Beta2 oscillations (23
–
30 Hz) in the mouse hippocampus
during novel object recognition
Arthur S. C. Francßa,1,2George C. do Nascimento,3 Vıtor Lopes-dos-Santos,1Larissa Muratori,1Sidarta Ribeiro,1 Bruno Lob~ao-Soares4andAdriano B. L. Tort1
1Brain Institute, Federal University of Rio Grande do Norte, Natal, RN 59056-450, Brazil 2Edmond and Lily Safra International Institute of Neuroscience of Natal, Natal, Brazil
3Department of Biomedical Engineering, Federal University of Rio Grande do Norte, Natal, Brazil 4Department of Biophysics and Pharmacology, Federal University of Rio Grande do Norte, Natal, Brazil
Keywords: brain rhythms,in vivoelectrophysiology, local field potential, novelty, spectral analysis
Abstract
The oscillatory activity of hippocampal neuronal networks is believed to play a role in memory acquisition and consolidation. Particu-lar focus has been given to characterising theta (4–12 Hz), gamma (40–100 Hz) and ripple (150–250 Hz) oscillations. Beyond these well-described network states, few studies have investigated hippocampal beta2 (23–30 Hz) activityin vivoand its link to behaviour. A previous sudy showed that the exploration of novel environments may lead to the appearance of beta2 oscillations in the mouse hippocampus. In the present study we characterised hippocampal beta2 oscillations in mice during an object recognition task. We found prominent bursts of beta2 oscillations in the beginning of novel exploration sessions (four new objects), which could be readily observed by spectral analysis and visual inspection of local field potentials. Beta2 modulated hippocampal but not neocortical neu-rons and its power decreased along the session. We also found increased beta2 power in the beginning of a second exploration ses-sion performed 24 h later in a slightly modified environment (two new, two familiar objects), but to a lesser extent than in the first session. However, the increase in beta2 power in the second exploration session became similar to the first session when we phar-macologically impaired object recognition in a new set of experiments performed 1 week later. Our results suggest that hippocampal beta2 activity is associated with a dynamic network state tuned for novelty detection and which may allow new learning to occur.
Introduction
The relation between neuronal oscillations and behaviour has been the focus of many studies over recent decades. Although also detected at the single-neuron level (Kamondiet al., 1998), neuronal oscillations are typically studied at the mesoscopic scale of local field potentials (LFPs; Buzsaki & Draguhn, 2004; Buzsaki, 2006), which represent the activity of an ensemble of nearby neurons (Buz-saki, 2004; Linden et al., 2011; Buzsaki et al., 2012; Reimann et al., 2013). In the hippocampus, many studies have focused on the role of theta (4–12 Hz; Vanderwolf, 1969; Winson, 1978; Buzsaki, 2002), gamma (30–100 Hz; Csicsvari et al., 2003; Montgomery & Buzsaki, 2007; Colgin et al., 2009), high-frequency (110–160 Hz; Scheffer-Teixeira et al., 2012; Tort et al., 2013) oscillations, and sharp-wave associated ripples (150–250 Hz; Buzsaki et al., 1992; Girardeau et al., 2009; Ego-Stengel & Wilson, 2010), in different behaviours and cognitive states. In addition, Berke et al. (2008) have called attention to prominent bursts of 23- to 30-Hz oscilla-tions that appear in CA1 and CA3 when mice explore novel envi-ronments. This rhythm, referred to as beta2 oscillations, has been shown to modulate hippocampal neurons and to depend on NMDA
transmission (Berke et al., 2008), which is known to be important for rapid learning (Nakazawa et al., 2003). Grossberg (2009) sug-gested that beta2 oscillations provide a transient plasticity signal able to solve the stability–plasticity dilemma, by which the ability of a neuronal network to learn rapidly must be compatible with sta-ble memory representations without catastrophic forgetting (Gross-berg, 1980, 1999, 2009).
The beta2 oscillations reported in Berke et al. (2008) are visible in unfiltered LFPs and, actually, may have amplitude similar to that of theta oscillations. This is by itself a remarkablefinding consider-ing that hippocampal oscillations constitute a major focus of research: why have such large-amplitude oscillations not been described before? And, to the best of our knowledge, why has there not been a second report showing similar hippocampal oscillations? Would the putative appearance of beta2 only during specific behav-iours account for the fact that these oscillations remained undetected for so long? If so, what kinds of behaviours elicit beta2 oscillations? Are they specific to novel spatial experiences? Can amnesic inter-ventions modulate beta2 appearance in the hippocampus?
In the present study we sought to confirm and extend the original findings of Berke et al. (2008). By recording from freely moving mice, we found that the exploration of novel objects leads to the transient appearance of prominent beta2 oscillations; no such activ-ity occurred when animals were recorded in the home cage before Correspondence: Adriano B. L. Tort, as above.
E-mail: [email protected]
and after the exploration session. Furthermore, beta2 power depended on the numbert of novel objects present in the arena, and pharmacologically blocking object memory consolidation was asso-ciated with higher beta2 power than when animals showed normal recognition of familiar objects. These results provide support to the proposal that hippocampal beta2 oscillations could play a role in novelty detection and may constitute a signal for new learning to occur (Berkeet al., 2008; Grossberg, 2009).
Materials and Methods
Animals
Five C57BL/6 mice were used in this study. Animals were housed individually, with a 12-h cycle of light and dark (lights on at 06.00 h), and no food or water restriction. All procedures followed guidelines of the National Institutes of Health and were approved by the Edmond and Lily Safra International Institute of Neuroscience of Natal Ethics Committee (protocol number 08/2010).
Task design
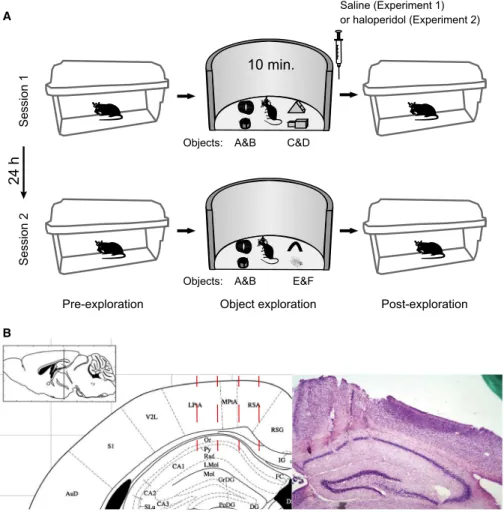
Animals were submitted to a novel object recognition task. Each experiment (saline or haloperidol protocol; see below) consisted of two sessions 24 h apart with three periods each: pre-exploration
(home cage), object exploration (open field), and post-exploration (home cage; Fig. 1A).
