Comparação da vegetação arbórea e características edáficas de um
cerradão e um cerrado
sensu stricto
em áreas adjacentes sobre solo
distrófico no leste de Mato Grosso, Brasil
1Ben Hur Marimon Junior2,4 e Mundayatan Haridasan3
Recebido em 10/09/2004. Aceito em 07/06/2005
RESUMO – (Comparação da vegetação arbórea e características edáficas de um cerradão e um cerrado sensu stricto em áreas adjacentes em solos distróficos no leste de Mato Grosso, Brasil). Em alguns casos, cerrado sensu stricto e cerradão ocorrem lado a lado e sob condições edáficas e topográficas semelhantes. Como os fatores que contribuem para a ocorrência de cerradão nessas situações ainda não estão totalmente esclarecidos, o objetivo do presente estudo foi comparar a composição florística e a fitossociologia de um cerradão e um cerrado sensu stricto no leste mato-grossense e verificar se maior fertilidade do solo estava contribuindo para a ocorrência do cerradão. Em cada comunidade foi realizado um levantamento florístico e fitossociológico em 50 parcelas de 10 m×10 m, onde foram medidas todas as árvores com diâmetro maior do que 5 cm a 30 cm do solo. Foram coletadas amostras de solos de um perfil de 2 m de profundidade em cada área e amostras superficiais (0-10 e 10-20 cm). As fitofisionomias foram distintas quanto a florística, estrutura e fitossociologia. A área basal (21,4 m2 ha-1) e altura média (6,4 m) do cerradão foram maiores do que as do cerrado sensu stricto (14,9 m2 ha-1 e 3,7 m). No cerradão, as três espécies mais importantes foram Hirtella glandulosa Spreg., Sclerolobium paniculatum Vog. e Xylopia
aromatica (Lam.) Mart., somando 29% do Índice do Valor de Importância (IVI) total e no cerrado sensu stricto, foram Qualea parviflora
Mart., Davilla elliptica A. St.-Hil.e Roupala montana Aubl., somando 20,5 do IVI total. Os solos sob ambas as fitofisionomias foram ácidos (pH < 5,0) e distróficos (Ca2+ < 0,4 cmol
c kg-1), com altos teores de alumínio trocável (Al3+ > 1,3 cmolc kg-1). A fertilidade do solo não apresentou diferenças para corroborar a hipótese de que a ocorrência do cerradão ao lado do cerrado sensu stricto era devida à maior fertilidade do solo. O solo sob o cerradão apresentou textura mais argilosa em todo o perfil, o que pode significar maior disponibilidade de água para as árvores durante todo o ano. Esse é um aspecto que merece investigações no futuro.
Palavras-chave: Savana, fitossociologia, fitofisionomia, fertilidade do solo
ABSTRACT– (A comparison of the woody vegetation and soil characteristics of a cerradão and a Cerrado sensu stricto in adjacents areas on dystrophic soils in eastern Mato Grosso State, Brazil). Cerrado sensu stricto and cerradão often occur side by side under similar edaphic and topographic conditions. The factors which contribute to the existence of cerradão in this situation are not well established. The objective of the present study was to compare the floristic composition and phytosociology of a Cerrado and cerradão on a dystrophic soil in eastern Mato Grosso and to determine whether higher soil fertility was contributing to the existence of the cerradão. Fifty 10 m×10 m plots were laid out in each vegetation type and a phytosociological survey was conducted of all trees with a minimum diameter of 5 cm at 30 cm above ground level. Soil samples were collected up to a depth of 2 m of soil profiles and from surface layer (0-10 and 10-20 cm) in each area. The two physiognomies showed distinct floristic, structural and phytosociological characteristics. The basal area (21.4 m2 ha-1) and the mean heights (6.4 m) of the cerradão were greater than that of the cerrado sensu stricto (14.9 m2 ha-1 and 3.7 m). The three most important species were Hirtella glandulosa Spreg., Sclerolobium paniculatum Vog. and Xylopia aromatica (Lam.) Mart. in the cerradão, representing 29% of the total Importance Value (IVI), and Qualea parviflora Mart., Davilla elliptica A. St.-Hil.and Roupala montana Aubl. in the cerrado sensu stricto, representing 21% of the total IVI. The soils of both areas were acid (pH < 5.0) and dystrophic (Ca2+ < 0.4 cmol
c kg-1) with high exchangeable aluminium levels (Al3+ > 1.3 cmolc kg-1). The fertility of the soils of the two areas was not different to support the hypothesis that the occurrence of the cerradão was due to the higher fertility of its soil. However, the cerradão soil showed higher percentages of clay than the cerrado soil at all depths up to 2 m, which could result in a higher availability of water throughout the year for the trees. This is an aspect worth investigating in future studies.
Key words: Savanna, phytosociology, phytophysiognomy, soil fertility
1 Parte da Dissertação de Mestrado do primeiro Autor
2 Universidade do Estado de Mato Grosso, BR-158, Km 145, CEP 70690-000, Nova Xavantina, MT, Brasil 3 Universidade de Brasília, Instituto de Biologia, Departamento de Ecologia, Brasília, DF
Introdução
O Bioma Cerrado brasileiro, com grande diversi-dade de formas fitofisionômicas, ocorre em 15 estados e o Distrito Federal, ocupando uma área de aproxima-damente dois milhões de km2, a qual corresponde a um quarto da superfície do país. A forma mais extensa, o cerrado sensu stricto, ocupava aproximadamente
65% da área geográfica do Bioma, enquanto que o cerradão ocupava apenas cerca de 1%. No restante da área original (34%), diversos outros tipos fitofisionômicos dividiam a paisagem.
A distribuição e a manutenção das diferentes fitofisionomias do Bioma Cerrado estão relacionadas com fatores edáficos e topográficos, além da ocorrência de fogo e perturbações antrópicas (Eiten 1972; RADAMBRASIL 1981; Oliveira-Filho et al.
1990). A profundidade efetiva, a drenagem, a presença de concreções no perfil, a profundidade do lençol freático e a fertilidade do solo são considerados os principais fatores determinantes da diversidade fitofisionômica (Eiten 1972; Haridasan 1992). O cerrado sensu stricto é uma vegetação savânica
composta por um estrato arbóreo-arbustivo e outro herbáceo-graminoso (Eiten 1994). Normalmente, ocorre sobre Latossolos e Neossolos Quartzarênicos profundos, bem drenados, distróficos, ácidos e álicos e raramente sobre solos mesotróficos (Haridasan 1992). O cerradão é uma vegetação florestal que ocorre tanto em solos distróficos quanto mesotróficos, sendo sua composição florística variável conforme a fertilidade do solo (Ratter 1971; Ratter et al. 1973; 1977; Araújo
& Haridasan 1988).
Os fatores determinantes do cerradão ainda não estão totalmente esclarecidos. Segundo Goodland (1971a; b) e Lopes & Cox (1977), a sua ocorrência estaria relacionada a um gradiente de fertilidade dos solos, onde o cerradão estaria ocupando sítios de fertilidade mais acentuada, mas não o suficiente para serem considerados solos mesotróficos. Ratter (1971) e Ratter et al. (1973) reconheceram duas situações
distintas no leste de Mato Grosso, onde um tipo de cerradão estaria relacionado a solos de alta fertilidade e outro a solos de baixa fertilidade, o qual denominaram “cerradão de Hirtella glandulosa”. Ribeiro &
Haridasan (1984) identificaram no Bioma Cerrado a ocorrência de cerradão em solos de fertilidade tão baixa quanto de cerrado sensu stricto. Costa & Araújo
(2001), trabalhando em um cerrado sensu stricto e
um cerradão adjacentes no Triângulo Mineiro, também verificaram que ambos ocorriam em solos distróficos,
com nível de fertilidade, pH e teores de Al muito semelhantes.
