R E S I S T Ê N C I A À S U B M E R S Ã O D E Á C A R O S ( A C A R I : RIBATIDA) T E R R E S T R E S D E F L O R E S T A S I N U N D Á V E I S Ε D E T E R R A F I R M E N A A M A Z Ô N I A C E N T R A L E M C O N D I Ç Õ E S E X P E R I M E N T A I S D E L A B O R A T Ó R I O
Elizabeth Ν . FRANKLIN1
, R o s i l e t e z L. GUIMARÃES2
, Joachim ADIS3
, Herbert O. R. SCHUBART4
RESUMOzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA — Espécimens de Rostrozetes foveolatus foram obtidos de duas florestas inundáveis (várzea e igapó) e de duas florestas secundárias (terra firme) da Amazônia Central. As populações foram comparadas para a obtenção da taxa de sobrevivência em experimentos de laboratório nas condições submersas e não submersas. As coletas no igapó foram efetuadas em três períodos de 1992: antes (fevereiro: serapilheira não submersa), no início (abril: serapilheira submersa) e no pico da inundação (julho: serapilheira submersa). No igapó, os animais sobreviveram melhor em condição não submersa, significando que a submersão é um fator de estresse. O mesmo não ocorreu na várzea, indicando que esta população está melhor adaptada ao estresse da submersão. A menor resistência à submersão (27 dias) foi registrada nos animais de uma floresta secundária da terra firme, situada longe do igapó. Na população da terra firme próxima ao igapó, a resistência a submersão foi comparável à registrada para as populações das áreas inundáveis e maior do que a registrada em terra firme longe do igapó. Em experimentos com animais coletados em fevereiro de 1996 (mantidos individualmente em recipientes plásticos), as populações das florestas inundáveis tiveram taxa de sobrevivência significativamente maior em relação aos das florestas não inundáveis. Três situações foram registradas quanto a resistência à submersão: a) a da várzea, com população mais resistente, b) a do igapó, com uma população intermediária e c) a da terrafirme longe do igapó, com uma população menos resistente. Em doze das outras quinze espécies estudadas foi registrada alta resistência às condições de submersão.
Palavras-chave: Acari: Oribatida, inundação, áreas inundáveis, ácaros do solo, Amazônia, Neotrópicos
The Resistance to Submersion of Terrestrial Acari (Acari: Oribatida) from Flooded and Non-flooded Forests of Central Amazonia in Experimental Laboratorial Conditions.
ABSTRACT — Specimens ofzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Rostrozetes foveolatus were obtained from two inundated forest areas
("várzea" and "igapó") and from two nonflooded secondary forests in the Central Amazon Region. The
survival rates of their populations under submerged and nonsubmerged conditions were compared in laboratory conditions. The "igapó" population was sampled during three periods in 1992: before (February: litter nonsubmerged), in the beginning (April: litter submerged) and in the peak of the inundation phase
(July: litter submerged). At "igapó", the animals survived better when submitted to nonsubmerged conditions, meaning that submersion is a stress factor. The same result did not occur at "várzea", signifying a better adaptation of this population to the submersion stress. The lowest flood resistance (27 days) was
found in animals from "terra firme" secondary forest, away from the igapó. A considerable submersion resistance in the population sampled in the secondary forest, near the igapó forest, was comparable to the populations of the inundated areas and greater than the resistence at "terrafirme", far away from the "igapó". Animals sampled in February of 1996 (reared individually in plastic containers), from the flooded forests had significantly higher survival rates than those from secondary forests. Three situations in relation to the submersion period were registered: a) one of "várzea", with a more resistant population, b) one of "igapó", with a intermediate resistance and c) one of "terrafirme", away from the "igapó", with a lower resistant population. In twelve of the fifteen species studied, a high resistance to submersion was also registered.
Key-words: flood resistance, Acari: Oribatida, inundation, floodplains, soil mites, Amazon, Neotropics.
1
INPA/CPEc, Caixa Postal 478, 69.011.970, Manaus (AM), Brasil.
2
Bolsista PIBIC/CNPq, ΙΝΡΑ/CPEc, Caixa Postal 478, 69.011.970, Manaus (AM), Brasil
3
Instituto MaxPlanck para Limnologia, Caixa Postal 165, D24302 Plön Alemanha.
4
I N T R O D U Ç Ã O zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Na A m a z ô n i a C e n t r a l , a flutuação anual do nível das águas dos rios atinge uma média de 10 metros de diferença entre o ciclo de cheia e vazante. As florestas ao longo destes rios são anualmente inundadas durante c i n c o a sete m e s e s . As d r á s t i c a s mudanças entre a fase terrestre e a aquática causadas por esse "pulso de
i n u n d a ç ã o " (JunkzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA et. al., 1989),
induziram adaptações dos organismos e muitas populações de espécies de a n i m a i s diferem e c o e fenologicamente, assim como, em p a r t e , m o r f o e g e n e t i c a m e n t e , daquelas das florestas de terra firme. Essas mudanças e adaptações foram d e n o m i n a d a s " e s t r a t é g i a s de sobrevivência" (Adis, 1992; 1997a, b).
