ISSN 0103-8478
Elizabeth Ann Veasey
I*Fernando Angelo Piotto
IWellington Ferreira do Nascimento
IJucelene Fernandes Rodrigues
IThiago Fonseca Mezette
IAline Borges
IIFelipe Almeida Biguzzi
IIFernanda Raquel Camilo dos Santos
IIIGraciela da Rocha Sobierajski
IVGustavo Henrique Recchia
IIJulio César Mistro
VProcessos evolutivos e a origem das plantas cultivadas
Evolutionary processes and the origin of crop plants
IDepartamento de Genética, Escola Superior de Agricultura “Luiz de Queiroz” (ESALQ), Universidade de São Paulo (USP). Av. Pádua Dias, 11, 13400-970, Piracicaba, SP, Brasil. E-mail: [email protected]. *Autor para correspondência.
IICentro de Energia Nuclear na Agricultura, Universidade de São Paulo (CENA/USP), Piracicaba, SP, Brasil.
IIICentro de Pesquisa e Desenvolvimento em Recursos Genéticos Vegetais, Instituto Agronômico/Agência Paulista de Tecnologia dos Agronegócios (IAC/APTA), Campinas, SP, Brasil.
IVCentro APTA Frutas, IAC/APTA, Jundiaí, SP, Brasil. VCentro de Café - IAC/APTA, Campinas, SP, Brasil.
RESUMO
A evolução das plantas cultivadas, que teve início há cerca de 13.000 anos, está sujeita aos mesmos processos evolutivos naturais, aliada à ação do homem de forma consciente ou inconsciente, levando à domesticação. Nesta revisão, são apresentados os principais fatores evolutivos, tais como mutação, hibridação, migração, seleção e deriva genética, que, de alguma maneira, estão envolvidos com a origem, evolução e domesticação de plantas cultivadas. São apresentados também exemplos de como esses processos influenciaram na diversidade intra e interespecífica de plantas cultivadas, com o aparecimento de novas variedades ou mesmo de novas espécies. De modo geral, tais processos atuaram na ampliação, na manutenção, bem como na redução da variabilidade genética das plantas cultivadas.
Palavras-chave: domesticação de plantas, mutação,
hibridação, migração, seleção, deriva genética.
ABSTRACT
The evolution of crop plants, which began at about 13,000 years ago, is subject to the same natural evolutionary processes, coupled with the action of man, consciously or unconsciously, leading to domestication. This review presents the main evolutionary factors such as mutation, hybridization,
migration, selection and genetic drift, which somehow are involved in the origin, evolution and domestication of crop plants. Examples of how these processes influenced in the intra and interespecific diversity of crop plants, with the uprise of new varieties or even of new species, are also presented. In general, these processes have worked well in the increase, maintenance, as well as in the reduction of genetic diversity of crop plants.
Key words: plant domestication, mutation, hybridization,
dispersion, selection, genetic drift.
INTRODUÇÃO
No Pleistoceno recente, pouco mais de
10,000 anos a.C., as primeiras populações humanas se
organizaram numa sociedade de caçadores e coletores
antes de iniciarem a prática agrícola no período Neolítico
(DIAMOND & BELLWOOD, 2003). Estas eram
caracterizadas por se deslocarem com maior frequência,
realizando o manejo dos recursos naturais, protegendo
espécies de interesse, podendo ainda alterar a
densidade e distribuição de espécies (BELLWOOD,
2005).
-Com o advento da agricultura, considerada
como um pré-requisito para o surgimento das
civilizações, acompan h ada de uma gr an de
transformação demográfica global (DIAMOND, 2002),
iniciou-se o processo de domesticação de plantas
cultivadas, sob a ação dos mesmos processos
evolutivos que ocorreram ao longo da evolução das
plantas em geral (LADIZINSKY, 1998). O diferencial,
no entanto, foi a ação do homem, que, de forma
consciente ou inconsciente, acabou tornando-as
dependentes da interferência humana e dos ambientes
criados por ele (FULLER, 2007).
Uma das consequências do processo de
domesticação de plantas cultivadas foi o surgimento
das ‘síndromes de domesticação’, ou seja, um conjunto
de caracteres que distinguem as plantas cultivadas dos
seus ancestrais selvagens (FULLER, 2007; BROWN et
al., 2009). O resultado foi o surgimento de inúmeras
novas variedades de diferentes espécies de plantas ou
mesmo o surgimento de novas espécies, resultando
num aumento da variabilidade genética, enquanto que
em outros casos o resultado foi uma diminuição da
variabilidade genética, como se observa, por exemplo,
no processo de seleção associado ao melhoramento
genético de plantas, ou em função da atuação da deriva
genética. Já em alguns casos, o resultado foi a
manutenção da variabilidade genética (LADIZINSKY,
1998).
Nesta r evisão, foram abor dados os
principais processos evolutivos que atuaram durante
a evolução e domesticação de plantas cultivadas.
Mutação
A mutação, um dos fatores evolutivos que
geram variabilidade genética, é definida como qualquer
alteração na sequência de nucleotídeos, bem como na
estrutura e número de cromossomos (FUTUYMA,
2006). Essas alter ações, que podem ocorr er
espontaneamente na célula ou pela ação de substâncias
mutagênicas, radiações ionizantes e ultravioletas,
causam substituições errôneas de bases nitrogenadas
e alter ações numéricas ou estruturais n os
cromossomos.
Mutações n os gen es r elacionados às
características de interesse agronômico podem ter
facilitado a domesticação de algumas espécies de
plantas cultivadas, principalmente em gramíneas, como
trigo, arroz e milho (LADIZINSKY, 1998). Essas culturas,
a partir de várias mutações gênicas, adquiriram maior
capacidade de retenção dos grãos na inflorescência
tornando-as mais dependentes da ação humana para
sua dispersão. Assim como as gramíneas, a uva (Vitis
vinifera) também sofreu diversas modificações
relacionadas à produção de frutos durante a sua
domesticação. Com o intuito de estudar a produção de
antocianina, principal responsável pela coloração dos
frutos da uva, THIS et al. (2007) analisaram a variação
do gene VvmybA em 200 acessos de uva e verificaram
que a inserção do elemento Gret1 na região promotora
permitiu o surgimento da coloração vermelha nos frutos
(Tabela 1).
Na geração F
3de um cruzamento entre
cebolas (Allium cepa) de coloração amarela, indivíduos
mutantes de coloração dourada foram identificados.
KIM et al. (2004) verificaram a presença de um códon
de terminação pré-maturo e adição de bases na região
que codifica a enzima Chalcone isomerase (CHI). Essa
mutação resultou n a in ativação do gen e CHI,
bloqueando a síntese de flavonóides e gerando um
acúmulo de derivados da Chalcone, culminando na
produção de uma cebola dourada. Em arroz, a deleção
do gene qSW5, responsável pelo controle do número
de células nas glumas e largura do grão, causou um
aumento na produção de sementes, favorecendo a
seleção artificial dessas variedades pelos primeiros
agricultores durante a domesticação da espécie
(SHOMURA et al., 2008). Na cultura do trigo (Triticum
aestivum), cromossomos não pareados, quebrados ou
perdidos, são anomalias cromossômicas bastante
observadas, o que dificulta a manutenção de linhagens
homozigotas. Essa instabilidade genética, geralmente
deletéria para outras culturas, na maioria das vezes não
provoca a morte das plantas de trigo devido ao seu
caráter hexaplóide (2n=6x=42) (FERNANDES et al.,
1991).