In the first session, four objects (A, B, C and D) were presented for 10 min during the exploration period in a circular arena (50 cm diameter and 30 cm high). Immediately after, animals were sub-jected to intraperitoneal (i.p.) injection of saline (saline protocol) before returning to the home cage. Twenty-four hours later (i.e., in the second session), animals explored two familiar objects (A and B from thefirst session) and two novel objects (E and F).
One week after thefirst experiment, four of the animals were sub-jected to a similar behavioural task but with different objects and with injection of haloperidol (0.3 mg/kg) immediately after thefirst exploration session (haloperidol protocol), which impairs object rec-ognition in the second session (Lob~ao-Soares et al., 2009). As in experiment 1, objects are also referred to as A, B, C, D, E and F for computing the novelty index (see below). Animals were video-recorded throughout the experiments.
Surgery
Animals were implanted with multielectrode arrays (dimensions 0.992.1 mm) composed of 50-lm-diameter tungsten wires, 1.5 mm in length; four electrodes targeted the primary motor cortex (M1), four electrodes targeted the somatosensory cortex (S1) andfive electrodes targeted the CA1 region of the dorsal hippocampus. The arrays were
Pre-exploration
24 h
Session 1
Session 2
10 min.
Saline (Experiment 1) or haloperidol (Experiment 2)
Object exploration Post-exploration
Objects:
Objects: A&B
A&B A
B
C&D
E&F
implanted through a rectangular opening in the skull (coordinates from begma, 0.55 and 1.65 mm mediolateral and 0.0 and 2.2 mm antero-posterior). Electrode placement was confirmed by inspecting histolog-ical brain sections stained with Cresyl Violet (Fig. 1B).
Recordings
Electrophysiological recordings were made using a multichannel acquisition processor (Plexon Inc., Dallas, TX, USA). LFPs were preamplified (10009),filtered (1–500 Hz) and sampled at 1000 Hz. A high-impedance homemade headstage and a PBX preamplifier (Plexon Inc., Dallas) model were used. Spikes from multiunit activ-ity were obtained by amplifying (10009),filtering at 500–8000 Hz, and sampling at 40 kHz.
Data analysis
Data analyses were performed using built-in and custom-written rou-tines in Matlab (MathWorks, Natick, MA, USA). The raw signal was first visually inspected and electrodes with prominent noise were discarded from further analysis.
Filter settings
Filtering was achieved with the eegfilt.m routine from the EEGLAB Matlab toolbox (http://sccn.ucsd.edu/eeglab/). This routine uses a linear finite response filter and applies the filter forward and then backwards to eliminate phase distortions. The instantaneous phase and amplitude of a filtered signal were obtained from the analytical representation of the signal using the hilbert.m routine from the Sig-nal Processing Toolbox. Theta and beta2 oscillations were obtained by bandpassfiltering at 4–12 Hz and 23–30 Hz, respectively.
Spectral analyses
The power spectrum density was computed by the pwelch.m routine from the Signal Processing Toolbox (50% overlapping
Hamming window of 4 s). Band power was defined as the mean over power values in the analysed frequency range. The time– fre-quency representation shown in Fig. 3A was obtained by the spec-trogram.m routine from the Signal Processing Toolbox. The latency to peak beta2 activity was estimated from the mean ampli-tude of beta2-filtered signals in 10-s sliding windows with 50% overlap. To detect beta2 bursts, for each electrode we first com-puted the mean amplitude of the beta2-filtered signal. A burst event was defined as occurring when the instantaneous amplitude was > 2 SD from the mean. Burst duration was defined as the per-iod > 1 SD from the mean. For the burst analysis shown in Fig. 7, each animal contributed with a single LFP, selected as the hippo-campal electrode with the highest number of beta2 bursts (elec-trodes in any individual animal had similar numbers of beta2 bursts; not shown).
Behavioural analysis
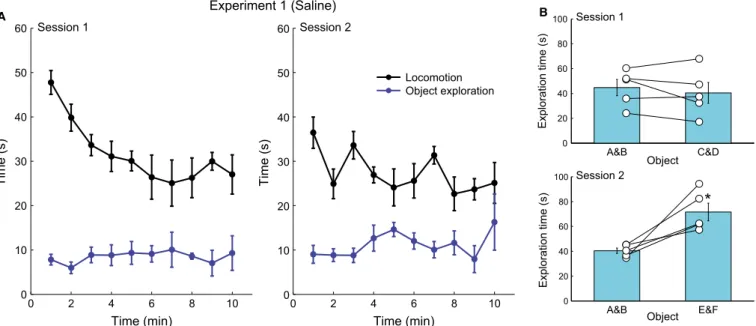
Object exploration time was considered to be the time animals spent with the whiskers or both front paws in contact with objects. Mice have a natural tendency to explore novel objects (Hughes, 1997, 2007; Dere et al., 2007; Heyser & Chemero, 2012), and spend roughly the same amount of time exploring each of the four objects if they are all novel. On the other hand, if only two objects are novel, animals spend less time exploring the familiar objects, giving rise to unequal exploration time between novel and familiar objects (Dere et al., 2007). Therefore, we defined the novelty index as the ratio of the time spent exploring the two objects that were the same in sessions 1 and 2 (objects A and B) divided by the time exploring the other two objects (objects C and D in session 1 and E and F in session 2). Notice that, under normal conditions, the novelty index should be lower in session 2 than 1 because objects A and B become familiar and are less explored.
The correlation between beta2 and the time spent in locomotion or exploring objects was obtained byfirst computing the mean (over electrodes) normalised beta2 power per animal, and then by pooling
A&B C&D
0 20 40 60 80 100
A&B E&F 0
20 40 60 80 100
0 2 4 6 8 10
0 10 20 30 40 50 60
Time (min)
Session 1 Session 1
*
Session 2 Session 2
Time (s)
Exploration time (s)
Exploration time (s)
Locomotion Object exploration
0 2 4 6 8 10
0 10 20 30 40 50 60
Object Time (min)
Object
Time (s)
A Experiment 1 (Saline) B
values among animals. We used 1-min non-overlapping windows; beta2 power in each window was normalised to the power value in the last window.