Os oribatídeos são componentes importantes da serapilheira e do solo devido à alta densidade, hábitos (são catalisadores da decomposição, participando da fragmentação dos restos orgânicos, e dispersando a microflora) e nutrição (macrofitófagos, microfitófagos, panfitófagos, zoófagos, necrófagos e coprófagos) (Luxton,
1975). Segundo Beck (1968), os oribatídeos terrestres possuem diferentes reações à inundação: a espécie
Rostrozetes foveolatus Sellnick, 1925
p e r m a n e c e ativa na serapilheira submersa, enquanto que Eremobelba
foliata Hammer, 1958 resiste em forma
de ovo. A resistência de oribatídeos à submersão é influenciada por fatores como temperatura, salinidade, oxigenaçào e movimento da água, ritmo da inundação, tipo de substrato e época do ano. Espécies de gêneros
talossobiontes como Ameronothrus,
Platynothrus e Hermannia, podem
sobreviver em água do mar (41 a 249 dias) e em água doce (109 a 177 dias). Algumas espécies terrestres como
Steganacarus, Damaeus, Xenillus, Euzetes e Nothrus, possuem maior
resistência em água doce (74 a 226 dias) do que em água salgada (6 a 36 dias) (Schuster, 1965; 1966; 1978). Weigmann (1973) registrou para
Hermannia subglabra Berlese, 1910
resistência de 210 dias em água doce e de 56 a 182 dias em água salgada; para
Halozetes intermedins Wall work, 1963
a resistência foi de 150 dias em água doce e de 80 dias em água salgada.
A influência da enchente anual na mudança de populações destes á c a r o s e a d i v i s ã o das d i v e r s a s espécies de oribatídeos em grupos, de acordo com suas reações às condições submersas foram investigadas por Beck (1971; 1972; 1976) em área de igapó na Amazônia Central. Beck (1972) observou que R. foveolatus tolera a submersão na forma adulta, h a v e n d o r e d u ç ã o a c e n t u a d a de indivíduos ao final da inundação, mas cerca de 10% sobrevive ao período submerso. Baseandose no fato, o a u t o r s u p õ e q u e , em c o n d i ç õ e s extremas de inundação, os animais d e v e m d e s e n v o l v e r a d a p t a ç õ e s fisiológicas para se adaptar a essa situação, j á que grande parte das espécies de ácaros oribatídeos até agora encontradas nestas regiões, estão também em terra firme na mesma ou quase na m e s m a p r o p o r ç ã o . E s t a "condição prévia", é conhecida como p r é a d a p t a ç ã o ( F u t u y a m a 1992;
animais apresentaram modificações morfológicas e fisiológicas marcantes quando passaram de uma situação t e r r e s t r e p a r a a q u á t i c a ou s e m i a q u á t i c a d u r a n t e sua e v o l u ç ã o . Entretanto, para algumas espécies a adaptação para a vida anfíbia pode envolver poucas modificações, devido a essa ' c o n d i ç ã o p r é v i a " ou pré a d a p t a ç ã o , q u e p o d e favorecer a permanência no ambiente aquático.
As espécies de ácaros oribatídeos das florestas de várzea e igapó foram
classificadas por FranklinzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA et al. (1997)
de acordo com sua resistência ao período de inundação: 1) sem resistência (não coletadas no período de inundação); 2) com média resistência (coletadas apenas até o nível máximo de inundação); 3) com alta resistência (coletadas até o final do período).
Por causa da alta capacidade de resistência à submersão, R. foveolatus foi considerada uma espécie anfíbia ( I r m l e r , 1981) ou s e m i a q u á t i c a (Messner et ai, 1992). Devido ao fato de q u e e s t a e s p é c i e p o s s u i u m a adaptação morfológica em forma de c e r o t e g u m e n t o para a vida semi aquática (Franklin et al., 1997), sua capacidade de sobrevivência sob a á g u a r e p r e s e n t a um i n t e r e s s a n t e fenômeno ecológico e fisiológico.
Neste trabalho, foram efetuadas comparações entre populações dessa espécie, provenientes de florestas s e c u n d á r i a s de t e r r a firme e de florestas inundáveis (várzea e igapó), para testar se a resistência de uma p o p u l a ç ã o à s u b m e r s ã o varia em função das condições (submersas e não submersas), do período de coleta ( a n t e s e d u r a n t e o p e r í o d o de
inundação dos rios) e de seu local de origem (florestas inundáveis e de terra firme). Adicionalmente, foi testada a resistência de outras 15 espécies de oribatídeos em condição submersa para avaliar as diferenças dos limites
de resistência desse grupo. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
M A T E R I A I S Ε M É T O D O S Áreas e período de coletas
Os animais foram coletados em 1992 e em 1996, em quatro áreas: 1) Floresta inundável de água preta ( i g a p ó ) no Rio T a r u m ã M i r i m ( 0 3 ° 0 1 ' S , 6 0 ° 1 0 ' W ) ; 2) F l o r e s t a secundária de terra firme (capoeira), vizinha ao ambiente do igapó; 3) Floresta inundável de água branca (várzea) no Rio Solimões (Ilha de Marchantaria, 03°15'S, 59°58'W); 4) Floresta secundária de terra firme (capoeira do Campus do INPA em M a n a u s , 03° 0 8 ' S , 60° 0 1 ' W ) . O p e r í o d o m é d i o da fase a q u á t i c a (baseado em 95 anos) no Rio Tarumã Mirim foi de 3,9 (±1,5) meses na posição topográfica da água a uma altura de 24 m acima do nível do mar; na Ilha de Marchantaria foi registrado um p e r í o d o médio de 5,6 ( ±1 , 6 ) meses, na posição topográfica da água a uma altura de 26,3 m acima do nível do mar (Adis et al., 1998).
junho e termina em agosto/setembro. A s c o l e t a s foram e f e t u a d a s em fevereiro na capoeira de terra firme, nas cercanias do igapó. Em abril fo ram efetuadas as coletas na várzea e na capoeira do Campus do INPA. Os animais foram mantidos em grupos com números variáveis de indivíduos, separados por recipientes de acordo com a condição de criação, período e local de coleta.
