RESUMO
Água residuária têxtil foi tratada com Aspergillus niger AN 400 em reator de bateladas sequenciais, em ciclos de 48 h para avaliar o efeito de fonte adicional de nitrogênio sobre a eficiência do sistema alimentado com efluente diluído (10% v/v) e glicose (3 g.L-1). O afluente recebeu fonte adicional de nitrogênio (etapa I), acarretando em excesso de nitrogênio amoniacal (192 mg.L-1) no meio, resultando em remoção de matéria orgânica de 85%, porém em apenas 55% de remoção de corante. Ao se diminuir a concentração média de amônia para 98 mg.L-1 (etapa II), a remoção de corante (83%) passou a ser superior à de matéria orgânica, de 69%, mostrando o efeito negativo de concentrações elevadas de amônia sobre o sistema, favorecendo ainda o crescimento de bactérias, o que provavelmente inibiu a produção de enzimas fúngicas e contribuiu para a perda de eficiência na remoção de corante e formação de subprodutos. Palavras‑chave: Índigo Carmim; bateladas sequenciais; efluente têxtil; fungos.
1Tecnóloga Ambiental. Mestre em Tecnologia e Gestão Ambiental pelo Programa de Pós-graduação em Tecnologia e Gestão Ambiental do Instituto Federal de Educação
Ciência e Tecnologia do Ceará (IFCE) – Fortaleza (CE), Brasil.
2Engenheiro Civil. Mestre em Saneamento Ambiental pela Universidade Federal do Ceará (UFC). Professor efetivo do Curso de Engenharia Ambiental do IFCE – Fortaleza (CE), Brasil. 3Farmacêutica. Doutora em Hidráulica e Saneamento pela Escola de Engenharia de São Carlos – Universidade de São Paulo (EESC‑USP). Professora efetiva do Departamento
de Química e Meio Ambiente do IFCE – Fortaleza (CE), Brasil.
4Aluna do curso de graduação em Tecnologia Ambiental do IFCE. Bolsista de trabalho em pesquisa científica do IFCE – Fortaleza (CE), Brasil..
5Aluna do curso de graduação em Tecnologia Ambiental do IFCE. Bolsista de iniciação científica do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) –
Fortaleza (CE), Brasil.
6Engenheira Civil. Doutora em Hidráulica e Saneamento pela EESC‑USP. Professora efetiva do Departamento de Química e Meio Ambiente do Instituto Federal de Educação
Ciência e Tecnologia do Ceará (IFCE) – Fortaleza (CE), Brasil.
Endereço para correspondência: Kelly Rodrigues – Rua Virgílio Paes, 2323, Cidade dos Funcionários – 60822‑465 – Fortaleza (CE), Brasil – E-mail: kellyarpessoa@gmail.com
Recebido: 19/11/12 – Aceito: 24/05/15 – Reg. ABES: 106578
Influência do excesso de nitrogênio amoniacal no
tratamento de efluente têxtil em reator de bateladas
sequenciais com
Aspergillus niger
AN 400
Influence of excess nitrogen in the treatment of the textile effluent in
sequential batch reactors with Aspergillus niger
AN 400
Karla Mayara Lima da Silva
1, Carlos Ronald Pessoa Wanderley
2, Glória Marinho
3,
Jéssica Cavalcante de Oliveira
4, Andreza Dnarla Oliveira dos Santos
5, Kelly Rodrigues
6ABSTRACT
Textile wastewater was treated by Aspergillus niger AN 400 in sequential batch reactor, which was operated in cycles of 48 h in order to evaluate the effect of an additional source of nitrogen on the system’s efficiency, which was fed with effluent diluted (10% v/v) and glucose (3 g.L-1). The influent received addition of an external source of nitrogen (phase I), in which there was excess ammonia (192 mg.L-1) in the medium, resulting in organic matter removal of 85%, but in only 55% removal of dye. In the phase II, when the average concentration of ammonia in the medium was lower (98 mg.L-1), removal of dye (83%) became higher than that of organic matter (69%), showing the negative effect of high concentrations of ammonia over the system and favoring the growth of bacteria, which probably inhibit the production of fungal enzymes and contributed to the loss of efficiency in the removal of dye and the formation of byproducts.
Keywords: Carmim Indigo, sequential batch, textile effluent, fungi.
INTRODUÇÃO
As indústrias têxteis possuem relevante importância na economia mundial e, no Brasil, tem se destacado como uma das principais em geração de empregos e faturamentos, uma vez que empregam mais de 1,7 milhões de trabalhadores, e juntas tem um faturamento anual de US$ 47 bilhões, como apresentado em Alves (2013).
Entretanto, apesar das divisas econômicas geradas, as atividades decorrentes do processamento têxtil são caracterizadas pelo consumo intenso de água e de corantes, cuja presença traz como consequência a
geração de efluentes com elevado nível de coloração (KILIÇ et al., 2007).
O uso de corantes no processo têxtil tem por função conferir cor
solúveis, não abrasivas e com capacidade elevada de absorção lumi‑ nosa, nos comprimentos de onda entre 400 e 800 nm. Porém, a mesma solubilidade que confere aos corantes a capacidade de interagir com a superfície do material, pode amplificar seu efeito tóxico (SARON & FELISBERTI, 2006), além de interferir na transparência da água, mesmo que em concentração baixa, pois a detecção de cor no meio é visível em concentrações tão baixas quanto 1 ppm (ZANONI & CARNEIRO, 2001). Desta forma, a presença de efluentes coloridos em corpos d’água acarreta uma problemática ambiental grave, por afetar a estética, a transparência da água, o processo fotossintético pelo fitoplâncton — com a diminuição da penetração da luz — e consequente diminuição do oxigênio dissolvido e ainda a solubilidade de outros gases presentes nos corpos d’água (COUTO, 2009).
Outro aspecto a ser considerado é em relação à saúde pública, pois a ingestão de água contendo corantes pode acarretar, no sistema diges‑ tivo, a formação de produtos mais nocivos do que o próprio corante por possuírem propriedades cancerígenas e de alto poder mutagênico (ANASTÁCIO FERRAZ, 2008). Essas características danosas são endos‑ sadas por Bolzon (2007), que alertou para os problemas decorrentes do contato entre corantes indigoides — de valor econômico elevado e de grande aplicação no setor industrial para o tingimento de roupas (blue jeans) — e seres humanos, ratificando a natureza carcinogênica desses compostos, que podem conduzir a toxicidades agudas e afe‑ tar o sistema reprodutivo, neurológico e o crescimento do indivíduo.
