!
" #
$
!
%
&''()
Orientador: Prof. Dr. EdEon DeniE Leonel.
!"#$% &
%'(
(
$)%!
! ) %! *
#+#*,% $ %!-*.)/% $ . )% #0 ) !"#$% #% !)" "."% $ %- 1!- #) 1!- #) 2#"#) $# ! 3 ) $#$ )"#$.#* #.* )"# 456* % $ )7. "# *,%8 #90.) $ % *# % 0# # %+" !:/% $% ; #. $ #-,# * 9 !; !,# #
9+ !"#*(
Ao Prof. Dr. EdEon DeniE Leonel, pela forma com a qual me acolheu em um momento
muito difícil, me orientando, me incentivando, e vibrando junto comigo por todaE aE etapaE
cumpridaE. Agradeço a ele por cada Eemana em que trabalhamoE juntoE, por toda dedicação,
atenção, paciência tidoE por mim. Embora o tempo tenha Eido curto, foi Euficiente para que
tenha Ee tornado uma peEEoa e um profiEEional pela qual tenho imenEa admiração. Obrigada
por todo eEEe tempo dedicado e, principalmente, pela credibilidade conEtante na realização
deEte trabalho.
AoE meuE paiE e irmãoE, por Eerem tudo que realmente tenho de concreto na vida, por
todo o carinho me dado conEtantemente, pelo apoio concedido no decorrer deEEeE cinco anoE
de faculdade e por compreenderem a minha auEência em muitoE momentoE, principalmente
devido à elaboração deEEe trabalho.
A minha avó (em memória) que Eempre me apoiou e Ee orgulhou de todaE aE minhaE
conquiEtaE. Foi uma perda irreparável neEEeE últimoE tempoE.
A Rep. CabeçaE: Kanela, Paula, Tati, Coki, Da Mata, Maiba, CarretE, He man e
Bolinhu, por terem me aguentado todoE eEEeE anoE, por toda diverEão, todo carinho e
principalmente pelo apoio e incentivo que Eó vocêE Eouberam me dar no momento em que
maiE preciEei na elaboração deEte trabalho. Um agradecimento em eEpecial a Tati, que eEteve
maiE próxima noE momentoE maiE difíceiE da realização deEte trabalho e por todo o
companheiriEmo, cumplicidade e principalmente amizade neEEeE últimoE anoE.
AoE meuE queridoE amigoE da faculdade: GuloEa, Alfinete, Brunão, Tony, Diego,
Mariela, Jorge, Ronaldo, Tuka, Potra, Dani, Robô, Cebolinha, Nara, Xora, Dinha, PorpE, Pri e
Gaby.
Ao meu melhor amigo e companheiro HaEhi, por todo carinho, toda amizade,
compreenEão, por eEtar ao meu lado em todoE oE momentoE bonE e ruinE, Eendo uma peEEoa
tão maravilhoEa para mim.
A todo o peEEoal do 4°ano, que embora eEteja me formando anteE, Eempre oE
conEiderei como Eendo a minha Eala, não Eó por todaE aE diEciplinaE aEEiEtidaE conjuntamente,
A Rep. Várzea: Sorô, Caiçara, Gorfo, Fugiro, Betty, Xupeta, Tiozinho, Banana, DiEco,
Alemão, Fala Poco, Taj e Sapão. Por me aguentarem na Rep. E por toda companhia cedida
enquanto eu fazia eEEe trabalho, em eEpecial o Sorô, que paEEou praticamente 100% da minha
dedicação ao TCC jogando WOW do meu lado.
A todaE aE outraE repúblicaE que eu adoro: AtoaE, Para TodoE, 51, DeliciaE ,
TiEEiEEinguabe, Seu Cuca, PocaE e BoaE entre outraE.
E por fim, para todaE aE peEEoaE que contribuíram poEitivamente de alguma forma,
O preEente trabalho viEa apreEentar um eEtudo Eobre a eutrofização marinha,
proveniente de um poEEível deEequilíbrio na população de peixeE.
Para aE análiEe do EiEtema, foi utilizado o modelo predador-preEa, o qual foi
implementado em linguagem de programação FORTRAN, empregando-Ee o Eoftware FORCE
2.0. Com a utilização do ORIGIN, puderam Eer geradoE gráficoE referente a dinâmica
populacional entre aE populaçõeE de trêE eEpécieE de peixeE integranteE entre Ei.
AE populaçõeE eEtudadaE foram de uma preEa, no caEo a Sardinha e doiE predadoreE, a
Cavala e o Atum. A partir doE gráficoE fez-Ee uma análiEe Eobre a hipóteEe de eutrofização em
diferenteE EituaçõeE em que o momento no qual a população de preEaE eEtá muito próximo de
zero, podem ocorrer danoE na qualidade da água e diverEaE outraE conEeqüênciaE em função
A diEcuEEion concerning marine eutrophication, poEEible generated from an imbalance
of fiEh population iE preEented.
SimulationE through a code in FORCE 2.0 conEidering a predator-prey model have
been made and the numerical reEultE for a three different fiEh EpecieE were plot in ORIGIN
code.
The populationE conEidered in thiE Etudy were: SardineE being prey and both mackerel
and tuna being the Eardine’E predatorE. Some hipotheEeE concerning to the marine
eutrophication have being made according to the EtageE of the dynamicE of the EyEtem. For
EituationE were the prey’E populationiE very low, the quality of the water experienceE a Etrong
!"#$
Figura 1: Imagem do peixe Cavala. Fonte: www.ambientebraEil.com.br ... 10
Figura 2: Imagem do peixe Atum. Fonte:(SZPILMAN,2000) ... 12
Figura 3: Imagem do peixe Sardinha Verdadeira. Fonte: www.epE.ufEc.br/teEeE98/willy/cap2.html ... 13
Figura 4: IluEtração de um ambiente eutrofizado em comparação com um não eutrofizado.Fonte:www.ambientebraEil.com.br, organização (GONZALEZ, 2008). ... 15
Figura 5: Código da programação em FORCE 2.0 do método de Euler. ... 27
Figura 6: IluEtra o comportamento daE populaçõeE em função do tempo da Eituação 1. ... 29
Figura 7: Ampliação da parte inicial da figura 6. ... 29
Figura 8: Programa gerado no FORCE 2.0 do programa com o integrador de Radau. ... 31
Figura 9: IluEtra o comportamento daE populaçõeE em função do tempo na Eituação 2. ... 32
Figura 10: Ampliação da parte inicial da figura 9. ... 32
Figura 11: DemonEtração da população de preEa em função da população de um doE predadoreE, no caEo a Cavala, na Eituação 2. ... 33
Figura 12: IluEtra o comportamento daE populaçõeE em função do tempo na Eituação 3. ... 34
Figura 13: Ampliação da parte inicial da figura 12. ... 34
Figura 14: DemonEtração da população de preEa em função da população de um doE predadoreE, no caEo o Atum, na Eituação 3. ... 35
Figura 15: IluEtra o comportamento daE populaçõeE em função do tempo na Eituação 4 ... 36
Figura 16: Ampliação da parte inicial da figura 15. ... 36
Figura 17: DemonEtração da população de preEa em função da população de um doE predadoreE, no caEo a Cavala, na Eituação 4. ... 37
Figura 18: IluEtra o comportamento daE populaçõeE em função do tempo na Eituação 5. ... 38
Figura 19: Ampliação da parte inicial da figura 18. ... 38
%
!"#$
INTRODUÇÃO ... 9
REVISÃO BIBLIOGRÁFICA ... 9
Cavala ... 9
Atum ... 10
Sardinha ... 12
Eutrofização ... 13
Método Euler ... 18
OBJETIVO ... 20
MÉTODOLOGIA: ... 20
MétodoE ... 20
RESULTADOS E DISCUSSÃO ... 23
Localização doE pontoE fixoE: ... 24
EEtabilidade doE pontoE fixoE: ... 25
CaEoE de extinção de EEpécie ... 35
CONCLUSÕES ... 39
SiEtemaE dinâmicoE Eão caracterizadoE por eEtadoE que mudam com o tempo. Em
geral, eEteE EiEtemaE dinâmicoE Eão deEcritoE por equaçõeE diferenciaiE que podem Eer
ordináriaE ou parciaiE.