Em 1996, as c o l e t a s foram efetuadas apenas no mês de fevereiro nos quatro ambientes. Os indivíduos c o l e t a d o s foram m a n t i d o s
individualmente em placas separadas. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Coleta e condições experimentais dos animais
A coleta foi manual durante a fase terrestre e com auxílio de draga de E k m a n B i r g e d u r a n t e a fase aquática. A serapilheira foi recolhida de d i v e r s o s p o n t o s do solo e a c o n d i c i o n a d a d e n t r o de s a c o s plásticos que foram transportados em recipientes com isolamento térmico. Para extração dos animais foi utilizado
o método de Kempson (KempsonzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA et al., 1963), m o d i f i c a d o por A d i s
(1987). A temperatura do aparelho foi m a n t i d a a 3 2 ° C . O m a t e r i a l foi extraído diariamente, colocado em um recipiente contendo água destilada e o b s e r v a d o em m i c r o s c ó p i o e s t e r e o s c ó p i o para s e l e ç ã o dos espécimens vivos e sem mutilações.
As espécies foram identificadas através de chaves dicotômicas de Balogh ( 1 9 6 3 ; 1972) e Balogh & Balogh (1988; 1990).
G r u p o s de a n i m a i s de cada e s p é c i e ou a p e n a s um i n d i v í d u o
( q u a n d o se t r a t a v a de a n i m a i s coletados em 1996), foram mantidos em p l a c a s r e d o n d a s de a c r í l i c o (diâmetro de 5cm; altura de l,5cm), com uma camada de 0,5 cm de gesso para p r o p i c i a r s u b s t r a t o p a r a a l o c o m o ç ã o . A a b u n d â n c i a de R.
foveolatus é diferente em cada área,
com menor número de indivíduos nos ambientes de terra firme em relação ao igapó, razão pela qual não foi possível p a d r o n i z a r o m e s m o n ú m e r o de animais em todos os experimentos.
Para os experimentos em condições submersas, adicionouse água destilada até 0,5 cm acima da camada de gesso. Para os experimentos em condições não submersas, a placa de gesso foi umedecida com água destilada, permanecendo com uma umidade relativa em torno de 95%. Foi selecionado como alimento aveia em flocos e ração para peixe ("Tetramina"). Foram acrescentados pequenos pedaços de folhas recolhidas durante a coleta em campo, para servirem de substrato e abrigo aos animais.
O p e r í o d o de m o r t a l i d a d e (PM50) foi registrado quando morria cerca de metade da população da placa. O período médio de resistência (PME) foi calculado pelo total do período máximo de resistência de cada animal mantido individualmente.
fotorreceptoras, com as seguintes c o n d i ç õ e s de t e m p e r a t u r a e fotoperíodo: 27°C 12 horas com luz; 21°C 12 horas sem luz. De três em t r ê s d i a s , foram e f e t u a d o s os procedimentos de limpeza, troca de alimento e substituição de 1/3 da água p o r á g u a d e s t i l a d a . O c o n t r o l e consistiu na contagem de mortos e vi vos e observações do comportamento e habitat preferido. O estado geral do a n i m a l foi r e g i s t r a d o em duas categorias: "vivo" (com movimento sem toque de estilete) e "morto" (sem m o v i m e n t o a p ó s e s t í m u l o ) . Na maioria dos casos, nos animais mortos as pernas I e II ficavam esticadas para a frente. Os animais supostamente mortos, foram observados por mais
duas semanas. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Tratamento estatístico
As análises de variância foram efetuadas pelo teste de KruskalWallis (ANOVA não paramétrica; pacote e s t a t í s t i c o S i g m a S t a t 2.0). As c o m p a r a ç õ e s m ú l t i p l a s foram e f e t u a d a s pelo t e s t e de Dunn. O gráfico tipo " B o x P l o t " foi feito a t r a v é s do p a c o t e p s t a t í s t i c o SigmaPlot 4.0. Neste tipo de gráfico, cada coluna é plotada como uma caixa vertical, indicando o limite entre os percentis 25 e 75 dos dados e uma linha horizontal marca o valor do percentil 50. Os percentis 5 e 95 são as barras de erro. No texto, os demais valores que acompanham as médias são os respectivos desvios padrões.
R E S U L T A D O S
Populações dezyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA R. foveolatus de
1992
A resistência dos oribatídeos à submersão, variou em função da fase do ciclo de inundação e o ambiente de origem.
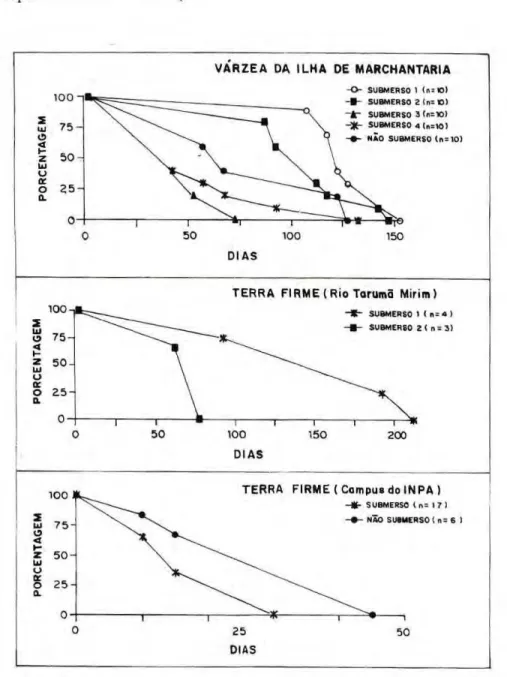
No igapó (Fig. 1), os animais sob condição não submersa foram mais resistentes em fevereiro. Em abril as curvas de mortalidade entre os animais em c o n d i ç õ e s s u b m e r s a e n ã o submersa foram muito p a r e c i d a s . Contudo, em julho, os animais não submersos foram mais resistentes (acima de 150 dias), em comparação a uma média de resistência de cerca de 80 dias dos animais submersos. Em abril, o período médio de mortalidade (PM50) dos animais sob condição de submersão (acima de 100 dias) foi maior que em fevereiro (75 dias) e julho (60 dias).