Results
Transient hippocampal beta2 oscillations appear during the exploration of novel objects
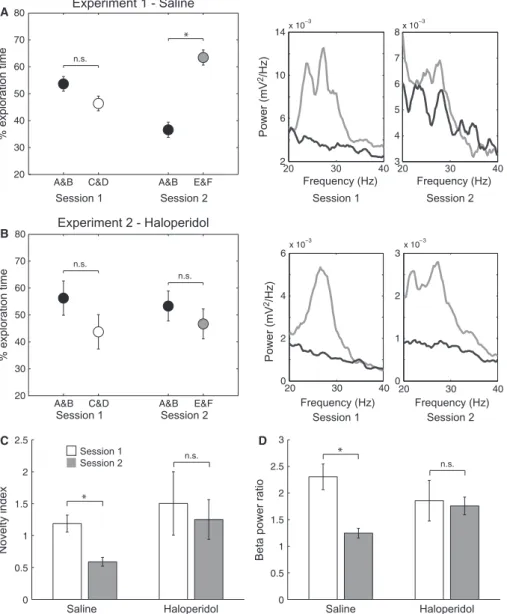
Each experiment consisted of two sessions separated by 24 h, in which animals were allowed to explore four objects in an openfield for 10 min (Fig. 1A). The four objects were novel in session 1, while only two objects were novel in session 2 (the other two objects were the same as in session 1). Behavioural analysis showed that animals explored the objects in sessions 1 and 2 of experiment 1 (saline injection after session 1; see Materials and Methods). In session 1, animals spent roughly the same amount of time exploring the four novel objects, while in session 2 animals spent more time exploring the only two novel objects (pairedt-test, t4= 3.83,P=0.0186; Fig. 2), which shows that animals recognised the two familiar objects.
We started the electrophysiological analyses by first characteris-ing the oscillatory content of hippocampal LFPs durcharacteris-ing the explora-tion of the four novel objects in session 1 of experiment 1. As expected, spectral analyses revealed robust theta oscillations throughout the 10-min exploration period (see Figs 3A and 4B). Interestingly, and consistent with Berke et al. (2008), we found prominent beta2 activity mostly during the beginning of session 1 (Fig. 3B). The mean latency to peak beta2 activity among animals was 559 s (range 35–80 s; Fig. 3B inset). Beta2 power in the first 100 s of object exploration was 2.300.24 times higher than in the last 100 s (paired t-test compared to 1, t16= 5.41, P< 0.0001; Fig. 4A). The power of other frequency bands such as theta and gamma was also higher in the beginning of the explora-tion session (Fig. 4), but to a much lower extent than that observed for beta2 [maximum power ratio of 1.310.07 (first 100 s/last 100 s) for theta oscillations]; indeed, the increase in power seen at the beginning of session 1 was statistically significantly larger for beta2 than for four other analysed frequency bands (F4,80= 15.44,
P< 0.0001, one-way ANOVA; Fig. 4). As shown in Fig. 5, the decrease in beta2 power along the session correlated with the decrease in locomotion (r= 0.49, P< 0.01), but not with the time animals spent exploring objects (P= 0.35; compare Figs 2A and 3B).
We next confirmed the results above by visual inspection of raw LFPs. To that end, wefirstfiltered the LFP into the beta2 band and localised the periods of high beta2 amplitude (which, consistent with the power analysis, occurred mostly at the beginning of the session; Figs 6A and 7A). We then examined the unfiltered LFP at these periods and found that both sustained theta oscillations and bursts of beta2 activity could be directly observed; the latter was characterised by high-amplitude, sharp LFP deflections (Figs 6B and 7B inset). The mean duration of beta2 bursts was 16742 ms (Fig. 7B). Therefore, beta2 oscillations are a genuine LFP activity and not har-monics or artifacts of the spectral analysis. In all, these results show that beta2 has a unique dynamics along the first object exploration session, with most beta2 bursts occurring at the beginning of the session.
Beta2 power is lower when animals explore two familiar and two novel objects
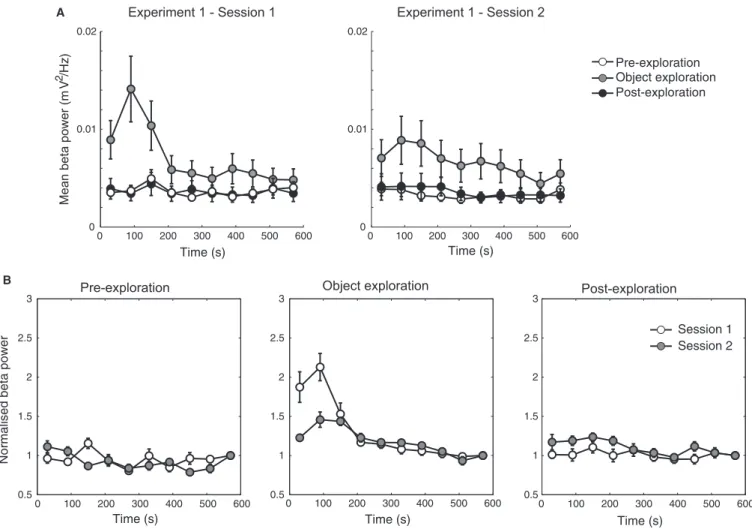
We next examined LFP power content in the second exploration session of experiment 1, as well as performed spectral analyses during the pre- and post-exploration periods in the home cage. As shown in Fig. 8A, we found that the transient increase in beta2 power occurred only during the object exploration period in both the first and second sessions (repeated-measures ANOVA,
F2,462= 25.19,P < 0.0001 andF2,393= 24.26, P< 0.0001, for the
first and second sessions, respectively), but not when animals were recorded in their home cage. Interestingly, when power values were normalised by dividing by the mean power in the last minute of exploration, we found that the transient increase in normalised beta2 power was larger in the first than in the second object exploration session (repeated-measures ANOVA, F1,283= 5325.16, P< 0.0001; Fig. 8B). These results suggest that a greater level of novelty in the first session compared to the second (four vs two novel objects, respectively) is associated with higher beta2 activity.
0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4 2.6
Time (s)
* *
*
*
Norm. beta power
Time (s)
Frequency (Hz)
50 100 150 200 250 300 350 400 450 500 550 0
10 20 30 40
Time (s)
Frequency (Hz)
40 60 80 100 120 140 15
20 25 30 35 40
0 0.1
A B
Power (mV
2/Hz)
Object exploration session 1 (Experiment 1) Object exploration session 1 (Experiment 1)
0 100 200 300 400 500 600
0 10 20 30 40 50 60 70 80
Latency to peak beta activity (s)
Beta2 oscillations modulate hippocampal but not S1 and M1 neurons
We next analysed LFP signals simultaneously recorded from S1 and M1. In contrast to hippocampal signals, S1 and M1 LFPs exhibited no power peak in the beta2 range (Fig. 9A). We then investigated whether spiking activity was coupled to beta2. Fig. 9B shows an example of multiunit activity in CA1, which was highly modulated by both theta and beta2 oscillations (see also Berkeet al., 2008). At the group level, while hippocampal beta2 modulated CA1 neurons (Rayleigh test,P < 0.0001), population activity recorded at neocorti-cal regions was not coupled to beta2 phase (Fig. 9C). Thus, nov-elty-related beta2 oscillations seem to occur and modulate spiking activity in the hippocampus but not in S1 or M1 (see Discussion).
