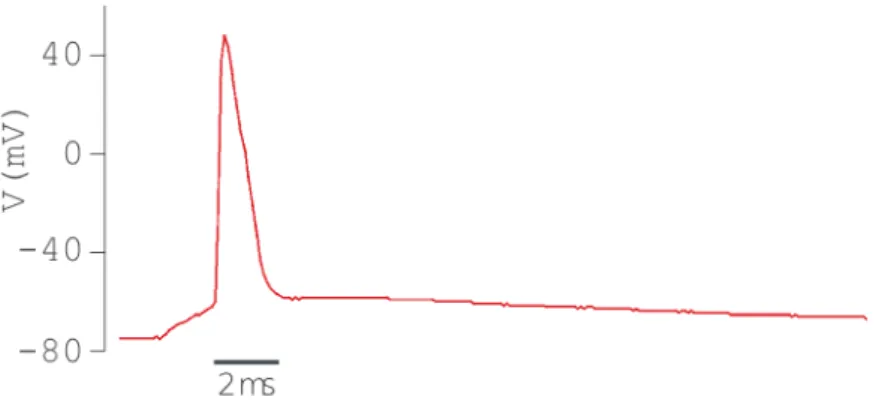
Quando um neurônio dispara, ele gera um pulso elétrico de amplitude aproximada de 100mV e duração em torno de 1 a 2ms. Esses pulsos elétricos são chamados de potencial de ação ou disparos (spikes), e mantém sua forma de onda inalterada enquanto percorre o interior do axônio.
Sempre existe uma diferença de potencial entre o interior e o exterior de uma célula neural, isso é causado pela distribuição desigual dos íons nos dois lados da membrana, e pela permeabilidade da membrana a esses íons. A diferença de potencial de um neurônio inativo é de aproximadamente -70mV sendo o interior mais negativo que o meio externo. Quando a membrana do neurônio é despolarizada além de um limiar de aproximada-mente -55mV, ocorre então o disparo e o pulso elétrico, de aproximadaaproximada-mente 40mV, é transmitido para as demais células ligadas ao seu axônio. Após o disparo a diferença de potencial no interior da membrana cai, durante um período de tempo conhecido como
período refratário, para um valor ainda mais negativo que na situação de equilíbrio, vol-tando novamente ao valor inicial (Figura 2.7).
Figura 2.7: Potencial de Ação de um neurônio piramidal do hipocampo em CA1.
Adap-tado de [Bean 2007].
O período refratário é dividido em período refratário absoluto, no qual é impossível haver um novo potencial de ação já que os canais de sódio ainda estão inativos, e período refratário relativo, no qual somente estímulos mais fortes (supralimiares) conseguem evo-car um novo potencial de ação. O efeito geral do período refratário é o da limitação da frequência dos potenciais de ação, e a unidirecionalidade de sua propagação ao longo do axônio.
2.2.3
Matriz de micro-eletrodos
2.2. ELETROFISIOLOGIA 11
lados da membrana celular. Fios inseridos no final da pipeta eram ligados a um ampli-ficador e posteriormente a um osciloscópio que mostrava a amplitude do potencial da membrana em volts. Uma vez que o diâmetro da ponta de um micro-eletrodo é muito pequeno, ele pode ser inserido no interior da célula com relativamente poucos danos à membrana celular [Kandel et al. 2012].
Figura 2.8: Ilustração de um implante de micro-eletrodo intracelular e extracelular.
Adap-tado de [Silver 2010]
Registros intracelulares são, em geral, feitos in vitro, como em experimentos com fatias de tecido neural. Existem também registros extracelulares, onde eletrodo está pró-ximo da célula mas não penetra sua membrana (figura 2.8). Estes são, em geral, feitos in vivo, especialmente em experimentos que envolvem o comportamendo do animal.
Figura 2.9: Matriz de micro-eletrodos. (a) Implante de uma matriz de micro-eletrodos no cérebro de um rato. (b) Ilustração do implante de duas matrizes de micro-eletrodos em regiões distintas do cérebro. (c) Imagem real de uma matriz de micro-eletrodos.
A abordagem atual para o registro extracelular de grandes populações de neurônios surgiu em meados de 1980 [Nicolelis & Lebedev 2009]. Ela faz uso intensivo de matrizes de micro-eletrodos que são implantadas em regiões específicas do cérebro.
figura 2.9.a e 2.9.b pode-se ver o implante de uma matriz de micro-eletrodos no cérebro de um rato.