Como a mutação espontânea ocorre com
baixa frequência na natureza, a técnica de indução
utilizando agentes mutagênicos químicos e físicos tem
sido bastante empregada, visando obter mutações
promissoras tanto para o melhoramento genético, como
para obtenção de variantes importantes para o estudo
de função gênica. ISHIY et al. (2006) identificaram uma
nova variedade de arroz, SCS Andosan 114, progênie
mutante da cultivar ‘IR 840’ após o tratamento com
150Gy de radiação gama. A nova variedade mostrou-se
mais produtiva e com maior teor de amido no grão,
além da maior tolerância a doenças e toxidez por ferro.
KIKUCHI et al. (2009), avaliando o efeito de diferentes
fontes de irradiação em trigo, observaram que elas
geraram recombinações não deletérias, aumentando
artificialmente a taxa de recombinações nesta cultura.
ligados à pr odução de frutos em pêr a (Pyrus
communis) (PREDIERI & ZIMMERMAN, 2001) e
obtenção de mutantes para estudos fisiológicos e
função gênica em tomateiro (Solanum lycopersicum)
(PINO-NUNES et al., 2009), dentre outros.
Hibridação e fluxo gênico
Dentre todos os fatores evolutivos, a
hibridação natural, definida como acasalamento natural
entre indivíduos de duas ou mais populações, é um
dos eventos de maior importância evolutiva (ARNOLD,
2004). Diversos estudos têm demonstrado os seus
efeitos ao longo do processo de domesticação das
plantas cultivadas, envolvendo fluxo gênico entre
espécies selvagens e cultivadas (DUPUTIÉ et al., 2007),
envolvendo também a introgressão de culturas
geneticamente modificadas (DALTON, 2009) e a
especiação híbrida (MALLET, 2007).
Diversos trabalhos têm demonstrado a
ocorrência de fluxo gênico entre espécies cultivadas e
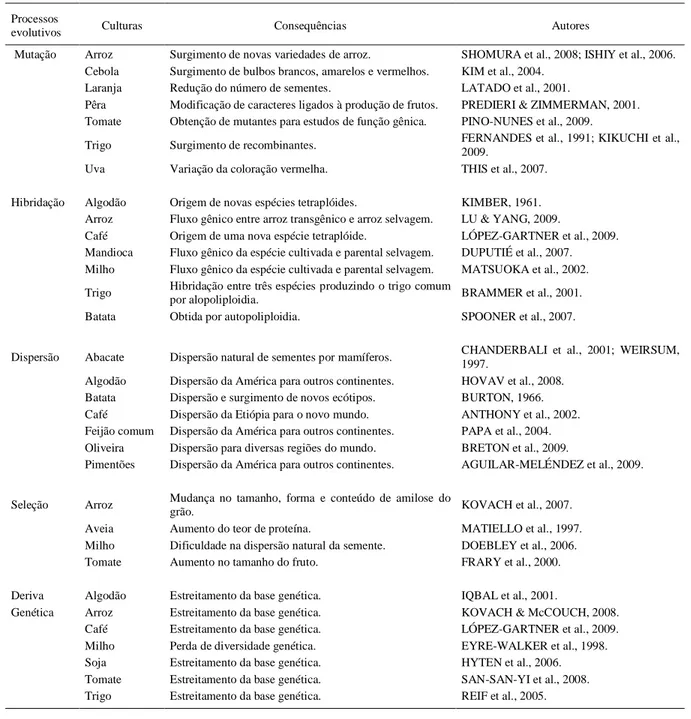
Tabela 1 - Consequências dos processos evolutivos na origem e domesticação de plantas cultivadas.
Processos
evolutivos Culturas Consequências Autores
Mutação Arroz Surgimento de novas variedades de arroz. SHOMURA et al., 2008; ISHIY et al., 2006.
Cebola Surgimento de bulbos brancos, amarelos e vermelhos. KIM et al., 2004.
Laranja Redução do número de sementes. LATADO et al., 2001.
Pêra Modificação de caracteres ligados à produção de frutos. PREDIERI & ZIMMERMAN, 2001.
Tomate Obtenção de mutantes para estudos de função gênica. PINO-NUNES et al., 2009.
Trigo Surgimento de recombinantes. FERNANDES et al., 1991; KIKUCHI et al.,
2009.
Uva Variação da coloração vermelha. THIS et al., 2007.
Hibridação Algodão Origem de novas espécies tetraplóides. KIMBER, 1961.
Arroz Fluxo gênico entre arroz transgênico e arroz selvagem. LU & YANG, 2009.
Café Origem de uma nova espécie tetraplóide. LÓPEZ-GARTNER et al., 2009.
Mandioca Fluxo gênico da espécie cultivada e parental selvagem. DUPUTIÉ et al., 2007.
Milho Fluxo gênico da espécie cultivada e parental selvagem. MATSUOKA et al., 2002.
Trigo Hibridação entre três espécies produzindo o trigo comum
por alopoliploidia. BRAMMER et al., 2001.
Batata Obtida por autopoliploidia. SPOONER et al., 2007.
Dispersão Abacate Dispersão natural de sementes por mamíferos. CHANDERBALI et al., 2001; WEIRSUM,
1997.
Algodão Dispersão da América para outros continentes. HOVAV et al., 2008.
Batata Dispersão e surgimento de novos ecótipos. BURTON, 1966.
Café Dispersão da Etiópia para o novo mundo. ANTHONY et al., 2002.
Feijão comum Dispersão da América para outros continentes. PAPA et al., 2004.
Oliveira Dispersão para diversas regiões do mundo. BRETON et al., 2009.
Pimentões Dispersão da América para outros continentes. AGUILAR-MELÉNDEZ et al., 2009.
Seleção Arroz Mudança no tamanho, forma e conteúdo de amilose do
grão. KOVACH et al., 2007.
Aveia Aumento do teor de proteína. MATIELLO et al., 1997.
Milho Dificuldade na dispersão natural da semente. DOEBLEY et al., 2006.
Tomate Aumento no tamanho do fruto. FRARY et al., 2000.
Deriva Algodão Estreitamento da base genética. IQBAL et al., 2001.
Genética Arroz Estreitamento da base genética. KOVACH & McCOUCH, 2008.
Café Estreitamento da base genética. LÓPEZ-GARTNER et al., 2009.
Milho Perda de diversidade genética. EYRE-WALKER et al., 1998.
Soja Estreitamento da base genética. HYTEN et al., 2006.
Tomate Estreitamento da base genética. SAN-SAN-YI et al., 2008.
seus parentais selvagens. MATSUOKA et al. (2002),
avaliando o fluxo gênico entre variedades de milho e
seu parental selvagem, o teosinto (Zea mays ssp.
mexicana), observaram que esse mecanismo afetou a
estrutur a gen ética das var iedades cultivadas,
possibilitando a sua sobrevivência em ambientes
extremos, como elevadas altitudes. Por outro lado, o
fluxo gênico da planta domesticada para seu parental
selvagem pode reduzir sua aptidão e comprometer sua
persistência em seu habitat natural, além de causar a
desestruturação genética do táxon e sua extinção, já
que o híbrido originado pode apresentar heterose e
competir com as formas selvagens (ELLSTRAND et
al., 1999). DUPUTIÉ et al. (2007) avaliaram a possível
ocorrência de uma zona híbrida entre a mandioca
cultivada (Manihot esculenta) e um parental selvagem
(Manihot ssp.) e observaram a ocorrência de híbridos
n atur ais que comparados aos seus par en tais
apresentaram heterose. Esse tipo de situação é
preocupante quando se considera a questão da
conser vação das espécies selvagens, já que a
ocorrência de fluxo gênico da planta cultivada para o
seu progenitor selvagem pode levar ao aparecimento
de plantas daninhas mais agressivas (MAGNUSSEN
& HAUSER, 2007) ou a introdução de genes da planta
cultivada no genoma da espécie selvagem, que poderia
levar à extinção das populações selvagens por mistura
completa (ELLSTRAND et al., 1999).
A preocupação com a conservação dos
recursos genéticos vegetais é ainda maior com o
surgimento das plantas transgênicas, já que a
ocorrência de fluxo gênico pode afetar a organização
da diversidade genética das variedades cultivadas e/
ou dos parentes silvestres. A verificação do fluxo
gênico entre espécies que apresentam a forma
transgênica é importante para inferir a possibilidade de
trocas de alelos que representam uma ameaça para a
preservação dos tipos selvagens (GEPTS & PAPA,
2003). Em uma revisão sobre o tema, LU & YANG (2009)
avaliaram vários estudos de fluxo gênico entre espécies
de arroz selvagem e transgênico e concluíram que o
escape destes para populações selvagens ou daninhas
pode representar consequências ecológicas em
potencial.
A especiação híbrida é aquela em que a
hibridação teve um importante papel na formação de
uma nova espécie (MALLET, 2007). Em geral, uma nova
espécie é formada quando duas delas se cruzam e o
genoma do híbrido é duplicado, formando um
alopoliplóide. Esse processo pode se repetir de modo
a formar espécies com mais de dois genomas diferentes.
Diversos exemplos de alopoliplóides são descritos na
literatura, entre eles o caso do trigo (Triticum aestivum),
formado por três genomas distintos (AABBDD),
originário do cruzamento entre T. monococcum (AA),
T. searssi (BB), sendo que o híbrido F
1 teve o seunúmero de cromossomos duplicados, formando o T.
turgidum (AABB) que se hibridizou com o T. tauschii
(DD) (BRAMMER et al., 2001). Outras espécies de
plantas cultivadas que sofreram o mesmo mecanismo é
o algodão tetraplóide (Gossypium hirsutum e G.
barbadense), formado a partir do cruzamento de uma
espécie diplóide (genoma A) originária da Ásia com
outra espécie diplóide (genoma D) originária do Novo
Mundo (KIMBER, 1961); e também o café tetraplóide
(Coffea arabica), originário do cruzamento entre as
espécies Coffea eugenioides e C. canephora
(LÓPEZ-GARTNER et al., 2009), entre outros (Tabela 1).
Contudo, além da alopoliploidia n a
especiação híbrida, os autopoliplóides também tiveram
um papel muito importante na evolução das plantas
cultivadas, pois o efeito “giga” proporcionado pelo
aumento no número de cópias dos cromossomos em
uma mesma célula ajudou o homem a selecionar plantas
com raízes, tubérculos, caules, folhas, frutos e sementes
maiores. No entanto, este processo tem sido associado
a uma redução na fertilidade devido a problemas no
pareamento da meiose, o que explica o fato da
autopoliploidia espontânea estar mais associada às
plantas de reprodução vegetativa, ou àquelas em que
as partes vegetativas são utilizadas (LADIZINSKY,
1998). Na história da domesticação das plantas, alguns
autopoliplóides merecem destaque, como é o caso da
batata (Solanum tuberosum), a qual pode ser diplóide
(2n=2x=24), triplóide (2n=3x=36), tetraplóide (2n=4x=48)
e até pentaplóide (2n=5x=60) (SPOONER et al., 2007).
Podemos citar também os poliplóides induzidos, como
é o caso do autotetraplóide artificial de melancia
(Citrullus vulgaris) com 2n=4x=44 cromossomos
(JASKANI et al., 2004), o qual tem sido utilizado em
cruzamentos com linhagens diplóides (2n=2x=22) para
obtenção de híbridos triplóides (2n=3x=33) sem
sementes.
Dispersão, migração e colonização
con dições edafoclimáticas diferentes daquelas
apresentadas em seus centros de origem, diversidade
ou domesticação. A cultura da batata (Solanum
tuberosum), levada da América do Sul à Europa, sofreu
restrições ao florescimento devido ao fotoperíodo do
Velho Mundo, com dias mais longos. No entanto, com
o conhecimento da importância desse fator, a batata
pôde ser explorada, permitindo a ocorrência da troca
de sementes entre os agricultores e o surgimento de
uma nova diversidade de ecótipos, adaptados aos dias
mais longos (BURTON, 1966).
A história da ocupação e deslocamento
humano pelos continentes também foram decisivos para
o surgimento das espécies domesticadas. O abacate
(Persia americana), por exemplo, que teve seu
ancestral disperso durante a era glacial no Pleistoceno,
chegando a atingir a Mesoamérica através da
movimentação de grandes mamíferos e dispersão
natural das sementes, foi domesticado após a chegada
do homem à América, por volta de 8000 a 7000 a.C.
(CHANDERBALI et al., 2001). Posteriormente, o
abacate teria sido levado ao sul do continente
acompanhando as rotas de colonização humana
pré-colombiana (WEIRSUM, 1997). Em eventos mais
recentes de dispersão, a cultura foi levada a outros
continentes, predominantemente pelos colonizadores
europeus, assim como ocorreu nos séculos 16 e 17
com diversas culturas também originadas na América,
tais como o feijão comum (Phaseolus vulgaris) (PAPA
et al., 2004), pápricas e pimentões (Capsicum annuum)
(AGUILAR-MELÉNDEZ et al., 2009), amendoim
(Arachis hypogaea) (WILLIAM, 2004) e algodão
(Gossypium hirsutum) (HOVAV et al., 2008) (Tabela 1).
Outro exemplo que podemos citar é a cultura do abacaxi
(Ananas comosus), originário da América tropical e
subtropical e, muito provavelmente, do Brasil (MEDINA
et al., 1978; CRESTANI et al., 2010), sendo disperso
para vários países americanos pelo intercâmbio entre
tribos, e tendo sido levado para a Europa, Ásia e África
após o descobrimento da América, disseminando-se
rapidamente pelos países desses continentes (CTENAS
& QUAST, 2000).
Dados moleculares têm sido eficientes
ferramentas para o estudo da origem e domesticação
de espécies cultivadas (BROWN et al., 2009).
Marcadores microssatélites foram fundamentais para
definirem a ocor rência de locais distin tos de
domesticação da oliveira (Olea europaea subsp.
europaea), resultantes de eventos de fluxo gênico da
espécie selvagem associado à migração humana. A
espécie selvagem de oliva, domesticada na região do
Oriente Médio, Norte da África e oeste da Europa, foi
levada pelo homem para diversas regiões do mundo
com clima Mediterrâneo (África, Austrália, Ásia e
América) (BRETON et al., 2009). Na cultura do café
(Coffea arabica), foi possível entender os eventos de
seleção negativa e deriva genética, que resultaram no
estreitamento da base genética da espécie, constituindo
apen as dois conjun tos gên icos bem defin idos
originários de duas subpopulações selvagens do
sudoeste da Etiópia (ANTHONY et al., 2002).