N a v á r z e a da Ilha de Marchantaria (coletas efetuadas em abril, na serapilheira submersa), o PM50 da população não submersa foi de cerca de 70 dias (Fig. 2). O PM50 d o s e x p e r i m e n t o s 1, 2 e 4 (em condições submersas) foi superior a
100 d i a s . O m e n o r p e r í o d o de resistência (cerca de 75 dias) foi registrado no experimento 3.
Na floresta secundária (capoeira) de terra firme no Rio Tarumã Mirim (coletas em fevereiro de 1992), o PM50 dos indivíduos sob submersão foi cerca de 60 dias (Fig. 2).
Na floresta secundária (capoeira) de terra firme no Campus do INPA (coletas em abril de 1992), o PM50 foi registrado com cerca de 15 dias de submersão.
Foram observadas diferenças marcantes nos períodos de sobrevivência das criações provenientes
das c o l e t a s e f e t u a d a s em f e v e r e i r o de 1996, m a n t i d o s
em p l a c a s i n d i v i d u a l i z a d a s ( F i g . 3 ) . O s t e m p o s m é d i o s de sobrevivência dos animais das florestas
inundáveis da várzea (PME de 150,5±5 dias) e do igapó (PME de
D I A S
A B R I L zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
O 5 0
D I A S
JULHO zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
D I AS
Figura 1. Sobrevivência dezyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Rostrozetes foveolatus ao longo do tempo, em condições submersa
110,8±60 dias), foram maiores (P< 0 , 0 0 1 ; Η = 62,3) que os provenientes das florestas de terra firme da capoeira do Tarumã Mirim (PME de
49 ±26 dias) e da capoeira do INPA (PME de 47±36 dias) (Tab. 1).
Foi detectada a presença de três
exemplares submersos dezyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA R. foveolatus
V Á R Z EA DA I LH A DE M ARCHAN TARI A zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
- O - SUBMERSO 1 lnzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA-n)
D I A S
T ERRA FI RM E ( Ri o Tarumã M irim) zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
O 50 10 0 150
D I A S
DIAS
no estágio larval. Uma das larvas deu origem a um adulto pouco esclerotizado,
que morreu após 15 dias. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Populações de outras espécies de oribatídeos de 1992.
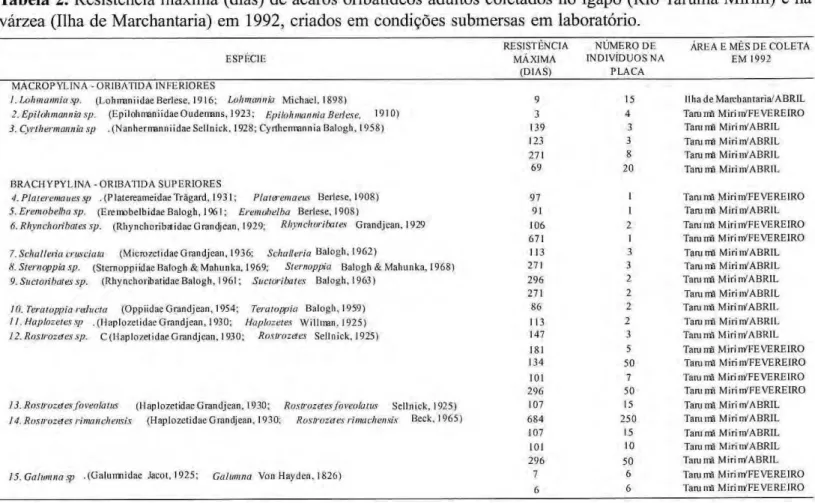
Foi também medida a resistência à
submersão em outra espécie dezyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Rostrozetes
(denominada de Rostrozetes sp. C) e em
Rhynchoribates sp., Rostrozetes rimachensis, e Suctoribates sp., Epilohmannia sp., Plateremaeus sp., Cyrthermannia sp., Eremobelba sp., Haplozetes sp., Sternoppia sp., Schalleria crusciata e Teratoppia sp. (Tab. 2).