2.3
Conectividade funcional entre neurônios
Uma das questões chave na pesquisa em neurociências é o entendimento de como as redes de neurônios interagem para codificar informação. Essas interações ocorrem em diferentes escalas temporais e espaciais [Stevenson & Körding 2010]. De uma maneira geral os neurônios não disparam de forma independente uns dos outros, exibindo padrões de disparos correlacionados [Perkel et al. 1967]. Isto se dá por diferentes fatores [Perkel et al. 1967, Stevenson et al. 2008] (I) Os neurônios podem interagir monossinapticamente ou polissinapticamente. (II) Eles podem codificar propriedades similares do ambiente ou do movimento planejado. (III) Eles podem receber a mesma entrada de outros neurônios não registrados do sistema nervoso.
Em muitos casos, por exemplo em registros eletrofisiológicos usando matrizes de micro-eletrodos, não é possível determinar as conexões anatômicas existentes entre os neurônios. Neste caso podemos apenas inferir as conexões funcionais baseado em medi-das que estimam os disparos neuronais a partir dos disparos dos demais neurônios (Figura 2.10). Se um determinado neurônio influencia o comportamento de outro, seja
aumen-tando ou diminuindo a probabilidade de disparos, diz-se que há umaconexão funcional
entre eles.
Uma das formas mais simples de inferir conexão funcional é através de medidas de correlação para caracterizar as interações entre os neurônios [Brown et al. 2004, Dayan & Abbott 2005, Kass et al. 2005, Bassett et al. 2011]. Estas medidas são usadas em
cor-relogramas cruzados [Perkel et al. 1967] ejoint peri-stimulus time histograms[Gersteins
& Perkel 1962], por exemplo. A principal desvantagem dos métodos baseados em corre-lação é que os mesmos só inferem as relações lineares entre os neurônios. Outras abor-dagens mais modernas fazem uso das componentes principais da matriz de contagem
de disparos neuronais 1 [Nicolelis et al. 1995, Peyrache et al. 2009, Lopes-Dos-Santos
et al. 2011]. Porém tais métodos não permitem detalhar a topologia das redes de conexão funcional, informando apenas a existência de assembléias entre os neurônios envolvidos. Uma alternativa mais recente são os métodos baseados em modelos de máxima verossi-milhança [Paninski et al. 2004, Okatan et al. 2005]. Estes métodos permitem não apenas a identificação das assembléia neuronais, mas também como se dá a interação entre os neurônios dessas assembléias, possibilitando assim a construção dos grafos de conexão funcional (Figura 2.10).
1É um erro comum achar que essas componentes principais são da matriz de correlação. Na verdade
2.3. CONECTIVIDADE FUNCIONAL ENTRE NEURÔNIOS 13
Figura 2.10: Comparativo entre correlação e métodos baseados em modelos na inferência da conexão funcional entre neurônios. (a) e (d) conexão real entre os neurônios. (b) e (e) estimativa das conexões usando correlograma cruzado. (c) e (f) estimativa das conexões usando método baseado em modelo de máxima verossimilhança. Em (b), (e), (c) e (f) fixando-se o trem de disparos do neurônio da coluna, mede-se a influência do neurônio da
linha variando-se o tempo.Adaptado de [Stevenson et al. 2008].
Na figura 2.10 pode-se ver um comparativo entre a estimativa da conexão funcional
entre neurônios utilizando-se de uma medida de correlação linear (correlação de
Pear-son) e um método baseado em modelos de máxima verossimilhança (GLM). Em 2.10.a e
2.10.d temos a conexão real e o trem de disparos associado a cada neurônio. Em 2.10.b percebe-se uma leve correlação entre A e C (painel amarelo) o que poderia levar a uma falsa estimativa de conexão. O mesmo resultado indesejado ocorre em 2.10.e entre C e B (painel amarelo). Já em 2.10.c e 2.10.f fazendo-se uso de métodos baseados em modelos as falsas conexões estimadas são descartadas do processo uma vez que, ao contrário dos métodos baseados em correlação linear que avaliam par-a-par as variáveis envolvidas, nos métodos baseados em modelos de máxima verossimilhança todas as variáveis são levadas em consideração ao mesmo tempo.
Figura 2.11: Modelos Lineares Generalizados para estimar os disparos de um determi-nado neurônio. O modelo proposto leva em consideração tanto a atividade recente do neurônio a ser estimado, quanto dos demais neurônios e também as variáveis externas
(movimento do braço).Adaptado de [Stevenson et al. 2008].
Na figura 2.11 os disparos neurais foram modelados usando modelos lineares genera-lizados (GLM). Neste caso o modelo contém a atividade recente do neurônio, dos demais neurônios e dos estímulos externos (movimento do controle mecânico). Estes parâmetros passam então por uma função não linear (função de ligação) da família das exponenciais, para que finalmente todos os parâmetros possam ser estimados maximizando o logaritmo da função de verossimilhança [Stevenson et al. 2008].