Na Reserva Biológica Mário Viana, no leste de Mato Grosso, o cerradão ocorre em manchas adjacentes ao cerrado sensu stricto sob condições
topográficas bastante semelhantes. O solo sob ambas as fitofisionomias não possui concreções ou qualquer outro tipo de impedimento, bem como nenhuma evidência de proximidade do lençol freático à superfície. O objetivo do presente estudo foi comparar a composição florística e a fitossociologia dessas duas vegetações e determinar se a ocorrência do cerradão está associada a uma maior fertilidade do solo.
Material e métodos
As fitofisionomias estudadas localizam-se na Reserva Biológica Municipal Mário Viana (14º41’S e 52º20’W), município de Nova Xavantina, leste do estado de Mato Grosso. É uma região de transição entre o bioma cerrado e a floresta amazônica, onde a vegetação predominante de cerrado sensu stricto está
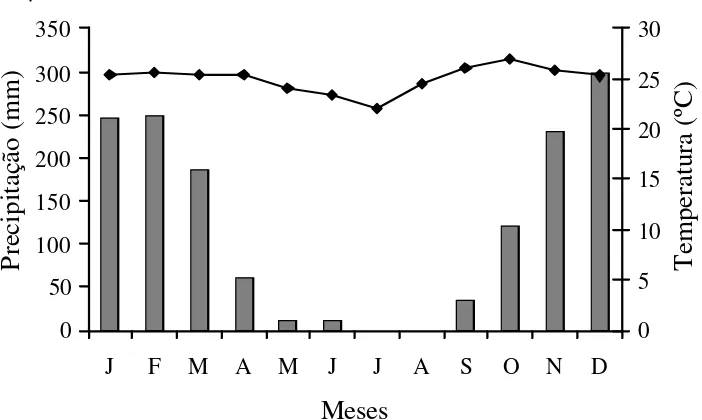
em contato com extensas áreas de matas e manchas de cerradão. A altitude média da reserva é de 250 m, o relevo é plano a suave-ondulado e os solos pre-dominantes são Latossolo Vermelho-Amarelo e Latossolo Amarelo, na maioria distróficos, álicos, profundos, bem drenados e de textura média, com material de origem da Cobertura Sedimentar do Bananal (RADAMBRASIL 1981). O clima é do tipo Aw de Köppen, (Camargo 1963), com precipitação média anual de 1.600 mm (Nimer 1989) e temperatura média anual de 24,4 ºC (Fig. 1).
Foram estabelecidas em cada área 50 parcelas de 10×10 m (0,5 ha) para o levantamento fitossociológico. Foram medidos o diâmetro e a altura
Figura 1. Precipitação e temperaturas médias na Reserva Mário Viana em Nova Xavantina, MT, Brasil. Dados médios de 1995 a 2000 obtidos na Estação Meteorológica da UNEMAT.
0 50 100 150 200 250 300 350
J F M A M J J A S O N D 0 5 10 15 20 25 30 Meses
das árvores, vivas e mortas, com diâmetro mínimo de 5 cm na altura de 30 cm do solo. A diversidade florística das duas comunidades foi calculada pelo índice de Shannon, sendo que a uniformidade (ou equabilidade) foi verificada pelo índice de Pielou. Para avaliar a similaridade florística entre as duas comunidades foi utilizado o índice de Sørensen e de Morisita (Horn 1966).
Para comparação das propriedades do solo, foi aberto um perfil em cada fitofisionomia e retiradas amostras em sete profundidades: 0-15; 15-30; 30-45; 45-70; 70-100; 100-140 e 140-200 cm. Para determinar as variações do solo dentro das fitofisionomias, foram selecionadas 25 parcelas aleatórias em cada área e escolhidos dois pontos aleatórios em cada parcela para coletas de amostras compostas nas profundidades de 0-10 e 10-20 cm. A textura foi determinada através do método de Bouyoucos; o pH foi medido em água e em KCl (1 mol L-1); o teor de N total foi obtido pelo método de Kjeldahl; o teor de matéria orgânica pelo método de Walkey-Black; os teores de Ca, Mg e Al trocáveis foram obtidos em KCl a 1 mol L-1 e os teores de P e K no extrato de Mehlich (H2SO4 a 0,0125 1 mol L-1; HCl a 0,051 mol L-1) (Tan 1996). O alumínio foi determinado por titulação com NaOH, o fósforo por colorimetria utilizando-se molibdato de amônia e ácido ascórbico (3%) e os demais por espectrofotometria de absorção atômica e emissão de chama (Allen 1974).
Resultados e discussão
Florística e fitossociologia – As espécies mais importantes no cerradão foram, por ordem de IVI, H. glandulosa (41,7), Sclerolobium paniculatum (24,2), Xylopia aromatica (20,8), Eriotheca gracilipes (11,5)
e Emmotum nitens (11,4), as quais representaram
36,5% do IVI total. No cerrado sensu stricto foram Qualea parviflora (27,8), Davilla elliptica (19,9), Roupala montana (13,8), Syagrus flexuosa (11,2) e Qualea grandiflora (11,1), representando 28% do IVI
total. Os principais gêneros e espécies amostrados evidenciam o padrão florístico típico observado para cerrado sensu stricto em outras áreas e/ou Estados
(Ribeiro et al. 1985; Furley et al. 1988; Nascimento
& Saddi 1992; Oliveira-Filho et al. 1990; Marimon et al.
1998; Costa & Araújo 2001; P.E. Nogueira, dados não publicados). A composição florística do cerradão enquadrou-se no tipo descrito por Ratter et al. (1973)
como “cerradão de Hirtella glandulosa”, em função
da espécie de maior IVI. A composição de espécies difere da encontrada por Ribeiro & Haridasan (1984)
em um cerradão em Planaltina, DF, onde Emmotum nitens, Ocotea spixiana e Sclerolobium paniculatum
var. rubiginosum foram as espécies mais importantes.
As diferenças são ainda maiores em relação ao cerradão amostrado por Costa & Araújo (2001) em Uberlândia, MG, onde as espécies de maior IVI foram Qualea grandiflora, Vochysia tucanorum e Matayba guianensis.
Foi encontrado o mesmo número de espécies no cerrado sensu stricto e no cerradão (77), sendo que
apenas uma do cerrado (Ficus sp.) não foi identificada.
A curva espécies × área foi semelhante em ambas as comunidades; 87% de todas as espécies do levanta-mento, tanto para o cerrado sensu stricto quanto para
o cerradão, foram encontradas na primeira metade (2.500 m2) das 50 parcelas (Fig. 2). Assim, a área total de amostragem (5.000 m2) foi suficiente para a comparação da composição florística e da estrutura das fitofisionomias.
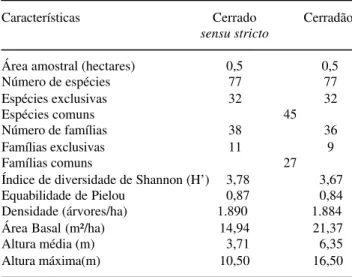
As duas fitofisionomias mostraram-se distintas quanto a composição florística, diâmetro médio, altura e área basal (Tab. 1). O cerradão apresentou indivíduos que atingiram mais de 14 metros de altura, dossel fechado e um padrão tipicamente florestal, com presença de cipós e vegetação graminosa-herbácea muito rara (Fig. 3). A área basal foi 21,4 m² ha-1, sendo que, deste total, 67,3% refere-se às dez espécies mais importantes (Tab. 2). A espécie Hirtella glandulosa,
de maior IVI, participa com 23,6% da área basal total. O cerrado sensu stricto apresentou padrão tipicamente
savânico, com estrato herbáceo-graminoso abundante e espécies que atingiram altura máxima de 7,5 metros (Fig. 4). A área basal foi de 14,9 m² ha-1, sendo 46% referente às dez espécies mais importantes e 11,7% à
Qualea parviflora, espécie de maior IVI (Tab. 3).