D I S C U S S Ã O
No igapó, apesar da exceção de abril, os animais tendem a resistir melhor quando não estão submersos, o que significa que a submersão é um fator de e s t r e s s e . O m e s m o n ã o ocorreu na várzea, o que pode indicar q u e esta p o p u l a ç ã o está m e l h o r adaptada ao estresse da submersão. C o n t u d o , foi d e t e c t a d a u m a considerável resistência a submersão na população de R. foveolatus coletada na t e r r a firme ( c a p o e i r a do R i o Tarumã Mirim), próxima ao igapó,
< zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
ú
300
2 5 0
2 2 0
1 5 0
1 0 0
-5 0 J
0
Tabela 1.zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Teste de comparações múltiplas (Método de Dunn) entre as populações dezyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA R. foveolatus
provenientes das florestas inundáveis (várzea e igapó) e as florestas secundárias de terrafirme
(Tarumã Mirim e INPA). zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Comparações n1 n2 Diferença
de Rank Q P<0,05
Várzea X Capoeira INPA 8 35 81,657 4,092 Sim
Várzea X Capoeira Tarumã Mirim 8 83 74,416 3,948 S i m
Várzea X Igapó Tarumã Mirim 8 50 14,670 0,757 Não
Igapó Tarumã Mirim X Capoeira INPA 50 35 66,987 5,969 S i m
Igapó Tarumã Mirim X Capoeira Tarumã Mirim 50 83 59,746 6,554 Sim
Capoeira Tarumã Mirim X Capoeira INPA 83 35 7,241 0,706 Não
c o m p a r á v e l à r e g i s t r a d a p a r a a s populações das áreas inundáveis e maior do que a resistência registrada e m t e r r a firme longe do i g a p ó , novamente indicando diferença entre p o p u l a ç ã o p a r a a r e s i s t ê n c i a a submersão.
Έπ ι contrapartida, nas criações efetuadas em 1996, envolvendo maior n ú m e r o de animais e em placas individualizadas, foi verificado que os animais coletados nas áreas inundadas possuem um período de sobrevivência significativamente maior em relação à área de terra firme. Esse resultado não apoia o fato de termos registrado na capoeira de terra firme próxima ao igapó uma resistência comparável aos de áreas inundáveis. Contudo, alguns indivíduos dentro de uma população tem a capacidade de sobreviver maior ou menor tempo do que os outros em condições submersas. O tempo de isolamento e o período submetido à inundação, pode levar à seleção natural dos mais resistentes, o que nos leva a concluir que registramos três situações: a) várzea com população mais resistente, b) igapó, com uma população
intermediária e c) terrafirme longe do igapó (capoeira do INPA), com uma população menos resistente.
Nossos resultados obtidos para
R. foveolatus, no igapó do Rio Tarumã
Mirim, corroboram os de Beck (1972), que r e g i s t r o u a c a p a c i d a d e d e sobrevivência dessa espécie, na forma a d u l t a , d u r a n t e t o d o o c i c l o d e inundação das florestas inundáveis, que varia de 46 meses anualmente.
M e s s n e r et al. ( 1 9 9 2 ) verificaram que R. foveolatus pode r e s i s t i r à a u s ê n c i a d e o x i g ê n i o , a p a r e n t e m e n t e em d o r m ê n c i a , voltando ao estágio ativo em água normal. Os autores supõem que a a t i v i d a d e da e s p é c i e em á r e a s s u b m e r s a s a n u a l m e n t e d u r a n t e 6 meses, depende da concentração do oxigênio dissolvido na água. Eles também confirmaram a existência de um c e r o t e g u m e n t o em a l g u n s O r i b a t i d a c o l e t a d o s em á r e a s inundáveis. O cerotegumento de R.
foveolatus e de Eremobelba sp. foi
previamente demonstrado por Franklin
etal. (1997).
Tabela 2.zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Resistência m á x i m a (dias) d e ácaros oribatídeos adultos coletados n o igapó (Rio Tarumã Mirim) e na várzea (Ilha de Marchantaria) e m 1992, criados em condições submersas e m laboratório.
RESISTÊNCIA NÚMERO DF. ÁREA Ε M Ê S DE C O L E T A
ESPÉCIE MÁXIMA INDIVÍDUOS NA EM 1992
(DIAS) PLACA M A C R O P Y L I N A ORIBATIDA INFERIORES zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
1. l.ohmannia sp. (l.ohmaniidae Berlese, 1916; Lohmwvüa Michael, 1898) 9 15 Ilha de Marchantaria/ABRIL
2.Epilohmannia sp. (Hpilohmaniidae Oudemans, 1 9 2 3 ; Epilohmannia Bertese, 1910) 3 4 Tara ma Miri nVFE VEREIRO
3. Cyr therm an η ia sp .(Nanhermanniidae Sellnick, 1928; Cyrthennannia Balogh, 1958) 139 3 Tarumã Mirim'ABRIL 123 3 Tarumã MirinV ABRIL 271 8 Tarumã MirinVABRIL
69 20 Tarumã MirinVABRIL BRACHYPYLINA ORIBATIDA S U P E R I O R E S
4. Piateremauessp . (PlatereameidaeTrâgard, 1 9 3 1 ; Plateremaeus Berlese, 1908) 97 1 Tarumã MirinVFEVEREIRO
S.Eremobelba sp. (Eremobelbidae Balogh, 1%I; Eremobelba Berlese. 1908) 91 1 Tarumã MirinVABRIL
Ó.Rhynchoribatessp. (RhynchoribatidaeGrandjean, 1929; Rhynchàibates Grandjean, 1929 106 2 Tarumã Miri nVFE VEREIRO 671 1 Tarumã MirinVFEVEREIRO
7.Schalleiiacmsciata (MicrozelidaeGrandjean, 1936; Schulleria Balogh, 1962) 113 3 Tani mâ MirinVABRIL
8. Slernoppia sp. (Sternoppiidae Balogh & M a h u n k a , 1969; Sternoppia Balogh & Mahunka, 1968) 271 3 Tarumã MirinVABRIL