Um pressuposto importante dos métodos baseados em modelos é que as variáveis explicativas devem ser independentes e as observações da variável resposta não devem ser explicadas pelo passado da própria variável. Estes pressupostos não são verdadeiros para os dados que estamos analisando, uma vez que em redes de conexão funcional a atividade elétrica de um neurônio não é totalmente independente da atividade dos demais neurônios nas regiões corticais aqui estudadas. Sendo assim, neste trabalho optamos por
inferir a conectividade funcional usando a correlação dePearson.
2.4
Redes complexas
2.4. REDES COMPLEXAS 15
suíçoLeonhard Paul Eulerao tentar resolver o problema das sete pontes de Königsberg.
Somente dois séculos depois, por volta de 1950, o estudo das redes complexas deu-se início quando sociólogos tentavam caracterizar os fenômenos que ocorrem durante a in-teração de populações humanas [Granovetter 1973].
Na seção 2.4.1 será definida um tipo de rede complexa conhecida como rede de mundo pequeno. As seções 2.4.2, 2.4.3, 2.4.4, e 2.4.5 concentram-se em definir algumas propri-edades importantes da topologia de tais redes.
2.4.1
Redes de Mundo Pequeno
Mundo pequeno é uma topologia de rede caracterizada pela presença de aglomera-dos, densamente conectaaglomera-dos, que são interligados por algumas poucas conexões de longo alcance.
Seja N o conjunto de todos os nós da rede, en a quantidade de nós existentes. L o
conjunto de todas as arestas da rede, el a quantidade de arestas existentes. (i,j)a aresta
que interliga os nósi e j (parai,j∈N). ai j o estado da conexão entrei e j: quando a
conexão(i,j)existe,ai j =1, caso contrárioai j =0. ti o número de arestas que existem
entre os vizinhos de um nó i. Essa topologia é alcançada quando, partindo de um grafo
inteiramente regular, onde todos os nós possuem o mesmo grau k, ou seja, o mesmo
número de arestas:
ki=
∑
j∈Nai j. (2.1)
Aumenta-se gradativamente a probabilidade pde que uma aresta seja reconectada a dois
nós quaisquer do mesmo grafo (Figura 2.12.a). O caminho mínimo entre dois nós é defi-nido comodi j =∑auv∈gi↔jauv, ondegi↔j é a distância geodésica entre ie j. A distância geodésica entre dois nós em um grafo é definida como sendo o número de arestas em um caminho mínimo que os conectam.
Para caracterizar a rede de mundo pequeno duas propriedades de sua topologia são
fundamentais, ocomprimento do caminho característico L(p) (average shortest path
length), que é a média do número de arestas no caminho mínimo entre cada par de nó (Figura 2.13.a):
L= 1
ni
∑
∈NLi=1
ni
∑
∈N∑j∈N,j6=idi j
n−1 . (2.2)
E o coeficiente de agrupamentoC(p) (cluster coefficient), que é a média do número
de vizinhos de um determinado nó que também são vizinhos entre sí, para todos os nós (Figura 2.13.b):
C= 1
ni
∑
∈NCi=1
ni
∑
∈N2ti
ki(ki−1). (2.3)
Na equação 2.2,di j é a distância geodésica entre os nósie j. Já na equação 2.3,ti é
o número de arestas que atualmente existem entre oskivizinhos do nói, eki(ki−1)é o
número total de arestas possíveis entre aqueles nós [Albert & Barabási 2002].
Entre a regularidade,p=0, e a completa aleatoriedade, p=1, existe um intervalo de
causada por algumas poucas conexões que interligam os diferentes aglomerados (Figura 2.12.b). Nestas condições, onde há uma alta densidade de nós interconectados e um baixo comprimento do caminho característico, a rede é chamada de mundo pequeno, em analogia ao fenômeno de mesmo nome apontado por Stanley Milgram [Milgram 1967].
Figura 2.12: Redes de mundo pequeno estão entre a ordem e a aleatoriedade. (A) Par-tindo de uma rede perfeitamente regular, onde todos os nós tem o mesmo número de
arestas, e aumentando a probabilidade, p, de uma determinada aresta ser reconectada a
dois nós quaisquer, haverá um intervalo de valores de ponde o fenômeno mundo pequeno
acontece. (B) Este fenômeno é caracterizado por um rápido decréscimo no comprimento
do caminho característico L(P), por isso a escala logarítmica é usada no eixo y,
cau-sado pela introdução de conexões de longa distância (atalhos), enquanto o coeficiente de
agrupamentoC(p) mantém-se elevado para valores de p<<1. Adaptado de [Watts &
2.4. REDES COMPLEXAS 17
Figura 2.13: Representação de uma rede complexa usando grafos. Os nós são repre-sentados por padrões circulares e as arestas por linhas que interligam os nós. (A) O comprimento do caminho entre dois nós, em amarelo, é definido como o número mínimo de arestas percorridas de um nó ao outro, ou seja, a distância em arestas, entre dois nós.