Impairing object recognition is associated with resurgence of prominent beta2 activity
One week after the first set of recordings (experiment 1) we sub-jected animals to a similar protocol, but using different objects (experiment 2). In addition, in experiment 2 animals were treated
with haloperidol (0.3 mg/kg i.p.) instead of saline immediately after the first object exploration session; haloperidol is amnesic in this paradigm (Lob~ao-Soares et al., 2009; A.S.C. Francßa, B. Lob~a o-Soares, L. Muratori, G.C. do Nascimento, J. Winne, C.M. Pereira, S.M.B. Jeronimo, S. Ribeiro, unpublished observations). While ani-mals treated with saline after the first session of experiment 1 showed lower preference for the two familiar objects (A and B) in the second session (pairedt-test, t4=4.759, P=0.0089; Fig. 10A left panel), haloperidol injected after the first session of experi-ment 2 led animals to display no object preference in the second session (Fig. 10B left panel). Thus, in the saline protocol the nov-elty index in the second exploration session was lower than in the first session (paired t-test, t4=4.262, P=0.0130; Fig. 10C, left bars), while in the haloperidol protocol the novelty index was not different between sessions 1 and 2 (Fig. 10C, right bars), suggesting that amnesic animals perceived the four objects as equally novel in the second session.
Interestingly, while animals displayed a lower increase in beta2 power in the beginning of the second exploration session of experi-ment 1 (c.f. Fig. 8), animals treated with haloperidol exhibited B
4–12 Hz
Spectral band
First 100 s Last 100 s
14–20 Hz 23–30 Hz 40–100 Hz 150–250 Hz 0
0.5 1 1.5 2 2.5 3
Power ratio (First 100 s/Last 100 s)
5 15 25 35 45 55 65 75 0
0.3
Frequency (Hz)
20 30 40 0
0.04
Frequency (Hz)
5 15 25 35 45 55 65 75 0
0.1
Frequency (Hz)
20 30 40
0 0.015
Frequency (Hz)
5 15 25 35 45 55 65 75 0
0.3
Frequency (Hz)
20 30 40
0 0.03
Frequency (Hz)
A
Power (mV
2/Hz)
Object exploration session 1 (Experiment 1)
#
*
*
*
*
Fig. 4. Beta2 oscillations increase more in power than other hippocampal oscillations at the beginning of the exploration session. (A) Power ratios forfive fre-quency ranges, as labeled. The power ratio is defined as the mean power in the analysed frefre-quency band in thefirst 100 s of the session divided by the mean band power in the last 100 s. Bars denote meanSEM;*P< 0.05,t-test against 1, Bonferroni-corrected forfive comparisons;#P< 0.0001 compared to all
other spectral bands, one-wayANOVAfollowed by Tukey’spost hoctest. (B) Power spectrum density for thefirst (light gray) and last (dark gray) 100 s of object exploration in three representative animals. Insets show the beta2 range.
0 10 20 30 40 50 60
0.5 1 1.5 2 2.5 3 3.5
r = 0.49
P < 0.01
Time in locomotion (s)
0 5 10 15 20 25
0.5 1 1.5 2 2.5 3 3.5
P = 0.35
Norm. beta power
Norm. beta power
Time exploring objects (s) Object exploration session 1 (Experiment 1)
prominent beta2 activity in both the first and second sessions of experiment 2 (see Fig. 10A and B, right panels, for representative examples). Thus, the beta2 power ratio (first 100 s/last 100 s) decreased from 2.3 in thefirst session of experiment 1 to 1.2 in the second session (t-test, t31=4.011, P< 0.001; Fig. 10D, left bars), while it was close to 1.8 in both thefirst and second explora-tion sessions of experiment 2 (Fig. 10D, right bars). In all, these results suggest that changes in beta2 power within sessions parallel
changes in behaviour (compare Fig. 10C and D), with greater beta2 activity in sessions associated with a greater number of novel objects as putatively perceived by the animals.
Discussion
We have characterised hippocampal beta2 oscillations through extra-cellular recordings, pharmacological intervention and behavioural
1 s
1 mV
A
B
Experiment 1 - Session 1
Beta (23–30 Hz)
Theta (4–12 Hz)
Fig. 6.Beta2 oscillations have large amplitude and are observable in unfilteredfield potentials. (A) Raw LFP (black) along with beta2- (red) and theta-filtered (blue) signals for 15-s epochs recorded at the beginning (left) and end (right) of the object exploration session of experiment 1. Notice prominent beta2 activity at the beginning of the session, while robust theta oscillations exist throughout the session (see also Figs 3B and 4B). (B) Expanded view of a 1-s-long period of the traces shown in A. Note a clear burst of beta2 waves in the raw LFP at the beginning of the session.
0 60 120 180 240 300 360 420 480 540 600
0 5 10 15 20 25 30 35
Time (s)
Number of beta bursts
0 100 200 300 400 500 600 700
200 ms
1 mV
0 5 10 15 20 25 30
Burst duration (ms)
%
A B
Beta (23–30 Hz)
Theta (4–12 Hz)
Experiment 1 - Session 1
analysis in a novel-object recognition task. We found prominent bursts of beta2 oscillations when mice were allowed to explore four novel objects in an open field. The occurrence of beta2 bursts was transient and concentrated at the beginning of the exploration session. Compared to basal levels, beta2 activity was also higher in the beginning of a second session in which animals explored two novel and two familiar objects, but to a lower extent than in thefirst exploration session with four novel objects. Interestingly, however, the transient increase in beta2 activity was higher in the second ses-sion of experiment 2, when animals putatively perceived familiar objects as novel due to the injection of an amnesic drug immedi-ately after the first exploration session. Taken together, our results suggest that the appearance of beta2 activity in the hippocampus is associated with novel experience.
The hippocampus plays a key role in memory formation (Squire, 1992; Eichenbaum, 2004), spatial navigation (O’Keefe & Dostrovsky, 1971; O’Keefe & Nadel, 1978), context discrimination (Frankland et al., 1998; Mizumori et al., 2007; Tort et al., 2011),
‘match–mismatch’ operations and novelty detection (Knight, 1996; Lisman & Otmakhova, 2001; Kumaran & Maguire, 2007; Duncan et al., 2012). Growing evidence indicates that network oscillations are important for the hippocampus to execute its functions (Mont-gomery & Buzsaki, 2007; Tort et al., 2009; Jutras et al., 2013). Given the large amplitude of the beta2 oscillations observed here,
which allow their direct observation by visual inspection of raw field potentials, it is quite surprising that (to the best of our knowl-edge) these hippocampal oscillations have only been reportedin vivo once (Berkeet al., 2008). This could be due to a selective appear-ance of beta2 in specific behavioural states of mice. In contrast, theta and gamma oscillations always accompany exploratory activ-ity, while ripple oscillations are common during consummatory behaviour such as drinking and grooming (Buzsakiet al., 2003).
Our results corroborate several of the beta2 characteristics first reported in Berkeet al.(2008), such as their large amplitude, transient appearance in the beginning of a novel exploration session, and modu-lation of hippocampal units. Interestingly, in both studies peak beta2 activity was not at the onset of the novel experience but occurred after a latency period. This latency period may reflect the time animals take to perceive the experience as novel (or to generate a mismatch from previous expectations; Grossberg, 2009), and/or for stable placefield representations to emerge. It should be noted that, while we found a positive correlation between beta2 power and locomotion activity (as both decreased along the session), we believe there is no causal rela-tionship between them: peak locomotion tended to occur at the very onset of the exploration session and did not coincide with peak beta2 activity. Moreover, Berkeet al.(2008) reported the disappearance of beta2 activity after a couple laps in a novel rectangular arena, despite the fact that animals continued to run. The decrease in beta2 activity Pre-exploration
Experiment 1 - Session 1 Experiment 1 - Session 2
Post-exploration Object exploration
0 100 200 300 400 500 600
0.5 1 1.5 2 2.5 3
Time (s)
0 100 200 300 400 500 600
0.5 1 1.5 2 2.5 3
Time (s)
Normalised beta power
0 100 200 300 400 500 600
0.5 1 1.5 2 2.5 3
Time (s)
Session 1 Session 2
0 100 200 300 400 500 600
0 0.01 0.02
Time (s)
0 100 200 300 400 500 600
0 0.01 0.02
Time (s)
Mean beta power (m
V
2 /Hz)
Pre-exploration Object exploration Post-exploration
B
A
with time, along with no changes in spatial context or animals’ behav-iour, reported in Berkeet al.(2008) is consistent with our findings showing no correlation between beta2 and the time animals spent exploring the objects.
It should be mentioned that Berke et al. (2008) and the present study obtained results from different mouse strains. Specifically, Berkeet al. (2008) observed beta2 bursts in a genetically modified mouse strain (‘fNR1 mouse’; Tsien et al., 1996) and pointed to the importance of examining whether theirfindings would hold true for wild-type animals. Here we show that this is indeed the case. However, whether similar hippocampal beta2 activity exists in other species such as rats remains to be demonstrated. In addition to cor-roborating previousfindings, here we went on to demonstrate that blocking familiar object recognition is associated with the reappear-ance of prominent of beta2 oscillations as in the first exploration session. This result goes well with previous theoretical accounts (Grossberg, 2009) and further supports a role for hippocampal beta2 oscillations in novel experience.
We found that hippocampal but not S1 or M1 recordings exhib-ited beta2 oscillations, and, moreover, beta2 phase-modulated only CA1 but neither M1 nor S1 neurons. These results suggest a certain specificity of novelty-related beta2 activity to the hippocampus. Nevertheless, beta2 oscillations were also recently reported in the basal forebrain of rats during an associative learning task (Quinn et al., 2010). In this study, beta2 power was higher in thefirst day
of learning in which the object–reward pairs were novel than in sub-sequent days when pairings became familiar (Quinn et al., 2010). Moreover, within the first day of learning, beta2 power was lowest in thefirst trials but increased with later encounters with the objects (Quinn et al., 2010), akin to the latency period for maximal beta2 power observed here. In contrast, however, Quinn et al. (2010) did not observe a disappearance of beta2 activity after objects became familiar. The basal forebrain possesses cholinergic, glutamatergic and GABAergic neurons that project to widespread regions of the cortex, including the hippocampus (McKinney et al., 1983; Mesu-lamet al., 1983; Grittiet al., 1997; Mannset al., 2003). Basal fore-brain projections modulate synaptic plasticity in the cortex (Kilgard & Merzenich, 1998; Conneret al., 2005) and are involved in atten-tional processes at the behavioural level (Muir et al., 1993; Voytko et al., 1994; Chiba et al., 1995). Beta2 oscillations could be thus involved in modulating the saliency of novel stimuli in downstream areas; nevertheless, whether the beta2 oscillations described in Quinn et al. (2010) are related to the beta2 in the hippocampus remains to be determined.
Several types of oscillatory activity can be obtained in the hippo-campus in vitro, and many believe that these would correspond to theirin vivocounterparts (Traubet al., 1996; Whittington & Traub, 2003). Previous work has shown that the internal circuits of the hip-pocampus are able to produce theta, gamma and ripple frequency oscillations (Whittington et al., 2000; Maieret al., 2003;
Whitting-0 180 360 540 720
0 0.1 0.2
P = 1.45e−25
Firing probability
Theta phase (Deg)
0 180 360 540 720
P = 3.94e−06
Beta phase (Deg)
0 180 360 540 720
0.06 0.08 0.1 0.12 0.14
Firing probability
CA1 beta phase (Deg)
0 180 360 540 720
CA1 beta phase (Deg)
0 180 360 540 720
CA1 beta phase (Deg)
CA1 M1 S1
P = 0.29
P = 0.89
P = 2.14e−05
20 25 30 35
0 1 2 3 4 5 6
7x 10
−3
Frequency (Hz)
Power (mV
2/Hz)
S1 CA1
M1
A B
C
ton & Traub, 2003; Colginet al., 2004, 2005; Gloveli et al., 2005; Goutagny et al., 2009). For instance, the cholinergic agonist carba-chol generates 25–40 Hz oscillations in hippocampal slices and these are considered to be related to hippocampal gamma oscillations observed in vivo (Fisahn et al., 1998; Traub et al., 2000; Dickinson et al., 2003; Palhalmi et al., 2004). However, our results prompt the speculation that the ‘gamma’ oscillations observed in hippocampal slices during cholinergic activation would actually correspond to a faster version (due to the experimental preparation) of in vivobeta2 oscillations, and not to in vivogamma oscillations as previously suggested. Consistent with this hypothesis, the sharp wave shape of‘gamma’in the slice as well as its narrow power peak (Fisahn et al., 1998; Gloveli et al., 2005; Le~aoet al.,
2009) resemblein vivobeta2 oscillations more thanin vivogamma. Moreover, in vivo hippocampal beta2 oscillations seem to depend on CA3 (Berkeet al., 2008), similarly to‘gamma’in the slice (Fis-ahn et al., 1998). Acetylcholine release has been linked to novelty and saliency detection (Acquas et al., 1996), which is putatively associated with beta2 bursts, whereas gamma oscillations exist even in the absence of salient or novel stimuli. Furthermore, as men-tioned above, the basal forebrain is a major source of acetylcholine and also produces beta2 oscillations in vivo (Quinn et al., 2010). Finally, it should be noted that oscillations in the beta2 frequency range have actually been observed in hippocampal slices under cho-linergic activation in similar protocols as used for generating
‘gamma’ (Shimono et al., 2000; Colgin, 2006), but they probably Experiment 1 - Saline
A
Session 1 Session 2
A&B C&D A&B E&F 20
30 40 50 60 70 80
% exploration time
*
A&B C&D A&B E&F 20
30 40 50 60 70 80
% exploration time
Experiment 2 - Haloperidol
B
0 0.5 1 1.5 2 2.5
Novelty index
*
Session 1
Session 1 Session 2
Session 2
C
20 30 40
2 6 10 14x 10
−3
Frequency (Hz)
Power (mV
2/Hz)
20 30 40
3 4 5 6 7 8x 10
−3
Frequency (Hz)
20 30 40
0 2 4 6
Frequency (Hz)
x 10−3
20 30 40
0 1 2 3
Frequency (Hz)
x 10−3
Power (mV
2/Hz)
Session 1 Session 2
Session 1 Session 2
0 0.5 1 1.5 2 2.5 3
*
Beta power ratio
D
Saline Haloperidol Saline Haloperidol
n.s.
n.s.
n.s.
n.s.
n.s.
received less attention than gamma due to the lack of its in vivo correspondent. Further work combining in vitro and in vivo tech-niques should test whether ‘gamma’ in hippocampal slice prepara-tions might in reality correspond to the novelty-related beta2 activity reported here.
An influential model proposes that the hippocampus and the ven-tral tegmental area (VTA) form a loop that controls the formation of long-term memory (Lisman & Grace, 2005). Accordingly, the hip-pocampus would be responsible for detecting new information and sending novelty signals to the VTA, which would integrate this information with others (such as saliency) and in turn release dopa-mine in the hippocampus, enhancing LTP and learning. Such a model is consistent with electrophysiological, molecular and behav-iouralfindings (Gasbarri et al., 1996; Otmakhova & Lisman, 1996; Lemon & Manahan-Vaughan, 2006; Morice et al., 2007; Terry et al., 2007; Rossatoet al., 2009). However, there is not much liter-ature about the influence of D2 antagonists on the novel object rec-ognition task used here. We have recently found that haloperidol blocks object memory consolidation (A.S.C. Francßa, B. Lob~a o-Soares, L. Muratori, G.C. do Nascimento, J. Winne, C.M. Pereira, S.M.B. Jeronimo, S. Ribeiro, unpublished observations), as observed in the present study. These findings are consistent with a role of dopaminergic projections to the hippocampus in controlling the for-mation of long-term memory (Lisman & Grace, 2005; Rossato et al., 2009). Whether hippocampal beta2 oscillations would take part in the communication between the hippocampus and VTA remains to be established.
In summary, our results demonstrate the appearance of transient beta2 oscillations in the hippocampus during exploration of novel objects. As hypothesised by Grossberg (1999), thesefindings suggest that beta2 may be involved in signaling specific time periods for new plasticity to occur. However, much yet should be done to characterise their biophysical mechanisms of generation, regions of occurrence, as well as cognitive roles. In particular, it would be interesting to know whether disruption of beta2 activity during memory acquisition affects behaviour and LFP spectral content when testing memory retrieval. Most importantly, thefield would benefit from independent labs reporting a similar pattern of organised electrical activity; we concur with many others (Buttonet al., 2013) in the belief that repli-cation is key for constructing solid knowledge.
Acknowledgments
Supported by grants of Conselho Nacional de Desenvolvimento Cientıfico e Tecnologico (CNPq)/Ministerio da Ci^encia e Tecnologia (MCT), Coorde-nacß~ao de Aperfeicßoamento de Pessoal de Nıvel Superior (CAPES), Fundacß~ao de Apoioa Pesquisa do Estado do Rio Grande do Norte (FAPERN), Financi-adora de Estudos e Projetos (FINEP), Pro-Reitoria de Pos-Graduacß~ao da Uni-versidade Federal do Rio Grande do Norte (UFRN), Programa de Apoio a Nucleos Emergentes (PRONEM 003/2011 FAPERN/CNPq), and Pew Latin American Fellows Program in the Biomedical Sciences. The authors thank Cesar Renno-Costa, Richardson Le~ao and Jurij Brankack for helpful discus-sions. The authors declare no competingfinancial interests.
Abbreviations
LFP, localfield potential; M1, primary motor cortex; S1, primary somatosen-sory cortex.
References
Acquas, E., Wilson, C. & Fibiger, H.C. (1996) Conditioned and unconditioned stimuli increase frontal cortical and hippocampal acetylcholine release: effects of novelty, habituation, and fear.J. Neurosci.,16, 3089–3096.
Berke, J.D., Hetrick, V., Breck, J. & Greene, R.W. (2008) Transient 23-30 Hz oscillations in mouse hippocampus during exploration of novel environments.Hippocampus,18, 519–529.
Button, K.S., Ioannidis, J.P., Mokrysz, C., Nosek, B.A., Flint, J., Robinson, E.S. & Munafo, M.R. (2013) Power failure: why small sample size under-mines the reliability of neuroscience.Nat. Rev. Neurosci.,14, 365–376. Buzsaki, G. (2002) Theta oscillations in the hippocampus.Neuron,33, 325–
340.
Buzsaki, G. (2004) Large-scale recording of neuronal ensembles.Nat. Neuro-sci.,7, 446–451.
Buzsaki, G. (2006) Rhythms of The Brain. Oxford University Press, New York.
Buzsaki, G. & Draguhn, A. (2004) Neuronal oscillations in cortical networks. Science,304, 1926–1929.
Buzsaki, G., Horvath, Z., Urioste, R., Hetke, J. & Wise, K. (1992) High-fre-quency network oscillation in the hippocampus.Science,256, 1025–1027. Buzsaki, G., Buhl, D., Harris, K., Csicsvari, J., Czeh, B. & Morozov, A.
(2003) Hippocampal network patterns of activity in the mouse. Neurosci-ence,116, 201–211.
Buzsaki, G., Anastassiou, C.A. & Koch, C. (2012) The origin of extracellular fields and currents - EEG, ECoG, LFP and spikes. Nat. Rev. Neurosci., 13, 407
–420.
Chiba, A.A., Bucci, D.J., Holland, P.C. & Gallagher, M. (1995) Basal fore-brain cholinergic lesions disrupt increments but not decrements in condi-tioned stimulus processing.J. Neurosci.,15, 7315–7322.
Colgin, L.L. (2006) Investigation of network phenomena in hippocampal slices using multi-electrode recording arrays. In Taketani, M. & Baudry, M. (Eds), Advances in Network Electrophysiology. Electrophysiology, Springer, pp. 425–453.
Colgin, L.L., Kubota, D., Jia, Y., Rex, C.S. & Lynch, G. (2004) Spontaneous waves in the dentate gyrus of slices from the ventral hippocampus. J. Physiol.,558, 953–961.
Colgin, L.L., Jia, Y., Sabatier, J.M. & Lynch, G. (2005) Blockade of NMDA receptors enhances spontaneous sharp waves in rat hippocampal slices. Neurosci. Lett.,385, 46–51.
Colgin, L.L., Denninger, T., Fyhn, M., Hafting, T., Bonnevie, T., Jensen, O., Moser, M.-B. & Moser, E.I. (2009) Frequency of gamma oscilla-tions routes flow of information in the hippocampus. Nature,462, 353– 357.
Conner, J.M., Chiba, A.A. & Tuszynski, M.H. (2005) The basal forebrain cholinergic system is essential for cortical plasticity and functional recov-ery following brain injury.Neuron,46, 173
–179.
Csicsvari, J., Jamieson, B., Wise, K.D. & Buzsaki, G. (2003) Mechanisms of gamma oscillations in the hippocampus of the behaving rat.Neuron,37, 311–322.
Dere, E., Huston, J.P. & De Souza Silva, M.A. (2007) The pharmacology, neuroanatomy and neurogenetics of one-trial object recognition in rodents. Neurosci. Biobehav. R.,31, 673–704.
Dickinson, R., Awaiz, S., Whittington, M., Lieb, W. & Franks, N. (2003) The effects of general anaesthetics on carbachol-evoked gamma oscilla-tions in the rat hippocampus in vitro.Neuropharmacology,44, 864
–872. Duncan, K., Ketz, N., Inati, S.J. & Davachi, L. (2012) Evidence for area
CA1 as a match/mismatch detector: a high-resolution fMRI study of the human hippocampus.Hippocampus,22, 389–398.
Ego-Stengel, V. & Wilson, M.A. (2010) Disruption of ripple-associated hip-pocampal activity during rest impairs spatial learning in the rat. Hippocam-pus,20, 1–10.
Eichenbaum, H. (2004) Hippocampus: cognitive processes and neural repre-sentations that underlie declarative memory.Neuron,44, 109–120. Fisahn, A., Pike, F.G., Buhl, E.H. & Paulsen, O. (1998) Cholinergic
induc-tion of network oscillainduc-tions at 40 Hz in the hippocampus in vitro.Nature, 394, 186
–189.
Frankland, P.W., Cestari, V., Filipkowski, R.K., McDonald, R.J. & Silva, A.J. (1998) The dorsal hippocampus is essential for context discrimination but not for contextual conditioning.Behav. Neurosci.,112, 863–874. Franklin, K.B. & Paxinos, G. (2007)The Mouse Brain in Stereotaxic
Coordi-nates. Academic Press, San Diego.
Gasbarri, A., Sulli, A., Innocenzi, R., Pacitti, C. & Brioni, J.D. (1996) Spa-tial memory impairment induced by lesion of the mesohippocampal dopa-minergic system in the rat.Neuroscience,74, 1037–1044.
Girardeau, G., Benchenane, K., Wiener, S.I., Buzsaki, G. & Zugaro, M.B. (2009) Selective suppression of hippocampal ripples impairs spatial mem-ory.Nat. Neurosci.,12, 1222–1223.
arrangement of rhythm-generating microcircuits in the hippocampus.Proc. Natl. Acad. Sci. USA,102, 13295
–13300.
Goutagny, R., Jackson, J. & Williams, S. (2009) Self-generated theta oscilla-tions in the hippocampus.Nat. Neurosci.,12, 1491–1493.
Gritti, I., Mainville, L., Mancia, M. & Jones, B.E. (1997) GABAergic and other noncholinergic basal forebrain neurons, together with cholinergic neurons, project to the mesocortex and isocortex in the rat.J. Comp. Neu-rol.,383, 163–177.
Grossberg, S. (1980) How does a brain build a cognitive code? Psychol. Rev.,87, 1–52.
Grossberg, S. (1999) The link between brain learning, attention, and con-sciousness.Conscious. Cogn.,8, 1–44.
Grossberg, S. (2009) Beta oscillations and hippocampal place cell learning during exploration of novel environments.Hippocampus,19, 881–885. Heyser, C.J. & Chemero, A. (2012) Novel object exploration in mice: not all
objects are created equal.Behav. Process.,89, 232–238.
Hughes, R.N. (1997) Intrinsic exploration in animals: motives and measure-ment.Behav. Process.,41, 213–226.
Hughes, R.N. (2007) Neotic preferences in laboratory rodents: issues, assess-ment and substrates.Neurosci. Biobehav. R.,31, 441–464.
Jutras, M.J., Fries, P. & Buffalo, E.A. (2013) Oscillatory activity in the monkey hippocampus during visual exploration and memory formation. Proc. Natl. Acad. Sci. USA,110, 13144–13149.
Kamondi, A., Acsady, L., Wang, X.-J. & Buzsaki, G. (1998) Theta oscillations in somata and dendrites of hippocampal pyramidal cells in vivo: activity-dependent phase-precession of action potentials.Hippocampus,8, 244–261. Kilgard, M.P. & Merzenich, M.M. (1998) Cortical map reorganization
enabled by nucleus basalis activity.Science,279, 1714–1718.
Knight, R.T. (1996) Contribution of human hippocampal region to novelty detection.Nature,383, 256–259.
Kumaran, D. & Maguire, E.A. (2007) Which computational mechanisms operate in the hippocampus during novelty detection? Hippocampus,17, 735–748.
Le~ao, R.N., Tan, H.M.T. & Fisahn, A. (2009) Kv7/KCNQ Channels control action potential phasing of pyramidal neurons during hippocampal gamma oscillations in vitro.J. Neurosci.,29, 13353–13364.
Lemon, N. & Manahan-Vaughan, D. (2006) Dopamine D1/D5 receptors gate the acquisition of novel information through hippocampal long-term poten-tiation and long-term depression.J. Neurosci.,26, 7723–7729.
Linden, H., Tetzlaff, T., Potjans, T.C., Pettersen, K.H., Grun, S., Diesmann, M. & Einevoll, G.T. (2011) Modeling the spatial reach of the LFP. Neu-ron,72, 859
–872.
Lisman, J.E. & Grace, A.A. (2005) The hippocampal-VTA loop: controlling the entry of information into long-term memory.Neuron,46, 703–713. Lisman, J.E. & Otmakhova, N.A. (2001) Storage, recall, and novelty
detec-tion of sequences by the hippocampus: elaborating on the SOCRATIC model to account for normal and aberrant effects of dopamine. Hippocam-pus,11, 551–568.
Lob~ao-Soares, B., Muratori, L., Francßa, A., Pereira, C., Nascimento, G., Nicolelis, M. & Ribeiro, S. (2009) REM sleep suppression by dopamine D2 receptors blockade leads to learning impairment.Behav. Pharmacol., 20, S82–S103.
Maier, N., Nimmrich, V. & Draguhn, A. (2003) Cellular and network mecha-nisms underlying spontaneous sharp wave-ripple complexes in mouse hip-pocampal slices.J. Physiol.,550, 873–887.
Manns, I.D., Alonso, A. & Jones, B.E. (2003) Rhythmically discharging basal forebrain units comprise cholinergic, GABAergic, and putative gluta-matergic cells.J. Neurophysiol.,89, 1057–1066.
McKinney, M., Coyle, J.T. & Hedreen, J.C. (1983) Topographic analysis of the innervation of the rat neocortex and hippocampus by the basal fore-brain cholinergic system.J. Comp. Physiol.,217, 103–121.
Mesulam, M., Mufson, E.J., Levey, A.I. & Wainer, B.H. (1983) Cholinergic innervation of cortex by the basal forebrain: cytochemistry and cortical connections of the septal area, diagonal band nuclei, nucleus basalis (sub-stantia innominata), and hypothalamus in the rhesus monkey. J. Comp. Neurol.,214, 170–197.
Mizumori, S.J., Smith, D.M. & Puryear, C.B. (2007) Hippocampal and neo-cortical interactions during context discrimination: electrophysiological evi-dence from the rat.Hippocampus,17, 851–862.
Montgomery, S.M. & Buzsaki, G. (2007) Gamma oscillations dynamically couple hippocampal CA3 and CA1 regions during memory task perfor-mance.Proc. Natl. Acad. Sci. USA,104, 14495–14500.
Morice, E., Billard, J.M., Denis, C., Mathieu, F., Betancur, C., Epelbaum, J., Girus, B. & Nosten-Bertrand, M. (2007) Parallel loss of hippocampal LTD
and cognitiveflexibility in a genetic model of hyperdopaminergia. Neuro-psychopharmacology,32, 2108
–2116.
Muir, J.L., Page, K.J., Sirinathsinghji, D., Robbins, T.W. & Everitt, B.J. (1993) Excitotoxic lesions of basal forebrain cholinergic neurons: effects on learning, memory and attention.Behav. Brain Res.,57, 123–131. Nakazawa, K., Sun, L.D., Quirk, M.C., Rondi-Reig, L., Wilson, M.A. &
Tonegawa, S. (2003) Hippocampal CA3 NMDA receptors are crucial for memory acquisition of one-time experience.Neuron,38, 305–315. O’Keefe, J. & Dostrovsky, J. (1971) The hippocampus as a spatial map:
pre-liminary evidence from unit activity in the freely-moving rat.Brain Res., 34, 171
–175.
O’Keefe, J. & Nadel, L. (1978) The Hippocampus as a Cognitive Map. Oxford University Press, New York.
Otmakhova, N.A. & Lisman, J.E. (1996) D1/D5 dopamine receptor activa-tion increases the magnitude of early long-term potentiaactiva-tion at CA1 hippo-campal synapses.J. Neurosci.,16, 7478–7486.
Palhalmi, J., Paulsen, O., Freund, T. & Hajos, N. (2004) Distinct properties of carbachol-and DHPG-induced network oscillations in hippocampal slices.Neuropharmacology,47, 381–389.
Quinn, L.K., Nitz, D.A. & Chiba, A.A. (2010) Learning-dependent dynamics of beta-frequency oscillations in the basal forebrain of rats.Eur. J. Neuro-sci.,32, 1507
–1515.
Reimann, M.W., Anastassiou, C.A., Perin, R., Hill, S.L., Markram, H. & Koch, C. (2013) A biophysically detailed model of neocortical localfield potentials predicts the critical role of active membrane currents.Neuron, 79, 375–390.
Rossato, J.I., Bevilaqua, L.R.M., Izquierdo, I., Medina, J.H. & Cammarota, M. (2009) Dopamine controls persistence of long-term memory storage. Science,325, 1017–1020.
Scheffer-Teixeira, R., Belchior, H., Caixeta, F.V., Souza, B.C., Ribeiro, S. & Tort, A.B. (2012) Theta phase modulates multiple layer-specific oscilla-tions in the CA1 region.Cereb. Cortex,22, 2404–2414.
Shimono, K., Brucher, F., Granger, R., Lynch, G. & Taketani, M. (2000) Origins and distribution of cholinergically induced beta rhythms in hippo-campal slices.J. Neurosci.,20, 8462–8473.
Squire, L.R. (1992) Memory and the hippocampus: a synthesis fromfindings with rats, monkeys, and humans.Psychol. Rev.,99, 195–231.
Terry, A.V., Gearhart, D.A., Warner, S., Hohnadel, E.J., Middlemore, M.-L., Zhang, G., Bartlett, M.G. & Mahadik, S.P. (2007) Protracted effects of chronic oral haloperidol and risperidone on nerve growth factor, choliner-gic neurons, and spatial reference learning in rats. Neuroscience, 150, 413–424.
Tort, A.B., Komorowski, R.W., Manns, J.R., Kopell, N.J. & Eichenbaum, H. (2009) Theta-gamma coupling increases during the learning of item-con-text associations.Proc. Natl. Acad. Sci. USA,106, 20942–20947. Tort, A.B., Komorowski, R., Kopell, N. & Eichenbaum, H. (2011) A
mecha-nism for the formation of hippocampal neuronalfiring patterns that repre-sent what happens where.Learn. Memory,18, 718–727.
Tort, A.B., Scheffer-Teixeira, R., Souza, B.C., Draguhn, A. & Brankack, J. (2013) Theta-associated high-frequency oscillations (110–160 Hz) in the hippocampus and neocortex.Prog. Neurobiol.,100, 1
–14.
Traub, R., Whittington, M., Colling, S., Buzsaki, G. & Jefferys, J. (1996) Analysis of gamma rhythms in the rat hippocampus in vitro and in vivo. J. Physiol.,493, 471–484.
Traub, R.D., Bibbig, A., Fisahn, A., LeBeau, F.E., Whittington, M.A. & Buhl, E.H. (2000) A model of gamma-frequency network oscillations induced in the rat CA3 region by carbachol in vitro.Eur. J. Neurosci.,12, 4093–4106.
Tsien, J.Z., Chen, D.F., Gerber, D., Tom, C., Mercer, E.H., Anderson, D.J., Mayford, M., Kandel, E.R. & Tonegawa, S. (1996) Subregion-and cell type-restricted gene knockout in mouse brain.Cell,87, 1317–1326. Vanderwolf, C. (1969) Hippocampal electrical activity and voluntary
move-ment in the rat.Electroen. Clin. Neuro.,26, 407–418.
Voytko, M.L., Olton, D.S., Richardson, R.T., Gorman, L.K., Tobin, J. & Price, D. (1994) Basal forebrain lesions in monkeys disrupt attention but not learning and memory.J. Neurosci.,14, 167–186.
Whittington, M.A. & Traub, R.D. (2003) Interneuron Diversity series: inhibi-tory interneurons and network oscillations in vitro.Trends Neurosci., 26, 676–682.
Whittington, M., Traub, R., Kopell, N., Ermentrout, B. & Buhl, E. (2000) Inhibition-based rhythms: experimental and mathematical observations on network dynamics.Int. J. Psychophysiol.,38, 315–336.