Mediante esse contexto, nos últimos anos, regulamentações rigo‑ rosas têm sido estabelecidas em muitos países em relação ao descarte de efluentes coloridos, especialmente nos países desenvolvidos e em desenvolvimento, no que diz respeito à remoção de cor dos efluentes industriais. Sua aplicação continuará a assegurar que indústrias têx‑ teis e de outros corantes tratem seus efluentes para que se ajustem aos padrões requeridos, minimizando o impacto ambiental e danos à saúde humana (ANJANEYULU; CHARY; RAJ, 2005).
Dentre as múltiplas estratégias aplicadas para a eliminação de coran‑ tes têxteis, a utilização de fungos demonstra ser uma alternativa viável
para a remoção desses compostos no meio ambiente (BANAT et al.,
1996; GIANFREDA & RAO, 2004), em vista de não resultar na presença de aminas aromáticas não degradáveis, como ocorre quando do emprego de processos anaeróbios (VAN DER ZEE & VILLAVERDE, 2005; ISIK & SPONZA, 2005), e também pela existência de dados promissores apresenta‑
dos em várias pesquisas, como as de Sanghi et al. (2009); Yang et al. (2009)
e Senthilkumar, Perumalsamy e Janardhana Prabhu (2014), entre outros.
Sanghi et al. (2009) estudaram a remoção de cor de efluente têxtil sin‑
tético pelo fungo Coriolus versicolor em reator em bateladas sequenciais.
Na presença de 100 g.L‑1 do corante Remazol Violeta Brilhante foram tes‑
tados diferentes cossubstratos: glicose (10 g.L‑1), sabugo de milho (5 g.L‑1)
e lactose (5 g.L‑1). Foram obtidas remoções de matéria orgânica, em ter‑
mos de demanda química de oxigênio (DQO), variando de 50 a 60%.
A remoção de cor foi elevada, chegando a 95 e 94%, respectivamente, quando do uso da glicose e do sabugo de milho, sendo esse percentual menor (45%) quando a lactose foi o cossubstrato adicionado ao meio.
Yang et al. (2009) utilizaram reator de leito fixo e escoamento con‑
tínuo inoculado com fungos para tratar efluente têxtil in natura, ope‑
rado com tempo de detenção hidráulica (TDH) de 12 h, com adição
de 0,5 g.L‑1 de glicose e de 0,1 g.L‑1 de (NH
4)2SO4. Foi alcançada remo‑
ção de cor de 89%. Houve predominância de Candida sp. no biofilme
em relação à população bacteriana a partir da contagem do número de colônias, com proporção mínima de 6,8:1 (fungos:bactérias), consta‑
tando‑se a não inibição da atividade enzimática dos fungos.
Senthilkumar, Perumalsamy e Janardhana Prabhu (2014) avaliaram
a remoção de cor de meio contendo 1 g.L‑1 do corante Amido Black
10B por Phanerochaete chrisosporyum. A glicose foi o cossubstrato
adicionado ao meio nas concentrações de 0,05; 0,1; 0,5; 1 e 2% (w/w). Também foi utilizada mistura de fosfato dihidrogênio de amônio e cloreto de amônio, na razão de 5:1, a qual foi adicionada nas concen‑ trações de 0,01; 0,05; 0,1; 0,5 e 1% (w/w). A maior eficiência de remo‑
ção de cor (97%) ocorreu quando da adição de 0,5 g.L‑1 de glicose e de
0,5% da fonte de nitrogênio, em seis dias de batelada.
Assim, tendo em vista o potencial dos fungos filamentosos para o tratamento de efluentes têxteis em reatores biológicos, esta pesquisa foi desenvolvida com o propósito de remover o corante Índigo Carmim
de efluente têxtil in natura diluído a 10% (v/v), utilizando reator em
bateladas sequenciais (RBS) inoculado com biomassa imobilizada de
Aspergillus niger AN 400, verificando a influência da presença e ausên‑ cia de fonte adicional de nitrogênio no meio têxtil.
METODOLOGIA
Água residuária têxtil in natura diluída a 10% (v/v)
A água residuária in natura foi coletada do tanque de equalização de
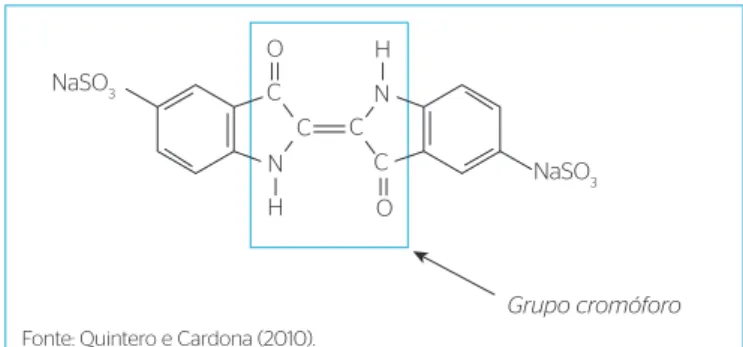
uma indústria têxtil localizada na região Metropolitana de Fortaleza, Ceará, e continha em sua composição o corante Índigo Carmim, com estrutura química representada na Figura 1. A coleta do efluente foi
NaSO3
NaSO3
Grupo cromóforo O
O C
C
C C
N
N
H
H
Fonte: Quintero e Cardona (2010).
realizada por meio de um frasco de polietileno de 5 L, previamente descontaminado com ácido clorídrico a 10% (v/v).
A água residuária têxtil in natura que preencheu o reator bio‑
lógico foi diluída a 10% (v/v) e foi, posteriormente, acrescida de
macronutrientes, na seguinte composição (em g.L‑1): (NH
4)2SO4
(0,2), NaNO3 (1,0), mgSO4 (0,25), K2HPO4 (0,2), CaCl2.2H2O (0,01),
CuSO4.7H2O (0,08), H2MoO4 (0,05), MnSO4.5H2O (0,05), Fe2(SO4)3
(0,05), ZnSO4.7(H2O) (0,04) e 1 mL.L‑1 de solução de contendo micro‑
nutrientes (solução de Vishniac).