O comprimento do caminho característicoL(p)é a média das distâncias entre todos os
pares de nós. (B) O coeficiente de agrupamentoC(p)é a fração dos vizinhos de um nó
que também são vizinhos entre si (em amarelo). (C) O grauk de um nó é o número de
nós conectados a ele. Um hub é um nó cujo grau é muito maior que o grau médio. O grau médio da rede é 3.3, portanto os nós com grau 6 são os nós hub (em amarelo). Adaptado de [Bassett & Bullmore 2006].
Afim de avaliar o quão distante da aleatoriedade está a rede estudada é necessário comparar os valores dos atributos dela extraídos com os valores dos atributos extraídos de modelos nulos, que são redes aleatórias contruídas de forma a ter o mesmo número de nós, arestas e distribuição do grau da rede de interesse [Humphries et al. 2006, Bullmore & Bassett 2011]. Em um grafo aleatório o comprimento do caminho característico é em geral curto,
Laleatorio´ ∼ln N/ln([K/N]−1), (2.4)
e o coeficiente de agrupamento assume valores pequenos [Albert & Barabási 2002],
Caleatorio´ ∼(K/N)/N. (2.5)
Para redes de mundo pequeno, por definição [Bassett & Bullmore 2006], temos que
λ=L/Laleatorio´ ≈1 (2.6)
e
A partir disso, uma medida do quão mundo pequeno é a rede pode ser definida como
S= γ
λ. (2.8)
Redes mundo pequeno apresentamS>1 [Humphries et al. 2006].
2.4.2
Eficiência
EficiênciaE é um conjunto de medidas do quão eficiente o fluxo de informação
per-corre a rede [Latora & Marchiori 2001]. Essas medidas são análogas a L e C, com a
vantagem de admitir a existência de nós completamente desconectados na rede. A eficiência global de transmissão de informação é definida como:
Eglobal= 1n
∑
i∈NEi =1
n i
∑
∈N∑j∈N,j6=i di j−1
n−1 (2.9)
OndeN é o conjunto de todos os nós da rede,né a quantidade de nós,Eié a eficiência do
nói. Já a eficiência local é definida como:
Elocal= 1n
∑
i∈NElocal,i=1n
∑
i∈N∑j,h∈N,j6=i ai jaih[djh(Ni)]−1
ki(ki−1) (2.10)
2.4.3
Centralidade
Centralidade é uma classe de medidas que avaliam a importância de um dado nó
ou uma determinada aresta na rede [Freeman 1977]. Podemos destacar a centralidade
de intermediação(betweenness centrality) e acentralidade de proximidade(closeness
centrality) como sendo duas grandes abordagens na determinação do grau de importância de um determinado nó ou uma determinada aresta.
A centralidade de intermediação de nó (node betweenness centrality) é definida
como sendo a quantidade de caminhos mínimos, entre todos os pares de nós, que passam pelo nó em questão [Freeman 1977, Freeman & Smith 1978]. É uma medida da influência de um nó sobre o fluxo de informação entre os demais nós de uma comunidade [Girvan & Newman 2002].
Acentralidade de intermediação de aresta(edge betweenness centrality) é definida
como sendo o número de caminhos mínimos, entre todos os pares de nós, que passam pela aresta em questão [Girvan & Newman 2002]. A centralidade de intermediação de aresta se preocupa com as arestas que são menos centrais com relação a uma comunidade. Ou seja as arestas que são mais ’intermediadoras’ entre duas comunidades [Girvan & Newman 2002].
A partir da determinação das arestas com a maior centralidade de intermediação é
possível encontrar o número de comunidades (number of community) através da
2.4. REDES COMPLEXAS 19
Newman 2002, O’Madadhain et al. 2003]. Uma vez determinadas as comunidades
exis-tentes na rede é possível mensurar o diâmetro do agrupamento(cluster diameter)
cal-culando o diâmetro de cada comunidade. Odiâmetro é definido como sendo a distância
máxima sobre todos os pares de nós no caminho mínimo (shortest path).
A centralidade de proximidade (closeness centrality) é definida como sendo o
in-verso da soma das distâncias geodésicas entre todos os pares de nós [Freeman & Smith 1978]. É uma medida que indica o quão central é um nó para uma determinada comuni-dade em termos de proximicomuni-dade.
Componentes fracas(weak components) formam o maior subgrafo no qual todos os
pares de nós podem ser acessados a partir de qualquer nó do subgrafo [Shiloach 1980].
2.4.4
Escores
O grau de importância de um determinado nó pode ser medido por diferentes
esco-res ou pontuações. A voltagemconsidera o grafo como uma malha elétrica de
resisto-res e pontua um nó de acordo com uma solução aproximada das equações de Kichorff
[Newman & Girvan 2004, Wu & Huberman 2004].
Já obaricentropontua o nó de acordo com a soma das distâncias para todos os outros
nós [O’Madadhain et al. 2003].