0 10 20 30 40 50 60 70 80 90
0 500 1000 1500 2000 2500 3000 3500 4000 4500 5000
Área (m2)
Número de espécies
Figura 2. Curva espécies × área (curva do coletor) do cerrado
sensu stricto e do cerradão na Reserva Mário Viana, Nova
A distribuição dos diâmetros apresentou diferenças inerentes às duas comunidades (Fig. 5). O cerrado
sensu stricto apresentou maior número de indivíduos
com diâmetros menores do que 10 cm, enquanto que
o cerradão apresentou maior número de indivíduos acima de 10 cm de diâmetro. A distribuição dos diâmetros nas duas comunidades apresentou a forma de J-invertido, sendo que mais de 50% dos indivíduos apresentaram-se com diâmetro inferior a 10 cm. Este padrão indica que o cerrado sensu stricto e o cerradão
são fitofisionomias autoregenerativas (Felfili & Silva-Júnior 1988). A distribuição das alturas foi quase unimodal nas duas comunidades (Fig. 6), ajustando-se ao que foi observado por Felfili (1997) para uma distribuição natural.
As áreas basais de ambas as comunidades mostraram-se maiores do que as encontradas por diversos autores no Bioma Cerrado. Felfili et al. (1993),
comparando onze áreas de cerrado sensu stricto das
Chapadas Pratinha e dos Veadeiros, encontraram valores para cerrado sensu stricto que variaram entre
5,8 e 11,3 m²ha-1. Costa & Araújo (2001) encontraram 17,1 m²ha-1 para um cerradão e 9,63 m²ha-1 para um cerrado sensu stricto em Uberlândia-MG, enquanto
que Felfili et al. (2002) encontraram 7,5 m²ha-1 em
um cerrado sensu stricto em Água Boa, MT. A área Tabela 1. Características fitossociológicas do cerrado sensu stricto
na Reserva Mário Viana, Nova Xavantina, MT, Brasil.
Características Cerrado Cerradão
sensu stricto
Área amostral (hectares) 0,5 0,5
Número de espécies 77 77
Espécies exclusivas 32 32
Espécies comuns 45
Número de famílias 38 36
Famílias exclusivas 11 9
Famílias comuns 27
Índice de diversidade de Shannon (H’) 3,78 3,67
Equabilidade de Pielou 0,87 0,84
Densidade (árvores/ha) 1.890 1.884
Área Basal (m²/ha) 14,94 21,37
Altura média (m) 3,71 6,35
Altura máxima(m) 10,50 16,50
Distância (m) 12
8
4
0 5 10 15 20
Altura (m)
Figura 3. Perfil esquemático do cerradão na Reserva Mário Viana, Nova Xavantina, MT, Brasil. Espécies: 1 - Vatairea macrocarpa;
2 - Hirtella glandulosa; 3 - Lutzelburgia praecox; 4 - Syagrus flexuosa; 5 - Erythroxylum daphnites; 6 - Myrcia sellowiana; 7 - Hirtella
Tabela 2. Parâmetros fitossociológicos das espécies arbóreas do cerrado sensu stricto da Reserva Municipal Mário Viana, Nova Xavantina, MT, Brasil. Ni = número de indivíduos; DR = Densidade relativa; FA = Freqüência absoluta; FR = Frqüência relativa; Doa = Dominância absoluta; DoR = dominância relativa; IVI = Índice do Valor de Importância; IVC = Índice do Valor de Cobertura.
Espécies Ni DR FA FR DoA DoR IVC IVI
Hirtella glandulosa Spreng. 112 11,89 41 6,24 5,042 23,59 35,48 41,72
Sclerolobium paniculatum Vog. 61 6,48 31 4,72 2,778 13,00 19,47 24,19
Xylopia aromatica (Lam.) Mart. 71 7,54 38 5,78 1,604 7,51 15,04 20,83
Eriotheca gracilipes (K. Schum.) A. Robyns 31 3,29 21 3,20 1,071 5,01 8,30 11,50
Emmotum nitens (Benth.) Miers. 24 2,55 19 2,89 1,260 5,89 8,44 11,33
Guapira graciliflora (Mart. ex Schmidt) Lund 46 4,88 28 4,26 0,449 2,10 6,99 11,25
Roupala montana Aubl. 39 4,14 25 3,81 0,693 3,25 7,39 11,19
Myrcia sellowiana Berg 41 4,35 25 3,81 0,545 2,55 6,90 10,71
Aspidosperma multiflorum A. DC. 34 3,61 25 3,81 0,545 2,55 6,16 9,97
Matayba guianensis Aubl. 28 2,97 21 3,20 0,388 1,82 4,79 7,99
Hirtella gracilipes (Hook. f.) Prance 29 3,08 20 3,04 0,270 1,26 4,34 7,39
Heisteria ovata Benth. 27 2,87 20 3,04 0,236 1,11 3,97 7,02
Tapirira guianensis Aubl. 19 2,02 12 1,83 0,422 1,98 3,99 5,82
Mezilaurus crassiramea (Meissn.) Taub. 11 1,17 10 1,52 0,620 2,90 4,07 5,59
Vatairea macrocarpa (Benth.) Ducke 19 2,02 15 2,28 0,249 1,17 3,18 5,47
Guapira noxia (Netto) Lund 19 2,02 12 1,83 0,265 1,24 3,26 5,08
Pseudobombax longiflorum (Mart. & Zucc.) A. Robyns 15 1,59 12 1,83 0,323 1,51 3,11 4,93
Erythroxylum daphnites Mart. 17 1,80 14 2,13 0,122 0,57 2,38 4,51
Protium heptaphyllum (Aubl.) March. 14 1,49 12 1,83 0,224 1,05 2,54 4,36
Alibertia elliptica (Cham.) K. Schum. 15 1,59 13 1,98 0,119 0,56 2,15 4,13
Antonia ovata Pohl 14 1,49 12 1,83 0,102 0,48 1,96 3,79
Luetzelburgia praecox (Harms.) Harms. 10 1,06 9 1,37 0,287 1,34 2,40 3,77
Sorocea klotzchiana Baill 13 1,38 12 1,83 0,093 0,44 1,82 3,65
Terminalia argentea Mart. & Zucc. 10 1,06 9 1,37 0,241 1,13 2,19 3,56
Syagrus flexuosa L. 15 1,59 10 1,52 0,089 0,42 2,01 3,53
Alchornea schomburgkii Klotz 12 1,27 11 1,67 0,116 0,55 1,82 3,49
Agonandra brasiliensis Benth. & Hook. f. 9 0,96 6 0,91 0,185 0,87 1,82 2,74