O pH do meio aquoso in natura foi previamente ajustado para 5,
com ácido sulfúrico PA para fornecer ao Aspergillus niger pH ótimo
para seu metabolismo (GRIFFIN, 1994).
Preparo do inóculo
A espécie utilizada para o tratamento foi o Aspergillus niger AN 400 e o inóculo
foi obtido por meio da produção de esporos, cultivados em placas de Petri
esterilizadas, contendo meio de cultura Agar‑Saboraud a 30oC, tendo sido
o meio previamente esterilizado em autoclave a 121ºC, durante 15 minutos. As placas inoculadas com fungos permaneceram por cinco dias em
incubadora microbiológica à temperatura de 28ºC, tendo‑se, ao final
desse período, observado o crescimento dos esporos por toda a placa,
de modo que eeses foram então removidos, adicionando‑se em cada
placa 5 mL de solução Tween 80, e transferidos para tubos de ensaio. Posteriormente, 50 µL da suspensão de esporos foi agitada em
vórtex com adição de 950 µL de solução Tween 80, retirando‑se então
20 µL dessa mistura para leitura do número de esporos por mL, fazendo uso de câmara de Neubauer, em microscópico óptico com aumento de 400x em 16 campos.
Imobilização da biomassa em meio suporte
A espécie Aspergillus niger AN 400 foi imobilizada em espuma de
poliuretano, cortada em cubos de 1 cm de aresta, fazendo uso de fras‑ cos Erlenmeyer de 250 mL, contendo meio de crescimento, o qual era
constituído por (mg.L‑1): glicose (5000), sulfato de alumínio (0,8);
nitrato de sódio (4); sulfato de magnésio (1); fosfato de potássio dibásico anidro (0,8); cloreto de cálcio (0,04); sulfato de cobre (0,32); ácido molibídico (0,2); sulfato de manganês (0,2); sulfato férrico (0,2); e sulfato de zinco (0,16). Foi ainda adicionado ao meio a solução de
Vishiniac (1 mL.L‑1).
Quinze gramas de espumas foram previamente esterilizadas em autoclave por 20 min a 121ºC e depois colocadas em saquinhos de polietileno que foram distribuídos em 3 Erlenmeyers com volume útil de 250 mL, cada qual contendo 5 g da espuma juntamente com 150 mL do meio de crescimento.
Em seguida, inoculou‑se a suspensão de esporos, obtida do preparo
do inóculo, na concentração de 2x106 esporos.mL‑1, sendo o procedi‑
mento realizado próximo ao bico de Bunsen, para evitar contaminação.
Os Erlenmeyers foram mantidos em uma mesa agitadora horizontal,
sob agitação de 150 rpm e ±28ºC, durante 16 dias, ressaltando‑se que
dentro desse período, a cada 48 h, o meio contido no interior do reator era substituído por um meio recém preparado. Após a etapa de imobi‑ lização, as espumas foram transferidas para o reator em batelada para a partida do mesmo.
Reator em bateladas sequenciais
Foi montado reator aeróbio, com volume total de 5 L e volume reacio‑ nal de 4 L, operado em regime de bateladas sequenciais, com a biomassa de Aspergillus niger AN 400, imobilizada em espumas de poliuretano. O reator era de vidro e foi coberto por saco preto de polietileno, para
evitar degradação fotolítica. O ar era fornecido por meio de mini‑com‑
pressor, na vazão de 150 L.h‑1.
A alimentação do reator com a água residuária in natura diluída a
10% (v/v) era feita no início de cada semana e o mesmo operado em ciclos com tempo reacional de 48 h. O experimento compreendeu um total de 43 ciclos, perfazendo 86 dias de operação.
Além disso, a glicose (3 g.L‑1) foi adicionada ao meio como fonte
primária de carbono, uma vez que o próprio corante Índigo também representava outra fonte de carbono, porém de maior complexidade. A partir do 25º ciclo, foi suspensa a adição de fonte adicional de nitrogê‑ nio — sulfato de amônia e nitrato de sódio — ao efluente diluído, cons‑ tituindo assim a etapa I (1º ao 24º ciclo) e a etapa II (25º ao 43º ciclo). Foi adicionado ainda o antibiótico Megacilin Super Plus, na con‑
centração de 0,12 g.L‑1, para minimizar a contaminação por bactérias.
Análises físico‑químicas
As análises físico‑químicas foram realizadas em amostras da água resi‑
duária diluída, do afluente e efluente do reator. Foram efetuadas as deter‑ minações das variáveis: corante, DQO bruta e dissolvida e pH. Todos os métodos para determinação das variáveis mencionadas seguiram proce‑ dimentos descritos em APHA (2005), exceto a determinação de corante. A determinação da concentração de corante foi realizada com base
em Rodrigues et al. (2011), a partir da medida da absorção do grupo
cromóforo do Índigo Carmim, no comprimento de onda (λ) de 640 nm,
uma vez que o cromóforo é a estrutura responsável pela característica de coloração do corante.
Para se ter a relação entre a absorbância obtida e seu equivalente em termos de concentração de corante, foi construída curva de calibração, com o uso de solu ções de concentrações conhecidas do corante Índigo Carmim,
variando de 0 (branco) a 50 mg.L‑1. Antes da leitura, em espectrofotôme‑
tro, as amostras foram centrifugadas a 3.500 rpm, durante 15 minutos.
Desta forma, a partir dos dados de absorbância versus concentra‑
E(%) = 100 x (Co – C)/Co (1)
Co: concentração afluente (mg.L‑1); C: concentração efluente (mg.L‑1).
Verificação de co‑produtos da
degradação via espectroscopia UV-Vis
Foi realizado detalhamento espectroscópico das bandas de absorção do corante Índigo Carmim em amostras do efluente e afluente, visando verificar a possível formação de subprodutos resultantes da degrada‑ ção do corante. Para tanto, foram executadas “varreduras” entre 200
e 800 nm das alíquotas em um espectrofotômetro UV‑Vis (Thermo
Evolution 100). O corante Índigo Carmim possui uma larga banda de absorção na região do visível (640 nm), referente ao seu grupo cromó‑ foro (–C=C– ligado a CO e NH no centro da molécula) e duas outras bandas de absorção na região do ultravioleta, uma em 205 nm e outra em 265 nm, referentes à fração aromática do corante.
Devido aos problemas laboratoriais somente foram realizadas var‑ reduras na etapa I, nos ciclos C16 e C19.
Contagem das unidades formadoras
de colônias de micro‑organismos
A determinação das unidades formadoras de colônias ocorreu no final
das etapas I e II, procedendo‑se à diluição em série da amostra do
efluente de cada uma das etapas. Após a diluição, fez‑se o plaqueamento
em placas de Petri contendo meios seletivos: meio Martin, usado para
a contagem de colônias de fungos, e meio nutriente‑ágar, para conta‑
gem do número de colônias de bactérias.
O meio de Martin era composto de (g.L‑1): K
2HPO4 (1); peptona (5);
KH2PO4 (0,5); mgSO4.7H2O (0,5); dextrose (10); extrato de levedura (0,5);
rosa bengala (0,033); ágar (18). Além disso, foi adicionado 3 µg.mL‑1
de estreptomicina, antibiótico usado para evitar contaminação do meio
por bactérias. O meio nutriente‑ágar possuía (g.L‑1): bacto‑agar (15);
extrato de carne (2); peptona (5); NaCl (8).
Desta forma, a amostra do efluente (10 mL) foi misturada em 90 mL de solução salina (0,89%) em tubos, os quais foram posteriormente agitados em vórtex por 10 min. Então, alíquotas de 1 mL foram reti‑ radas desses tubos e transferidas para 9 mL de solução salina e, dessa forma, o mesmo procedimento foi feito sucessivamente até obter con‑
centrações de 10‑2, 10‑3 e 10‑4. Em seguida, as diluições em série obtidas
foram plaqueadas nos meios seletivos de Martin e nutriente‑ágar, com
ajuda de alça de Drigalski.
Ensaio de adsorção do corante no biofilme
A quantidade de Índigo Carmim presente no biofilme foi estimada reti‑
rando‑se um cubo contendo biomassa imobilizada do reator, após o último
ciclo, sendo então transferido para Erlenmeyer de 250 mL, adicionado
de 100 mL de solução salina e pérolas de vidro. Em seguida, o conjunto foi submetido à agitação vigorosa para propiciar a remoção de biomassa do meio suporte. O meio foi então centrifugado a 3.500 rpm e o sobrena‑ dante foi separado da biomassa. O metanol foi utilizado como solvente para remover o corante adsorvido, de modo que 10 mL foram transferi‑ dos juntamente com a biomassa para tubos, permanecendo em contato durante 10 min. Depois desse período, foi adicionado mais 5 mL de meta‑ nol, seguido de homogeneização e leitura espectrofotométrica a 640 nm.
Ensaio de adsorção no meio suporte
Foi utilizado frasco de vidro de 2 L, ao qual foram adicionados 15 g do material suporte previamente seco em estufa. Esse recipiente recebeu 1 L de água residuária diluída (20% v/v).
A capacidade máxima de adsorção do corante na espuma de poliure‑ tano, empregada como material suporte, foi monitorada a partir da sua
concentração inicial no meio durante um período de 1,83 h, retirando‑se
amostras do meio a cada 10 minutos para determinação da concentra‑ ção de saturação e a respectiva massa de corante adsorvida à mesma.
RESULTADOS E DISCUSSÕES
Na etapa I, a concentração média de corante no afluente foi de 10 mg.L‑1,
variando de 3 a 20 mg.L‑1, pois o efluente foi oriundo de processamento
têxtil, apresentando alterações em função do tipo e quantidade de pro‑ duto produzido, bem como de técnicas empregadas. Nessa etapa, foi observado decaimento da concentração inicial do Índigo Carmim,
registrando‑se remoção média do corante de 55%, máxima de 97%
(ciclo C24) e mínima de 3% (ciclo C6).
É importante mencionar que, em metade dos ciclos operacionais da etapa I, a eficiência de remoção do Índigo Carmim não foi boa, ocor‑ rendo ciclos em que as remoções foram inferiores à média registrada para a etapa (55%) e outros em que não houve remoção do corante, nos quais a concentração do mesmo no efluente foi superior à do afluente (Figura 2). A perda da eficiência foi atribuída à suplementação do meio com os compostos nitrogenados, pois concentrações elevadas de nitrogê‑
nio amoniacal, como as encontradas no presente trabalho (192 mg.L‑1),
inibiram a assimilação do corante pelos fungos (KHELIFI et al., 2009;
De acordo com Debrassi, Largura e Rodrigues (2011), o acúmulo de corante no meio é propiciado pela competição por sítios de adsorção
entre o corante e íons OH‑ e, conforme há o aumento do valor do pH
no meio, o número de sítios carregados positivamente diminui, o que resulta na redução da adsorção do corante devido à repulsão eletrostática.
Na presente pesquisa, na etapa I, o valor médio do pH foi 8 (variando de 5,9 a 9), ao passo que na etapa II, esse valor foi de 5,6 (variando de 4 a 7,6). Os maiores valores de pH registrados na etapa I estariam rela‑ cionados com as reações de conversão de nitrato a nitrito pelos fungos,
ocorrendo liberação de OH‑, o que favorece a dessorção do poluente
(GRIFFIN, 1994).
Quando o micélio está protonado, o que pode ocorrer em meio ácido, a velocidade de adsorção de corante tende a aumentar. Chaves
et al. (2008), ao utilizarem biomassa morta de Aspergillus niger verifica‑ ram maior adsorção do corante quando a superfície do micélio estava
protonada, após tratamento com solução ácida de H2SO4, e perda da
capacidade biossortiva da massa micelial quando esta foi tratada com solução alcalina de NaOH.
De fato, na etapa II, com os valores mais baixos de pH, influencia‑ dos pela retirada da fonte externa de nitrogênio, houve, possivelmente, diminuição da tendência à dessorção do corante, com a produção de enzimas atuantes na degradação de corantes e outros poluentes sendo
favorecida (KHELIFI et al., 2009; SWAMY & RAMSAY, 1999). Isso
foi evidenciado pela melhora nos percentuais de remoção do Índigo Carmim, com eficiência média de 83%, máxima de 97% (ciclos C26 e C40) e mínima de 69% (ciclo C39). Nessa etapa, a concentração média
de corante afluente foi de 10 mg.L‑1, variando de 5 a 19 mg.L‑1.
É importante mencionar que embora tenha sido observada a adsor‑ ção de corante ao material suporte (0,064 g) e no biofilme (0,32 g),
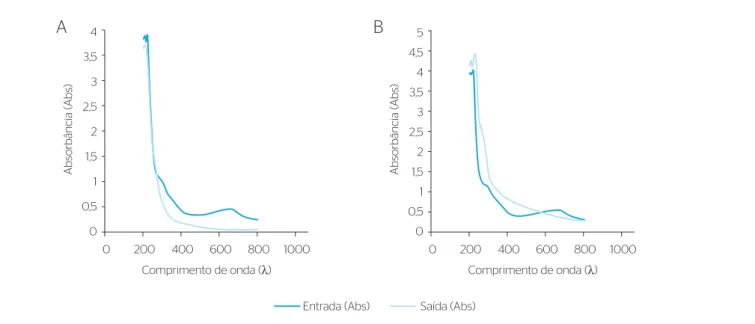
perfazendo massa de corante adsorvida de 0,38 g, essa parcela foi infe‑ rior à massa total de corante removido pelo reator (2,11 g), indicando que a via biológica foi o principal mecanismo de remoção do corante. Na Figura 2 são apresentadas as varreduras realizadas no afluente e
efluente dos ciclos C16 e C19, contatando‑se diminuição na área rela‑
tiva ao grupo cromóforo da molécula do corante Índigo Carmim, as quais foram de 51 e 57%, respectivamente.
Nos ciclos C16 e C19, apesar de ter ocorrido ruptura da ligação
química do grupo cromóforo, –C=C– (λ=640 nm), não houve a uti‑
lização efetiva do anel benzeno (λ=205 nm) que compõe a molécula
do corante, tendo‑se, no ciclo C16, obtido remoção muito pequena
(Figura 2A), de apenas 4%. Paralelamente, detectou‑se acréscimo de
compostos aromáticos no comprimento de onda de λ=265 nm.
Já no ciclo C19, registrou‑se aumento do valor das absorbâncias
para as duas bandas espectrais (λ=205 e 265 nm), como demonstrado
na Figura 2B, o que estaria relacionado à formação de subprodutos decorrentes da ruptura da própria molécula do corante, sendo que o
efluente obtido no final dos ciclos dessa etapa apresentava‑se turvo.
Verificou‑se que, apesar das oscilações de matéria orgânica no
afluente, em decorrência do próprio processo industrial — tratando‑se
o meio de efluente “in natura” —, estas não influenciaram na remoção
de DQO bruta e dissolvida, evidenciando a fácil adaptação da biomassa fúngica às condições do meio.
Na etapa I, a remoção média de matéria orgânica dissolvida foi 85%, com máxima de 99% (ciclo 16) e mínima de 70% (ciclo C22),
o que representou em termos de concentração no efluente, 23 mg.L‑1
(ciclo C16) e 561 mg.L‑1 (ciclo C22).
Para a DQO bruta, a eficiência média de remoção para a etapa I foi de 75%, com máxima de 94% (ciclo 14) e mínima de 51% (ciclo C22),
Figura 2 – Varredura do corante Índigo Carmim no ciclo C16 (A) e ciclo 19 (B), durante a operação do reator em bateladas sequenciais na etapa I. Comprimento de onda (λ)
Entrada (Abs) Saída (Abs)
Absorbância (Abs)
4
A
B
3,5 3 2,5 2 1,5 1 0,5 0
0 200 400 600 800 1000
Comprimento de onda (λ)
Absorbância (Abs)
4 3,5 3 2,5 2 1,5 1 0,5 0
0 200 400 600 800 1000
resultando em termos de concentração para o efluente, 208 mg.L‑1
(ciclo C14) e 1155 mg.L‑1 (ciclo C22).
Nessa etapa, de modo geral, a eficiência de degradação do corante foi inferior à registrada para remoção de matéria orgânica dissolvida, sendo que, nos ciclos C2, C6, C7, C9, C17 e C18, os percentuais de remoção obtidos para o corante foram muito baixos em relação às efi‑ ciências tanto de matéria orgânica bruta quanto dissolvida, indicativo de que a maior parte da matéria orgânica removida estaria relacionada com o uso da glicose e outros constituintes da água residuária pelos micro‑organismos, tendo o Índigo Carmim sido removido parcialmente.
Nos ciclos C1, C3, C5, C11, C14, C15, C23 e C24 os percentuais de eficiências de degradação do corante acompanharam os de matéria orgânica dissolvida, fato que permitiu inferir que nos ciclos menciona‑ dos houve não somente a remoção do cromóforo, mas, provavelmente, melhor assimilação de outros componentes da estrutura molecular do corante, bem como menor formação de excretas pelos fungos.
Por outro lado, nessa mesma etapa, embora nos ciclos C10, C12, C13, C21 e C22 não tenha ocorrido eficiência de remoção do corante, com aumento da concentração do Índigo Carmim no efluente ao final dos ciclos, foram registradas remoções elevadas de matéria orgânica dissolvida, respectivamente, 88; 77; 81; 84 e 70%. Pode ter ocorrido desprendimento em maior quantidade de parte do corante que estaria adsorvido ao biofilme/material suporte, indi‑ cando que na etapa I houve instabilidade do sistema, com momen‑ tos de eficiência baixa ou nenhuma remoção de corante na maior parte da operação do RBS.
A instabilidade do sistema observada na etapa I foi atribuída ao excesso de nitrogênio, o que afeta diretamente o funcionamento do sistema enzimático e causa alterações no metabolismo secundário da célula (LEATHAM & KIRK, 1983), inibindo a assimilação do corante pelos fungos (SWAMY & RAMSAY, 1999).
Vahabzadeh, Mehranian e Saatari (2004) relataram que a remo‑ ção de cor é afetada pela concentração de nitrogênio amoniacal, pois a velocidade de ruptura da molécula diminui quando esse íon encon‑
tra‑se presente em excesso.
A importância da fonte de nitrogênio sobre a remoção de cor e degradação do corante também foi relatada por Levin, Melignani e Ramos (2010), que apontaram para sua influência sobre a produção
de enzimas lignolíticas pelos fungos Trametes trogii, Trametes villosa
e Coriolus versicolor var. antarcticus, em meio mineral contendo gli‑
cose (20 g.L‑1) e variadas fontes de nitrogênio, as quais foram utiliza‑
das separadamente, sendo adicionadas ao meio na concentração de
0,75 g.L‑1. Verificou‑se que ácido glutâmico seguido de peptona foram
excelentes para a produção de maiores quantidades das enzimas lac‑ case e manganês peroxidase, importante arsenal para a degradação de
poluentes orgânicos complexos, chegando‑se à produção de, respecti‑
vamente, a 1188,3 e 4,5 U.mL‑1.
Senthilkumar, Perumalsamy e Janardhana Prabhu (2011) estu‑ daram o efeito da concentração de nitrogênio sobre a degradação
de água residuária têxtil sintética contendo 1 g.L‑1 de Amido Black
10 pelo fungo Phanerochaete chrysosporium. Os autores utilizaram
NH4H2PO4 e NH4Cl como fonte de nitrogênio, compreendendo mis‑
tura na razão de 5:1, que foi adicionada à água residuária sintética a fim de perfazer concentrações de 0,01; 0,05; 0,1; 0,5 e 1% (w/w). Embora se tenha constatado aumento da remoção de cor do meio com o aumento da concentração de 0,01 a 0,5%, houve redução acentuada da remoção de cor na concentração de 1%, ratificando o efeito nega‑ tivo do excesso de nitrogênio sobre a remoção de cor e de corante, o qual na pesquisa mencionada também se encontrava na forma de nitrogênio amoniacal.
Niebisch et al. (2010) avaliaram o efeito de diferentes fontes de nitro‑
gênio — nitrato de sódio, ureia, tartarato de amônia, cloreto de amônio, peptona e oxalato de amônio —, com concentrações variando de 5 a
15 g.L‑1, sobre a remoção de cor de meio contendo corante Azul Reativo
220 (0,1 g.L‑1), na presença de glicose (10 g.L‑1), por Lentinus crinitus.
O experimento foi conduzido ao longo de 15 dias, a 28ºC, na ausência
de luz, e o oxalato de amônio (10 g.L‑1) foi a melhor fonte de nitrogê‑
nio, resultando na maior remoção de cor, em torno de 90% (10º dia).
Eficiências tão elevadas quanto à encontrada por Niebisch et al.
(2010) também foram registradas na presente pesquisa, quando da suspensão da adição de fonte externa de nitrogênio (etapa II), com remoção máxima de Índigo Carmim, de 97%, em apenas 48 h, em
menor concentração de glicose (3 g.L‑1) e em maior concentração ini‑
cial do corante (4 a 20 mg.L‑1), ainda que se trate de corantes diferentes.
Ainda na etapa II, apesar da melhora acentuada na remoção de Índigo Carmim, foi registrada diminuição da remoção de matéria orgânica. A eficiência média de DQO bruta foi de 64%, com máxima de 90% (ciclos C33 e C34) e mínima de 23% (ciclo C26). Em relação à matéria orgânica dissolvida, em termos de DQO, a média de remoção alcançada nessa etapa foi 69%, com máxima de 90% (ciclos 33 e 34), e mínima de 43% (ciclo C23).
Observou‑se que na etapa II ocorreu diminuição na média de
remoção de DQO dissolvida em comparação com a etapa I (85%),
registrando‑se percentual médio de 69%. Contudo, houve aumento
da eficiência média de remoção de corante, com remoção média de corante de 83%, máxima de 97% (ciclos C26 e C40) e mínima de 69% (ciclo C39), o que indicou a melhor resposta do sistema após a retirada da fonte adicional de nitrogênio.
Semelhantemente, houve também a diminuição da eficiência do tratamento em relação à remoção média de matéria orgânica bruta (64%) em comparação com a etapa I, quando foi registrada eficiência média de remoção de 75%.
Assim, na etapa II, possivelmente tenha ocorrido melhor atividade enzimática, a qual antes teria sido fortemente inibida pela concentração
elevada de nitrogênio amoniacal no afluente (192 mg.L‑1) que diminuiu
para 98 mg.L‑1. Nessa condição, ocorreu o melhor desempenho do sistema,
com o alcance de maior remoção do poluente e, provavelmente, de seus subprodutos, apresentando efluente muito clarificado no final dos ciclos. Na referida etapa, o efluente apresentou os menores valores de pH, entre 4 e 7,5, cujo valor médio foi de 5,6, sendo que nessa condi‑ ção ocorre maior produção de enzimas, conforme relatado por Spier (2005), o qual frisou ainda que a produção de enzimas atinge seu ótimo em valores de pH em torno de 5.
Na etapa I, a adição de 3 g.L‑1 de glicose teria resultado na produ‑
ção de ácidos orgânicos (ZNAD; MARKOS; BALES, 2004) que devem ter sido prontamente consumidos do meio pelos micro‑organismos, o que, aliado ao excesso de compostos com nitrogênio adicionados na
água residuária têxtil in natura, como nitrato de sódio e sulfato de amô‑
nio, contribuiu para a elevação do pH do meio nos ciclos operacionais.
Sabe‑se que esses compostos regulam equilíbrio metabólico da célula
fúngica, variando de acordo com suas necessidades (SINGH, 2006).
Neste trabalho, a espécie Aspergillus niger AN 400, utilizada como
inóculo no reator em bateladas sequenciais, encontrava‑se no meio
ao final da operação das etapas I e II, sendo que, na etapa I, foi regis‑ trado maior número de colônias de bactérias em relação às de fungos,
20x107 e 15x107 unidades formadoras de colônia por mL (UFC.mL‑1),
respectivamente, resultando em relação de 1:0,75 (bactérias:fungos).
Já na etapa II, o número de colônias de fungos foi de 157x103 UFC.mL‑1,
inferior ao valor encontrado na etapa I (17x107 UFC/mL), porém supe‑
rior à população de bactérias verificada para essa mesma etapa (etapa
II), a qual foi de 90x103 UFC.mL‑1. Assim, na etapa II foi obtida a rela‑
ção bactérias:fungos de 0,57:1, indicando que na condição de menor
concentração de nitrogênio amoniacal (98 mg.L‑1), os fungos tiveram
melhores condições de se sobressair em relação às bactérias e de meta‑ bolizar os substratos presentes, provavelmente, pela não inibição de seu sistema enzimático, fato este que teria ocorrido na etapa I, devido ao excesso de nitrogênio.
Ainda, a concentração na qual o antibiótico foi usado na presente pesquisa não inibiu o crescimento da população de bactérias. Porém, a população bacteriana não cresceu predominando no meio, ao ponto de exercer efeito negativo sobre os fungos, fato que poderia ocasionar perda da eficiência do sistema, motivado também pela repressão do
sistema enzimático fúngico (YANG et al., 2009).
Contudo, de acordo com Lopes et al. (2011), é importante ressaltar
que o tratamento biológico aplicado ao efluente não visa à operação de reatores sob condições assépticas, pois no tratamento de águas resi‑ duárias, diferentemente da indústria de fermentação, isto é impensável. Além do mais, ainda segundo os autores, a ação em conjunto de fungos e bactérias é favorável ao processo de biorremediação, pois estes são os maiores decompositores encontrados na natureza e são responsáveis pela mineralização de vários poluentes, muitos deles com maior eficiên‑ cia de remoção quando metabolizados em culturas mistas (GRIFFIN,
1994; HAI et al., 2008; SINGH, 2006).
De acordo com Yang et al. (2009), fungos e bactérias podem esta‑
belecer uma relação de simbiose, na qual os fungos atuam na ruptura de moléculas complexas, as quais as bactérias presentes no meio não são capazes de utilizar ao ponto de se obter um grau de mineralização elevado, e então os subprodutos oriundos da ruptura dessas moléculas estariam disponíveis para serem utilizados por fungos e bactérias que atuariam em conjunto no tratamento do despejo.
CONCLUSÕES
Na presença de concentração elevada de amônia (192 mg.L‑1), houve
eficiência baixa de remoção de corante e de matéria orgânica e, nessa condição, maior acúmulo de subprodutos no meio, decorrentes da utilização das fontes de carbono, com grande persistência do benzeno oriundo da ruptura da molécula do Índigo Carmim.
Quando a fonte adicional de nitrogênio deixou de ser adicionada ao meio, ocorreu melhor desempenho de sistema e maior número de colônias de fungos em relação ao de bactérias, favorecendo a não repressão do sistema enzimático fúngico, importante na degradação dos poluentes.
Assim, na etapa II, marcada pela supressão da fonte adicional de nitrogênio, a população de fungos predominou no reator indicando que os mesmos teriam encontrado melhores condições de cresci‑ mento e de assimilação dos substratos, sendo importante ressaltar a necessidade de otimizar o sistema e a realização de mais estudos sobre mecanismos que propiciem obter maior grau de mineralização da molécula do corante.
AGRADECIMENTOS
Os autores agradecem a Deus, que nos permitiu chegar até aqui; ao Conselho Nacional de Pesquisa (CNPq) pela concessão das bolsas de iniciação científica e financiamento da pesquisa pelo Processo
no. 475831/2010‑1 e à Coordenação de Aperfeiçoamento de Pessoal de
ALVES, F.C. (2013) Estudo dos processos de adsorção utilizando argilas como adsorventes para remoção do corante verde malaquita.
Dissertação (Mestrado em Agroquímica), Universidade Federal de Lavras, Minas Gerais.
ANASTÁCIO FERRAZ, E.R. (2008) Comparação da mutagenicidade dos azo corantes Disperse Red 1, Disperse Orange 13 e Disperse Red 13 utilizando o teste de mutagenicidade Salmonella. Dissertação (Mestrado). Universidade de São Paulo, Ribeirão Preto.
ANJANEYULU, Y.; CHARY, N.S.; RAJ, D.S.S. (2005) Descolourization of industrial effluents – available methods and emerging technologies – a review. Reviews in Environmental Science and Biotechnology, v. 4, n. 4, p. 245‑273.
APHA – AMERICAN PUBLIC HEALTH ASSOCIATION. (2005) Standard methods for the examination of water and wastewater. 21st edition. Washington: APHA.
BANAT, I.M.; NIGAM, P.; SINGH, D.; MARCHANT, R. (1996) Microbial decolorization of textile‑dyecontaining effluents: a review. Bioresource Technology, v.58, n.3, p.217‑227.
BOLZON, L.B. (2007) N2b2O5 como fotocatalizador para a degradação de índigo carmina. Dissertação (Mestrado em Química), Universidade de Brasília, Brasília.
CHAVES, K.O.; MONTEIRO, C.R.L.; MUNIZ, C.R.; GOMES, R.B.; BUARQUE, H.L.B. (2008) Adsorção de índigo carmim em biomassas mortas de
Aspergillus niger. Engenharia Sanitária e Ambiental, v. 13, n. 4, p. 351‑355.
COULIBALY, L.; GOURENE, G.; AGATHOS, S.N. (2003) Utilization of fungi for biotreatment of raw wastewaters. African Journal of Biotechnology, v. 2, n. 12, p. 620‑630.
COUTO, R.S. (2009) Dye removal by immobilised fungi. Biotechnology Advances, v. 27, n. 3, p. 227‑235.
DEBRASSI, A.; LARGURA, M.C.T.; RODRIGUES, C.A. (2011) Adsorção do corante vermelho congo por derivados da o‑carboximetilquitosana hidrofobicamente modificados. Química Nova, v. 34, n. 5, p. 764‑770.
GIANFREDA, L. & RAO, M.A. (2004) Potential of extra cellular enzymes in remediation of polluted soils: a review. Enzyme and Microbial Technology, v. 35, n. 4, p. 339‑354.
GRIFFIN, D.H. (1994) Fungal physiology. 2nd ed. New York: Wiley‑Liss. 458 p.
HAI, F.I.; YAMAMOTO, K.; NAKAJIMA, F.; FUKUSHI, K. (2008) Removal of structurally different dyes in submerged membrane fungi reactor– biosorption/PAC‑adsorption and biodegradation. Journal of Membrane Science, v. 325, n. 1, p. 395‑403.
ISIK M.; SPONZA, D.T. (2005) Effects of alkalinity and co‑substrate on the performance of an upflow anaerobic sludge blanket (UASB) reactor through decolonization of Congo Red azo dye. Bioresource Technology, v. 96, n. 5, p. 633‑643.
KHELIFI, E.; AYED, L.; BOUALLAGUI, H.; TOUHAMI, Y.; HAMDI, M. (2009) Effect of nitrogen and carbon sources on Indigo and Congo red decolourization by Aspergillus alliaceus strain 121C. Journal of Hazardous Materials, v. 163, n. 2‑3, p. 1056‑1062.
KILIÇ, N.K.; NIELSEN, J.L.; YÜCE, M.; DÖNMEZ, G. (2007) Characterization of a simple bacterial consortium for effective treatment of wastewaters with reactive dyes and Cr(VI). Chemosphere, v. 67, n. 4, p. 826‑831.
LEATHAM, G.F.; KIRK, T.K. (1983) Regulation of ligninolytic activity by nutrient nitrogen in white‑rot basidiomycetes. FEMS Microbiology Letters, v. 16, n. 1, p. 65‑67.
LEVIN, L.; MELIGNANI, E.; RAMOS, A.M. (2010) Effect of nitrogen sources and vitamins on ligninolytic enzyme production by some white‑rot fungi: dye decolorization by selected culture filtrates. Bioresource Technology, v. 101, n. 12, p. 4554‑4563.
LOPES, M.S.S.; OLIVEIRA, P.C.C; ANDRADE, M.V.F.; ARAÚJO, R.S.; MARINHO, G.; RODRIGUES, K. (2011) Remoção de macronutrientes de efluente da indústria de castanha de caju por uso de reator aeróbio em batelada com inóculo fúngico. Engenharia Sanitária e Ambiental, v. 16, n. 1, p. 17‑26.
NIEBISCH, C.H.; MALINOWSKI, A.K.; SCHADECK, R.; MITCHELL, D.A.; KAVA‑CORDEIRO, V.; PABA, J. (2010) Decolorization and biodegradation of reactive blue 220 textile dye by Lentinus crinitus extracellular extract.
Journal of Hazardous Materials, v. 180, n. 1‑3, p. 316‑322.
QUINTERO, L.; CARDONA, S. (2010) Tecnologías para la decoloración de tintes índigo e índigo carmín. Revistade la Facultad de Minasde la UniversidadNacional de Colombia, v. 77, n. 162, p. 371‑386.
RODRIGUES, K.; SILVA, K. M. L.; SILVA, G. M. M.; LIMA, P. C. C.; WANDERLEY, C. R. P.; SILVA, G. M. (2011) Remoção de corante por uso de Aspergillus niger AN400 em reator em bateladas sequenciais. Química Nova, v. 34, n. 7, p. 1119‑1123.
SANGHI, R.; DIXIT, A.; VERMA, P.; PURI, S. (2009) Design of reaction conditions for the enhancement of microbial degradation of dyes in sequential cycles. Journal of Environmental Sciences, v. 21, n. 12, p. 1646‑1651.
SANGHI, R.; DIXIT, A.; GUHA, S. (2006) Sequential batch culture studies for the decolorization of reactive dye by Coriolus versicolor.
Bioresource Technology, v. 97, n. 3, p. 396‑400.
SARON, C. & FELISBERTI, M.I. (2006) Ação de colorantes na degradação e estabilização de polímeros. Química Nova, v. 29, n. 1, p. 124‑128.
SENTHILKUMAR, S.; PERUMALSAMY, M.; JANARDHANA PRABHU, H. (2014) Decolourization potential of white‑rot fungus Phanerochaete chrysosporium on synthetic dye bath effluent containing Amido Black 10B. Journal of Saudi Chemical Society, v. 18, n. 6, p. 845‑853.
SINGH, H. (2006) Mycoremediation: fungal bioremediation. Canada: Wiley. 617 p.
SPIER, M.R. (2005) Produção de enzimas amilolíticas fúngicas alfa‑amilase e amiloglucosidase por fermentação no estado sólido. Dissertação (Mestrado em Tecnologia de Alimentos), Universidade Federal do Paraná, Curitiba.
SWAMY, J. & RAMSAY, J.A. (1999) Effects of glucose and NH4 +
concentrations on sequential dye decoloration by Trametes versicolor.
Enzyme and Microbial Technology, v. 25, n. 3‑5, p. 278‑284.
VAN DER ZEE, F. & VILLAVERDE, S. (2005) Combined anaerobic‑aerobic treatment of azo dyes: a short review of bioreactor studies. Water Research, v. 39, n. 8, p. 1425‑1440.
VAHABZADEH, F.; MEHRANIAN, M.; SAATARI, A.R. (2004) Color removal ability of Phanerochaete chrysosporium in relation to lignin peroxidase and manganese peroxidase produced in molasses wastewater. World Journal of Microbiology and Biotechnology, v. 20, p. 859‑864.
YANG, Q.; LI, C.; LI H.; LI, Y.; YU, N. (2009) Degradation of synthetic reactive azo dyes and treatment of textile wastewater by a fungi consortium reactor. Biochemical Engineering Journal, v. 43, n. 3, p. 225‑230.
WANG, B.E. & HU, Y.Y. (2008) Bioaccumulation versus adsorption of reactive dye by immobilized growing Aspergillus fumigatus beads.
Journal of Hazardous Materials, v. 157, n. 1, p. 1‑7.
ZNAD, H.; MARKOS, J.; BALES, V. (2004) Production of gluconic acid from glucose by Aspergillus niger: growth and non‑growth conditions.
Process Biochemistry, v. 39, n. 11, p. 1341‑1345.