2.4.5
Buracos estruturais
A partir da ideia dos buracos estruturais e suas respectivas conexões não redundantes
[Burt 1995] algumas propriedades de redes podem ser extraídas. O tamanho efetivo
(effective size) é definido como sendo a diferença entre o tamanho da rede e a quantidade de conexões redundantes:
ESi=
∑
j[1−
∑
q
piqmjq],q6=i,j (2.11)
Onde piq é proporção da energia de i investida no relacionamento com q. Já mjq é
i-ésima interação com o nóqdividido pelo j-ésimo relacionamento mais forte entre todos
os nós.
Arestrição(constraint) é uma medida que quantifica o grau de importância das
cone-xões entre um indivíduo e seus parceiros dado que os mesmos também estão conectados uns com os outros. Se os parceiros de um indivíduo também são parceiros de todos os parceiros desse indivíduo então o indivíduo em questão tem sua liberdade restrita. Por outro lado, se os parceiros de um determinado indivíduo não possuem outras alternati-vas na vizinhança desse indivíduo então estes não podem restringir o comportamento do indivíduo em questão.
CTi j =pi j−
∑
qpiqmq j,q6=i,j (2.12)
Onde pi j é a proporção da energia deiinvestida no relacionamento com j. Já piq é a
onde N é o tamanho da rede. O nó j restringeina medida quei investiu em j e possui poucos buracos estruturais.
Neste sentido a restrição agregada (aggregated constraint) é o somatório de todas as
restrições da rede.
CTagregada=
∑
N
CTi j (2.13)
Onde N é o número de conexões de um determinado nó.
A eficiência estrutural é definida como sendo a razão entre o tamanho efetivo e o
grau do nó em questão. É uma medida que nos diz o quão eficiênte é a rede em termos de gasto de energia para manter conexões com diferentes agrupamentos. Uma rede onde há muitas conexões para um mesmo agrupamento é menos eficiênte pois há um gasto energético para manter essas conexões redundantes. Essa energia poderia ser aplicada para manter conexões com novos agrupamentos ainda não conectados, o que tornaria a rede mais eficiênte. A eficiência estrutural aumenta quando o número de conexões não redundantes cresce com o número de nós da rede.
A hierarquia é o grau no qual a restrição agregada de um nó está concentrada no
relacionamento de um único contato.
Hierarquia=
∑i(CT CTi j
agregada/N)ln(
CTi j
CTagregada/N)
N ln(N) (2.14)
Capítulo 3
Revisão de Literatura
Neste capítulo será apresentada uma revisão da literatura que trata dos principais con-ceitos utilizados neste trabalho.
Em 1973 Granovetter propos que se um indivíduo A se relaciona com B e com C e este relacionamento é forte, então há uma grande probabilidade de B vir a se relacionar diretamente com C no futuro [Granovetter 1973]. Além disso, o fato de B se relacionar com C torna os relacionamentos entre A e B, e A e C menos independentes (Figura 3.1). Formando uma tríade de conexão social entre indivíduos que seria um bloco básico na formação das redes sociais.
Figura 3.1: Tríade social. (a) Os eventos A está com B, e A está com C são independentes. (b) Se as conexões entre A-B e A-C são fortes então a probabilidade de B-C aumenta e os eventos A-B e A-C tornam-se menos independentes.
Essa premissa básica permite o surgimento de comunidades (aglomerados) onde cada indivíduo está fortemente conectado aos demais membros da sua comunidade. Alguns indivíduos conectam-se a outras comunidades através de conexões fracas. Granovetter
(1973) foca-se “na força das conexões fracas“ mostrando o quão elas são
importan-tes pois permitem o fluxo de informação por entre diferenimportan-tes comunidades de uma rede [Granovetter 1973]. Para Granovetter (1973) as conexões fortes são pouco informativas pois são redundantes e portanto são menos importantes para um indivíduo que procura por recursos ou informação. As conexões fracas, que interligam círculos sociais diferentes, desempenham papel fundamental na manutenção de toda a rede, uma vez que permitem a troca de recursos, informação, ideias, e influências entre diferentes comunidades.
entre estas comunidades de forma que a informação flui com facilidade entre elas, não havendo necessidade de se investir em novas rotas de acesso. Porém existem casos onde não há redundância na conexão entre comunidades. Tais redes tornam-se intimamente dependentes de tais conexões não redundantes que passam a atuar como intermediadoras de recursos e informação, formando o que foi chamado buracos estruturais [Burt 1995]. A partir de tal observação Burt (1992) cria um modelo de competição social onde a im-portância de um indivíduo não está na força de sua conexão (seja ela forte ou fraca) mas sim na quantidade de buracos estruturais que ele interliga (Figura 3.2).
Conexões não redundantes que interligam buracos estruturais são mais econômicas uma vez que a informação já flui com facilidade dentro de uma mesma comunidade. Apenas uma única conexão entre duas comunidades distintas garante a mesma eficiência que várias conexões redundantes entre tais comunidades. Sendo assim, em termos de custo de manutenção de tais conexões, as conexões não redundantes garantem uma boa solução de compromisso entre fluxo de informação e custo de conexão (Figura 3.2).
Figura 3.2: Buracos estruturais podem ocorrer entre indivíduos e/ou entre comunidades de uma rede. Em uma rede esparsa, onde há pouca conexão entre indivíduos, as conexões que interligam os buracos estruturais garantem eficiência na transmissão de informação ao passo que a não redundância minimiza o custo em manter tais conexões (A). Uma maior densidade de conexão implica numa maior eficiência na troca de informação, ao passo que a redundância nas interligações entre buracos estruturais não aumenta a eficiência do fluxo de informação (B) e (C). As ligações não redundantes entre buracos estruturais são
mais econômicas.Adaptado de [Burt 1995]
Em 1998 Watts e Strogatz, em um dos artigos mais citados da história de uma das
revistas mais importantes da atualidade: Nature, propuseram a existência de uma
topo-logia de redes denominada redes de mundo pequeno (small-worlds networks) [Watts &
Strogatz 1998]. Duas medidas da topologia da rede foram propostas com o intuito de
ca-racterizar uma uma rede mundo pequeno, o comprimento do caminho característicoL(p)
(Equação 2.2), e o coeficiente de agrupamento C(p) (Equação 2.3). Redes de mundo
pequeno são caracterizadas por um alto coeficiente de agrupamento e um baixo compri-mento do caminho característico (Figura 2.12.a). Uma das vantagens desse tipo de topo-logia é manutenção da relação de compromisso entre a facilidade do fluxo de informação
pela rede [Watts & Strogatz 1998], já queL(p)é reduzido, e uma diminuição dos custos
metabólicos de fiação [Bullmore & Sporns 2012], já queC(p)é alto (Figura 3.3). Essas
in-23
ternet e naWorld Wide Web[Albert & Barabási 2002]. São portanto um fenômeno geral
de redes complexas [Watts & Strogatz 1998, Albert & Barabási 2002].
Figura 3.3: Topologia das redes cerebrais minimiza o custo metabólico de fiação enquanto maximiza a integração entre áreas. Redes construídas visando apenas a minimização do custo de fiação terão uma topologia em grade pois os vizinhos topológicos também são os vizinhos espaciais (imagem esquerda). Essa topologia não facilita a integração entre diferentes áreas. Uma topologia aleatória irá maximizar a integração entre áreas, porém a um alto custo de fiação (imagem direita). A topologia de mundo pequeno das redes cerebrais (imagem central) facilita a integração entre áreas ao mesmo tempo que minimiza o custo de fiação devido a existência de algumas poucas conexões de longo alcançe. Adaptado de [Bullmore & Sporns 2012].
Em 2001LatoraeMarchioriredefiniram o conceito de redes de mundo pequeno em
termos da eficiênciaE (Equação 2.9 e 2.10) com a quão o fluxo de informação percorre
a rede [Latora & Marchiori 2001]. Uma primeira aproximação da eficiência global pode
ser vista comoEglobal∼1/L se não houver grande diferença entre as distâncias na rede.
A fundamental diferença entreEglobal eL, é queEglobal é a eficiência de um sistema em
paralelo, onde os nós da rede trocam informação concorrentemente, enquanto 1/L é a
eficiência de um sistema em série, onde apenas um único pacote de informação percorre
a rede. Já a eficiência local pode ser vista comoElocal ∼C se a maioria dos subgrafos
Dados L C λ γ Eglobal Elocal
Córtex visual de Macaco 1.73 0.53 1.04 1.47 – –
Córtex inteiro de Macaco 2.38 0.46 1.17 3.06 0.52 0.70
Córtex de Gato 1.81 0.55 1.06 1.77 0.69 0.83
Tabela 3.1: Parâmetros topológicos, em redes de conectividade anatômica, que definem as redes de pequeno mundo para dados de conectividade anatômica do córtex visual e
do córtex inteiro de macaco, como também do córtex de gato. L,C, λeγ mostram que
essas redes são de mundo pequeno [Hilgetag et al. 2000]. Eglobal eElocalmedidos para os
mesmos dados em outro estudo [Latora & Marchiori 2003], corroborando a característica mundo pequeno de tais redes. Adaptado de [Bullmore & Sporns 2012]
O estudo das propriedades que descrevem a topologia de redes provenientes de dados reais é conhecido como análise de redes complexas [Strogatz 2001, Newman 2003, Ru-binov & Sporns 2010]. Ao contrário da teoria dos grafos tradicional, que estuda redes em sua maioria uniformes, aleatórias e simples, a análise de redes complexas estuda re-des da vida real de grande volume, cuja topologia é complexa [Rubinov & Sporns 2010]. As redes cerebrais são um exemplo de redes complexas que compartilham uma série de propriedades, tais como mundo pequeno (Tabela 3), com outras redes biológicas e físicas [Watts & Strogatz 1998, Strogatz 2001, Rubinov & Sporns 2010].
De acordo com a técnica utilizada para fazer o mapeamento cerebral a rede represen-tará um tipo de conectividade específico. Redes de conectividade estrutural são obtidas a partir de técnicas de histologia, tal como traçador de vias, ou de neuroimagem, tal como imagem de difusão por ressonância magnética (MRI de difusão). Já as redes de conecti-vidade funcional são obtidas a partir de técnicas de neuroimagem, tal como imageamento funcional por ressonância magnética (fMRI), ou neurofisiologia, tais como Eletroence-falografia (EEG), MagnetoeletroenceEletroence-falografia (MEG). Tais redes são representadas por matrizes de adjacências onde cada entrada da matriz guarda a força da conexão estimada de acordo com a técnica utilizada, podendo representar o tamanho, densidade, ou coerên-cia se a conexão for estrutural, ou a magnitude das correlações ou interações causais, se a conexão for funcional (Figura 3.4).
25
Figura 3.4: Construção de redes de conectividade cerebral. As redes de conectividade estrutural, em geral, advém de técnicas de histologia (traçador de vias), ou neuroimagem (MRI de difusão). As redes de conectividade funcional, em geral, advém de técnicas de neuroimagem (fMRI) ou neurofisiologia (EEG, MEG). Essas redes são representadas por matrizes de adjacências. Com o objetivo de simplificar a análise as redes são binarizadas, ou seja, os pesos de cada aresta serão desconsiderados, a rede é então transformada numa rede não ponderada e simétrica. Adaptado de [Rubinov & Sporns 2010].
Tais simplificações não são regra geral na análise de redes complexas do cérebro. Em 2011 Danielle Basset [Bassett et al. 2011] em um estudo da dinâmica de reconfiguração das redes cerebrais em humanos durante a aprendizagem de uma tarefa motora, fez uso de redes de conexão funcional, a partir do cálculo da correlação de pearson em dados de fMRI, onde os pesos das conexões foram mantidos (Figura 3.5). Neste estudo mostrou-se que é possível predizer o aprendizado dos indivíduos a partir da propriedade flexibilidade
27
Figura 3.6: Flexibilidade e aprendizagem. O experimento consistiu em três sessões de uma tarefa de aprendizagem motora. (A) Há um aumento da flexibilidade nas primeiras sessões, seguido por um decréscimo nas sessões finais. (B) Predição da aprendizagem a partir da correlação entre a flexibilidade medida na sessão 1 e a aprendizagem da sessão 2, e entre a flexibilidade na sessão 2 e a aprendizagem na sessão 3. (C) Regiões cerebrais onde a flexibilidade da sessão 1 predisse a aprendizagem da sessão 2. (D) Regiões cere-brais onde a flexibilidade da sessão 2 predisse a aprendizagem da sessão 3. Nas figuras
(C) e (D) as cores indicam o coeficiente de correlação de postos deSpearman, r, entre
Capítulo 4
Materiais e Métodos
Neste capítulo apresentaremos a metodologia adotada na solução do problema. Na seção 4.1 é detalhada a origem dos dados analisados neste trabalho. Na seção 4.2 é ex-plicada como a taxa dos disparos neurais é estimada. A seção 4.3 trata sobre como foram identificados os estados comportamentais do ciclo sono-vigília. Na seção 4.4 é descrito como são construídos os grafos variantes no tempo e extraídas suas propriedades topoló-gicas. Já na seção 4.5 os experimentos controle realizados neste trabalho são explicados.
4.1
Origem dos dados
O experimento que deu origem aos dados [Ribeiro et al. 2007] foi realizado no
labo-ratório do Professor Miguel Nicolelis, no Departamento de Neurobiologia daDuke
Uni-versity. Ratos da linhagemLong Evanscriados em um ambiente controlado, sem contato com objetos geometricamente complexos, foram submetidos a uma cirurgia de implante
de três matrizes de micro-eletrodos (35µm) no hipocampo (HP), córtex somestésico
pri-mário (S1) e córtex visual pripri-mário (V1).
Pré-EXP (2 h) Exposição (20 min) Pós-EXP (3 h)
(a) (b) (c)
SWS REM
Vigília
SWS REM
Vigília
Figura 4.1: Visão geral do experimento. O animal implantado é colocado em uma caixa inicialmente vazia durante o período de pré-exposição (a). Após 2h inicia-se o período de exposição (b) onde 4 objetos completamente novos para o animal são colocados na caixa por 20min e o animal pode os explorar livremente. Em seguida, no período de pós-exposição (c), os objetos são retirados e o animal continua na caixa por um período de 3h. Adaptado de [Ribeiro et al. 2007].
Durante o experimento tem-se o registro da atividade elétrica neuronal, feito através de uma matriz de micro-eletrodos, e processado pelo mecanismo de seleção e registro. Cada eletrodo pode registrar a atividade elétrica de até 4 neurônios ao mesmo tempo, sendo papel do mecanismo de seleção e registro identificar e separar cada disparo neuronal (Figura 4.2). Como resultado final do processamento feito pelo mecanismo de seleção e
4.2. ESTIMATIVA DA TAXA DE DISPARO 31
Figura 4.2: Sistema de seleção e registro: (a) Animal implantado ligado ao sistema. (b) Visão geral do sistema. (c) Identificação dos disparos de cada neurônio baseada na forma de onda do sinal.
4.2
Estimativa da taxa de disparo
A maneira como a informação sensorial é codificada entre os neurônios ainda é objeto de estudo da neurociência [Dayan & Abbott 2005, Hung et al. 2005, Vasconcelos et al. 2009]. Uma das possibilidades é que a informação seja codificada através da taxa dos disparos neuronais. De fato, como mostrado por [Nicolelis & Lebedev 2009], é possível encontrar informação suficiente para predizer o movimento dos membros superiores de macacos utilizando a taxa de disparo de um conjunto de neurônios.
Existem pelo menos duas formas de se avaliar a taxa dos disparos neuronais [Dayan & Abbott 2005] (Figura 4.3): Uma é a simples contagem do número de disparos, dentro
de uma janela (bin) de tamanho específico, ocorridos ao longo do tempo (spike counting),
Figura 4.3: Trem de disparos: cada linha horizontal representa um neurônio, cada barra vertical representa um disparo neuronal ao longo do tempo. As taxas de disparo de cada
neurônio de (a) e (b) são iguais, porém seu ISI é diferente. Adaptado de [Dayan &
Abbott 2005]
A contagem do número de disparos neuronais é feita dividindo-se o espaço amostral em janelas (bins) de tamanho específico. Ou seja, os intervalos de contato são divididos em bins e posteriormente o número de disparos que ocorrem dentro de cada um dos bins é contado. Como pode ser observado na Figura 4.3, a contagem do número de disparos desconsidera a estrutura temporal presente no trem de spikes e, portanto, é menos acurada. Por outro lado, a contagem de disparos garante que o vetor de contagem terá o mesmo tamanho para cada neurônio, pois nos bins onde o neurônio não disparou seu valor será zero. O mesmo já não ocorre com o ISI. Os vetores de ISI podem ter tamanhos diferentes para cada neurônio.
Neste trabalho, a contagem de disparos, em bins de 20ms, foi a abordagem utilizada para estimar a taxa de disparos.
4.3
Obtenção dos estados comportamentais do ciclo
sono-vigília
Os estados do ciclo de sono-vigília são detectados automaticamente de formaoff-line,
no trabalho que deu origem aos dados [Ribeiro et al. 2007], por um algoritmo espec-tral quantitativo [Gervasoni et al. 2004] para classificação dos estados comportamentais baseado nos potenciais de campo local (LFP) do córtex e hipocampo. Afim de acessar
o estado comportamental do animal uma janela de 1s move-se sem sobreposição sobre
todo o sinal do LFP, e em cada janela o algoritmo espectral quantitativo é utilizado para estimar tais estados. Isso significa que existe uma marcação do estado comportamental do animal para cada janela de 1 segundo durante todo o experimento. Tal marcação é
denotadahipnograma.
4.4
Construção dos grafos variantes no tempo
Uma janela de 1s é movida sem sobreposição por todo o trem de disparos (Figura
4.5.a) e em cada uma dessas janelas o coeficiente de correlação dePearson, entre a taxa


![Figura 2.6: Partes básicas de um neurônio. Adaptado de [Kandel et al. 2012]. A membrana exterior de um neurônio forma vários ramos chamados de dendritos, que são os responsáveis por coletar os sinais elétricos vindos de outras células neuronais, e uma estr](https://thumb-eu.123doks.com/thumbv2/123dok_br/15614124.106769/25.892.184.755.564.806/neurônio-adaptado-neurônio-chamados-dendritos-responsáveis-elétricos-neuronais.webp)
![Figura 2.8: Ilustração de um implante de micro-eletrodo intracelular e extracelular. Adap- Adap-tado de [Silver 2010]](https://thumb-eu.123doks.com/thumbv2/123dok_br/15614124.106769/27.892.344.584.311.603/figura-ilustração-implante-micro-eletrodo-intracelular-extracelular-silver.webp)