Ouratea spectabilis (Mart.) Engl. 9 0,96 9 1,37 0,052 0,25 1,20 2,57
Pterodon pubescens (Benth.) Benth. 5 0,53 4 0,61 0,299 1,40 1,93 2,54
Annona coriacea Mart. 9 0,96 7 1,07 0,107 0,50 1,46 2,52
Curatella americana L. 5 0,53 5 0,76 0,227 1,06 1,59 2,36
Aspidosperma macrocarpon Mart. 7 0,74 7 1,07 0,113 0,53 1,27 2,34
Salvertia convallariaeodora St. Hil. 9 0,96 7 1,07 0,063 0,30 1,25 2,32
Alibertia edulis (L.C. Rich.) A. Rich. ex DC. 7 0,74 7 1,07 0,048 0,22 0,97 2,03
Qualea parviflora Mart. 7 0,74 6 0,91 0,072 0,34 1,08 2,00
Coccoloba mollis Casar 8 0,85 5 0,76 0,080 0,38 1,23 1,99
Qualea multiflora Mart. 6 0,64 6 0,91 0,086 0,40 1,04 1,95
Syagrus comosa (Mart.) Becc. 7 0,74 5 0,76 0,095 0,45 1,19 1,95
Maprounea guianensis (Aubl.) M. Arg. 6 0,64 6 0,91 0,067 0,31 0,95 1,86
Dipteryx alata Vog. 4 0,42 4 0,61 0,174 0,82 1,24 1,85
Siparuna guianensis Aubl. 6 0,64 6 0,91 0,062 0,29 0,93 1,84
Euplassa inaequalis (Pohl) Engl. 5 0,53 5 0,76 0,111 0,52 1,05 1,81
Dimorphandra mollis Benth. 6 0,64 5 0,76 0,084 0,40 1,03 1,79
Qualea grandiflora Mart. 6 0,64 6 0,91 0,051 0,24 0,88 1,79
Vochysia rufa Mart. 5 0,53 5 0,76 0,091 0,43 0,96 1,72
Lacistema aggregatum (Berg.) Rusby 6 0,64 6 0,91 0,028 0,13 0,77 1,68
Astronium fraxinifolium Schott. 4 0,42 4 0,61 0,094 0,44 0,87 1,48
Strychnos pseudoquina A. St.-Hil. 2 0,21 2 0,30 0,204 0,96 1,17 1,47
Eugenia geminiflora O. Berg 5 0,53 5 0,76 0,037 0,18 0,71 1,47
Machaerium acutifolium Vog. 4 0,42 4 0,61 0,037 0,17 0,60 1,21
Styrax camporum Pohl 3 0,32 3 0,46 0,065 0,31 0,63 1,08
Vochysia haenkeana Mart. 2 0,21 2 0,30 0,092 0,43 0,64 0,95
Tabebuia aurea (Manso) Benth. & Hook. f. ex S. Moore 3 0,32 3 0,46 0,032 0,15 0,47 0,93
Brosimum gaudichaudii Tréc. 3 0,32 3 0,46 0,031 0,15 0,47 0,92
Andira vermifuga Mart. ex Benth. 3 0,32 3 0,46 0,022 0,11 0,42 0,88
basal por hectare do cerradão foi cerca de 1,4 vezes superior à do cerrado sensu stricto e a altura média em
torno de 1,7 vezes, o que sugere maior biomassa do cerradão em relação ao cerrado sensu stricto. A
participação de H. glandulosa no total dessa biomassa
é duas vezes maior do que a participação de
Q. parviflora no total do cerrado sensu stricto. Essas
diferenças evidenciam um padrão estrutural distinto
Tabela 2 (continuação)
Espécies Ni DR FA FR DoA DoR IVC IVI
Byrsonima coccolobifolia H.B. & K. 2 0,21 2 0,30 0,062 0,29 0,51 0,81
Hymenaea stigonocarpa Mart. ex Hayne 3 0,32 2 0,30 0,023 0,11 0,43 0,73
Tetrapterys glabra (Spreng.) Griseb 2 0,21 2 0,30 0,037 0,17 0,39 0,69
Copaifera langsdorffii Desf. 2 0,21 2 0,30 0,025 0,12 0,33 0,63
Dalbergia miscolobium Benth. 2 0,21 2 0,30 0,024 0,11 0,33 0,63
Bowdichia virgilioides H.B. & K. 2 0,21 2 0,30 0,022 0,11 0,32 0,62
Plathymenia reticulata Benth. 2 0,21 2 0,30 0,015 0,07 0,28 0,59
Couepia grandiflora (Mart. & Zucc.) Benth. & Hook. f. 2 0,21 2 0,30 0,014 0,07 0,28 0,59
Tapura amazonica Poepp. & Endl. 2 0,21 2 0,30 0,014 0,07 0,28 0,58
Mimosa laticifera Rizzini & Mattos 2 0,21 2 0,30 0,011 0,05 0,27 0,57
Cardiopetalum calophyllum Schlecht. 2 0,21 2 0,30 0,010 0,05 0,26 0,57
Ficus sp. 2 0,21 2 0,30 0,009 0,04 0,26 0,56
Platypodium elegans Vog. 1 0,11 1 0,15 0,056 0,26 0,37 0,52
Simarouba amara Aubl. 1 0,11 1 0,15 0,049 0,23 0,34 0,49
Magonia pubescens A. St.-Hil. 1 0,11 1 0,15 0,032 0,15 0,26 0,41
Andira cuiabensis Benth. 1 0,11 1 0,15 0,022 0,11 0,21 0,37
Cybistax antisiphilitica (Mart.) Mart. 1 0,11 1 0,15 0,019 0,09 0,20 0,35
Peltogyne confertiflora (Hayne) Benth. 1 0,11 1 0,15 0,011 0,06 0,16 0,31
Miconia albicans (Sw.) Triana 1 0,11 1 0,15 0,007 0,04 0,14 0,29
Byrsonima basiloba A. Juss. 1 0,11 1 0,15 0,005 0,03 0,13 0,29
Myrcia camapuanensis N.F.E. Silveira 1 0,11 1 0,15 0,005 0,03 0,13 0,28
Aspidosperma nobile M. Arg. 1 0,11 1 0,15 0,004 0,02 0,13 0,28
942 100 1.314 100 21,377 100 200 300
Figura 4. Perfil esquemático do cerrado sensu stricto na Reserva Mário Viana, Nova Xavantina, MT, Brasil. Espécies: 1 - Euplassa
inaequalis; 2 - Qualea multiflora; 3 - Curatella americana; 4 - Davilla elliptica; 5 - Hymenaea stigonocarpa; 6 - Myrcia lanuginosa;
7 - Emmotun nitens; 8 - Guapira noxia; 9 - Magonia pubescens; 10 - Roupala montana; 11 - Lafoensia pacari; 12 - Aspidosperma
tomentosum; 13 - Qualea grandiflora; 14 - Erythroxylum suberosum; 15 - Caryocar brasiliense.
Distância (m) 6
8
4
0 5 10 15 20
Altura (m)
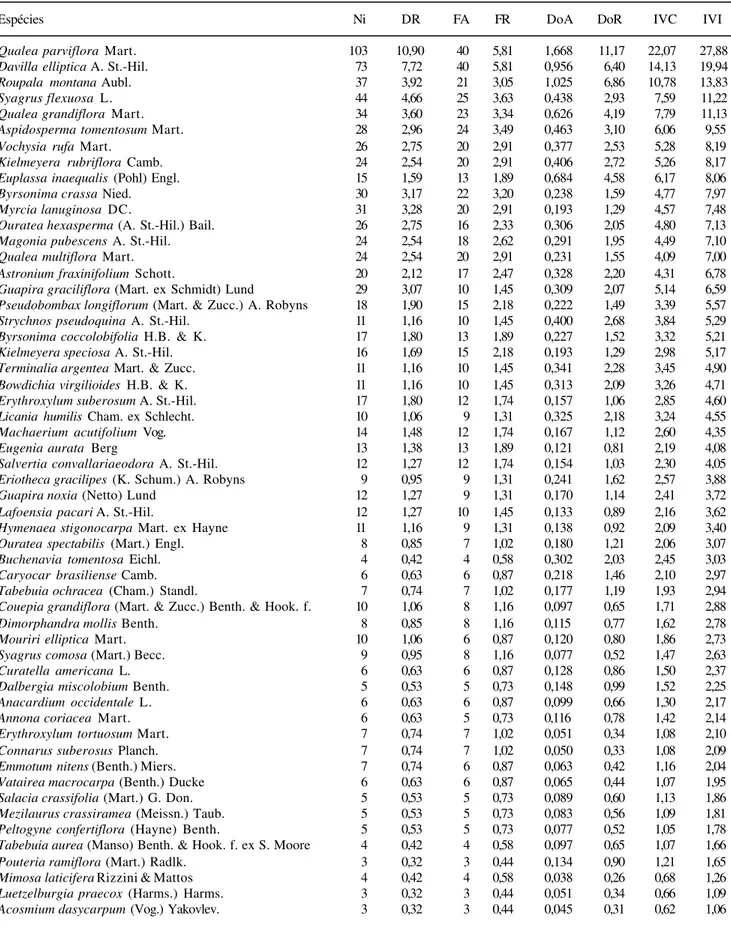
Tabela 3. Parâmetros fitossociológicos das espécies arbóreas do cerradão da Reserva Municipal Mário Viana, Nova Xavantina, MT, Brasil. Ni = número de indivíduos; DR = Densidade relativa; FA = Freqüência absoluta; FR = Frqüência relativa; Doa = Dominância absoluta; DoR = dominância relativa; IVI = Índice do Valor de Importância; IVC = Índice do Valor de Cobertura.
Espécies Ni DR FA FR DoA DoR IVC IVI
Qualea parviflora Mart. 103 10,90 40 5,81 1,668 11,17 22,07 27,88
Davilla elliptica A. St.-Hil. 73 7,72 40 5,81 0,956 6,40 14,13 19,94
Roupala montana Aubl. 37 3,92 21 3,05 1,025 6,86 10,78 13,83
Syagrus flexuosa L. 44 4,66 25 3,63 0,438 2,93 7,59 11,22
Qualea grandiflora Mart. 34 3,60 23 3,34 0,626 4,19 7,79 11,13
Aspidosperma tomentosum Mart. 28 2,96 24 3,49 0,463 3,10 6,06 9,55
Vochysia rufa Mart. 26 2,75 20 2,91 0,377 2,53 5,28 8,19
Kielmeyera rubriflora Camb. 24 2,54 20 2,91 0,406 2,72 5,26 8,17
Euplassa inaequalis (Pohl) Engl. 15 1,59 13 1,89 0,684 4,58 6,17 8,06
Byrsonima crassa Nied. 30 3,17 22 3,20 0,238 1,59 4,77 7,97
Myrcia lanuginosa DC. 31 3,28 20 2,91 0,193 1,29 4,57 7,48
Ouratea hexasperma (A. St.-Hil.) Bail. 26 2,75 16 2,33 0,306 2,05 4,80 7,13
Magonia pubescens A. St.-Hil. 24 2,54 18 2,62 0,291 1,95 4,49 7,10
Qualea multiflora Mart. 24 2,54 20 2,91 0,231 1,55 4,09 7,00
Astronium fraxinifolium Schott. 20 2,12 17 2,47 0,328 2,20 4,31 6,78
Guapira graciliflora (Mart. ex Schmidt) Lund 29 3,07 10 1,45 0,309 2,07 5,14 6,59
Pseudobombax longiflorum (Mart. & Zucc.) A. Robyns 18 1,90 15 2,18 0,222 1,49 3,39 5,57
Strychnos pseudoquina A. St.-Hil. 11 1,16 10 1,45 0,400 2,68 3,84 5,29
Byrsonima coccolobifolia H.B. & K. 17 1,80 13 1,89 0,227 1,52 3,32 5,21
Kielmeyera speciosa A. St.-Hil. 16 1,69 15 2,18 0,193 1,29 2,98 5,17
Terminalia argentea Mart. & Zucc. 11 1,16 10 1,45 0,341 2,28 3,45 4,90
Bowdichia virgilioides H.B. & K. 11 1,16 10 1,45 0,313 2,09 3,26 4,71
Erythroxylum suberosum A. St.-Hil. 17 1,80 12 1,74 0,157 1,06 2,85 4,60
Licania humilis Cham. ex Schlecht. 10 1,06 9 1,31 0,325 2,18 3,24 4,55
Machaerium acutifolium Vog. 14 1,48 12 1,74 0,167 1,12 2,60 4,35
Eugenia aurata Berg 13 1,38 13 1,89 0,121 0,81 2,19 4,08
Salvertia convallariaeodora A. St.-Hil. 12 1,27 12 1,74 0,154 1,03 2,30 4,05
Eriotheca gracilipes (K. Schum.) A. Robyns 9 0,95 9 1,31 0,241 1,62 2,57 3,88
Guapira noxia (Netto) Lund 12 1,27 9 1,31 0,170 1,14 2,41 3,72
Lafoensia pacari A. St.-Hil. 12 1,27 10 1,45 0,133 0,89 2,16 3,62
Hymenaea stigonocarpa Mart. ex Hayne 11 1,16 9 1,31 0,138 0,92 2,09 3,40
Ouratea spectabilis (Mart.) Engl. 8 0,85 7 1,02 0,180 1,21 2,06 3,07
Buchenavia tomentosa Eichl. 4 0,42 4 0,58 0,302 2,03 2,45 3,03
Caryocar brasiliense Camb. 6 0,63 6 0,87 0,218 1,46 2,10 2,97
Tabebuia ochracea (Cham.) Standl. 7 0,74 7 1,02 0,177 1,19 1,93 2,94
Couepia grandiflora (Mart. & Zucc.) Benth. & Hook. f. 10 1,06 8 1,16 0,097 0,65 1,71 2,88
Dimorphandra mollis Benth. 8 0,85 8 1,16 0,115 0,77 1,62 2,78
Mouriri elliptica Mart. 10 1,06 6 0,87 0,120 0,80 1,86 2,73
Syagrus comosa (Mart.) Becc. 9 0,95 8 1,16 0,077 0,52 1,47 2,63
Curatella americana L. 6 0,63 6 0,87 0,128 0,86 1,50 2,37
Dalbergia miscolobium Benth. 5 0,53 5 0,73 0,148 0,99 1,52 2,25
Anacardium occidentale L. 6 0,63 6 0,87 0,099 0,66 1,30 2,17
Annona coriacea Mart. 6 0,63 5 0,73 0,116 0,78 1,42 2,14
Erythroxylum tortuosum Mart. 7 0,74 7 1,02 0,051 0,34 1,08 2,10
Connarus suberosus Planch. 7 0,74 7 1,02 0,050 0,33 1,08 2,09
Emmotum nitens (Benth.) Miers. 7 0,74 6 0,87 0,063 0,42 1,16 2,04
Vatairea macrocarpa (Benth.) Ducke 6 0,63 6 0,87 0,065 0,44 1,07 1,95
Salacia crassifolia (Mart.) G. Don. 5 0,53 5 0,73 0,089 0,60 1,13 1,86
Mezilaurus crassiramea (Meissn.) Taub. 5 0,53 5 0,73 0,083 0,56 1,09 1,81
Peltogyne confertiflora (Hayne) Benth. 5 0,53 5 0,73 0,077 0,52 1,05 1,78
Tabebuia aurea (Manso) Benth. & Hook. f. ex S. Moore 4 0,42 4 0,58 0,097 0,65 1,07 1,66
Pouteria ramiflora (Mart.) Radlk. 3 0,32 3 0,44 0,134 0,90 1,21 1,65
Mimosa laticifera Rizzini & Mattos 4 0,42 4 0,58 0,038 0,26 0,68 1,26
Luetzelburgia praecox (Harms.) Harms. 3 0,32 3 0,44 0,051 0,34 0,66 1,09
Acosmium dasycarpum (Vog.) Yakovlev. 3 0,32 3 0,44 0,045 0,31 0,62 1,06
Tabela 3 (continuação)
Espécies Ni DR FA FR DoA DoR IVC IVI
Sclerolobium aureum (Tul.) Benth. 3 0,32 3 0,44 0,033 0,22 0,54 0,98
Callisthene fasciculata Mart. 3 0,32 3 0,44 0,031 0,21 0,53 0,97
Rourea induta Planch. 3 0,32 3 0,44 0,017 0,12 0,43 0,87
Xylopia aromatica (Lam.) Mart. 2 0,21 2 0,29 0,052 0,35 0,56 0,85
Myrcia camapuanensis N.F.E. Silveira 3 0,32 2 0,29 0,024 0,16 0,48 0,77
Aspidosperma multiflorum A. DC. 2 0,21 2 0,29 0,031 0,21 0,42 0,71
Dipteryx alata Vog. 2 0,21 2 0,29 0,025 0,17 0,38 0,67
Casearia sylvestris Sw. 2 0,21 2 0,29 0,017 0,11 0,33 0,62
Simarouba versicolor A. St.-Hil. 2 0,21 2 0,29 0,017 0,11 0,33 0,62
Vernonia ferruginea Less. 2 0,21 2 0,29 0,013 0,09 0,30 0,59
Himatanthus obovatus (M. Arg.) R.E. Woodson 2 0,21 2 0,29 0,010 0,07 0,28 0,57
Pterodon pubescens (Benth.) Benth. 1 0,11 1 0,15 0,046 0,31 0,42 0,56
Antonia ovata Pohl 2 0,21 2 0,29 0,008 0,05 0,27 0,56
Heisteria ovata Benth. 1 0,11 1 0,15 0,045 0,30 0,41 0,56
Myrcia sellowiana Berg 2 0,21 1 0,15 0,008 0,06 0,27 0,42
Styrax camporum Pohl 1 0,11 1 0,15 0,017 0,12 0,22 0,37
Austroplenckia populnea Reiss. 1 0,11 1 0,15 0,015 0,11 0,21 0,36
Andira cuiabensis Benth. 1 0,11 1 0,15 0,013 0,09 0,19 0,34
Myrtaceae-NI 1 0,11 1 0,15 0,011 0,08 0,19 0,33
Tocoyena formosa (Cham. & Schlecht.) K. Schum. 1 0,11 1 0,15 0,009 0,06 0,17 0,31
Palicourea rigida Kunth 1 0,11 1 0,15 0,005 0,04 0,14 0,29
Brosimum gaudichaudii Tréc. 1 0,11 1 0,15 0,005 0,04 0,14 0,29
945 100 688 100 14,945 100 200 300
Figura 5. Distribuição de diâmetros de árvores em um cerrado sensu stricto e um cerradão na Reserva Mário Viana, Nova Xavantina, MT, Brasil. –¯– Cerrado sensu stricto; –u– = Cerrado.
5--| 8 8--| 11 11--| 14--| 17 17--| 20 20--| 23 23-| 26 26--| 29 29--|32 32--| 35 35--| 38 38--| 54 500
Classes de diâmetro (cm)
Número de indivíduos
450
400
350
300
250
200
150
100
50
0
5--|8 8--|11 11--|14 14--|17 17--|20 20--|23 23--|26 26--|29 29--|32 32--|35 35--|38 38--|54 entre as duas comunidades e apontam para uma
dominância mais marcante de H. glandulosa no
cerradão do que Q. parviflora no cerrado sensu stricto.
O índice de similaridade de Sørensen (0,58) e Morisita (0,21) indicam baixa similaridade florística, reforçando as diferenças também nesse aspecto. Os valores de
similaridade ficaram dentro de uma faixa de variação encontrada por diversos autores (Costa & Araújo 2001; Ribeiro et al. 1985; Felfili et al. 1993).
8,5% da densidade total. Vinte e três espécies apresentaram apenas um ou dois indivíduos, representando cerca de 30% do total de espécies e cerca de 4% do total de indivíduos. As espécies de maior freqüência foram H. glandulosa e Xylopia aromatica, que ocorreram em 82% e 76% das
parcelas, respectivamente, sendo que dez espécies ocorreram em apenas uma parcela. No cerrado sensu stricto foram amostrados 945 indivíduos pertencentes
a 77 espécies, 68 gêneros e 38 famílias. Os indivíduos mortos (em pé) perfizeram 11% da densidade total. As espécies raras, com apenas um ou dois indivíduos, representaram 23% do total de espécies e 3% da densidade total. Q. parviflora e Davilla elliptica
foram as espécies mais freqüentes, ocorrendo em 80% das parcelas, sendo que 10 espécies ocorreram em apenas uma parcela. O índice de diversidade de espécies de Shannon (H’) e a equabilidade de Pielou (J) para o cerrado sensu stricto (Tab. 1) foram,
respectivamente, 3,78 e 0,870. Para o cerradão, os mesmo índices foram 3,67 e 0,846.
De acordo com Ratter et al. (1977), a estrutura
fitossociológica de um cerradão pode variar em função do tipo de solo. Os autores mencionaram que cerradão sobre solo mesotrófico é caracterizado por Magonia pubescens, Luehea paniculata, Physocalymma scaberrimum e Callisthene fasciculata, entre outras,
enquanto o cerradão indiferenciado apresenta Qualea grandiflora e Vochysia tucanorum como espécies
principais. Já no caso do cerradão sobre solos distróficos, as principais espécies em IVI são Hirtella
glandulosa, Emmotum nitens, Sclerolobium paniculatum e Vochysia haenkeana. Ribeiro &
Haridasan (1984) encontraram Emmotum nitens, Ocotea spixiana e Sclerolobium paniculatum var. rubiginosum como as espécies mais importantes em
um cerradão sobre solos distróficos no Distrito Federal. No presente estudo, somente V. haenkeana não
estava entre as de maior IVI.
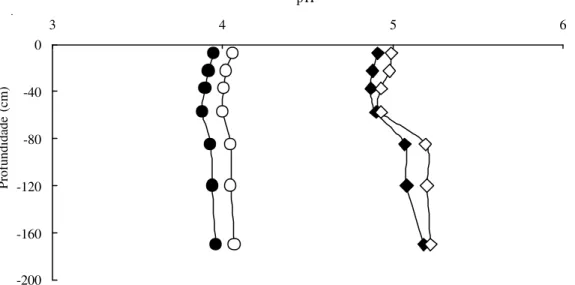
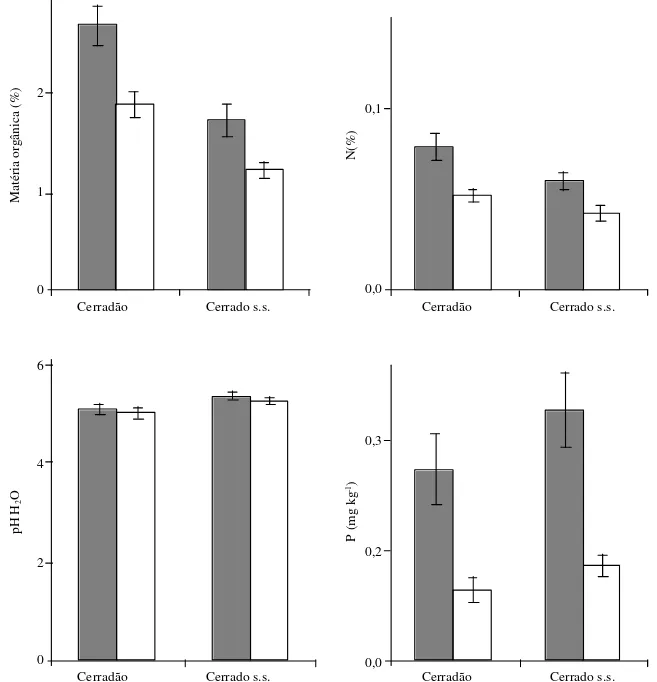
Solos – O solo sob ambas fitofisionomias apresentou-se distrófico (Ca < 0,4 cmolc kg-1), álico (Al > 1,3 cmolc kg-1) e ácido (pH < 5,0) ao longo do perfil (Fig. 7, 8). Os resultados de análises dos dois perfis do solo foram confirmados pelas análises das 25 amostras superficiais (Fig. 9, 10). O Al trocável mostrou-se um pouco mais alto ao longo do perfil no cerradão, com valor máximo de aproximadamente 2,6 cmolc kg-1 contra 1,4 cmolc kg-1 no perfil do cerrado sensu stricto. Em relação aos teores de cálcio e magnésio no perfil do solo, as diferenças entre as áreas não foram significativas, sendo inferiores a 0,02 cmolc kg-1 e 0,102 cmolc kg-1 (Fig. 7), respectivamente, além do pH, que apresentou pequenas diferenças em água (<0,3) e em KCl (<0,14).
Os teores de macronutrientes, alumínio, os percentuais de matéria orgânica, carbono e nitrogênio, bem como os valores de pH em água e em KCl encontrados no solo sob as duas comunidades estudadas apresentaram-se muito próximos aos encontrados em vários outros solos distróficos para vegetação de cerrado sensu stricto e cerradão em diferentes regiões
Figura 6. Distribuição de alturas de árvores em um cerrado sensu stricto e um cerradão na Reserva Mário Viana, Nova Xavantina, MT, Brasil. –¯– Cerrado sensu stricto; –u– = Cerrado.
500
450
400
350
300
250
200
150
100
50
0
Classes de altura (m)
Número de indivíduos
-200 -160 -120 -80 -40 0
0,0 0,5 1,0 1,5 2,0 2,5 3,0
Figura 7. Cátions trocáveis no solo sob o cerradão e o cerrado sensu stricto na Reserva Mário Viana, Nova Xavantina, MT, Brasil. –u– = K - cerradão; –¯– = K - cerrado; –˜– = Ca - cerradão; –š– = Ca - cerrado; –p– = Mg - cerradão; –r– = Mg - cerrado; –¢– = Al cerradão; ––£– = Al - cerrado.
cmolc kg-1
Profundidade (cm)
-200 -160 -120 -80 -40 0
3 4 5 6
Figura 8. pH no solo sob o cerradão e o cerrado sensu stricto na Reserva Mário Viana, Nova Xavantina, MT, Brasil. –u– = pH em água
- cerradão; –˜– = pH em KCl - cerradão; –r– = pH em água cerrado; –š– = pH em KCl - cerrado. pH
Profundidade (cm)
do Bioma Cerrado (J.F. Ribeiro, dados não publicados; Ribeiro & Haridasan 1984; Haridasan 1987; Furley & Ratter 1988; Furley et al. 1988; Haridasan 1992; Sassaki et al. 1999; Haridasan 2000; Costa & Araújo 2001;
Ruggiero et al. 2002).
Como os valores dos macronutrientes, alumínio e pH do solo não diferiram entre as áreas, tanto nos perfis quanto nas amostras superficiais, o presente estudo obteve resultados distintos daqueles verificados por
Alvim & Araújo (1952), Goodland (1971b), Goodland & Pollard (1973) e Lopes & Cox (1977) em outras localidades do Bioma Cerrado, onde os autores concluíram por uma maior fertilidade nos solos sob cerradão. Nossos resultados assemelham-se mais aos de Askew et al. (1971), Gibbs et al. (1983), J.F. Ribeiro
Figura 9. Características das camadas superficiais do solo sob o cerrado sensu stricto e o cerradão na Reserva Mário Viana, Nova Xavantina, MT, Brasil. As barras representam o desvio padrão da média (25 amostras). ¢ = 0 a 10 cm; £ = 10 a 20 cm.
Como o cerradão apresenta maior biomassa que o cerrado sensu stricto, a quantidade maior de nutrientes
requerida para a sua manutenção pode ter existido no solo no passado, mantendo-se até o presente através da ciclagem de nutrientes (Haridasan 1987; 199; 2000). Desta forma, não obstante outros fatores, como presença de fogo, diferenças genéticas e/ou possíveis diferenças no regime hídrico do solo em função de sua textura, é provável que o cerradão do presente estudo permaneça confinado em seu sítio atual por não existir quantidade suficiente de nutrientes nas áreas adjacentes
para que esta vegetação possa sobstituir o cerrado sensu stricto.
Originalmente, o cerrado sensu stricto era a
vegetação predominante na paisagem da região leste de Mato Grosso e o cerradão ocorria apenas em manchas esparsas e geralmente nas áreas próximas do contato entre cerrado e floresta de transição pré-amazônica. O solo sob estas manchas de cerradão pode ter sido originado, pelo menos em parte, de rochas calcáreas, cujos afloramentos são muito comuns na região. Esta poderia ter sido a origem de sítios um
3
2
1
0
0,1
0,0
6
4
2
0
0,3
0,2
0,0
Cerradão Cerrado s.s. Cerradão Cerrado s.s.
Cerradão Cerrado s.s. Cerradão Cerrado s.s.
pH H
2
O
Matéria orgânica (%)
P (mg kg
-1)
Figura 10. Cátions trocáveis nas camadas superficiais do solo sob o cerrado sensu stricto e o cerradão na Reserva Mário Viana, Nova Xavantina, MT, Brasil. As barras representam o desvio padrão da média (25 amostras). ¢ = 0 a 10 cm; £ = 10 a 20 cm.
pouco mais ricos em nutrientes, o suficiente para permitir o estabelecimento dos cerradões em detrimento do cerrado sensu stricto no passado.
Entre as duas fitofisionomias, as variações relevantes foram relativas à composição granulométrica dos solos. O solo sob o cerradão apresentou-se consideravelmente mais argiloso ao longo de todo o perfil. Entre 0-20 cm de profundidade, o teor de argila foi de aproximadamente 28%, aumentando para mais de 40% entre 45-200 cm, contra 18% entre 0-20 cm e
28% entre 45 e 200 cm no cerrado sensu stricto
(Fig. 11).
Como a textura foi o único fator edáfico que apresentou diferenças entre as áreas, sugerimos que sejam conduzidos estudos para a verificação do comportamento da água em ambos os solos e as relações hídricas das principais espécies arbóreas. Além disso, seria importante a investigação sobre o material de origem dos solos nessas fitofisionomias, o que poderia indicar a história geológica de ambos e o
0,1
0,0
0,4
0,2
0,0
0,6
0,3
0,0
1,2
0,6
0,0
Cerradão Cerrado s.s.
Mg (cmol
c
kg
-1)
Matéria orgânica (%)
Al (cmol
c
kg
-1)
Ca (cmol
c
kg
-1)
Cerradão Cerrado s.s.
papel desta no fornecimento diferencial de nutrientes no passado.
Conclui-se que a hipótese de que o solo sob cerradão seria mais fértil, com menores teores de alumínio trocável, não foi confirmada. Uma das explicações plausíveis em relação à existência das duas fitofisionomias está associada à composição granulomé-trica dos solos. Presume-se que o solo sob cerradão, mais argiloso, tem maior capacidade de retenção de água disponível. Portanto, é capaz de suprir mais adequadamente as plantas com água nos processos de síntese de biomassa, bem como possui maior fertilidade ativa, pois a disponibilidade de água regula a dinâmica dos nutrientes no solo e sua absorção pelas plantas.
Referências bibliográficas
Allen, S.E. 1974. Chemical analysis of ecological materials. Oxford, Blackwell Scientific Publications.
Alvin, P.T. & Araújo, W.A. 1952. El suelo como factor ecológico en el desarrollo de la vegetación en el centro-oeste del Brasil. Turrialba 2(4): 153-160.
Araújo, G.M. & Haridasan, M. 1988. A comparison of the nutrients status of two forests on dystrophic and mesotrophic soils in the cerrado region of central Brazil. Communications in Soil Science and plant analysis 19: 1075-1089.