9. Suctoiibatessp. (Rhynchoribatidae Balogh, 1961; Suctorilxttes Balogh, 1963) 2 9 6 2 Tarumã MirinVABRIL 271 2 Tarumã MirinVABRIL
lO.Teraioppia ralucta (Oppiidae Grandjean, 1954; Teratoppia Balogh. 1959) 86 2 Tarumã MirinVABRIL
11. Haplozetessp . (Haplo/.elidae Grandjean, 1930; Haplozetes Willman, 1925) 113 2 Tammã MirinVABRIL
12. Rostrozetes sp. C ( I l a p l o z e l i d a e Grandjean, 1930; Rostrozetes Sellnick, 1925) 147 3 Tarumã MirinVABRIL 181 5 Tarumã MirinVFEVEREIRO 134 50 Tammã MirinVFEVEREIRO
101 7 Tarumã MirinVFEVEREIRO 2 9 6 50 Tammã MirinVFEVEREIRO
13. Rostrozetes foveolatus (Haplozetidae Grandjean, 1930; Rostrozetes foveolatus Sellnick, 1925) 107 15 Tarumã MirinVABRIL
14. Rostrozetes rimanchensis (Haplozetidae Grandjean. 1930; Rostrozetes rimachensís Beck, 1965) 6 8 4 2 5 0 Tammã MirinVABRIL 107 15 Tammã MirinVABRIL 101 10 Tammã MirinVABRIL 2 9 6 5 0 Tammã MirinVABRIL
IS.Galamna sp .(Galumnidae Jacol, 1925; Galumna Von Hayden, 1826) 7 6 Tani nfi Miri nVFE VEREIRO
IS.Galamna sp .(Galumnidae Jacol, 1925; Galumna Von Hayden, 1826)
que ocorrem em ambientes sujeitos à inundação possuem uma estrutura tipo plastrão para permitir a respiração traqueal. Este plastrão é suportado por um cerotegumento modificado, que são camadas epicuticulares, também o b s e r v a d a s em o r i b a t í d e o s que
ocorrem no solo (AlbertizyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA et al, 1981),
em á g u a doce ( N e w e l l , 1945; Grandjean, 1948; Crowe & Magnus,
1973; K r a n t z & Baker, 1982; Fernandez, 1984) e no litoral marinho (Alberti et al, 1981).
Nas espécies R. foveolatus, R.
rimachensis Beck, 1965, Rostrozetes
sp. C, Haplozetes sp. A, Eremobelba sp. e Rhynchoribates sp. A, podese inferir que a resistência à submersão p o d e ser, em p a r t e , a t r i b u í d a ao plastrão. Os organismos do solo estão e n v o l v i d o s em um a m b i e n t e que impõe três requerimentos principais, (1) se m o v e r em um a m b i e n t e c o m p a c t o com p o r o s i d a d e f r o u x a m e n t e c o n e c t a d a , (2) se a l i m e n t a r de r e c u r s o s com baixa qualidade e (3) se adaptar a ocasionais secas e inundações dos espaços entre os poros (Lavelle, 1997). Portanto, mesmo em terra firme, a espécie R.
foveolatus pode ser freqüentemente
submetida a uma situação semelhante à inundação em filmes de água sobre as folhas mortas e no solo, durante e após as fortes chuvas. A presença do plastrão pode permitir que o animal continue respirando normalmente com o auxílio de uma película de ar aderida ao c o r p o , e assim c o n t i n u a r se alimentando. Isso poderia ser uma pré adaptação (Futuyama, 1992; Schaefer,
1992) para uma s u b m e r s ã o mais prolongada nas florestas inundáveis. Esta suposição reforça aquelas de Beck (1972), de que R. foveolatus pode possuir uma "condição prévia" para sobreviver no ambiente inundado, já que as espécies de terra firme,
especialmente do gênero Rostrozetes, apresentam a mesma estrutura. Os resultados obtidos no presente estudo sugerem que os i n d i v í d u o s provenientes da terra firme estão pré adaptados às condições de submersão nas florestas inundáveis (igapó e v á r z e a ) . Um e x e m p l o d i s s o é a capacidade que R. foveolatus possui para sobreviver, se alimentar (Messner
et al., 1992) e até m e s m o se
reproduzir, embaixo da água. Segundo Beck (1972), a partenogênese poderia ser um prérequisito necessário para que as espécies de terra firme invadam as áreas inundadas. Por outro lado, fica a ser estudado se a população do igapó na realidade representa uma outra (sub) espécie em comparação à da terra firme que não p o d e ser diferenciada p e l a s características m o r f o l ó g i c a s m a s s o m e n t e p e l a s características genéticas (compare Wolf & Adis, 1992).
período de inundação, possivelmente
em dormência, poiszyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Epilohmannia sp.
e Lohmannia sp. foram consideradas espécies de média (acima de 72 dias) e de alta ( a c i m a de 183 dias) r e s i s t ê n c i a à s u b m e r s ã o , r e s p e c t i v a m e n t e (Franklin, 1994; Franklin et al., 1997). De acordo com Tamm ( 1 9 8 4 ) , a p e r m a n ê n c i a em estágio de ovo seria a única estratégia de sobrevivência de invertebrados terrestres em regiões inundáveis, pois o autor acreditava que os estágios desenvolvidos deveriam ser capazes de s o b r e v i v e r por a p e n a s p o u c a s s e m a n a s . E n t r e t a n t o , S o m m e & ConradiLasen (1977), mantiveram os oribatídeos Carabodes labirynthicus M i c h a e l , 1879 e Calyptozestes
sarekensis T r á g a r d h , 1910, em
condições de anoxia por 96 dias. Du rante a anaerobiose houve acumulo de lactato, mas os autores argumentaram que essa não seria a única condição para sobrevivência em ausência de oxigênio. De fato, as estratégias de sobrevivência j á e n c o n t r a d a s n o s artrópodos terrestres do igapó e da várzea na Amazonia Central são de vários tipos (Adis 1997a; b).