Eventos recentes de dispersão podem ser
relacionados com a troca de material vegetal entre áreas
agriculturáveis. Na região sudeste do Brasil, este tipo
de dispersão, característico da agricultura tradicional,
tem ampliado a base genética de culturas manejadas
em sistemas agrícolas (PERONI et al., 2007). Estas áreas
são caracterizadas por realizar constantes trocas entre
os locais de cultivo, estimulando a dispersão de
indivíduos e, consequentemente, possibilitando maior
fluxo gênico entre as populações mantidas neste
sistema agrícola. Além disso, os agricultores tendem a
manter alta variabilidade inter e intra-específica
(BRESSAN et al., 2005; VEASEY et al., 2008). Esses
fatores têm con tr ibuído para a manutenção e
amplificação da diversidade genética de muitas
espécies cultivadas.
Seleção
A seleção é a ação natural ou artificial
exercida em uma determinada população capaz de alterar
suas frequências alélicas, pelo fato de apenas alguns
indivíduos contribuírem para a formação das gerações
seguintes (FALCONER, 1987). Na seleção natural, as
populações são direcionadas no sentido de
tornarem-se mais adaptadas ao ambiente no qual estão intornarem-seridas,
decorrente do sucesso reprodutivo diferencial dos
indivíduos (HANCOCK, 2005). Na artificial, a seleção é
realizada de forma intencional ou inconsciente, visando
à melhoria das características vegetais para os interesses
e necessidades humanas. Este processo, conhecido
como domesticação, está diretamente relacionado à
evolução das plantas cultivadas (HARLAN, 1992).
espécie ancestral, apresenta maior dificuldade na
dispersão natural da semente, pois os grãos estão
aderidos ao sabugo e são envolvidos pela palha
(DOEBLEY et al., 2006). Assim como no milho, o arroz,
além das alterações estruturais e reprodutivas ocorridas
ao longo da domesticação, tem sofrido pressão de
seleção pelo homem para tornar os grãos mais atraentes,
com relação ao tamanho, forma, cor, fragrância e
conteúdo de amilose do grão (KOVACH et al., 2007).
Nesse mesmo contexto, o aumento do teor de proteína
bruta no grão de aveia para a alimentação humana tem
sofrido constante ação do homem, visto que a maioria
dos materiais selvagens possui baixo teor de proteína
(MATIELLO et al., 1997). Outro exemplo seria o do
tomateiro (Solanum lycopersicum), cujos frutos
sofreram um incrível aumento no tamanho ao longo de
sua domesticação, quando comparado com seu
ancestr al mais pr óximo S. lycopersicum var
cerasiforme. Esse processo no tomateiro, envolveu
principalmente a mutação e seleção do QTL (loco de
característica quantitativa) fw2.2, o qual é um loco
chave para a evolução do tamanho do fruto da cultura
(FRARY et al., 2000) (Tabela 1).
Devido às características envolvidas no
processo de domesticação serem principalmente
quantitativas, o uso de técn icas molecular es,
sobretudo o mapeamento de locos de caracteres
quantitativos, tem auxiliado nos estudo da evolução
das espécies vegetais, tanto na ação destes genes
envolvidos na domesticação quanto nos efeitos da
seleção neste processo (YAMASAKI et al., 2007). A
análise comparativa do polimorfismo de nucleotídeos
dos gen es r elacionados à domesticação tem
possibilitado determinar em que medida a seleção está
atuando em todo o genoma. Esta análise, denominada
selective sweep, permite observar os nucleotídeos que
foram eliminados em função da elevada pressão de
seleção nos materiais selvagens. No milho, o gene
teosinte branched1 (Tb
1
) foi identificado num QTL que
atua no controle da dormência dos meristemas apicais
e axilares, sendo ativo em seu ancestral selvagem, o
teosinto (Zea mays ssp. parviglumis) (DOEBLEY,
2004). O gene Q, envolvido na domesticação do trigo,
afeta um conjunto de características, incluindo a forma
alongada do trigo selvagem ou compacta como nas
espécies cultivadas, bem como a estr utura de
inflorescência e época de floração (SIMONS et al.,
2006). O shattering 4 (Sh
4
) é um QTL que controla a
liberação de sementes da planta no arroz selvagem ou
permite a sua maior fixação, como no arroz cultivado
(KOVACH et al., 2007). A comparação genômica das
plantas cultivadas com os seus respectivos parentais
selvagens permite compreender que a evolução do
genoma está relacionada à pressão de seleção em
deter min adas r egiões do mater ial gen ético,
r espon sáveis pela expr essão dos caracter es
relacionados à domesticação (BURKE et al., 2007).
Deriva genética
A der iva gen ética, defin ida como a
imprevisibilidade de variação nas frequências alélicas
em uma população de tamanho reduzido, pode ocorrer
tanto em populações naturais quanto em espécies
cultivadas (FUTUYMA, 2006). Em relação às plantas
cultivadas, a deriva genética pode ser representada
praticamente de duas formas: efeito do fundador,
quando a domesticação ocorre fora do seu centro de
origem, geralmente devido ao fato de uma amostra de
poucos indivíduos ser levada para colonizar outra
região e; gargalo de garrafa (bottleneck), quando o
tamanho de uma população é reduzido drasticamente
devido ao processo de seleção feito pelo homem, sendo
que sua recomposição é feita a partir de poucos
indivíduos e uma quantidade restrita de alelos
(HANCOCK, 2004).
Em geral, a magnitude dos efeitos da deriva
é mais forte quando a domesticação de uma planta
ocorre fora do seu centro de origem, pois a troca de
alelos com espécies selvagens torna-se inexistente.
Esse efeito é mais pronunciado ainda em espécies
autógamas, pois a endogamia, ao eliminar alelos
deletérios por seleção natural ou artificial, acaba também
eliminando alelos não deletérios que estão ligados a
estes. Este é o exemplo do tomateiro (Solanum
lycopersicum), planta autógama, domesticada no
México, fora de seu centro de origem na América do
Sul, no qual a principal consequência foi o estreitamento
da base genética da cultura (MILLER & TANKSLEY,
1990), tornando-a vulnerável ao ataque de pragas e
doenças, além da fragilidade a ambientes estressantes.
Processo semelhante observa-se na soja (Glycine max),
já que poucos acessos foram trazidos da Ásia e
introduzidos na América do Norte, no período de 1947
a 1988 (HYTEN et al., 2006). Já no caso do milho (Zea
mays), tal efeito foi menos pronunciado, uma vez que
esta é uma espécie alógama, a qual foi domesticada no
seu próprio centro de origem. Isso fez com que o milho
preservasse perto de 75% da sua diversidade genética
original, quando comparado com seus parentais
selvagens, Zea parviglumis e Zea luxurians
(EYRE-WALKER et al., 1998).
(SAN-SAN-YI et al., 2008), grão-de-bico (Cicer
arietinum) (ABBO et al., 2003), entre outras (Tabela 1).
Tal processo também foi importante na domesticação
de espécies arbóreas como a teca (Tectona grandis)
(VARGHESE et al., 2006) e de plantas daninhas que
coevoluiram com as plantas cultivadas, como é o caso
de Lythrum salicaria, a qual foi introduzida nos EUA
a partir da Europa (ECKERT et al., 1996). Estimativas da
intensidade dessa perda de diversidade genética
en con trada n os an cestr ais selvagen s variam
consideravelmente, desde 80% no milho (WRIGHT &
GAUT, 2005) para 40-50% no girassol (LIU & BURKE,
2006) e 10-20% no arroz (ZHU et al., 2007).
Em contraste, é possível, em menor escala,
fazer o caminho contrário ao processo de deriva,
atr avés da busca de novos alelos em espécies
selvagens e variedades crioulas. Vários programas de
melhoramento genético buscam novos alelos em
espécies selvagen s, pr in cipalmen te aqueles
relacionados a tolerâncias a diversas pragas e doenças.
Esse é o caso de espécies como o trigo (Triticum
aestivum) (REIF et al., 2005), o arroz (KOVACH &
McCOUCH, 2008) e tomateiro (PINO-NUNES et al.,
2009), às quais vem sendo reincorporados vários novos
alelos a par tir de cr uzamen tos com espécies
relacionadas.
CONCLUSÃO
Conclui-se com esta revisão que a origem e
a domesticação das plantas cultivadas estiveram
diretamente relacionadas, ao longo de suas histórias
evolutivas, com os processos evolutivos abordados
(mutação, hibridação, migração, seleção e deriva
genética), que têm atuado no sentido de manter,
aumentar ou mesmo diminuir a diversidade genética
das espécies cultivadas. São vários os exemplos (Tabela
1) de como esses processos influenciaram no
aparecimento de novas espécies de plantas, assim como
de novas variedades dentro de espécies domesticadas.
É bom lembrar que a divisão desses processos foi feita
de forma a tornar sua compreensão mais clara, uma vez
que todos eles ocorrem de forma simultânea, diferindo
somente na magnitude da importância de cada
fenômeno para a domesticação das diferentes espécies
cultivadas.
REFERÊNCIAS
ABBO, S. et al. Evolution of cultivated chickpea: four bottlenecks limit diversity and constrain crop adaptation. Functional Plant
Biology, v.30, n.10, p.1081-1087, 2003. Disponível em: <http:/
/www.publish.csiro.au/?act=view_file&file_id=FP03084.pdf>. Acesso em: 19 abr. 2011. doi: 10.1071/FP03084.
AGUILAR-MELÉNDEZ, A. et al. Genetic diversity and structure in semiwild and domesticated chiles (Capsicum annum; Solanaceae) from Mexico. American Journal of Botany, v.96, p.1190-1202, 2009. Disponível em: <http://www.amjbot.org/cgi/reprint/96/6/ 1190?maxtoshow=&hits=10&RESULTFORMAT=&author1=Aguilar-melendez&andorexacttitle=and&a ndorexacttitleabs=and&andore c t f u l l t e x t = a n d & s e a r c h i d = 1 & F I R S T I N D E X=0&sortspec=relevance&resourcetype=HWCIT>. Acesso em: 19 abr. 2011. doi: 10.3732/ajb.0800155.
ANTHONY, F. et al. The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers. Theoretical
and Applied Genetics, v.104, p.894-900, 2002. Disponível
em: <http://www.springerlink.com/content/4r9txcxh052l1gkk/ fulltext.pdf>. Acesso em: 19 abr. 2011. doi: 10.1007/s00122-001-0798-8.
ARNOLD, M.L. Natural hybridization and the evolution of domesticated pest and disease organisms. Molecular Ecology, v.13, p.997-1007, 2004. Disponível em: <http:// o n l i n e l i b r a r y . w i l e y . c o m / d o i / 1 0 . 1 1 1 1 / j . 1 3 6 5 -294X.2004.02145.x/pdf>. Acesso em: 19 abr. 2011. doi: 10.1111/j.1365-294X.2004.02145.x.
BELLWOOD, P. First farmers: the origins of agricultural societies. Blackwell, Malden, Mass, 2005. 360p.
BRAMMER, S.P. et al. A potencialidade de Agropyron,
espécie afim ao trigo cultivado, como fonte de introgressão de genes agronomicamente importantes.
Passo Fundo: Embrapa Trigo, 2001. 8p. (Embrapa Trigo Documentos online: 8)
BRESSAN, E.A. et al. Collecting yam (Dioscorea spp.) and sweet potato (Ipomoea batatas) germplasm in traditional agriculture small-holdings in the Vale do Ribeira, São Paulo, Brazil. Plant Genetic
Resources Newsletter, v.144, p.8-13, 2005. Disponível em:
<http://www2.bioversityinternational.org/publications/pgrnewsletter/ article.asp?lang=en&id_article=2&id_issue=144. Acesso em: 19 abr. 2011.
BRETON, C. et al. The origins of the domestication of the olive tree. Comptes Rendus Biologies, v.332, p.1059-1064, 2009. Disponível em: <http://www.sciencedirect.com/ s c ie n c e ? _ o b = M I mg & _ i m a g e k e y = B 6 X 1 F 4 X 5 B P 1 K 1 -7 &_ cdi= -7 2 4 1 &_ u ser = 5 6 -7 4 9 3 1 &_ pii= S1 6 3 1 0 6 9 1 0 9 0 0 1 8 9 9 &_ or igin= sea r ch&_ zone= r slt_ list_ item&_ cover D a te = 12 %2 F3 1 %2 F2 00 9 &_ sk =9 9 66 7 9 98 7 &wchp= dGLzVzb-zSkWA&md5=eba93d1e6ec7db84b13537b54527c910&ie=/ sdarticle.pdf>. Acesso em: 19 abr. 2011. doi:10.1016/ j.crvi.2009.08.001.
BURKE, J.M. et al. Crop evolution: from genetics to genomics.
Current Opinion in Genetics & Development, v.17,
p.525-532, 2007. Disponível em: <http://www.sciencedirect.com/ s c i e n c e ? _ o b = A r t i c l e U R L & _ u d i = B 6 V S 0 4 P Y J D Y K -4 & _ u s e r = 1 0 & _ c o v e r D a t e = 1 2 % 2 F 3 1 % 2 F 2 0 0 7 & _ r d o c = 1 3 & _ f m t = h i g h & _ o r i g = b r o w s e & _ o r i g i n = b r o w s e & _ z o n e = r s l t _ l i s t _ i t e m & _ s r c h = d o c -i n f o ( % 2 3 t o c % 2 3 6 2 4 8 % 2 3 2 0 0 7 % 2 3 9 9 9 8 2 9 9 9 3 % 2 3 6 7 5 6 8 6 % 2 3 F L A % 2 3 d i s p l a y % 2 3 V o l u m e)&_cdi=6248&_sort=d&_docanchor=&_ct=17&_acct=C00005022 1&_version=1&_urlVersion=0&_userid=10&md5=92196ad8f7c 2bbe51648d0d0ea366fb3&searchtype=a>. Acesso em: 20 abr. 2011. doi: 10.1016/j.gde.2007.09.003.
BURTON, W.G. The potato. Wageningen: Veenman and Zonen, 1966. 153p.
CHANDERBALI, A.S. et al. Phylogeny and historical biogeography of Lauraceae: evidence from chloroplast and nuclear genomes. Annals of the Missouri Botanical
Garden, v.88, p.104-134, 2001. Disponível em: <http://
www.biodiversitylibrary.org/item /86343#page/106/mode/1up>. Acesso em: 20 abr. 2011.
CRESTANI, M. et al. Das Américas para o Mundo - origem, domesticação e dispersão do abacaxizeiro. Ciência Rural, v.40, p.1473-1483, 2010. Disponível em: <http://www.scielo.br/ pdf/cr/v40n6/a620cr2584.pdf>. Acesso em: 20 abr. 2011. doi:
10.1590/S0103-84782010000600040.
CTENAS, M.L.B.; QUAST, D. Abacaxi. In: CTENAS, A.C. et al. (Eds.). Frutas das terras brasileiras. São Paulo: C2, 2000. p.41-45.
DALTON, R. Mexico´s transgenic maize under fire. Experimental planting scheme has insufficient controls to prevent gene flow to native crops, critics say. Nature, v.462, p.404, 2009. Disponível em:<http://www.nature.com/news/ 2009/091125/pdf/462404a.pdf>. Acesso em: 20 abr. 2011. doi:10.1038/462404b.
DIAMOND, J.M. Evolution, consequences and future of plant and animal domestication. Nature, v.418, p.700-707, 2002. Disponível em: <http://www.nature.com/nature/ journal/v418/ n6898/pdf/nature01019.pdf. Acesso em: 20 abr. 2011. doi:10.1038/nature01019.
DIAMOND, J.M.; BELLWOOD, P. Farmers and their languages: the first expansions. Science, v.300, p.597-603, 2003. Disponível em: <http://www.sciencemag.org/content/ 300/5619/ 597.full.pdf>. Acesso em: 20 abr. 2011. doi:10.1126/ science.1078208.
DOEBLEY, J.F. The genetics of maize evolution. Annual
Review of Genetics, v.38, p.37-59, 2004. Disponível em:
< h t t p : / / w w w. a n n u a l r e v i e w s . o r g / d o i / p d f / 1 0 . 1 1 4 6 / annurev.genet.38.072902.092425>. Acesso em: 20 abr. 2011. doi: 10.1146/annurev.genet.38.072902.092425.
DOEBLEY, J.F. et al. The molecular genetics of crop domestication. Cell, v.127, p.1309-1321, 2006. Disponível em: <http://www.cell.com/abstract/S0092-8674(06)01592-3>. Acesso em: 20 abr. 2011. doi:10.1016/j.cell.2006.12.006.
DUPUTIÉ, A. et al. Natural hybridization between a clonally propagated crop, cassava (Manihot escultenta Crantz).
Molecular Ecology, v.16, p.3025-3038, 2007. Disponível
em: <http://onlinelibrary.wiley.com/doi/10.1111/j.1365-294X.2007.03340.x/pdf>. Acesso em: 19 abr. 2011. doi: 10.1111/j.1365-294X.2007.03340.x.
ECKERT, C.G. et al. Genetic drift and founder effect in native versus introduced populations of an invading plant, Lythrum
salicaria (Lythraceae). Evolution, v.50, p.1512-1519, 1996.
Disponível em: <http://www.jstor.org/stable/2410888>. Acesso em: 20 abr. 2011.
ELLSTRAND, N.C. et al. Gene flow and introgression from domesticated plants into their wild relatives. Annual Review
of Ecology, Evolution and Systematics, v.30, p.539-563,
1999. Disponível em: <http://www.annualreviews.org/doi/pdf/ 10.1146/annurev.ecolsys. 30.1.539>. Acesso em: 20 abr. 2011. doi: 10.1146/annurev.ecolsys.30.1.539.
EYRE-WALKER, A. et al. Investigating the bottleneck leading to the domestication of maize. Proceedings of the National
Academy of Sciences, v.95, p.4441-4446, 1998. Disponível
em: <http://www.pnas.org/content/95/8/4441.full.pdf+html>. Acesso em 20 abr. 2011.
FALCONER, D.S. Introdução à genética quantitativa. Viçosa: UFV, 1987. 279p.
FERNANDES, M.I.B.M. et al. Fatores responsáveis pela desuniformidade varietal no trigo Triticum aestivum (L.) Thell e o papel da instabilidade cromossômica. Pesquisa
Agropecuária Brasileira, v.26, p.353-369, 1991. Disponível
em: <http://webnotes.sct.embrapa.br/pdf/pab1991/marco/ pab10_mar_91.pdf>. Acesso em: 20 abr. 2011.
FRARY, A. et al. Fw2.2: a quantitative trait locus key to the evolution of tomato fruit size. Science, v.289, p.85-88, 2000. Disponível em: <http://www.sciencemag.org/content/289/5476/ 85>. Acesso em: 20 abr. 2011. doi: 10.1126/ science.289.5476.85.
FULLER, D.Q. Contrasting patterns in crop domestication and domestication rates: recent archaeobotanical insights from the Old World. Annals of Botany, v.100, p.903-924, 2007. Disponível em: <http://aob.oxfordjournals.org/content/100/5/ 903.full.pdf+html>. Acesso em: 20 abr. 2011. doi: 10.1093/ aob/msm048.
FUTUYMA, D.J. Evolutionary biology. Sunderland: Sinauer Associates, 2006. 763p.
GEPTS, P.; PAPA, R. Possible effects of (trans) gene flow from crops on the genetic diversity from landraces and wild relatives. Environmental Biosafety Research, v.2, p.89-103, 2003. Disponível em: <http://www.ncbi.nlm.nih.gov/ pubmed/15612275>. Acesso em: 20 abr. 2011.
HAMARICK, J.L.; NASON, J.D. Consequences of dispersal in plants. In: RHODES, O.E. et al. Population dynamic in
ecological space and time. Chicago: University of Chicago,
1996. p.203-236.
HANCOCK, J.F. Plant evolution and the origin of crop
HANCOCK, J.F. Contributions of domesticated plant studies to our understanding of plant evolution. Annals of Botany, v.96, p.953-963, 2005. Disponível em: <http:// a ob.oxfor djou r nals.org/content/9 6 /6 /9 5 3.fu ll.pdf+ html> . Acesso em: 20 abr. 2011. doi: 10.1093/aob/mci259. HARLAN, J.R. Crops and man. Madison: American Society of Agronomy, 1992. 284p.
HOVAV, R. et al. Parallel domestication, convergent evolution and duplicated gene recruitment in allopolyploid cotton.
Genetics, v.179, p.1725-1733, 2008. Disponível em: <http:/
/www.genetics.org/cgi/reprint/179/3/1725>. Acesso em: 20 abr. 2011. doi: 10.1534/genetics.108.089656.
HYTEN, D.L. et al. Impacts of genetic bottlenecks on soybean genome diversity. Proceedings of the National Academy
of Sciences, v.103, p.16666-16671, 2006 . Disponível em:
<http://www.pnas.org/content/103/45/16666.full.pdf+html>. Acesso em: 20 abr. 2011. doi: 10.1073/pnas.0604379103. IQBAL, M.J. et al. A genetic bottleneck in the ‘evolution under domestication’ of upland cotton Gossypium hirsutum L. examined using DNA fingerprinting. Theoretical and Applied
Genetics, v.103, p.547-554, 2001. Disponível em: <http://
w w w. s p r i n g e r l i n k . c o m / c o n t e n t / 9 b g b 2 x y 1 4 q e 2 7 0 w r / fulltext.pdf>. Acesso em: 20 abr. 2011. doi: 10.1007/ PL0000 2908.
ISHIY, T. et al. Rice mutant cultivar SCS114 Andosan. Plant
Mutation Report, v.1, p.25, 2006. Disponível em: <http://
www-pub.iaea.org/MTCD/publications/PDF/Newsletters/PMR-01-02.pdf>. Acesso em: 20 abr. 2011.
JASKANI, M.J. et al. Induction and characterization of tetraploid watermelon. Journal of the Korean Society
Horticultural Science, v.45, p.60-65, 2004.
KIKUCHI, S. et al. Effects of heavy-ion beams on chromosomes of common wheat, Triticum aestivum. Mutation Research, v.669, p.63-66, 2009. Disponível em: <http://www.sciencedirect.com/ s c ie n c e? _ o b = M I m g& _ i ma g e k ey = B 6 T 2 C 4 W8 V W 8 R 1 3 & _ c d i = 4 9 1 5 & _ u s e r = 5 6 7 4 9 3 1 & _ p i i = S 0 0 2 7 5 1 0 7 0 9 0 01 61 4 &_ origin=ga tewa y&_ cover D ate=1 0%2 F0 2 %2 F2 0 09&_sk=993309998&view=c&wchp=dGLbVtbzSkWA&md5 =3c85a24fe0547c223f9a8ca213270906&ie=/sdarticle.pdf>. Acesso em: 20 abr. 2011. doi: 10.1016/j.mrfmmm.2009.05.001. KIM, S. et al. Gold color in onions (Allium cepa): a natural mutation of the chalcone isomerase gene resulting in a premature stop codon. Molecular Genetics and Genomics, v.272, p.411-419, 2004. Disponível em: <http://www.springerlink.com/ content/yfyj 5844d0m02mxw/fulltext.pdf>. Acesso em: 20 abr. 2011. doi: 10.1007/s00438-004-1076-7.
KIMBER, G. Basis of the diploid-like meiotic behaviour of polyploid cotton. Nature, v.191, p.98-100, 1961. Disponível em: <http://www.nature.com/nature/journal/v191/ n4783/abs/ 191098a0.html>. Acesso em: 20 abr. 2011. doi: 10.1038/ 191098a0.
KOVACH, M.J.; McCOUCH, S.R. Leveraging natural diversity: back through the bottleneck. Current Opinion in Plant
Biology, v.11, p.193-200, 2008. Disponível em: <http://
w w w . s c i e n c e d i r e c t . c o m / s c i e n c e ? _ o b = M I m g & _ i m a g e k e y = B 6 V S 4 4 R Y 9 1 X D 1
9 & _ c d i = 6 2 5 2 & _ u s e r = 5 6 7 4 9 3 1 & _ p i i = S 1 3 6 9 5 2 6 6 0 8 000186&_origin=gateway&_coverDate=04%2F30%2F2008&_sk=99 9 8 8 9 9 9 7 & v i e w = c & w c h p = d G L b V z b z S k W b & m d 5 = 8 2 7 a 8 f 9 7 6 5 0 6 2 c 6 a d e c f 4 e d 2 7 0 e d 2 9 2 5 & i e = / sdarticle.pdf>. Acesso em: 20 abr. 2011. doi: 10.1016/ j.pbi.2007.12.006.
KOVACH, M.J. et al. New insights into the history of rice domestication. Trends in Genetics, v.23, p.578-587, 2007. Disponível em: <http://www.sciencedirect.com/science? _ o b = M I m g & _ i m a g e k e y = B 6 T C Y 4 R 0 5 H Y K 1 9 & _ c d i = 5 1 8 3 & _ u s e r = 5 6 7 4 9 3 1 & _ p i i = S 0 1 6 8 9 525 0700 3046&_origin=gatewa y&_coverD ate=1 1%2F30% 2 F20 0 7 &_sk = 99 9 7 6 99 8 8 &view= c&wchp= dG LbVtbzSk z k & m d 5 = f d 5 d 0 d a 9 a 8 7 8 6 e 4 d c 6 4 3 9 1 e 6 b 6 0 f 0 2 8 c & i e = / sdarticle.pdf>. Acesso em: 20 abr. 2011. doi: 10.1016/ j.tig.2007.08.012.
LADIZINSKY, G. Plant evolution under domestication. London: Kluwer Academic, 1998. 254p.
LATADO, R.R. et al. Mutantes de laranja-‘pêra’ com número reduzido de sementes, obtidos através de mutações induzidas.
Revista Brasileira de Fruticultura, v.23, p.339-344, 2001.
Disponível em: <http://www.scielo.br/pdf/rbf/v23n2/7978.pdf>. Acesso em: 20 abr. 2011. doi:
10.1590/S0100-29452001000200028.
LIU, A.; BURKE, J.M. Patterns of nucleotide diversity in wild and cultivated sunflowers. Genetics, v.173, p.321-330, 2006. Disponível em: <http://www.genetics.org/cgi/reprint /173/1/ 3 2 1 ? m a x t o s h o w = & h i t s = 1 0 & R E S U L T F O R M A T = 1 & a n d o r e x a c t t i t l e = p h r a s e & a n d o r e x a c t t i t l e a b s = p h r a s e & a n d o r e x a c t f u l l t e x t = p h r a s e & s e a r c h i d = 1 & F I R S T I N D E X = 0 & s o r t s p e c = d a t e & v olume=173&firstpage=321&resourcetype=HWCIT>. Acesso em: 20 abr. 2011. doi: 10.1534/genetics.105.051110. LÓPEZ-GARTNER, G. et al. Analysis of genetic structure in a sample of coffee (Coffea arabica L.) using fluorescent SSR markers. Tree Genetics and Genomes, v.5, p.435-446, 2009. Disponível em: <http://www.springerlink.com/content/ j0q51761753810um/fulltext. html>. Acesso em: 25 abr. 2011. doi: 10.1007/s11295-008-0197-2.
LU, B.; YANG, C. Gene flow from genetically modified rice to its wild relatives: assessing potential ecological consequences.
Biotechnology Advances, v.27, p.1083-1091, 2009. Disponível em: <http://www.sciencedirect.com/ s c i e n c e ? _ o b = A r t i c l e U R L & _ u d i = B 6 T 4 X - 4 W B C 1 P T-7&_user=5674931&_coverDate=12%2F31%2F2009&_rdoc= 1&_fmt=high&_orig=gateway&_origin=gateway&_sort=d&_d ocanchor=&view=c&_searchStrId=1729954171&_rerunOrigin=google& _acct=C000049650&_version=1&_urlVersion=0&_userid=56749 31&md5=e7487794b63efd0d30dc6bad45810166&searchtype=a>. Acesso em: 25 abr. 2011. doi: 10.1016/j.biotechadv.2009.05.018. MEDINA, J.C. A cultura do abacaxi. In: MEDINA, J.C. et al.
Frutas tropicais 2. São Paulo: Canton, 1978. p.6-68.
MALLET, J. Hybrid speciation. Nature, v.446, p.279-283, 2007. Disponível em: <http://www.nature.com/nature/journal/ v446/n7133/pdf/nature05706.pdf>. Acesso em: 20 abr. 2011. doi:10.1038/nature05706.
MATIELLO, R.R. et al. Variabilidade genética para teor de proteína bruta em grãos de aveia. Ciência Rural, v.27, p.183-187, 1997. Disponível em: <http://www.scielo.br/pdf/cr/v27n2/ a02v27n2.pdf>. Acesso em: 20 abr. 2011. doi: 10.1590/S0103-84781997000200002.
MATSUOKA, Y. et al. A single domestication for maize shown by multilocus microsatellite genotyping. Proceedings of the
National Academy of Sciences, v.99, p.6080-6084, 2002.
Disponível em: <http://www.pnas.org/content/99/9/ 6080.full.pdf+html>. Acesso em: 20 abr. 2011. doi:10.1073/ pnas.052125199.
MILLER, J.C.; TANKSLEY, S.D. RFLP analysis of phylogenetic relationships and genetic variation in the genus Lycopersicon.
Theoretical and Applied Genetics, v.80, p.437-448, 1990.
Disponível em: <http://www.springerlink.com/content/ w290x81038054800/>. Acesso em: 25 abr. 2011. doi: 10.1007/ BF00226743.
PAPA, R. et al. The evolution of genetic diversity in Phaseolus
vulgaris L. In: MOTLEY, T.J. Darwin’s harvest: new
approaches to the origins, evolution, and conservation of crop. New York: Columbia University, 2004. p.121-142.
PERONI, N. et al. Molecular differentiation, diversity, and folk classification of “sweet” and “bitter” cassava (Manihot
esculenta) in Caiçara and Caboclo management systems (Brazil).
Genetic Resources and Crop Evolution, v.54, p.1333-1349,
2007. Disponível em: <http://www.springerlink.com/content/ q3827ug3p234m67q/fulltext.pdf.> Acesso em: 20 abr. 2011. doi: 10.1007/s10722-006-9116-2.
PINO-NUNES, L.E. et al. Induced mutagenesis and natural genetic variation in tomato ‘Micro-tom’. Acta Horticulturae, v.821, p.63-72, 2009.
PREDIERI, S.; ZIMMERMAN, R.H. Pear mutagenesis: In vitro treatment with gamma-rays and field selection for productivity and fruit traits. Euphytica, v.117, p.217-227, 2001. Disponível em: <http://www.springerlink.com/content/ m788800444g57x17/fulltext.pdf>. Acesso em: 20 abr. 2011. doi: 10.1023/A:1026594103277.
PURUGGANAN, M.D.; FULLER, D.Q. The nature of selection during plant domestication. Nature, v.457, p.843-848, 2009. Disponível em: <http://www.agri.ankara.edu.tr/fcrops/ 1289__SELECTION.pdf>. Acesso em: 20 abr. 2011. doi:10.1038/nature07895.
REIF, J. et al. Wheat genetic diversity trends during domestication and breeding. Theoretical and Applied
Genetics, v.110, p.859-864, 2005. Disponível em: <http://
www.springerlink.com/content/7fg8xeuccvxqlcw4/fulltext.pdf>. Acesso em: 20 abr. 2011. doi: 10.1007/s00122-004-1881-8.
SAN-SAN-YI, S.A. et al. Potential loss of unique genetic diversity in tomato landraces by genetic colonization of modern cultivars at a non-center of origin. Plant Breeding, v.127, p.189-196, 2008. Disponível em: <http://link.periodicos.capes.gov.br.ez72.periodicos.capes.gov.br/ s f x l c l 3 ? u r l _ v e r = Z 3 9 . 8 8 - 2 0 0 4 & u r l _ c t x _ f m t = i n f o f i / fmt:kev:mtx:ctx&ctx_enc=info:ofi/enc:UTF-8&ctx_ver=Z39.88-2004&rfr_id=info:sid/sfxit.com:azlist&sfx.ignore_date_thresh old=1&rft.object_id=954925488804&svc.fulltext=yes>. Acesso em: 20 abr. 2011. doi:10.1111/j.1439-0523.2007.01446.x.
SIMONS, K.J. et al. Molecular characterization on the major wheat domestication gene Q. Genetics, v.172, p.547-555, 2006. Disponível em: <http://www.genetics.org.ez72.periodicos.capes.gov.br/cgi/reprint/ 172/1/547>. Acesso em: 20 abr. 2011. doi: 10.1534/ genetics.105.044727.
SHOMURA, A. et al. Deletion in a gene associated with grain size increased yields during rice domestication. Nature
Genetics, v.40, p.1023-1028, 2008. Disponível em: <http://
link .per iodicos.capes.gov.br.ez7 2.per iodicos.capes.gov.br/ s f x l c l 3 ? u r l _ v e r = Z 3 9 . 8 8 - 2 0 0 4 & u r l _ c t x _ f m t = i n f o f i / f m t : k e v : m t x : c t x & c t x _ e n c = i n f o : o f i / e n c : U T F -8 & c t x _ v e r = Z 3 9 . -8 -8 - 2 0 0 4 & r f r _ i d = i n f o : s i d / s f x i t . c o m : a z l i s t & s f x . i g n o r e _ d a t e _ t h r e s h o l d =1&rft.object_id=954925598609&svc.fulltext=yes>. Acesso em: 20 abr. 2011. doi:10.1038/ng.169.
SPOONER, D. M. et al. Extensive simple sequence repeat genotyping of potato landraces supports a major reevaluation of their gene pool structure and classification. Proceedings
of the National Academy of Sciences, v.104, n.49, p.
19398-19403, 2007. Disponível em: <http://www.pnas.org/content/ 104/49/19398.full.pdf+html>. Acesso em: 20 abr. 2011. doi: 10.1073/pnas.0709796104.
THIS, P. et al. Wine grape (Vitis vinifera L.) color associates with allelic variation in the domestication gene VvmybA1.
Theoretical and Applied Genetics, v.114, p.723-730, 2007.
Disponível em: <https://springerlink3.metapress.com/content/ 7 0 1 1 2 7 3 8 8 g 7 4 0 5 0 1 / r e s o u r c e - s e c u r e d / ? t a r g e t = f u l l t e x t . p d f & s i d = 4 g h o 0 j 3 b f w 0 rwo452vsgid3p&sh=www.springerlink.com>. Acesso em: 20 abr. 2011. doi: 10.1007/s00122-006-0472-2.
VARGHESE, M. et al. Impact of fertility variation on gene diversity and drift in two clonal seed orchards of teak (Tectona
grandis Linn. f.). New Forests, v.31, p.497-512, 2006.
Disponível em: <https://springerlink3.metapress.com/content/ 1 5 6 5 v 7 6 5 n 4 5 3 2 3 l 4 / r e s o u r c e - s e c u r e d / ? t a r g e t = f u l l t e x t . p d f & s i d = 4 g h o 0 j 3 b f w 0 rwo452vsgid3p&sh=www.springerlink.com>. Acesso em: 20 abr. 2011. doi: 10.1007/s11056-005-2178-8.
WEIRSUM, K.F. Indigenous exploitation and management of tropical forest resources: an evolutionary continuum in forest-people interactions. Agriculture, Ecosystems and
Environment, v.63, p.1-16, 1997. Disponível em: <http://
w w w . s c i e n c e d i r e c t . c o m / s c i e n c e ? _ o b = A r t i c l e U R L & _ u d i = B 6 T 3 Y 3 R H M RY 1 -1&_u ser =1 0&_ coverDa te=05 %2 F31 %2 F19 97 &_ rdoc= 1& _fmt=high&_orig=gateway&_origin=gateway&_sort=d&_docanchor=& v i e w = c & _ a c c t = C 0 0 0 0 5 0 2 2 1 & _ v e r s i o n = 1 & _ u r l Ve r s i o n = 0 & _ u s e r i d = 1 0 & m d 5 = 2 b b 8 c 6 0 a 5 6296a390000afd32fc6cbd7&searchtype=a>. Acesso em: 20 abr. 2011. doi:10.1016/S0167-8809(96)01124-3.
WILLIAM, D.E. Columbian exchange: the role of analogue crops in the adoption and dissemination of exotic cultigens. In: GOODMAN, R. Encyclopedia of plant and crop science. New York: Marcel Dekker, 2004. p.292-296.
WRIGHT, S.I.; GAUT, B.S. Molecular population genetics and the search for adaptative evolution in plants. Molecular Biology
and Evolution, v.22, p.506-519, 2005. Disponível em: <http://
mbe.oxfordjournals.org/content/22/3/506.full.pdf+html>. Acesso em: 20 abr. 2011. doi:10.1093/molbev/msi035.
YAMASAKI, M. et al. Genomic screening for artificial selection during domestication and improvement in maize. Annals of
Botany, v.100, p.967-973, 2007. Disponível em: <http://
aob.oxfor djour na ls.or g/content/10 0/5/9 67 .full.pdf+ html>. Acesso em: 20 abr. 2011. doi:10.1093/aob/mcm173. ZHU, Q. et al. Multilocus analysis of nucleotide variation of
Oryza sativa and its wild relatives: severe bottleneck during