Askew, K.P.; Montgomery, R.F. & Searl, P.L. 1971. Soil and soil moisture as factors influencing the distribution of the vegetation formations of the Serra do Roncador, Mato Grosso. Pp.150-160. In: M.G. Ferri (ed.). III Simpósio sobre o Cerrado). São Paulo, Edgard Blücher.
Camargo, A.P. 1963. Clima do cerrado. Pp.75-59. In: M.G. Ferri (coord.). Simpósio sobre o Cerrado. São Paulo, EDUSP. Costa, A.A. & Araújo, G.M. 2001. Comparação da vegetação
arbórea de cerradão e cerrado na Reserva do Panga, Uberlândia, Minas Gerais. Acta Botanica Brasilica 15(1): 63-72.
Eiten, G. 1972. The cerrado vegetation of Brazil. The Botanical Review 38(2): 201-338.
Eiten, G. 1994. Vegetação do Cerrado. Pp. 17-75. In: M.N. Pinto (org.). Cerrado: caracterização, ocupação e perspectivas. Brasília, Editora da UnB.
Felfili, J.M. & Silva-Júnior, M.C. 1988. Distribuição dos diâmetros numa faixa de cerrado na Fazenda Água Limpa (FAL) em Brasília-DF. Acta Botanica Brasilica 2: 85-104. Felfili, J.M.; Silva-Júnior, M.C.; Rezende, A.V.; Machado, J.W.B.; Walter, B.M.T.; Nogueira, P.E. & Hay, J.D. 1993. Análise comparativa da florística e fitossociologia da vegetação arbórea do cerrado sensu stricto na Chapada pratinha, DF-Brasil. Acta Botanica Brasilica 6: 27-46. Felfili, J.M. 1997. Diameter and height distributions in a
gallery forest tree community and some of its main species in central Brazil over a six-year period (1985-1991). Revista Brasileira de Botânica 20: 155-162.
Felfili, J.M.; Nogueira, P.E.; Silva-Júnior, M.C.; Marimon, B.S. & Delitti, W.B.C. 2002. Composição florística e fitossociologia do cerrado sentido restrito no município de Água Boa-MT. Acta Botanica Brasilica 16(1): 103-112.
Furley, P.A.; Ratter, J.A. & Gifford, D.R. 1988. Observations on the vegetation of esatern Mato Grosso, Brazil III. The woody vegetation and soils of the Morro da Fumaça, Torixoréu. Phil. Trans. R. Soc. Lond. B 235: 259-280.
Furley, P.A & Ratter, J.A. 1988. Soil resources and plant communities of the central brazilian cerrado and their development. Journal of Biogeography 15: 97-108.
-200 -160 -120 -80 -40 0
0 10 20 30 40 50 60 70 80 90
cmolc kg-1
Profundidade (cm)
Figura 11. Textura do solo sob o cerradão e o cerrado sensu stricto na Reserva Mário Viana, Nova Xavantina, MT, Brasil. –u– = Argila
Gibbs, P.E.; Leitão-Filho, H.F. & Shepherd, G. 1983. Floristic composition and community structure in an area of cerrado in SE Brazil. Flora 173: 433-449.
Goodland, R. 1971a. The cerrado oxisols of the Triângulo Mineiro, central Brazil. Anais da Academia Brasileira de Ciências 43: 407-414.
Goodland, R. 1971b. A physiognomic analysis of the cerrado vegetation of central Brazil. Journal of Ecology 59: 411-419. Goodland, R. & Pollard, R. 1973. The Brazilian cerrado vegetation: a fertility gradient. Journal of Ecology 61: 219-224.
Haridasan, M. 1987. Distribution and mineral nutrition of aluminium-accumulating species in different plant communities of the cerrado region of the central Brazil. Pp. 309-348. In: J.J. San Jose & R. Montes (eds.). La Capacidad Bioreprodutiva de Sabanas. IVIC, Caracas, Venezuela.
Haridasan, M. 1992. Observations on soils, foliar nutrient concentration and floristic composition of cerrado sensu stricto and cerradão communities in central Brazil. Pp.171-184. In: P.A. Furley; J. Proctor & J.A. Ratter (eds.). Nature and Dynamics of Forest-Savanna Boundaries. London, Chapman & Hall Publishing.
Haridasan, M. 2000. Nutrição mineral das plantas nativas do Cerrado. Revista Brasileira de Fisiologia Vegetal 12(1): 54-64.
Horn, H.S. 1966. Measurement of “overlap” in comparative ecological studies. The American Naturalist 100: 419-424. Lopes, A.S. & Cox, F.R. 1977. Cerrado vegetation in Brazil: an
edaphic gradient. Agronomy Journal 69: 828-831. Marimon, B.S.; Varella, R.F. & Marimon-Júnior, B.H. 1998.
Fitossociologia de uma área de cerrado de encosta em Nova Xavantina, Mato Grosso. Boletim do Herbário Ezechias Paulo Heringer 3: 82-101.
Nascimento, M.T. & Saddi, N. 1992. Structure and floristic composition in an area of cerrado in Cuiabá-MT, Brazil. Revista Brasileira de Botânica 15: 47-55.
Nimer, E. 1989. Climatologia do Brasil. Rio de Janeiro, Instituto Brasileiro de Geografia e Estatística.
Oliveira-Filho, A.T.; Shepherd, G.J.; Martins, F.R. & Stubblebine, W.H. 1990. Environmental factors affecting physiognomic and floristic variation in an area of cerrado in central Brazil. Journal of Tropical Ecology 5: 413-451.
RADAMBRASIL. 1981. Levantamento de recursos naturais. Rio de Janeiro, Ministério das Minas e Energia 25, folha SD-22/Goiás.
Ratter, J.A. 1971. Some notes on two types of cerradão occuring in northeastern Mato Grosso. Pp.110-112. In: M.G. Ferri (ed.). III Simpósio Sobre o Cerrado. São Paulo, EDUSP/ Edgard Blücher.
Ratter, J.A.; Richards, P.W.; Argent, G. & Gifford, D.R. 1973. Observations on the vegetationof the northeastern Mato Grosso I. The woody vegetation types of the Xavantina-Cachimbo Expedition area. Phil. Trans. R. Soc. Lond. B 266:449-492.
Ratter, J.A.; Askew, G.P.; Montgomery, R.F. & Gifford, D.R. 1977. Observações adicionais sobre o cerradão de solos mesotróficos no Brasil central. Pp.303-316. In: M.G. Ferri (ed.). IV Simpósio sobre o Cerrado: Bases para a Utilização Agropecuária. São Paulo, EDUSP.
Ribeiro, J.F. & Haridasan, M. 1984. Comparação fitossociológica de um cerrado denso e um cerradão em solos distróficos no Distrito Federal. In: Anais do 35o Congresso Nacional de
Botânica. SBB, Manaus.
Ribeiro, J.F.; Silva, J.C.S. & Batmanian, G.J. 1985. Fitossociologia de tipos fisionômicos de cerrado em planaltina-DF. Revista Brasileira de Botânica 8: 131-142.
Ruggiero, P.G.C.; Batalha, M.A.; Pivelo, V.R. & Meireles, S.T. 2002. Soil-vegetation relationships in cerrado (Brazilian savanna) and semideciduous forest, Southeastern Brazil. Plant Ecology 160: 1-16.
Sassaki, R.M.; Rondon, J.N.; Zaidan, L.B.P. & Fellipe, G.M. 1999. Number of buried seeds and sedlings emergence in cerradão, cerrado and gallery forest soils at Pedregulho, Itirapina (SP), Brazil. Revista Brasileira de Botânica 22(2): 147-152.
Tan, K.H. 1996. Soil sampling, preparation and analisys. New York, Marcel Dekker.