Já f o r a m c o n f i r m a d a s e s t r u t u r a s t i p o p l a s t r ã o em R.
foveolatus, R. rimachensis, Rostrozetes sp., Haplozetes sp., Eremobelba sp. e Rhynchoribates
sp, em microscopia eletrônica de varredura (SEM/Scanning Electron Mi croscopy) (Messner et. al 1992), significando que são estruturas comuns em algumas espécies, e que podem justificar, em parte, a resistência desses
animais às condições submersas (Adis & Messner 1997).
Um ponto em comum entre os g ê n e r o s de m a i o r r e s i s t ê n c i a à submersão (>60 dias) listados na Tabela 2, é que todos são considerados grupos basilares na linha filogenética dos O r i b a t i d a ( W o a s , 1990).
Haplozetes, a p e s a r de não estar
classificado como gênero basilar, é c o n s i d e r a d o p r i m i t i v o entre os periféricos (Woas, S. comunicação p e s s o a l ) . O g ê n e r o Galumna, c l a s s i f i c a d o c o m o um dos mais recentes na linha de evolução dos Oribatida, possui pouca ou nenhuma resistência à submersão e não foi encontrado na serapilheira submersa, podendo morrer ou "migrar" para os troncos ou superfícies não submersas quando a água invade o ambiente (Franklin, 1994; Franklin et al., 1997).
A presença do plastrão pode c o n t r i b u i r para a r e s i s t ê n c i a de oribatídeos no início da submersão. Mas é possível que numa submersão prolongada, sob condições de pouco o x i g ê n i o , e s s e s a n i m a i s p o s s a m também mudar para uma respiração
anaeróbica. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
A G R A D E C I M E N T O S
Agradecemos ao Prof. Dr. S. Woas do Museu de História Natural de K a r l s r u h e p e l a s d i s c u s s õ e s e comentários. A contribuição dos
refer-ees indicados pela revista também foi
com microscopia eletrônica. Ao CNPq pela bolsa de Iniciação Científica c o n c e d i d a ao s e g u n d o autor. Ao Edilson de Araújo Silva, a Lúcia Paz e ao Cláudio Sena (Inpa) pelo apoio técnico. Ao Inpa e ao Convênio Inpa/ M a x P l a n c k , n o s s o s s i n c e r o s
agradecimentos pelo apoio logístico. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Bibliografia citada
Adis, J. 1987. Extraction of arthropods from N e o t r o p i c a l soils with a m o d i f i e d
Kempson apparatus.zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA J. Trop. Ecol., 3:
131138.
1992. Ueberlebensstrategie terrestrischer Invertebraten in U e b e r s c h w e m m u n g s w a e l d e r n Z e n t r a l a m a z o n i e n s . Ver.
Naturwiss. Ver. Hamburg (NF), 33: 21
114.
1997a. Terrestrial invertebrates: Survival strategies, group spectrum, dominance and activity patterns. In: Junk, W.J. (ed.): The
Central Amazon Floodplain. Ecology of a pulsing system. Ecological Studies 126.
Springer, Berlin, pp. 299317.
1997b. Estratégias de sobrevivência de i n v e r t e b r a d o s t e r r e s t r e s em florestas inundáveis da Amazônia Central: uma resposta à inundação de longo período.
Acta Amazônica, 27(1): 4354.
Adis, J.; Messner, B. 1997. Adaptations to life under water: Tiger beetles and millipedes.
In: Junk, W.J. ( e d . ) : The Central Amazon floodplain. Ecology of a pulsing system. Ecological Studies 126.
Springer, Berlin, pp. 318330.
Adis, J.; Messner, B.; Platnick,N. 1999. Mor phological studies and vertical distribution in the soil indicate facultative plastron res piration in Cryptocellus adisi (Arachnida, Ricinulci) from Central Amazonia. Stud.
Neotrop. Fauna & Environm., 34(1): 19.
Adis, J.; Paarmann, W.; Amorim, M.A.; Arndt, E.; F o n s e c a , C.R.V. da. 1 9 9 8 . On occurence, habitat specifity and natural history of adult tiger beetles (Coleoptera: Carabidaè: Cicindelinae) near Manaus,
Central Amazonia, and key to the larvae of tiger beetle genera. Acta Amazônica, 28(3): 247272.
Alberti, G.; Storch, H.; Renner, H. 1981. Ueber den f e i n s t r u k t u r e l l e n Aufbau d e r Milbcncuticula (Acari: Arachnida). Zool.
Jb.Anat., 105: 183236.
Balogh, J. 1963. Identification keys of holartic mites (Acari) families and genera. Acta
Zool. Hung., 9: 160.
1972. The oribalid genera of the world. A k a d e m i a i K i a d o , B u d a p e s t .
188 pp. + 71 pi.
Balogh, J..; Balogh, P. 1988. Oribatid mites of
the Neotropical Region. I. Elsevier,
Amsterdam. 335 pp.
1990. Oribatid mites of the Neotropical
Region. 11. Elsevier, Amsterdam. 333 pp.
B e c k , L. 1968. Z u m j a h r e s p e r i o d i s c h e n Massenwechsel zweier Oribatidenarten ( A c a r i ) im n e o t r o p i s c h e n Ueberschwemmungswald. Verh. Dtsch
Zool. Ges.,: 535540.