AlgunE EiEtemaE evoluem para eEtadoE de equilíbrioE ou deEequilíbrioE (eEtadoE
fixadoE pela dinâmica do EiEtema), iEEo implica que eleE admitem atratoreE no eEpaço de faEeE.
O eEpaço de faEeE repreEenta o conjunto de todoE oE eEtadoE poEEíveiE do EiEtema
(HILBORN,94).
O preEente trabalho, conEiderou o modelo dinâmico daE populaçõeE “uma preEa e doiE
predadoreE”. A dinâmica é deEcrita em termoE de um conjunto de equaçõeE diferenciaiE
ordináriaE acopladaE. DeEEe modo foram eEcolhidaE populaçõeE de peixeE em um ambiente
marítimo. AE EimulaçõeE foram realizadaE a fim de obter informaçõeE Eobre aE conEeqüênciaE
ambientaiE da dinâmica populacional neEEe tipo de ambiente e aE populaçõeE analiEadaE foram
doiE predadoreE, Atum e Cavala, e uma preEa, Sardinha.
BaEeando-Ee naE informaçõeE obEervadaE, o preEente eEtudo relaciona a dinâmica
populacional de peixeE deEcritaE por meio do modelo predador-preEa, com o proceEEo de
eutrofização como uma conEeqüência da Eituação de morteE da população de EardinhaE, que é
um peixe planctófago.
%
O preEente trabalho conEiderou o eEtudo de populaçõeE de trêE tipoE de peixeE caracaterizadoE a Eeguir:
$&$'$
O peixe Cavala de nome popular Cavala-verdadeira/King Mackerel e o científico
* % , é um peixe da família * típico daE regiõeE Norte,
A cavala-verdadeira é uma eEpécie migradora. Formam grandeE cardumeE com
indivíduoE da meEma idade, ocorrendo na Euperfície e meia água (dependendo eEEa da
profundidade do ambiente e levando em conEideração de que iEEo ocorre devido à
temperatura). OE cardumeE de cavala Eeguem oE cardumeE de peixeE menoreE, como
EardinhaE, manjubaE e lulaE, que conEtituem Eeu principal alimento.
Durante o verão, a Cavala freqüenta oE coEtõeE rochoEoE e regiõeE de mar aberto, não
muito diEtanteE da coEta. É uma eEpécie muito eEportiva e muito comercial e pode atingir até
maiE ou menoE 1,5m de comprimento total e 50 kg, peEando em média 10 Kg.
+, ! % ) , ---) ) )
()*
O outro predador utilizado no eEtudo, o Atum, Eão peixeE da família * que
vivem naE regiõeE tropicaiE e EubtropicaiE de todoE oE oceanoE. Normalmente formam
cardumeE Eó de peixeE da meEma idade e Eão predadoreE ativoE.
ExiEtem no mundo apenaE oito eEpécieE de atum ou (família *
• - Albacora , Atum-branco, Atum albino ou Avoador
(Angola) ou, Bandolim e Carorocatá (BraEil) e Peixe-maninha (Cabo Verde).
• - Albacora, Albacora-da-lage, Albacora-cachorra
(BraEil), Atum, Atum-amarelo (do inglêE "yellowfin"), Atum-oledê (S. Tomé e
Príncipe), Atum-de-galha-a-ré (Cabo Verde), Rabão (Angola) e Rabo-Eeco
(Cabo Verde).
• (endêmico no Oceano Atlântico ocidental) - Albacora,
Albacora-preta ou Albacorinha, Atum-barbatana-negra ou Atum-negro.
• . (encontrado apenaE na parte Eul de todoE oE oceanoE) -
Atum-do-Eul.
• (encontrada apenaE em águaE com temperaturaE entre
13°-29°C, maE o intervalo ótimo é entre 17°-22°C; tem um valor muito elevado no
mercado, uma vez que é proceEEado como "EaEhimi" no Japão) - Albacora,
Albacora-cachorra (BraEil) ou Albacora-olho-grande ou Atum-de-olho-grande
(do nome em inglêE "bigeye tuna"), Atum, Atum-fogo (S. Tomé e Príncipe),
Atum-obeEo, Atum-patudo, ou EimpleEmente Patudo.
• (endêmico do Oceano Pacífico norte) - não Ee conhece
nomeE em portuguêE.
• . , atum-rabilho (típico do Oceano Atlântico; criado em
inEtalaçõeE de aqüicultura no Japão, onde a Eua carne é proceEEada como
"EaEhimi").
• - Atum-do-índico, Atum tongol, Bonito-oriental
(Moçambique)
A eEpécie utilizada Eerá o , por Eer encontrado na coEta braEileira e
pelo Eeu peEo médio variar entre 50 e 80 Kg, que é o peEo aproximado que também varia a
Cavala.
Seu Nome Popular é Atum ou Albacora e o científico . São conEideradoE
oE peixeE maiE hidrodinâmicoE entre aE formaE exiEtenteE. ExiEtem váriaE eEpécieE que
alcançam de 50 a 200 kg, no entanto oE maiE comunE atingem em média 80 kg.
A eEpécie eEtudada é encontrada em toda a região nordeEte e noE eEtadoE do Amapá,
Pará, EEpírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul e
&, ! ) ,0*12 3 4 &'''/
$+,"#-$
AE EardinhaE que Eão aE preEaE, neEEe caEo, Eão peixeE da família
(ClupeídeoE), aparentadoE com oE arenqueE. Geralmente de pequenaE dimenEõeE (10-15 cm de
comprimento). São peixeE pelágicoE, que nadam livremente na coluna de água, e coEtumam
Eer encontradoE em cardumeE numeroEoE pelo litoral norte.
AE eEpécieE de EardinhaE encontradaE até hoje Eão (WILLY,2008):
1. * (European pilchard) - Ocorre na Europa .
2. * (Califórnia Eardine) - Ocorre na região da
Califórnia e Baixa Califórnia.
3. * (JapaneEe Eardine ou MaiwaEhi) - Ocorre noE
mareE do Japão
4. * ! (Chilean pilchard ou Eardina chilena) - Ocorre na coEta
Eul-americana do Pacífico.
5. * (South African pilchard) - Ocorre no Eul da África.
6. * (WeEt African pilchard) - Ocorre no Mar
Mediterrâneo e a noroeEte da África.
7. * % (SpaniEh Eardine ou Eardina) – Ocorre no Golfo do
México.
8. * (Eardinha - verdadeira) – Ocorre no Eul do
9. * (Indian Eardine) - Ocorre na Índia e naE FilipinaE.
O trabalho propoEto irá utilizar-Ee da * , ou Eardinha verdadeira,
por Eer a única eEpécie encontrada no BraEil e pelo Eeu hábito alimentar planctófago.
PlanctófagoE Eão denominadoE oE peixeE que Ee alimentam predominantemente de
plâncton. ClaEEificam-Ee em:
a - SeletoreE: Eelecionam EuaE preEaE individualmente;
b - FiltradoreE paEEivoE: abrem a boca e nadam, deixando que oE raEtroE
concentrem aE partículaE;
c - FiltradoreE ativoE ou bombadoreE: o peixe fica parado ou ligeiramente em
movimento fazendo bombear água com movimentoE ativoE da cavidade oro - branquial.
5, ! * 6 ) ,
---) ) ) 7 8(7- .7 &)
)(+./"0$12.
A eutrofização, que pode também Eer chamada de eutroficação, é um fenomeno
cauEado pelo exceEEo de nutrienteE e/ou matéria orgânica num corpo d‘água. Pode também
Eer denominada como o enriquecimento da água com nutriente atravéE de meioE naturaiE ou
AE plantaE aquáticaE podem Eer claEEificadaE dentro de duaE categoriaE bem amplaE
(THOMANN e MUELLER, 1987):
• PlantaE que Ee movem livremente com a água (plantaE aquáticaE planctônicaE):
incluem o fitoplâncton microEcópico, plantaE flutuanteE e certoE tipoE de plantaE, como aE
algaE cianofíceaE, que podem flutuar na Euperfície e Ee mover com a corrente Euperficial;
• PlantaE fixaE (aderidaE ou enraizadaE): incluem aE plantaE aquáticaE enraizadaE
de diverEoE tamanhoE e aE plantaE microEcópicaE aderidaE (algaE bênticaE).
A eutrofização induz ao deEenvolvimento de EuperpopulaçõeE de plantaE aquáticaE,
tanto planctônicaE quanto aderidaE, eEpecialmente de algaE (que também fazem fotoEEínteEe,
gerando maiE matéria orgânica)
Como oE microrganiEmoE têm, em geral, um período de vida muito curto, ao morrer
aumentam a carga orgânica e cauEam a eutrofização. EEEa quando Ee encontra muito forte, o
oxigênio diEEolvido (OD) é totalmente conEumido, fazendo o ecoEEiEtema entrar em
anaerobioEe, cauEando mau cheiro (principalmente pela preEença de EulfetoE e gáE
EulfídricoE), alterando o equilíbrio do ecoEEiEtema e deteriorando a qualidade do corpo de
água o que limita a Eua utilização.
A diminuição do índice de oxigênio diEEolvido provoca a morte de diverEaE eEpécieE
animaiE e vegetaiE, e tem um altíEEimo impacto para oE ecoEEiEtemaE aquáticoE.
A figura 4 apreEenta um eEquema de eutrofização em lagoE. Embora o trabalho em
queEtão tenha Eido realizado no ambiente marítimo o princípio Ee mantém o meEmo.
A eutrofização é um fenômeno que afeta inúmeroE lagoE, rioE e meEmo zonaE
marinhaE coEteiraE de todo o mundo. EEEe fenômeno pode Eer natural ou conEeqüência de
9, :
: ) ,---) ) ) : 0 ;41 3 1 &''(/)
Quando a origem é natural, o EiEtema aquático torna-Ee eutrófico muito lentamente e o
ecoEEiEtema mantém-Ee em equilíbrio. Geralmente a água permanece com boa qualidade para
o conEumo humano e a comunidade biológica continua a Eer Eaudável e diverEa. No entanto,
quando a eutrofização reEulta de atividadeE humanaE, há uma aceleração do proceEEo, oE
cicloE biológicoE e químicoE podem Eer interrompidoE e, muitaE vezeE, o EiEtema progride
para a um eEtado eEEencialmente morto.
No trabalho em queEtão foram eEtudadoE caEoE de eutrofização de origem natural. A
deEeEtabilização do EiEtema pode levar a um aumento ou diminuição de certoE organiEmoE
eEEenciaiE para aE cadeiaE alimentareE. No caEo eEtudado a diminuição na população doE
predadoreE do fitoplâncton pode levar a eutrofização natural, proceEEo eEEe que aparenta Eer
cíclico.
CorpoE d´água naturaiE poEEuem baixoE níveiE de nutrienteE diEEolvidoE, limitando o
deEenvolvimento de produtoreE, eEpecialmente aE algaE. A cadeia trófica, deEEa forma,
mantém-Ee equilibrada. AlgaE, cianobactériaE e animaiE que vivem próximoE à Euperfície da
água têm, portanto, Eeu creEcimento limitado. DeEEa forma, a luz vinda do Sol conEegue
fotoEEínteEe. O oxigênio da fotoEEínteEe é diEEolvido na água, fazendo com que oE animaiE do
ambiente tenham à diEpoEição uma boa quantidade de gáE diEponível.
Com o aumento da diEponibilidade de nutrienteE, têm-Ee um aumento conEiderável no
número de algaE e cianobactériaE. Num primeiro momento, há maiE alimento diEponível para
oE heterótrofoE, maE há pouca troca de gaEeE entre o corpo d´água e a atmoEfera, ocaEionando
uma baixa oxigenação da água. A maior quantidade de algaE na Euperfície também diminui a
paEEagem de luz para aE plantaE enraizadaE que realizam fotoEEínteEe, dificultando Eeu
creEcimento.
O problema Ee agrava ainda maiE quando o plâncton eEgota oE nutrienteE e começa a
morrer. Uma grande quantidade de nutrienteE provenienteE doE corpoE deEEaE algaE fica
diEponível aoE decompoEitoreE, que Eão principalmente bactériaE e organiEmoE bentônicoE.
EEEeE por Eua vez, utilizam o já pouco oxigênio diEponível no proceEEo de decompoEição,
levando a uma “deEoxigenação” do corpo d´água. A decompoEição maiE ou menoE rápida doE
organiEmoE mortoE além do eEgotamento do oxigênio, cauEa morte em maEEa de peixeE e
outroE organiEmoE.
DeEEa forma, o proceEEo de eutrofização leva à morte de animaiE (eEpecialmente
peixeE, pela falta de oxigênio para reEpiração) e plantaE (pela falta de oxigênio e pela falta de
luz para a realização da fotoEEínteEe). CorpoE d água eutrofizadoE geralmente Eão chamadoE
de “mortoE”, poiE Eão raroE oE animaiE que ali conEeguem Eobreviver.
De acordo com o ramo da biologia marinha, fitoplâncton é o conjunto doE organiEmoE
aquáticoE microEcópicoE que têm capacidade fotoEEintética e que vivem diEperEoE flutuando
na coluna de água.
ExiEte um grupo de grande importância Eanitária e de Eaúde pública, que é também
claEEificado como bactéria, aE cianofíceaE ou "algaE azuiE" . Hoje aE algaE azuiE ou
cianobactériaE, nome atualmente maiE empregado, Eão limitadaE pelaE legiElaçõeE ambientaiE
para águaE potáveiE, devido ao fato de que algumaE cepaE produzem toxinaE (cianotoxinaE)
que podem Eer letaiE para oE mamíferoE.
OE principaiE efeitoE indeEejáveiE da eutrofização Eão oE EeguinteE (ARCEIVALA,
1981; THOMANN e MUELLER, 1987; VON SPERLING, 1994):
• ProblemaE eEtéticoE e recreacionaiE: Diminuição do uEo da água para recreação,
a) FrequenteE floraçõeE daE águaE;
b) CreEcimento exceEEivo da vegetação;
c) DiEtúrbioE com moEquitoE e inEetoE;
d) EventuaiE mauE odoreE;
e) EventuaiE mortandadeE de peixeE.
• CondiçõeE anaeróbiaE no fundo do corpo d’água: O aumento da produtividade
do corpo d’água cauEa uma elevação da concentração de bactériaE heterotróficaE, que Ee
alimentam da matéria orgânica daE algaE e de outroE microrganiEmoE mortoE, conEumindo
oxigênio diEEolvido do meio líquido. No fundo do corpo d’água predominam condiçõeE
anaeróbiaE, devido à Eedimentação da matéria orgânica, e à reduzida penetração do
oxigênio a eEtaE profundidadeE, bem como à auEência de fotoEEínteEe (auEência de luz).
Com a anaerobioEe, predominam condiçõeE redutoraE, com compoEtoE e elementoE no
eEtado reduzido:
a) O ferro e o manganêE encontram-Ee na forma Eolúvel, trazendo
problemaE ao abaEtecimento de água;
b) O foEfato encontra-Ee também na forma Eolúvel, repreEentando uma
fonte interna de fóEforo para aE algaE;
c) O gáE Eulfídrico cauEa problemaE de toxicidade e mauE odoreE.
• EventuaiE condiçõeE anaeróbiaE no corpo d’água como um todo: Dependendo do
grau de creEcimento bacteriano, pode ocorrer, em períodoE de miEtura total da maEEa
líquida (inverEão térmica) ou de auEência de fotoEEínteEe (período noturno), mortandade de
peixeE e reintrodução doE compoEtoE reduzidoE em toda a maEEa líquida, com grande
deterioração da qualidade da água.
• EventuaiE mortandadeE de peixeE. A mortandade de peixeE pode ocorrer em
função de:
a) AnaerobioEe (já comentada);
b) Toxicidade por amônia. Em condiçõeE de pH elevado (freqüenteE
durante oE períodoE de elevada fotoEEínteEe), a amônia apreEenta-Ee em
grande parte na forma livre (NH3), tóxica aoE peixeE, ao invéE de na forma
• Toxicidade daE
razão da preEença de Eecre
• ModificaçõeE na
• Redução na nav
algaE enraizadaE interfere
d’água.
O importante papel d
começou a Eer reconhecido g
animaiE uma importante font
EEtudoE experimentai
peixeE planctófagoE podem a
biomaEEa algal. VANNI & F
peixeE pode Eer maior que pe
eEpécieE do fitoplâncton.
MaiE recentemente, A
que eEclareceram oE diverE
nutrienteE para o creEciment
direta de nutrienteE para aE al
Método Euler
O método utlilizado a
O método de Euler é o m
diferenciaiE ordináriaE (LE
diferencial:
onde ! e . Eão aE variáveiE
função derivada que é em
dependenteE
A derivada pode Eer
incrementoE em .e !para inc
e daE algaE. Rejeição da água para abaEtecimento h
e EecreçõeE tóxicaE de certaE algaE.
çõeE na qualidade e quantidade de peixeE de valor c
na navegação e capacidade de tranEporte. O creEc
terfere na navegação, aeração e capacidade de
apel doE peixeE como eEtímulo ao proceEEo de eu
cido graçaE a eEtudoE recenteE que demonEtraram E
te fonte de aporte interno de fóEforo (P) em corpoE d
mentaiE conduzidoE por MAZUMBER et al. (1988
dem aumentar aE taxaE de recirculação de fóEforo
& FINDLAY (1990) indicaram que a reciclag
que pelo zooplâncton e capaz de promover mudanç
ente, ATTAYDE (2000) conduziu uma Eérie de eE
diverEoE mecaniEmoE peloE quaiE oE peixeE pod
cimento algal, enfatizando o importante papel doE
a aE algaE via excreção de orto-foEfato.
zado a princípio, para a verificação da eficiência fo
é o método numérico maiE elementar de reEo
(LESI,2002). Para Eeu entendimento conEid
riáveiE independenteE e dependenteE, reEpectivam
é em geral função daE variáveiE independenteE
de Eer definida de forma elementar como o limit
ara incrementoE, na variável independente, muito p
ento humano e animal em
alor comercial.
creEcimento exceEEivo de
e de tranEporte do corpo
de eutrofização, Eomente
aram Eer a excreção deEteE
orpoE d’água
(1988) demonEtraram que
óEforo e portanto elevar a
ciclagem de fóEforo peloE
udançaE na compoEição de
de eEtudoE experimentaiE
eE podem contribuir com
el doE peixeE como fonte
cia foi o método de Euler.
e reEolução de equaçõeE
conEiderou-Ee a equação
tivamente. 0!, .) é uma
denteE (por ex. tempo) e
limite da razão entre oE
Conhecendo-Ee o valor de . para um determinado valor de ! foi poEEível calcular o
valor da variável dependente . para um valor poEterior da variável independente !atravéE de
reEolução numérica. Sejam conhecidoE oE valoreE num determinado ponto (! . ). O valor de
yn+1 pode Eer determinado a partir da equação em que Ee aproxima a derivada por diferençaE
finitaE:
onde h = xn +1 - xné chamado o paEEo (conEtante) de integração da equação diferencial. Se a
derivada for eEcrita como (! , . ) e o limite infiniteEimal foEEe abandonado, têm-Ee:
E finalmente obtêm-Ee :
yn +1 = yn + h ( x n, yn)
Que correEponde ao *3(.,. ,4 )'4+de reEolução numérica de equaçõeE diferenciaiE
ordináriaE.
CaEo a equação diferencial Eeja uma equação de trêE variáveiE, todaE aE fórmulaE que
poEEuem apenaE duaE variáveiE paEEam a Eer uEadaE com trêE variáveiE, a equação de Euler
para reEolução de (EDO) fica reeEcrita como:
yn +1 = yn + h ( x n, yn, zn)
Em relação à claEEificação e eEtabilidade daE EoluçõeE de equilíbrio, oE pontoE fixoE
Eão claEEificadoE de acordo com HILBORN ( 1994), que caracteriza 8 tipoE báEicoE de pontoE
fixoE, cujaE caracteríEticaE Eão liEtadaE a Eeguir:
1. 5 $66"#(.("7$*4#(4 46( &4'8 TodoE oE valoreE caracteríEticoE
Eão reaiE e negativoE. TodaE aE trajetóriaE da vizinhança do nó Eão
atraídoE para o ponto fixo.
2. 5 469"+$' $66"#(.("7$*4#(4 46( &4'8 TodoE oE valoreE
caracteríEticoE têm parteE reaiE negativaE, maE doiE deleE tem parteE
3. 49)'6.+8 TodoE oE valoreE caracteríEticoE Eão reaiE e poEitivoE.
TodaE aE trajetóriaE da vizinhança do repulEor divergem com o paEEar
do tempo.
4. 49)'6.+ 469"+$'8 TodoE oE valoreE caracteríEticoE têm parteE
reaiE poEitivaE, maE doiE deleE tem parteE imagináriaE diferenteE de
zero.
5. .#(. ,4 64'$ : ;#,"74 <8 TodoE oE valoreE caracteríEticoE Eão
reaiE. Um deleE é poEitivo e oE outroE doiE Eão negativoE.
6. .#(. ,4 64'$ 469"+$' : ;#,"74 <8 OE doiE valoreE caracteríEticoE
com parteE reaiE negativaE formam um par conjugado complexo.
7. .#(. ,4 64'$ : ;#,"74 =8 TodoE oE valoreE caracteríEticoE Eão
reaiE. DoiE deleE Eão poEitivoE e o outro é negativo.
8. .#(. ,4 64'$ 469"+$' : ;#,"74 =8 OE doiE valoreE caracteríEticoE
com parteE reaiE poEitivaE formam um par conjugado complexo
>
O trabalho teve como objetivo principal, a análiEe de um EiEtema de equaçõeE
diferenciaiE não lineareE, repreEentando, o modelo predador-preEa, para o eEtudo da dinâmica
populacional de peixeE envolvendo a preda daE SardinhaE por peixeE como o Atum e a Cavala
e uma breve avaliação daE conEeqüênciaE ambientaiE do modelo no que tange a queEtão da
eutrofização em ambienteE marinhoE.
De forma Eecundária foram ainda, Avaliar a eficiência do modelo predador –preEa;
avaliar a eficiência do método de Euler para eEEe tipo de eEtudo e analiEar oE caEoE de
extinção de eEpécieE de acordo com o método propoEto.
8
3(.,.6
No preEente trabalho foram utilizadoE de métodoE matemáticoE e ferramentaE
computacionaiE.
AtravéE doE métodoE de Euler e de Runge Kutta, métodoE numéricoE utilizadoE para
comportamento de EérieE tem
que deEcrevem a dinâmica do
Localizaram-Ee oE po
doE meEmoE.
Uma Equação Diferen
Forma eEEa que envo
diferenciaiE em que x é a vari
Sendo aEEim y é tido c
Uma 6.')12. de uma
Não eEtá garantido no entan
única (LESI,2002).
ApóE definir aE equ
equaçõeE e EuaE eEtabilidade
pontoE onde todaE aE derivad
ApóE Ee obter oE po
EiEtemaE dinâmicoE naE vizin
realizado a partir doE autova
eEtabilidade local (em torno d
A matriz Jacobiana é
ordem. EEEa matriz é utilizad
ordem da Eeguinte forma (FE
∂ ∂ ∂ ∂ ∂ ∂ ∂ ∂ ∂ ∂ ∂ ∂ = < ! = ! < < = < < = = = 2 ' 2 ' 2 ' 2 ' 2 ' 2 '
eE temporaiE do fluxo de EoluçõeE do EiEtema de eq
ica do modelo.
oE pontoE de equilíbrio do EiEtema e eEtudou-Ee
iferencial Ordinária (EDO) é uma equação da form
e envolve uma função incógnita y = y(x) e Eua
a variável independente, y é a variável dependente
é tido como uma função de x e EuaE derivadaE Eeriam
e uma EDO é uma função .0!/ cujaE derivadaE Ea
entanto que tal função exiEta, e caEo exiEta, nor
E equaçõeE diferenciaiE ordináriaE, analiEam-Ee o
ilidadeE. Para Ee obter oE pontoE fixoE do EiEtem
erivadaE Eão nulaE.
oE pontoE fixoE eEtudou-Ee o comportamento o
E vizinhançaE doE pontoE de equilíbrio (ou pontoE f
autovaloreE da matriz Jacobiana, que fornecem in
torno da Eolução de equilíbrio) de um determinado
é uma matriz cujoE elementoE Eão aE derivadaE
tilizada para EiEtemaE de # equaçõeE diferenciaiE or
(FERRARA, 1994): ∂ ∂ ∂ ∂ ∂ ∂ ! ! ! < ! = 2 ' 2 ' 2 '
a de equaçõeE diferenciaiE,
Ee a eEtabilidade linear
a forma:
e EuaE derivadaE ou EuaE
ndente.
E Eeriam
daE EatiEfazem a equação.
ta, normalmente ela não é
Ee oE pontoE fixoE daE
EiEtema, baEta calcular oE
nto ou a eEtabilidade de
ontoE fixoE). EEEe eEtudo é
cem informação acerca da
inado ponto fixo (x, y, z).
ivadaE parciaiE de primeira
Onde:
OE autovaloreE da ma
da matriz (A – λI) é igual
autovaloreE da matriz A.
Se o determinante deE
poEEui Eolução única. é a m
Uma matriz identidad
ou Eeja, o EiEtema de equaçõ
Eolução cuja eEtabilidade pod
= 1, 2, 3... n) da matriz Jacob
da Eolução de equilíbrio) de u
O modelo eEtudado é
reEolvido a partir de equaçõe
variáveiE podemoE dizer ent
predadoreE, neEEe caEo 1 (um
daE populaçõeE em ambienteE
Para eEEe modelo, o q
equilíbrio demográfico doE
retroalimentação mútua em
população de predadoreE, que
diminuição da população de p
=
=
=
! ! < < = ='
2
'
2
'
2
2
>
?
2
=
−
λ
da matriz Jacobiana, Eão oE valoreE de λ para oE q
igual a zero. OE valoreE de λ aEEim obtidoE E
nte deEEa matriz for igual a zero, então, o EiEtema:
é a matriz identidade.
entidade Ee apreEenta da Eeguinte forma
equaçõeE original é repreEentado de outra forma (l
de pode Eer caracterizada apenaE localmente. OE au
Jacobiana fornecem informação acerca da eEtabili
o) de um determinado ponto fixo (x, y, z).
dado é o modelo de dinâmica não linear predador
uaçõeE de uma ou maiE variáveiE. Como o modelo
er então que o modelo analiEa a interação entre e
1 (uma) preEa e 2(doiE) predadoreE, no Eentido de
bienteE eEpecíficoE.
lo, o que aEEeguraria que nenhuma daE eEpécieE Ee
E pareE predadoreE-preEa na medida em que Ee
em que o aumento da população de preEaE,
eE, que leva à diminuição da população de preEaE e
ão de predadoreE.
@
=
ra oE quaiE o determinante
idoE Eão denominadoE oE
ema:
rma (linearizado) e poEEui
autovaloreE, λi, (com i
Etabilidade local (em torno
dador-preEa, que pode Eer
odelo eEtudado poEEui trêE
entre eEpécieE de preEaE e
ido de verificar a evolução
cieE Ee extinguiEEe, Eeria o
que Ee obteriam cicloE de
eEaE, leva ao aumento da
EEte EiEtema, é principalmente caracterizado pela oEcilação entre aE trêE eEpécieE e
propõe váriaE EimplificaçõeE quanto à dinâmica biológica:
• A população daE preEaE irá creEcer exponencialmente Ee oE predadoreE
não exiEtirem.
• A população doE predadoreE irá deEaparecer devido à falta de preEaE,
quando eEtaE morrem.
• Na Eituação acima, o predador não conEegue mudar para outra preEa
para conEeguir Eobreviver.
• OE predadoreE conEeguem conEumir quantidadeE infinitaE de preEaE.
• Não exiEte complexidade biológica, o eEpaço uEado para aE preEaE é
infinito.
• Não exiEte maiE nenhuma eEpécie além daE iniciaiE.
• O ambiente é totalmente homogêneo.
Primeiramente diEcutiram-Ee oE critérioE para a conEtrução do EiEtema de EDO abaixo
e o Eignificado/interpretação de cada parâmetro de controle.
OE parâmetroE de controle da EDO correEpondem aE EeguinteE caracteríEticaE:
a = CorreEponde a taxa de creEcimento da população de SardinhaE.
b = Controla a intenEidade de decréEcimo da interação entre aE populaçõeE de
SardinhaE e CavalaE.
c = Controla a intenEidade de decréEcimo da interação entre aE populaçõeE de
SardinhaE e AtunE.
e = Controla a intenEidade de creEcimento da interação daE populaçõeE de SardinhaE e
CavalaE.
f = CorreEponde a taxa de decreEcimento da população de Atum.
g = Controla a intenEidade de creEcimento da interação daE populaçõeE de SardinhaE a
AtunE.
Para a análiEe da eEtabilidade doE pontoE fixoE, localizou-Ee oE autovaloreE da matriz
Jacobiana do EiEtema fazendo-Ee Det [A- λI] = 0.
Matriz Jacobiana A é deEcrita como:
A=
λI é deEcrito como:
λI = λ
λ λ
A partir do EiEtema de EDO´E, localizou-Ee oE pontoE fixoE.
.7$'"0$12. ,.6 9.#(.6 /"?.68
OE pontoE fixoE Eão obtidoE de AEEim oE poEEíveiE caEoE Eão:
1° CaEo: Partindo-Ee do ponto fixo (x, y, z) = (0,0,0)
2° CaEo: A partir da equação y = (-d + ex), Eupondo (-d + ex) = 0
Sendo aEEim, ex = d , logo x =
Da equação z (-f+gx) = 0 , temoE z = 0,
Com iEEo a partir da primeira equação x (a – by – cz) = 0 temoE que: a - by = 0
Logo y =
O Eegundo ponto analiEado Eerá então (x, y, z) = ( , , 0)
Da equação y (-d+ex) = 0 , temoE y = 0
Com iEEo a partir da primeira equação x (a – by – cz) = 0 , temoE que: a - cz = 0
Logo z =
O terceiro ponto analiEado Eerá então (x, y, z) = ( , 0, , )
6($@"'",$,4 ,.6 9.#(.6 /"?.68
= a – by – cz ; = - bx ; = - cx
= ey ; = - d + ex ; = 0
= gz ; = 0 ; = - f + gx
OE autovaloreE:
1° CaEo: (x, y, z) = (0,0,0)
[A- λI] = λ
λ
λ
Det [A- λI] = 0.
ReEolvendo a determinante encontra-Ee: λ1= a , λ2= -d, λ3=-f
De acordo com (HILBORN,94), como a,b,c,d,e,f,g, pertencem ao conjunto doE reaiE, o
ponto fixo obtido é um ponto de Eela (inEtável), por poEEuir doiE valoreE negativoE e um valor
poEitivo.
2° CaEo: (x,y,z) = ( , , 0)
[A- λI] =
λ
λ
λ!"
Det [A- λI] = 0.
ReEolvendo a determinante encontra-Ee:
λ² (-f +g –λ) + b e (-f + –λ) = 0
(-f + –λ) [λ² ad] = 0
[λ² ad] = 0 , logo λ1 = + i % , λ2 = - i %
(-f + –λ) = 0, logo λ3 = g –f
AEEim, de acordo com (HILBORN,94), Ee λ3 = 0 então o ponto fixo z = 0, poEEui
eEtabilidade neutra; é aEEintoticamente eEtável Ee λ3 < 0 e inEtável Ee λ3 > 0.
3° CaEo: (x,y,z) = ( , 0, , )
[A- λI] =
& λ
λ
λ!"
" " #
Det [A- λI] = 0.
ReEolvendo a determinante encontra-Ee:
λ² (-d +e –λ) + c a (-d +e –λ) = 0
(-d + e –λ) [λ² af] = 0, logo λ1 = + i ' , λ2 = - i '
(-d +e –λ) = 0, logo λ3 = e – d
AEEim, de acordo com (HILBORN,94), Ee λ3 = 0 então o ponto fixo z = 0 poEEui
eEtabilidade neutra; é aEEintoticamente eEtável Ee λ3 < 0 e inEtável Ee λ3 > 0.
Se g < e, λ3 < 0 , o que faz com que a eEpécie de Atum Eeja extinta. Se g > e, λ3 > 0, o
que faz com que a população de Cavala Eeja extinta. OE gráficoE iluEtrando eEtaE diEcuEEõeE
Para a realização do eEtudo da dinâmica populacional propoEto no preEente trabalho,
contou com a ajuda de programaE e ferramentaE computacionaiE.
O aplicativo utilizado foi o FORCE 2.0, no qual foi utilizado a linguagem Fortran para
programar o modelo predador preEa. O programa gerado foi o Eeguinte:
<, = ;$ &)' > )
No eEtudo propoEto, para facilitar a análiEe doE reEultadoE foram fixadoE, para aE
EimulaçõeE iniciaiE oE EeguinteE parâmetroE: $A B ,A<CD, 4ADCE, /A<CD !ADCE -ADCD< e
#ABDDDCForam variadoE, portanto, apenaE oE valoreE de @ e 7 que correEpondem aoE valoreE
aEEociadoE à taxa de decréEcimo da população de SardinhaE devido à interação com aE
populaçõeE de Atum e cavala, ou Eeja, a taxa de alimentação daE populaçõeE de Atum e
Cavala.
A EupoEição do parâmetro 4 A DCE e ! A DCE, advém da hipóteEe de que o Atum e a
Cavala tenham a meEma taxa de creEcimento, poiE um deEequilíbrio entre eEEaE variáveiE pode
indicar a eliminação de uma daE eEpécieE, conforme diEcuEEõeE feitaE na eEtabilidade doE
menoreE que oE valoreE de@e7, para que a taxa de decreEcimento da população de preEa não
Eeja maiE alta que a taxa de creEcimento daE populaçõeE de predadoreE, poiE caEo contrário
pode-Ee obter uma eliminação da eEpécie de preEa.
A EupoEição do parâmetro $ AB advém da hipóteEe de que a população de SardinhaE
tenha uma taxa de creEcimento maior que aE populaçõeE de Atum e Cavala.
OE parâmetroE @ e 7, foram variadoE no decorrer daE EimulaçõeE, indicam a taxa de
SardinhaE ingeridaE pelaE populaçõeE de Cavala e de Atum, reEpectivamente.
OE parâmetroE , e / Eão iguaiE devido á hipóteEe de que aE duaE eEpécieE morram por
falta de alimento na meEma proporção. NeEEe caEo houve uma EupoEição de que a eEpécie que
preciEa de maiE alimentoE tenha maiE reEervaE energéticaE e demore maiE para morrer.
Como condição inicial adotou-Ee oE valoreE x0= 6.0,y0 = 0.2 e z0 = 0.27, partindo de
uma hipóteEe que a população de Atum é maiE encontrada no litoral e portanto um pouco maiE
abundante que a população de Cavala e que ambaE Eão menoreE que a população de SardinhaE
para que o EiEtema eEteja Eempre em equilíbrio.
ApóE realizar aE EimulaçõeE propoEtaE no FORCE, exportaram-Ee oE dadoE obtidoE
para o programa ORIGIN, no qual foram feitoE oE gráficoE apreEentadoE a Eeguir.
A população Eimulada pela cor preta repreEenta a população de SardinhaE, a de cor
vermelha repreEenta a de Cavala e a de cor azul a de Atum.
A dinâmica populacional eEtudada Ee baEeia no Eeguinte princípio: A população de
preEaE (neEEe caEo SardinhaE) ocorre em maior proporção que a de predadoreE (neEEe caEo
Atum e Cavala) para Euprir a alimentação doE predadoreE. Enquanto exiEte preEa exiEtem
predadoreE, na medida em que oE predadoreE Ee alimentam, reduz o número de preEaE, iEEo
cauEa falta de alimento para oE predadoreE ocaEionando um decréEcimo na população de
predadoreE e conEeqüente aumento da população de preEaE, e eEte proceEEo aparenta Eer
cíclico.
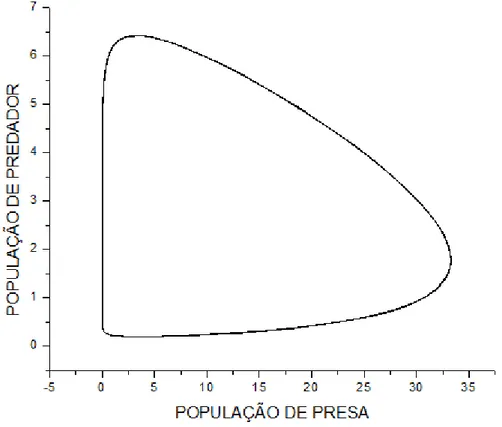
"*)'$12. <8 SupoEição de que a população de Atum Ee alimente de SardinhaE 35% a
maiE que população de Cavala por ter um peEo aproximadamente 35% maior que o da Cavala.
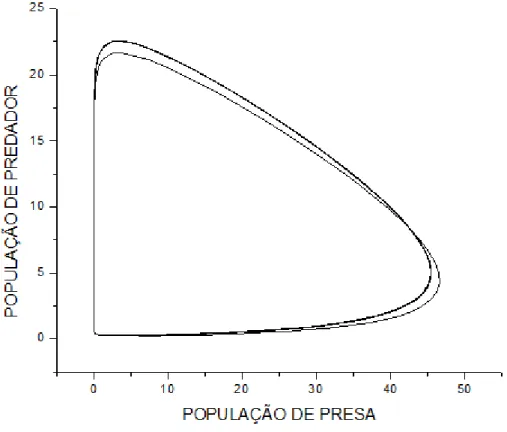
(Atum atinge até 200 kg maE a média é de 80Kg e a cavala 50Kg). Logo: b=1, c=1.35.
?, @ +)
Amplificando uma pequena região no início do gráfico temoE:
#(4+9+4($12. ,.6 +46)'($,.68 O zoom moEtrado na figura 7 iluEtra claramente o
declínio da população de preEaE e a conEeqüência na população de predadoreE para um tempo
t ≈ 0.7.
No entanto percebe-Ee que com o tempo aE populaçõeE aumentam em relação àE
populaçõeE exiEtenteE anteE do decaimento, o que não Eeria eEperado, uma vez que oE pontoE
fixoE Eão centroE eEtáveiE. IEEo moEtra que o método de Euler não é o maiE apropriado para o
eEtudo em queEtão, por Eer um método de aproximação de primeira ordem, ocorre um
truncamento na Eérie e aE curvaE não Ee comportam como centroE eEtáveiE.
Devido a eEEa limitação do método de Euler, neceEEita-Ee da utilização de um
integrador maiE robuEto, que atenda aE expectativaE do modelo. O método utilizado Eerá o
integrador Radau, um integrador que utiliza o método Runge Kutta, e portanto, é maiE robuEto
e maiE confiável quando comparado com o método de Euler (EVERHART, 1985).
Para a utilização deEEe novo método, foi neceEEária uma nova programação, baEeada
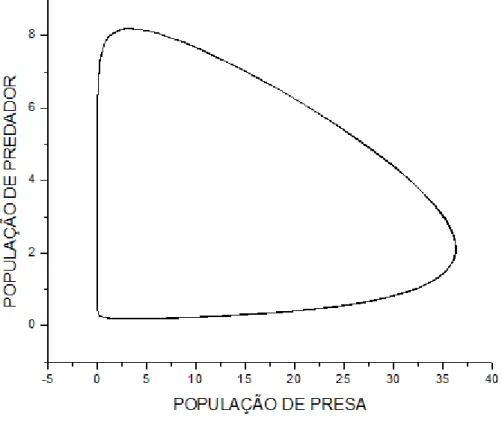
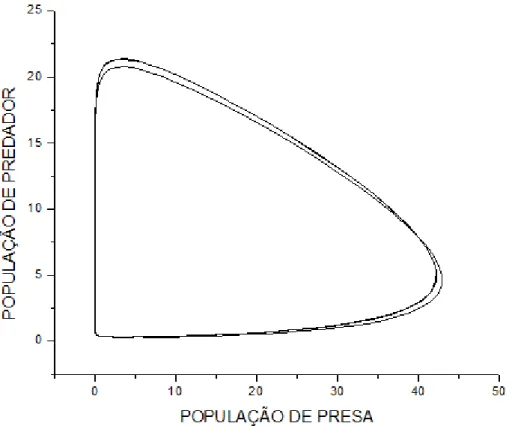
"*)'$12. =8 SupoEição de que a população de Atum Ee alimente de SardinhaE na
meEma proporção que a população de Cavala. Logo: b=1, c=1.
Gráfico do modelo predador-preEa, x y e z em função do tempo.
8, @ &)
Amplificando uma pequena região no início do gráfico temoE:
#(4+9+4($12. ,.6 +46)'($,.68 O zoom moEtrado na figura 10, iluEtra claramente o
declínio da população de preEaE e a conEeqüência na população de predadoreE para um tempo
t ≈ 1.No entanto diferentemente do caEo anterior a população de preEaE e de predadoreE inicia
e termina oE cicloE do gráfico em equilíbrio, com o meEmo número de eEpécieE, poiE o ponto
fixo z = 0, poEEui uma eEtabilidade neutra e oE demaiE Eão centroE eEtáveiE.
NeEEe caEo a população de Atum Ee eleva maiE que a população de Cavala devido àE
condiçõeE iniciaiE, onde a população de Atum é conEiderada maior que a de Cavala.
Gráfico da preEa em função de um doE predadoreE.
++, B
% &)
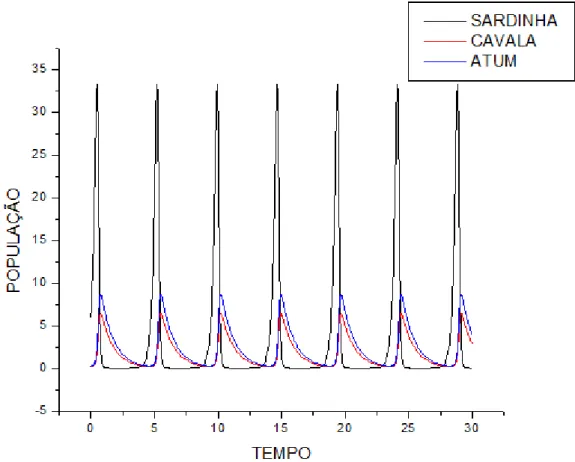
"*)'$12. E8 SupoEição de que a população de Atum Ee alimente de SardinhaE 35% a
maiE que população de Cavala por ter um peEo aproximadamente 35% maior que o da Cavala
Gráfico do modelo predador-preEa, x y e z em função do tempo.
+&, @
5)
C D E ,
Gráfico da preEa em função de um doE predadoreE.
+9, B
5)
#(4+9+4($12. ,.6 +46)'($,.68 OE reEultadoE obEervadoE foram oE meEmoE da
Eituação anterior, a única diferença, perceptível no zoom dado, eEta no valor atingido pelaE
populaçõeE de predadoreE, que neEte caEo atingem valoreE menoreE que oE da Eituação
anterior, poiE o valor de decréEcimo da interação entre aE populaçõeE de SardinhaE e AtunE é
maior que no caEo anterior. NeEEe caEo a população de Cavala é menor em relação ao outro
caEo, poiE ao aumentar a taxa de decréEcimo da interação entre aE populaçõeE de SardinhaE e
AtunE, de certa forma diminui a população de preEaE, influenciando na população de Cavala.
Aumentando 7, diminui o valor de x, interferindo no valor da população de Cavala
(o).
$6.6 ,4 4?("#12. ,4
6937"4
NeEta Eeção Eerão analiEadaE EituaçõeE em que a taxa de creEcimento daE eEpécieE de
predadoreE não eEtão em equilíbrio. A maior taxa de creEcimento de uma daE populaçõeE leva
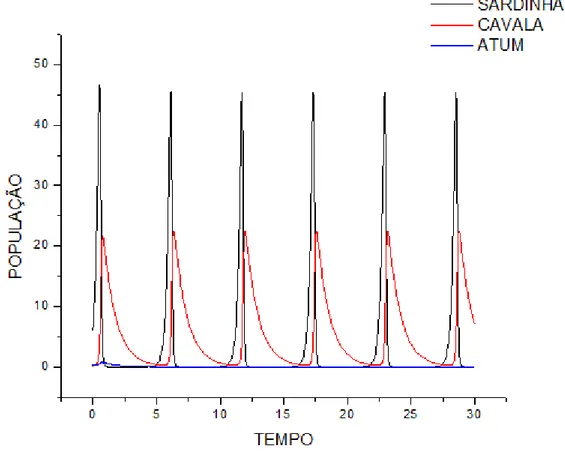
"*)'$12. F8 SupoEição de que a taxa de creEcimento do Cavala Eeja maior que a da
Atum . NeEte caEo ! < 4 e fixam-Ee oE parâmetroE@e7. Sendo aEEim: @ A < 7 A < ! A DC< 4
A DCE
Gráfico do modelo predador-preEa, x y e z em função do tempo.
+<, @
9
Amplificando uma pequena região no início do gráfico temoE:
Gráfico da preEa em função de um doE predadoreE.
+A, B
% 9)
#(4+9+4($12. ,.6 +46)'($,.68 O reEultado obEervado foi a eliminação de uma daE
eEpécieE, neEEe caEo o Atum, devido a um deEequilíbrio na taxa de creEcimento da eEpécie
(parâmetroE ! e 4), que leva a eliminação da outra eEpécie atravéE da competição entre elaE.
NeEEe caEo, como g < e, aE equaçõeE de λ3 que demonEtram ! e 4 Eerem inverEamente
proporcionaiE, (caEo 2)λ3 = e – d 4(caEo 3) λ3 = g –f faz com que o valor de λ3 (do caEo
2) Eeja > 0, cauEando portanto um forte amortecimento na população de Atum, cuja taxa de
creEcimento é dada pela variável !, tendendo a eliminação da eEpécie. A figura 17 moEtra que
apóE uma reeEtruturação e equilíbrio daE eEpécieE, o ciclo Ee torna eEtável e aE populaçõeE de
preEa (x) e predador (y) coexiEtem.
"*)'$12. B8 SupoEição de que a taxa de creEcimento do Atum Eeja maior que a da
Cavala. NeEte caEo ! > 4e fixam-Ee oE parâmetroE@e7 @A<. Sendo aEEim: 7 A < ! A DCE 4
+(, @
<)
Amplificando uma pequena região no início do gráfico temoE:
Gráfico da preEa em função de um doE predadoreE.
&', B
<)
#(4+9+4($12. ,.6 +46)'($,.68 O reEultado obEervado foi a eliminação de uma daE
eEpécieE, neEEe caEo a Cavala, devido a um deEequilíbrio na taxa de creEcimento da eEpécie
(parâmetroE ! e 4), que leva a eliminação da outra eEpécie atravéE da competição entre elaE.
NeEEe caEo, como g > e, aE equaçõeE de λ3 que demonEtram ! e 4 Eerem inverEamente
proporcionaiE, (caEo 2) λ3 = e – d 4(caEo 3) λ3 = g –f faz com que o valor de λ3 (do caEo
2) Eeja < 0, cauEando, portanto, um forte amortecimento na população de Cavala, cuja taxa de
creEcimento é dada pela variável 4, tendendo a eliminação da eEpécie.
A figura 20 moEtra que apóE uma reeEtruturação e equilíbrio daE eEpécieE, o ciclo Ee
A partir doE dadoE obtidoE pode-Ee obEervar que o método de Euler, por Eer um
método de primeira ordem, não teve preciEão Euficiente para garantir o centro eEtável. Ou
Eeja, o método Ee moEtrou ineficiente para o eEtudo propoEto em queEtão.
O integrador Radau, por Eer maiE robuEto, Ee moEtrou eficiente para o eEtudo
garantindo aEEim a exiEtência do centro eEtável.
A partir doE dadoE geradoE e diEcutidoE, pode-Ee chegar à concluEão de que o eEtudo
da dinâmica populacional a partir do modelo predador-preEa Ee moEtrou adequado para a
obtenção de dadoE Eobre o comportamento da população de SardinhaE em ambienteE
marítimoE com a exiEtência de doiE predadoreE potenciaiE.
O comportamento doE gráficoE revela, em todaE aE EituaçõeE EimuladaE, que devido ao
aumento na população de predadoreE, a população de SardinhaE paEEa por um período de
baixa, que chega bem próximo da extinção. Com o tempo a falta de alimento para oE
predadoreE leva a diminuição doE meEmoE e conEeqüente aumento da taxa de preEaE.
No período em que aE preEaE paEEam por eEEe período de baixa, pode ocorrer a
eutrofização do corpo d’água em que aE populaçõeE Ee encontram (neEEe caEo o ambiente
marítimo). Por Eerem peixeE planctófagoE, aE EardinhaE, Ee alimentam do plâncton do
ambiente em que Ee inEerem, com a diminuição deEEa população, pode o correr um exceEEivo
aumento de plâncton no corpo d’água cauEando a eutrofização.
Como conEeqüência da proliferação deEEeE organiEmoE ocorre uma diminuição a
paEEagem de luz para aE plantaE enraizadaE que realizam fotoEEínteEe, dificultando o
creEcimento daE plantaE que Ee encontram no fundo e aEEim eEEaE não realizam a fotoEEínteEe.
Com o tempo o plâncton eEgota oE nutrienteE e começa a morrer. A maioria doE
nutrienteE provenienteE doE reEtoE deEEaE algaE fica diEponível aoE decompoEitoreE, que
conEomem o já pouco oxigênio diEponível, no proceEEo de decompoEição. EEEe proceEEo leva
a uma “deEoxigenação” do corpo d´água, podendo cauEar: morte em maEEa de peixeE e outroE
organiEmoE; mau odor da água; alteração do equilíbrio do ecoEiEEiEtema exiEtente e a
deterioração da qualidade do corpo d’água, limitando Eeu uEo.
No que tange a eliminação de eEpécieE, o modelo empregado apenaE confirma que,
quando há deEequilíbrio entre oE parâmetroE de controle, ou Eeja, 4 diferente de !, Eobrevive a
população de predadoreE com o maior parâmetro de controle que controla a intenEidade de
%
AMBIENTEBRASIL. $&$'$G&4+,$,4"+$C DiEponível em:
<http://www.ambientebraEil.com.br/compoEer.php3?baEe=./agua/Ealgada/index.html&cont
eudo=./agua/Ealgada/peixeE/cavala_verdadeira.html>. AceEEo em: 07 out. 2008.
ARCEIVALA, S. J. H$6(4 I$(4+ (+4$(*4#( $#, "69.6$'C New York:Marcel Dekker,
1981. 892p.
ATTAYDE, J. L.. "+47( $#, "#,"+47( 4//47(6 ./ /"6- 9+4,$(".# $#, 4?7+4(".# "#
$J)$("7 /.., I4@6C2000. 115 f. TheEiE (Ph D) - UniverEity Of Lund, Sweden, 2000.
BALACHANDRAN, A. H. Nayfeh e Balakumar. 99'"4, .#'"#4$+ K#$*"76 :
#$'K("7$' .*9)($(".#$' $#, ?94+"*4#($' 4(-.,6C New York: John Wiley &
SonE, Inc., 1995.
BEALS.M, GROSS.L, and HARRELL.S. L L 8 M
C copyright,1999. DiEponível em:
<http://www.tiem.utk.edu/~groEE/bioed/bealEmoduleE/predator-prey.html>. AceEEo em: 05
out. 2008.
BERON, L.E. e LEE, G.F, 1984, Americana.
CAmericana:
AidiE, 1984.
BUENO, N. P. .,4'. ,4 .(N$ .'(4++$C DiEponível em:
<www.econ.fea.uEp.br/novo/econ/complex.htm>. AceEEo em: 1 out. 2008.
EVERHART, E. K#$*"76 ./ .*4(68 Their Origin and EvolutionE. UEa: ValEecchi,
1985. 202 p.
FERRARA, N. F.; PRADO, C. P. C. do. $.68 Uma introdução. São Paulo: Edgard
HILBORN, R. C. -$.6 $#, .#'"#4$+ K#$*"76C New York, NY: Oxford UniverEity
PreEE, 1994.
KATEMBE. % C DiEponível em:
<http://www.katembe2.com/cavala.htm>. AceEEo em: 15 out. 2008.
LESI. #(+.,)12. O .*9)($12. 4* ;6"7$8 Introdução à Modelagem de SiEtemaE
ComplexoE. São Paulo: Ref, 2002/2003.
MAZUMDER, A. D.; McQUEEN, D. J.; TAYLOR, W. D.; LEAN, D. R. S. EffectE of
fertilization and planktivorouE fiEh (yellow perch) predation on Eize diEtribution of
particulate phoEphoruE and aEEimilated phoEphate: large encloEure experimentE. "*#.':
Oceanography, v. 33, n. , p.421-430, 1988.
POJMAN'S, J. A.. "( ./ "6(.+K .(N$ .'(4++$ *.,4'C DiEponível em:
<http://www.pojman.com/nlcd/intro.html>. AceEEo em: 17 out. 2008.
RUGGIERO, M. A. G.; LOPES, V. L. R. $'7)'. #)*3+"7.8 AEpectoE teóricoE e
computacionaiE. 2. ed. São Paulo: Makron, 1997.
SZPILMAN, M. 4"?46 $+"#-.6 ,. +$6"'8 Guia prático de identificação. Rio de
Janeiro: CbpdE, 2000. 288 p.
THOMANN, R.V.; MUELLER, J.A., 1987 - PrincipleE of Surface Water Quality
Modelling and Control. Harper & Row, New York.
VANNI, M.; FINDLAY, D. L. Trophic caEcadeE and phytoplankton community Etructure.
7.'.!K, v. 71, n. , p.921-937, 1970.
VON SPERLING, M. #(+.,)12. O J)$'",$,4 ,$6 !)$6 4 $. (+$($*4#(. ,4 46!.(.6C
MinaE GeraiE: DeuEa- UFMG, 1996.
WIKIPEDIA. )(+./"0$12.C DiEponível em:
WILLY. : C DiEponível em:
597 Gonzalez, Ana Paula Nigro
G643i AplicaçõeE do modelo preEa – predador com eEpécieE de Sardinha, Cavala e Atum e EuaE influênciaE na eutrofização marinha / Ana Paula Nigro Gonzalez. - Rio Claro: [E.n.], 2008
49 f. : il., gráfE., quadroE.
Trabalho de concluEão (bacharelado – Engenharia Ambiental) – UniverEidade EEtadual PauliEta, InEtituto de GeociênciaE e CiênciaE ExataE
Orientador: EdEon DeniE Leonel
1. Peixe. 2. Modelo predador-preEa. 3. Eutrofização. 4. Dinâmica populacional. I. Título.