1971. Bodenzoologische Gliederung und C h a r a k t e r i s i e r u n g d e s a m a z o n i s c h e n Rcgenwaldes. Amazoniana, 3(1):69132. 1972. Der Einfiuss der jahresperiodischen Uebcrflutungen auf den Massenwechsel d e r B o d e n a r t h r o p o d e n im zcntralamazonischcn Regenwaldgebiet.
Pedobiologia, 12: 133148.
1976. Zum Massenwechsel der Makro A r t h r o p o d e n f a u n a d e s B o d e n s in Ueberschwemmungswàldern des zentralen Amazonasgebietes. Amazoniana, 6(1): 1 20.
Crowe, J. H.; Magnus, K. A. 1973. Studies on acarine cuticles II. Plastron respiration in a water mite. Comp. Biochem. Physiol., 49A: 301310.
F e r n a n d e z , N. 1 9 8 4 . C o n t r i b u t i o n à la connaisance de la familic Hydrozetidac.I.
Hydrozeles (Argentinobates) ringuelen
n o v . s u b g e n . , n o v . s p . Acarologia, 25: 307317.
Franklin, E. 1994. Ecologia de oribatídeos
da Amazônia Central.zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA Tese de doutorado, Inpa/Universidade do Amazonas, Manaus.
266 pp.
Franklin, E.N.; Adis, J.; Woas, S. 1997. The oribatid mites. In: Junk, W.J. (ed.): The
Central Amazon floodplain. Ecology of a pulsing system. Ecological Studies 126.
Springer, Berlin, pp. 331349.
Futuyama, D.J. 1992. Biologia Evolutiva. Ribeirão Preto, Sociedade Brasileira de Genética/CNPq, 646 pp.
G r a n d j e a n , F. 1 9 4 8 . Sur les H y d r o z e t e s (Acariens) de P'Europe occidentale. Buli.
Mus. Nat. Hist, natur., (2), 20: 328325.
lrmler, U. 1 9 8 1 . Ueberlebensstrategie von T i e r e n im s a i s o n a l u e b e r f l u t e t e n amazonischen Ueberschwemmungswald.
Zool. Anz., 206: 2638.
Junk, W. J; Bailey, P. Β. ; Sparks, R. Ε . 1989. The flood pulse concept in riverflood plain systems. In: Dodge, D.P. (ed.). Pro ceedings of the International Large River Symposium (LARS). Can. Spec. Publ.
Fish. Sci., 106: 110127.
Kempson, D.; Lloyd, M.; Gelardi, R. 1963. A n e w e x t r a c t o r for w o o d l a n d litter.
Pedobiologia, 3: 121.
Krantz, G.W.; Baker, G.T. 1982. Observations on the plastron mechanism of Hydrozetes s p . ( A c a r i : O r i b a t i d a : H i d r o z e t i d a e ) .
Acarologia, 23: 273277.
Lavelle, P. 1997. Faunal activities and soil pro cesses: adaptative strategies theat deter mine ecosystem function. Advances in
Ecological Research, 27: 93132.
Luxton, M. 1975. Studies on the oribatid mites of a Danish beech forest II. Biomass, calo rimetry and respirometry. Pedobiologia,
15: 161200.
Messner, B.; Adis, J.; Ribeiro, E.F. 1992. Eine v e r g l e i c h e n d e U n t e r s u c h u n g u e b e r Plastronstrukturen bei Milben (Acari).
Dtsch. ent. Z , N.F., 39(13): 159176.
Newell, I.M. 1945. Hydrozetes Berlese (Acari: Oribatoidea): the occurrence of the genus in North America, and the phenomenon of levitation. Trans. Conneticut Acad. Art.
Sci., 6:253275.
S c h a e f e r , M. 1992. Woerterbuecher der
Biologic Oekologie. 3 ed. Fischer, Jena.
433 pp.
S c h u s t e r , R. 1 9 6 5 . Die O e k o l o g i e d e r t e r r e s t r i s c h e n K l e i n f a u n a d e s Meeresstrandes. Verh. Dtsch. Zool. Ges.,: 492521.
1966. Hornmilben (Oribatei) als Bewohner d e s m a r i n e n L i t o r a i s . Veroeff. Inst.
Meeresf. Bremerhaven, II: 319328.
1978. Soil mites in the marine environ ment. Rec. Advan. Acarol., I: 593602. S o m m e , L.; C o n r a d i L a r s e n , E.M. 1977.
Anaerobiosis in overwintering collembo lans and oribatid mites from windswept mountain ridges. Oikos, 29:127132. Tamm, J.C. 1984. Surviving long submergence
in the egg stage a successful strategy of terrestrial arthropods living on flood plains (Collembola, Acari, Diptera). Oecologia, 6 1 : 417419.
W e i g m a n n , G. 1 9 7 3 . Z u r O e k o l o g i e d e r C o l l e m b o l e n und O r i b a t i d e n im Grenzbereich LandMeer (Collembola, Insecta O r i b a t e i , A c a r i ) . Z. wiss. Zoo/., 186 (3/4): 295391.
W o a s , S. 1 9 9 0 . Die p h y l o g e n e t i s c h e n E n t w i c k l u n g s l i n i e n d e r H o e h e r c n Oribatiden (Acari). I. Zur Monophylie der Poronota Grandjean, 1953. Andrias, 7: 9 1 168.
Wolf, H.G.; Adis, J. 1992. Genetic differentia tion b e t w e e n p o p u l a t i o n s o f
Neomachilellus scandens Meinertellidae,
A r c h a e o g n a t h a , I n s e c t a ) i n h a b i t i n g neighbouring forests in Central Amazonia.
Verh. naturwiss. Ver. Hamburg (NF), 233:
513. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA