ANGELO MIGUEL SANTIN
CONTROLE BIOLÓGICO DA PLANTA INVASORA
Miconia calvescens POR UMA NOVA
ESPÉCIE DE Ditylenchus
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Fitopatologia, para obtenção do título de Magister Scientiae.
VIÇOSA
Ficha catalográfica preparada pela Seção de Catalogação e Classificação da Biblioteca Central da UFV
T
Santin, Angelo Miguel, 1980-
S235c Controle biológico da planta invasora Miconia calvescens 2008 por uma nova espécie de Ditylenchus / Ângelo Miguel Santin.
– Viçosa, MG, 2008.
xii, 80f.: il. (algumas col.) ; 29cm.
Orientador: Rosangela D’Arc de Lima Oliveira.
Dissertação (mestrado) - Universidade Federal de Viçosa. Inclui bibliografia.
1. Miconia calvescens - Controle biológico.
2. Ditylenchus. 3. Controle biológico. 4. Fitopatologia - Aspectos epidemiológicos. I. Universidade Federal de Viçosa. II.Título.
ANGELO MIGUEL SANTIN
CONTROLE BIOLÓGICO DA PLANTA INVASORA
Miconia calvescens POR UMA NOVA
ESPÉCIE DE Ditylenchus
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Fitopatologia, para obtenção do título de Magister Scientiae.
APROVADA: 30 de abril de 2008.
__________________________________ __________________________________ Prof. Robert Weingart Barreto Prof. Luiz Antonio Maffia
(Co-orientador) (Co-orientador)
__________________________________ __________________________________ Dr. Dartanhã José Soares Prof. Silamar Ferraz
_________________________________ Profa Rosângela D’Arc de Lima Oliveira
Aos meus pais, Valdir e Lourdes. À minha namorada, Nadia. À minha irmã, Carla.
AGRADECIMENTOS
À Universidade Federal de Viçosa e ao Departamento de Fitopatologia, pela oportunidade de realização do Programa de Pós-Graduação.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela concessão da bolsa de estudo.
Aos professores Rosângela D’Arc de Lima Oliveira, Robert Weingart Barreto e Luiz Antonio Maffia, pela orientação, pela dedicação, pelos ensinamentos e pela amizade.
À professora Aristéa Alves Azevedo, pelas sugestões e informações necessárias à realização deste trabalho.
À Dalila Ceni, pela dedicação e pelo companheirismo na realização dos trabalhos.
Ao Dr. Dartanhã José Soares, pelas sugestões e informações necessárias à correção deste trabalho.
Aos servidores Célio e Eloi, pela ajuda e pelas informações.
Aos colegas Roseli, Rodrigo, Naylor, Renata, Patrícia, Douglas, Aline, Déborah, Henrique, Davi, Bruno, Elizangela, Marcelo, Geraldo, Flávio, Fabiano e Rodrigo.
BIOGRAFIA
ANGELO MIGUEL SANTIN, filho de Valdir Antonio Santin e Lourdes Santin, nasceu em 6 de dezembro de 1980, em Faxinal dos Guedes, Estado de Santa Catarina.
Em 2000, iniciou o Curso de Agronomia na Universidade Federal de Viçosa, graduando-se em 25 de julho de 2005.
SUMÁRIO
Página
RESUMO... ix
ABSTRACT... xi
1. INTRODUÇÃO GERAL... 1
REFERÊNCIAS BIBLIOGRÁFICAS ... 7
CAPÍTULO 1... 10
GAMA DE HOSPEDEIROS DE Ditylenchus gallaeformans sp. n., POTENCIAL AGENTE DE BIOCONTROLE DE Miconia calvescens (MELASTOMATACEAE)... 10
1. INTRODUÇÃO ... 10
2. MATERIAL E MÉTODOS ... 12
2.1. Determinação indireta da gama de hospedeiros de Ditylenchus gallaeformans ... 12
2.2. Determinação da gama de hospedeiro por meio de inoculações ... 15
2.3. Verificação da infectividade de Ditylenchus gallaeformans a Miconia calvescens do biótipo do Havaí em condições de campo... 16
3. RESULTADOS ... 17 3.1. Determinação indireta da gama de hospedeiros de Ditylenchus
Página
3.2. Determinação da gama de hospedeiro por meio de inoculações
artificiais ... 17
3.2.1. Descrição dos sintomas por espécie... 19
3.3. Verificação da infectividade de Ditylenchus gallaeformans a Miconia calvescens do biótipo do Havaí em condições de campo 23 4. DISCUSSÃO ... 24
REFERÊNCIAS BIBLIOGRÁFICAS ... 28
CAPÍTULO 2... 30
HISTOPATOLOGIA DE GALHAS INDUZIDAS POR Ditylenchus gallaeformans sp. n. EM FOLHAS DE Miconia ibaguensis 30 1. INTRODUÇÃO ... 30
2. MATERIAL E MÉTODOS ... 32
2.1. Coleta e processamento do material coletado no campo ... 32
2.2. Histopatologia de folhas de M. ibaguensis inoculadas com Ditylenchus gallaeformans ... 33
3. RESULTADOS ... 34
3.1. Cronologia do desenvolvimento das galhas em tecidos inoculados artificialmente... 37
4. DISCUSSÃO ... 40
REFERÊNCIAS BIBLIOGRÁFICAS ... 43
CAPÍTULO 3... 45
ASPECTOS BIOLÓGICOS E ECOLÓGICOS DE Ditylenchus gallaeformans sp. n., POTENCIAL AGENTE DE BIOCONTROLE DE Miconia calvescens (MELASTOMATACEAE) ... 45
1. INTRODUÇÃO ... 45
2. MATERIAL E MÉTODOS ... 47
2.1. Ciclo de vida de Ditylenchus gallaeformans ... 47
2.1.1. Período de emberiogênese ... 47
Página
2.2. Verificação do estádio de anidrobiose de Ditylenchus gallaeformans ... 49
2.2.1. Verificação do estádio de anidrobiose de Ditylenchus gallaeformans em galhas secas encontradas no campo ... 49
2.2.2. Sobrevivência de Ditylenchus gallaeformans em tecidos vegetais campo... 49
2.3. Dinâmica temporal da população de Ditylenchus gallaeformans em plantas de Miconia ibaguensis ... 50
3. RESULTADOS ... 51
3.1. Determinação da duração do ciclo de vida de D. gallaeformans ... 51
3.1.1. Período de embriogênese ... 51
3.1.2. Período de desenvolvimento pós-embriogênico ... 52
3.2. Verificação do estádio de anidrobiose de Ditylenchus gallaeformans ... 52
3.2.1. Verificação do estádio de anidrobiose de Ditylenchus gallaeformans em galhas secas encontradas no campo ... 52
3.2.2. Sobrevivência de Ditylenchus gallaeformans em tecidos vegetais .... 53
3.3. Dinâmica temporal da população de Ditylenchus gallaeformans em plantas de Miconia ibaguensis ... 54
4. DISCUSSÃO ... 57
REFERÊNCIAS BIBLIOGRÁFICAS ... 60
CAPÍTULO 4... 62
DANOS CAUSADOS POR Ditylenchus gallaeformans sp. n., EM PLANTAS DE Miconia calvescens (MELASTOMATACEAE) ... 62
1. INTRODUÇÃO ... 62
2. MATERIAL E MÉTODOS ... 64
2.1. Métodos de inoculação de Ditylenchus gallaeformans ... 64
2.2. Impacto da aplicação de concentrações crescentes de inóculo de Ditylenchus gallaeformans em plantas de Miconia calvescens ... 65
2.3. Análise estatística ... 65
3. RESULTADOS ... 68
Página
3.2. Impacto da aplicação de concentrações crescentes de inóculo de
Ditylenchus gallaeformans em plantas de Miconia calvescens...
RESUMO
SANTIN, Angelo Miguel, M.Sc., Universidade Federal de Viçosa, abril de 2008.
Controle biológico da planta invasora Miconia calvescens por uma nova espécie de Ditylenchus. Orientadora: Rosângela D’Arc de Lima Oliveira. Co-orientadores:
Robert Weingart Barreto e Luiz Antonio Maffia.
Levantamentos exploratórios de fitopatógenos associados à Miconia calvescens para uso em controle biológico apontaram o nematóide Ditylenchus gallaeformans, agente causal de galhas e deformações em tecidos aéreos de melastomatáceas, como tendo potencial para este fim. Considerando a interação de melastomatáceas com este nematóide, objetivou-se estudar os aspectos biológicos e ecológicos da nova espécie; a histopatologia das galhas induzidas em seus hospedeiros; a gama de hospedeiros; e o impacto do nematóide em plantas de M. calvescens. A gama de hospedeiros de
D. gallaeformans ficou restrita às plantas da família Melastomataceae e as do gênero Miconia foram consideradas as mais suscetíveis. D. gallaeformans foi patogênico à M. calvescens, tanto do biótipo do Havaí quanto do Brasil. D. gallaeformans apresentou
D. gallaeformans foram originadas da hiperplasia de células do meristema fundamental
ABSTRACT
SANTIN, Angelo Miguel, M.Sc., Universidade Federal de Viçosa, april, 2008.
Biological control of the invasive weed Miconia calvescens using a new species of
Ditylenchus. Adviser: Rosângela D’Arc de Lima Oliveira. Co-advisers: Robert
Weingart Barreto and Luiz Antonio Maffia.
Exploratory assessments of phytopathogens associated with Miconia calvescens to be used for biological control have pointed the nematode Ditylenchus gallaeformans, a causal agent of galls and deformations in aerial tissues of Melastomataceae as being potentially fit for such purpose. The description of the new species with the proposition of the name D. gallaeformans is in submission phase for the publication. Considering the interaction of Melastomataceae with this nematode, the aim of this work was to study the biological and ecological aspects of the new species; galls histopathology induced in their hosts; host range; and the impact of the nematodes on plants of
M. calvescens. D. gallaeformans host range was restricted to the plants of the family
correlate with the climatic variables. The foliar galls formed by D. gallaeformans originated from hyperplasia of the fundamental meristem cells in the leaf primordia. The galls presented central cavities housing the nematodes and connected to the external environment. These cavities were surrounded by layers of dense cells responsible for supplying food to the nematodes. The most efficient inoculation method was the deposition of nematode suspension on the leaf primordia. A positive correlation was found among incidence, severity, and nematode population in plants inoculated at different concentrations of D. gallaeformans inoculum. The third leaf pair emitted after inoculation presented high correlation with incidence and severity, being considered the ideal point for evaluation of these variables. There was a decrease in the development of plants of M. calvescens with increased concentration of the inoculum of
D. gallaeformans. It was concluded that D. gallaeformans is specific for the family
1. INTRODUÇÃO GERAL
A disseminação antrópica de plantas involuntárias ou deliberadas tem aumentado assustadoramente em todos os continentes; muitas se tornaram indesejáveis, daninhas e, ou, invasoras. Algumas se multiplicaram de forma explosiva e promoveram invasões biológicas, que resultaram em alterações ambientais graves em novos ecossistemas, inclusive ameaçando de extinção espécies nativas de plantas e animais (PEDROSA-MACEDO, 2004). O potencial de espécies exóticas em modificar sistemas naturais é tão significativo que as plantas invasoras exóticas são atualmente consideradas como a segunda causa mundial de perda de diversidade biológica (IUCN, 2000).
Dessa forma, a contaminação biológica está sendo equiparada às mudanças climáticas e à ocupação do solo como um dos mais importantes agentes de mudança global de causa antrópica (ZILLER et al., 2004).
As invasões biológicas de origem vegetal envolvem plantas exóticas que se distribuem naturalmente, sem auxílio direto de pessoas, em ambiente natural ou seminatural, alterando a composição, a estrutura ou os processos do ecossistema. A partir desta definição, as plantas invasoras de áreas altamente perturbadas pelo homem são excluídas da lista de plantas envolvidas em invasões biológicas (CRONK e FULLER, 1995).
Miconia calvescens é nativa dos neotrópicos, mas, na região que compreende o
verde-escuro e face abaxial violeta, sendo este biótipo o responsável pelas invasões biológicas nos arquipélagos do Pacífico (MEDEIROS et al., 1997). No Brasil ocorre um biótipo diferente, com 3 a 4 m de altura, folhas menores entre 30 e 40 cm e com as duas faces verdes. O nome M. calvescens foi originalmente proposto para o biótipo que ocorre no Brasil, mas, apesar das evidentes diferenças entre os dois biótipos, as autoridades no gênero insistem em considerá-las co-específicas.
Entre os ecossistemas mais suscetíveis e vulneráveis às invasões biológicas estão os encontrados nas ilhas oceânicas. Por causa de uma serie de fatores únicos encontrados nestes locais, as ilhas oceânicas apresentam baixo número de espécies nativas, resultado do isolamento geográfico. Nestes locais, normalmente não há duas espécies nativas competindo por um mesmo nicho ecológico, o que leva a uma fraca capacidade de competição; além disso, os baixos níveis de fluxo de genes, devido à pouca variabilidade genética dentro das populações, as tornam mais sensíveis a mudanças ambientais. Espécies exóticas, adaptadas a estes ambientes e livres de seus inimigos naturais, apresentam nítida vantagem sobre as espécies nativas destes ecossistemas (CRONK e FULLER, 1995).
Miconia calvescens é reconhecida hoje como uma das plantas invasoras mais
Em 1937, M. calvescens foi introduzida como planta ornamental no Taiti, e hoje está distribuída em 65 % dos 1.045 km2 da ilha. Destes, 25 % ocorre de forma densa, afetando diretamente o ecossistema da ilha ao provocar sombreamento excessivo, o que as espécies nativas não toleram. Desta forma, ela se sobressai em relação às espécies nativas (MEDEIROS et al., 1997). As estimativas são de que no Taiti existam de 70 a 100 espécies vegetais, das quais 35 a 45 são espécies endêmicas da Polinésia Francesa e estão diretamente ameaçadas pela invasão de M. calvescens (MEYER, 1996).
No Havaí, esta espécie foi introduzida por volta de 1961, como planta ornamental, e desde 1992 está presente na lista de plantas invasoras nocivas ao arquipélago. Está atualmente distribuída nas ilhas do Havaí, Maui, Oahu, e Kauai, no Arquipélago Havaiano (MEYER, 1996). Nestes locais, uma série de medidas foi tomada visando a sua erradicação. Entre 1993 e 1995, cerca de 20 mil plantas foram destruídas manualmente, porém, a grande capacidade de regeneração, graças ao considerável banco de sementes, inviabilizou esta estratégia. Em áreas de difícil acesso, tentativas de controle por pulverização aérea fracassaram, pois não houve mortalidade total das plantas, além do favorecimento da germinação do banco de sementes (LOOPE, 1997).
A remoção física e o controle químico obtiveram sucesso na contenção da sua expansão para novas áreas, porém, o controle completo e duradouro requer a introdução de agentes patogênicos por meio de controle biológico clássico (MEDEIROS et al., 1997). O controle biológico é um meio natural de controlar as pragas, pois utiliza seus inimigos naturais sem os inconvenientes causados pelos agrotóxicos, mas exige estudos aprofundados antes de ser empregado, para que sua introdução não cause desequilíbrios ambientais. Wikler (2004), referindo-se aos insetos, afirma que
para um agente de controle biológico ter sucesso, é importante que ele se adapte ao ambiente, tenha grande capacidade de busca da planta-alvo, capacidade própria de dispersar-se entre as plantas-alvo e reaja rapidamente às mudanças na população da planta invasora.
Os agentes de controle biológico são partes ou a íntegra da solução para as invasões biológicas, porém, nem sempre são baratos nem fáceis de serem encontrados, mas os seus efeitos são duradouros e não-poluentes (PEDROSA-MACEDO, 2004).
método clássico ou inoculativo de controle biológico de plantas invasoras envolve a busca e seleção de patógenos (inimigo natural) da planta-alvo no seu centro de origem e a sua introdução na nova área de distribuição da planta que se encontra livre de seus inimigos naturais, visando restabelecer o equilíbrio (BARRETO, 2004).
Desde 1995, após o estabelecimento de um convênio entre a FUNARBE (Fundação Arthur Bernardes) da Universidade Federal de Viçosa e o RCUH (Research
Corporation of the University of Hawaii) da Universidade do Havaí tem-se estudado
uma série de fitopatógenos e insetos associados à M. calvescens no Brasil e em outros países da América do Sul e Central. Em 1997, o fungo Colletotrichum gloeosporioides f. sp. miconiae, detectado no Estado de Minas Gerais, após avaliação e demonstração de grande potencial para o biocontrole de M. calvescens, foi introduzido no Havaí, tornando-se o primeiro exemplo de fungo fitopatogênico brasileiro a ser introduzido no exterior para o controle biológico clássico de uma planta invasora (BARRETO et al., 2001). Outros organismos mostraram-se promissores para a utilização no controle biológico desta planta, como o fungo Coccodiella miconiae (SEIXAS, 2002). Em 2005, uma nova espécie de nematóide, pertencente ao gênero Ditylenchus, foi constatada causando deformações foliares, galhas e formação de filódios sobre folhas, hastes e inflorescências de M. calvescens e outras melastomatáceas, apresentando grande potencial para ser utilizado como agente de biocontrole de melastomatáceas (OLIVEIRA et al., 2005).
Todos os nematóides estudados como possíveis agentes de controle biológico de plantas, pertencem à família Anguinidae e apresentam como característica comum a capacidade de induzir em seus hospedeiros deformações e galhas nos tecidos aéreos de seus hospedeiros (PARKER, 1991). Os principais gêneros da família Anguinidae que atacam tecidos aéreos de plantas são: Anguina, Subanguina e Ditylenchus.
(criptobiose/anidrobiose) induzido pela desidratação do corpo. Desta forma, sobreviverão até a próxima estação, quando iniciarão um novo ciclo da doença (WATSON, 1986). Esta espécie foi introduzida nos Estados Unidos e no Canadá e se tornou uma das principais invasoras nestes países (WATSON, 1986). Por causa da severidade dos danos provocados, o nematóide S. picridis apresenta-se como um agente de grande potencial para ser usado no controle biológico de knapweed russo, pois, sua gama de hospedeiros é restrita a espécies da tribo Cynareae da família Asteraceae (KRALL, 1991).
Brzeski (1981), após revisar a família Anguinidae, propôs a inclusão do gênero
Ditylenchus, com 81 espécies válidas. A maioria das espécies vive no solo como
parasitas de plantas ou alimentando-se de micélio de fungos, entre as espécies fitopatogênicas, algumas apresentam relações muito específicas com suas plantas hospedeiras, podendo ser utilizadas no biocontrole de plantas (STURHAN e BRZESKI, 1991).
Seixas et al. (2004) detectaram a presença de Ditylenchus drepanocercus Goodey, agente causal da mancha-angular em M. calvescens, durante buscas por agentes de biocontrole desta espécie de melastomatácea no Brasil. Esse nematóide apresenta alto nível de especificidade com seus hospedeiros (STURHAN e BRZESKI, 1991). D. drepanocercus é um endoparasita migrador que vive alojado no parênquima lacunoso, provoca hipertrofia das células que aumentam até 13 vezes em tamanho. Este nematóide não tem muita resistência à dessecação. As lesões ocorrem apenas nas folhas, principalmente nas mais velhas e na superfície abaxial. As lesões são angulares e delimitadas pelas nervuras; inicialmente, são de coloração amarela progredindo até a necrose do tecido, tamanho entre 1 e 5 cm, podendo coalecer, atingindo grandes proporções do limbo foliar. A severidade da doença no campo é afetada pela temperatura e umidade (SEIXAS, 2002). As semelhanças nas condições climáticas entre Brasil e Havaí, onde M. calvescens é invasora e a limitada gama de hospedeiros, sugerem algum potencial deste nematóide como agente de biocontrole desta invasora (SEIXAS et al., 2004).
Outra espécie, o nematóide D. phyllobius Brzeski, 1991 (= Anguillulina
phyllobia (Thorne, 1934), = Nothanguina phyllobia (Thorne, 1934), = Orrina phyllobia
(Thorne, 1934)) apresenta grande potencial para o biocontrole da invasora Solanum
elaeagnifolium (Solanaceae) (ROBINSON et al., 1979), uma vez que apresenta alto
vida deste nematóide depende da emergência de juvenis infecciosos de quarto estádio, sobreviventes em galhas de folhas caídas no solo (ROBINSON, 1985). Durante períodos de alta umidade relativa (UR) e temperaturas amenas, os juvenis saem do estádio de quiescência e escalam o hospedeiro até atingirem os tecidos aéreos da região apical, penetrando e desenvolvendo-se ativamente (ROBINSON et al., 1978). A atividade de movimentação dos juvenis infecciosos é afetada pela temperatura e umidade relativa do ar, sendo a temperatura ótima encontrada por volta de 25 ºC, e quanto maior a UR mais rápida é a movimentação do nematóide, permitindo que ele atinja a região apical rapidamente, com menor risco de dessecação (ROBINSON et al., 1981; 1984). Os juvenis infectam o limbo e o pecíolo de folhas jovens no ápice do broto, provocando hipertrofia e hiperplasia do parênquima cortical e vascular e formam uma região de confluência entre limbo, pecíolo e talo, que resultarão em galhas com volume de até 25 cm3. As galhas imaturas possuem coloração verde-clara e formato irregular, sendo observadas, primeiramente, na superfície abaxial da folha; já galhas maduras são de formato esférico, com uma cavidade central formada por espaços intercelulares, onde se encontram fileiras de células com tamanho maior que o normal com vacúolos grandes, indicando que grande quantidade de energia da planta é usada pelo nematóide (SKINNER et al., 1980).
Apesar das três espécies citadas acima como possíveis agentes de controle biológico de plantas, não há nenhum registro de sua efetiva introdução como agente de controle biológico para as suas plantas-alvo.
Este trabalho teve por objetivo estudar os aspectos biológicos e ecológicos de
D. gallaeformans, a histopatologia das galhas induzidas em seus hospedeiros, a gama de
REFERÊNCIAS BIBLIOGRÁFICAS
ANDREAS, L. A.; GOEDEN, R. D. Biological control of weeds. In: PROCEEDINGS OF THE AAAS SYMPOSIUM ON BIOLOGICAL CONTROL. Boston: Massachussets, 1969. 511 p.
BARRETO, R. W.; SEIXAS, C. D. S.; KILLGORE, E. Colletotrichum gloeosporioides f. sp. miconiae: o primeiro fungo fitopatogênico brasileiro a ser introduzido no exterior para o controle biológico clássico de uma planta invasora: (Miconia calvescens). In: SIMPÓSIO DE CONTROLE BIOLÓGICO, 7., 2001. Poços de Caldas. Livro de
Resumos, Poços de Caldas, 2001. p.109.
BARRETO, R. W. Uso de agentes patogênicos no controle biológico de plantas invasoras. In: PEDROSA-MACEDO, J. H.; BREDOW, E. A. Princípios e rudimentos
do controle biológico de plantas, 2004. p. 143-160.
BRZESKI, M. W. The genera of Anguinidae (nematoda, Tylenchida). Revue
Nematologie, v. 4, p. 23-34, 1981.
CRONK, Q. C. B.; FULLER, J. L. Plant invaders – The threat to natural ecosystems. London: Chapman e Hall, 1995. 239 p.
INTERNATIONAL UNION FOR CONSERVATION OF NATURE – IUCN. 100 of
the World’s Worst Invasive Alien Species. Auckland invasive species specialist
group, 2000. Disponível em: <http://www. iucn.org/ >.
LOOPE, L. L. Miconia calvescens D.C. HNIS Report for Miconia calvescens. A product of the Hawaiian Ecossistems at Risk Project, 1997. 8 p.
MEYER, J. Y. Mecanismes d'invasion de Miconia calvescens en Polynesie Français. Ph.D. Thesis. L'Université de Montpellier II Sciences et Techniques du Languedoc. Montpellier, France, 1994.
MEYER, J. Y. Status of Miconia calvescens (Melastomataceae), a dominant invasive tree in the Society Islands (French Polynesia). Pacific Science, v. 50, n. 1, p. 66-76, 1996.
MEYER, J. Y. Observations on the reproductive biology of Miconia calvescens DC. (Melastomataceae) an alien invasive tree on the islands of Tahiti (South Pacific Ocean).
Biotropica, v. 30, n. 4, p. 609-624, 1998.
OLIVEIRA, R. D. L.; BARRETO, R. W.; SANTIN, A. M. A new species of
Ditylenchus causing galls on the invasive weed Miconia calvecens. In: ANNUAL
MEETING OF THE ORGANIZATION OF NEMATOLOGIST OF TROPICAL AMERICA (ONTA), 37., 2005, Viña Del Mar, Chile. Resumos, Viña Del Mar, Chile, 2005. 90 p.
PARKER, P. E. Nematodes as biological control agents of weeds. Microbial Control of Weeds. New York: Chapman Hall, 1991. p. 58-68.
PEDROSA-MACEDO, J. H. Princípios e rudimentos do controle biológico de plantas. In: PEDROSA-MACEDO, J. H.; BREDOW, E. A. Princípios e rudimentos do
controle biológico de plantas/coletânea, 2004. p. 115-128.
ROBINSON, A. F. Observations of molting and population development by Orrina
phyllobia. Journal of Nematology, v. 17, p. 239-241, 1985.
ROBINSON, A. F.; ORR, C. C.; HEINTZ, C. E. Some factors affecting survival of desiccation by infective juveniles of Orrina phyllobia. Journal of Nematology, v. 16, p. 86-91, 1984.
ROBINSON, A. F.; ORR, C. C.; HEINTZ, C. E. Effects of oxygen and temperature on the activity and survival of Nothanguina phyllobia. Journal of Nematology, v. 13, p. 528-535, 1981.
ROBINSON, A. F.; ORR, C. C.; ABERNATHY, J. R. Behavioral response of
Nothonguino phyllobio to selected plant species. Journal of Nematology, v. 11, p.
73-77, 1979.
ROBINSON, A. F.; ORR, C. C.; ABERNATHY, J. R. Distribution of Nothanguina
phyllobia and its potential as a biological control agent for silver-leaf nightshade. Journal of Nematology, v. 10, n. 4, p. 362-367, 1978.
SEIXAS, C. D. S. Controle biológico de Miconia calvescens (Melastomatácea) com
fitopatógenos. 2002. 123 f. Tese (Doutorado em Fitopatologia) – Universidade Federal
de Viçosa, Viçosa, 2002.
SEIXAS, C. D. S.; BARRETO, W. R.; FREITAS, L. G.; MONTEIRO, F. T.; OLIVEIRA, R. D. L. Ditylenchus drepanocercus rediscovered in the neotropics causing angular leaf spots on Miconia calvescens. Journal of Nematology, v. 36, n. 4, p. 481-486, 2004.
SKINNER, J. A.; ORR, C. C.; ROBINSON, A. F. Histopathogenesis of the galls induced by Nothanguina phyllobia in Solanum elaeagnifolium. Journal of
Nematology, v. 12, n. 2, p. 142-150, 1980.
STURHAN, D.; BRZESKI, M. W. Stem and bulb nematodes, ditylenchus spp. In: NICKLE, W. R. (Ed.) Manual of Agricultural Nematology. New York, 1991. p.423-459.
WATSON, A. K. Biology of Subanguina picridis, a potential biological control agent of Russian Knapweed. Journal of Nematology, v. 18, n. 2, p. 154-158, 1986.
WATSON A. K. Host range of, and plant reaction to, Subanguina picridis. Journal of
Nematology, v. 18, n. 1, p. 112-120, 1986.
WIKLER, C. Princípios e rudimentos do controle biológico de plantas. In: PEDROSA-MACEDO, J. H.; BREDOW, E. A. (Ed.) Princípios e rudimentos do controle
biológico de plantas/coletânea, 2004. Cap. 7, p. 115-128.
CAPÍTULO 1
GAMA DE HOSPEDEIROS DE Ditylenchus gallaeformans sp. n.,
POTENCIAL AGENTE DE BIOCONTROLE DE Miconia calvescens
(MELASTOMATACEAE)
1. INTRODUÇÃO
Miconia calvescens DC (Melastomataceae) é nativa desde o sul do México ao
sul do Brasil e norte da Argentina (MEDEIROS et al., 1997). É reconhecida hoje como uma das plantas invasoras mais ameaçadoras dos ecossistemas florestais do Havaí e Polinésia Francesa, onde foi introduzida para uso ornamental (MEYER, 1994; MEDEIROS et al., 1997). M. calvescens apresenta todos os atributos de uma espécie invasora agressiva, como crescimento rápido, tolerância ao sombreamento, que permite sua germinação e permanência em baixos níveis de luz, alta taxa de produção e dispersão de sementes (MEYER, 1998). Para casos de invasões biológicas como estes, a medida que se apresenta mais viável para o seu controle é a introdução de inimigos naturais, por meio de controle biológico clássico (MEDEIROS et al., 1997).
Entre as várias etapas do processo de estudo de viabilidade de um agente de controle biológico, uma das mais importantes é a verificação de sua gama de hospedeiros. Somente depois de determinada a especificidade do agente de biocontrole é possível a sua liberação, pois agentes inespecíficos são inadequados para introduções em ambientes exóticos (SIMBERLOFF e STILING, 1996).
Os procedimentos mais utilizados para verificação da especificidade são com base em Wapshere (1974) e consistem na formação de um conjunto de espécies de plantas-teste selecionadas em função de sua relação filogenética com a planta-alvo. Estas plantas são expostas ao potencial agente de biocontrole em condições controladas. Watson (1986) estudou a gama de hospedeiros de Subanguina picridis (Kirjanova) Brzeski um nematóide investigado como potencial agente de biocontrole de Acroptilon
repens (L.) DC., pelo método citado acima, inoculando plantas de 66 espécies
pertencentes a 34 famílias em condições controladas, com suspensão de nematóides. Desta lista de plantas-teste, o nematóide infectou apenas sete espécies pertencentes a duas subtribos da família Asteraceae. Mais tarde, foi verificado, no entanto, que muitas outras espécies de plantas podiam ser infectadas por este nematóide (PARKER, 1991), por causa da pouca representatividade das espécies de plantas usadas no teste centrífugo-filogenético.
Outra forma de avaliar a especificidade de um agente de controle biológico é por meio da utilização do método indireto de avaliação, descrito por Seixas (2002). Neste método são selecionadas espécies de plantas pertencentes a diversos gêneros e famílias com ênfase nas espécies mais próximas filogeneticamente da planta-alvo, no entorno de plantas-alvo infestadas pelo agente, em condições naturais de campo favoráveis ao desenvolvimento da doença. Parte-se da pressuposição de que qualquer outra planta presente na mesma área estiver constantemente exposta ao possível ataque do patógeno e se a infecção não ocorrer, é por que estas plantas não são hospedeiras do agente.
Neste capítulo, objetivou-se determinar a gama de hospedeiros de
D. gallaeformans, pelos dois métodos, direto e indireto, e a verificação da capacidade
2. MATERIAL E MÉTODOS
2.1. Determinação indireta da gama de hospedeiros de Ditylenchus gallaeformans
Quadro 1 – Plantas avaliadas no teste de especificidade indireta de D. gallaeformans Divisão Magnoliophyta
Classe Magnoliopsida
Ordem Família Espécie Coleta*
Leandra sp. C
Clidemia hirta D. Don C/D/G
Tibouchina granulosa (Desr.) Cogn. C/D
Tibouchina herbacea (DC). Cogn. G
Miconia calvescens DC. C/G
Miconia sp. 1 G
Miconia sp. 2 C
Miconia albicans Steud. C/D/G Miconia cinnamomifolia Triana C/D
Miconia ibaguensis Triana C/D/G
Melastomataceae
Miconia latecrenata Naudin C/D
Psidium guajava L. C/G
Eugenia uniflora L. C
Myrtaceae
Eucalyptus sp. D
Myrtales
Lythraceae Não identificada C
Malpighiaceae Lophanthera lactescens Ducke C
Clusiaceae Vismia brasiliensis Choisy C/G
Phyllanthaceae Phyllanthus sp. D
Passiflora sp. G
Malpighiales
Passifloraceae
Passiflora edulis Sims C
Desmodium sp. C/D
Fabaceae
Piptadenia sp. G
Fabales
Polygalaceae Diclidanthera sp. C
Joaneria sp. G Manihot esculenta Crantz C
Croton floribundus Spreng D/G
Croton sp. C
Euphorbiales Euphorbiaceae
Mabea sp. C/G
Erythroxylum sp. G
Linales Erythroxylaceae
Erythroxylum pelleterianum A. St.-Hil. C
Trichilia pallida Sw. D
Toona sp. D
Meliaceae
Guarea macrophylla Vahl G
Citrus sp. C
Rutaceae
Zanthoxylum sp. D
Anacardiaceae Schinus terebinthifolius Raddi C/G
Paullinia sp. D
Sapindales
Sapindacea
Cardiospermum sp. C
Geraniales Balsaminaceae Impatiens walleriana Hook.f. D
Eriobotrya japonica (Thunb.) Lindl. C
Rosales Rosaceae
Rubus rosifolius Stokes C
Centella sp. G
Apiales Umbelliferae
Centella asiatica (L.) Urb. C
Nectandra rigida (Kunth ) Nees D
Lauraceae
Persea americana Mill. C
Laurales
Monitiniaceae Siparuna guianensis Aubl. G
Ranunculales Menispermaceae Não identificada D
Magnoliales Annonaceae Malmea sp. C
Piper sp. D
Piper aduncum L. C/G
Piperales Piperaceae
Pothomorphe umbellata (L.) Miq. C
Quadro 1, Cont.
Divisão Magnoliophyta Classe Magnoliopsida
Ordem Família Espécie Coleta*
Moraceae Ficus sp. D
Urticaceae Urtica sp. D
Urticales
Cecropiaceae Cecropia sp. C/D/G
Pavonia communis A. St.-Hil. C
Malvaceae
Sida sp. D/G
Luehea grandiflora Mart. C
Malvales
Tiliaceae
Luehea sp. G
Violales Curcubitaceae Cucurbita sp. C
Solanum sp. C
Solanaceae
Solanum cernuum Vell. G
Ipomoea sp. C
Solanales
Convolvulaceae
Ipomoea cairica ( L. ) Sweet C
Gentianales Apocynaceae Peschiera laeta Miers C
Verbenaceae Lantana camara L. C/G
Boraginaceae Borago sp. G
Gesneriaceae Sinningia sp. C
Lamiaceae Schizonepeta sp. C
Lamiales
Plantaginaceae Plantago sp. C
Coffea arabica L. C/D Spermacoce latifólia Aubl. C
Psychotria sp. G
Rubiales Rubiaceae
Richardia brasiliensis Gomes C
Thunbergia alata Bojer ex Sims G
Acanthaceae
Hypoestes sanguinolenta Hook. D
Tabebuia sp.(1) G
Tabebuia sp.(2) D
Scrophulariales
Bignoneaceae
Macfadyena unguiscati (L.) D
Mikania sp. C
Chromolaena odorata L. C
Baccharis sp. G
Asterales Asteraceae
Vernonia sp. C
Caryophyllales Amaranthaceae Gomphrena sp. C
Classe Liliopsida
Alismatales Araceae Xanthosoma sagittifolium K.Koch C
Subclasse Commelinidae
Commelinales Commelinaceae Commelina benghalensis Wall G/C
Arecales Arecaceae Euterpe sp. C
Bromeliaceae Tillandsia sp. D
Cyperaceae Fimbristylis sp. D
Panicum maximum Nees G
Melinis minutiflora P.Beauv. C
Poales
Poaceae
Imperata brasiliensis Trin. G
Maranthaceae Calathea sp. D
Heliconiaceae Heliconia sp. C
Zingiberales
Musaceae Musa sp. C
Orchidales Orchidaceae Oeceoclades maculata L. D
Liliales Smilacaceae Smilax sp. D
Hypoxidaceae Hypoxis decumbens L. C
Asparagales
Ruscaceae Sansevieria sp. D
Quadro 1, Cont.
Divisão Cycadophyta Classe Cycadopsida
Ordem Família Espécie Coleta*
Cycadales Cycadaceae Cycas sp. D
Divisão Pteridophyta Classe Pteridopsida
Gleicheniaceae Gleichenia sp. C
Filicales
Schizaeaceae Anemia phyllitidis (L.) Sw C
Divisão Lycopodiophyta Classe Selaginellopsida
Lycopodiales Selaginellaceae Selaginella sp. G
* Local da coleta: C = Cristais, Viçosa, MG; D = Dendrologia, Campus da UFV, Viçosa, MG; G = Guaraciaba, MG.
2.2. Determinação da gama de hospedeiro por meio de inoculações
O inóculo foi obtido de folhas e inflorescências coletadas em plantas de
M. Ibaguensis, naturalmente infectadas no campo. As galhas foram retiradas, levadas ao
laboratório e trituradas por 20 s no liquidificador; posteriormente, foram passadas em peneiras de 60 e 500 mesh, para separar os nematóides dos restos vegetais. A suspensão obtida da peneira de 500 mesh foi deixada no funil de Baermann por 24 horas, para separar os nematóides vivos dos mortos. A suspensão recolhida do funil de Baermann foi calibrada para 1.500 nematóides/mL, com auxílio da câmara de Peters.
Foram testadas 39 espécies, com ênfase em espécies relacionadas filogenética-mente com a família Melastomataceae, seguindo-se o método centrífugo-filogenético de Wapshere (1974) e incluindo de importância econômica (Quadro 2).
Cada planta foi inoculada com 2 mL da suspensão contendo os nematóides. A inoculação foi realizada pela deposição da suspensão de nematóides sobre pedaços de algodão nos primórdios foliares, com auxílio de uma pipeta. Após a inoculação, as plantas foram mantidas em câmara úmida a 25 °C, por 48 horas e fotoperíodo de 12 horas em câmaras de crescimento. Posteriormente, foram transferidas para casa de vegetação do Departamento de Fitopatologia da Universidade Federal de Viçosa, recebendo os tratos culturais necessários ao bom desenvolvimento.
laboratório e as áreas que continham possíveis sintomas da presença do nematóide, como deformações e galhas foliares, foram recortadas e pesadas. Nas espécies em que não houve o desenvolvimento de sintomas, foi retirada, aleatoriamente, uma amostra representativa da parte aérea da planta e, posteriormente, realizou-se a extração, como descrito no item 1. Após a extração, as suspensões referentes a cada amostra foram examinadas em microscópio estereoscópico para verificação da presença de nematóides. Nas amostras em que se verificou a presença de nematóides foi realizada a montagem de lâminas para identificação dos nematóides em microscópio de luz e, posteriormente, foi realizada a sua contagem com auxílio da câmara de Peters. Após a determinação do número total de nematóides, por amostra, foi calculado o fator de reprodução (FR = população final/população inicial), segundo Oostenbrink (1966).
O experimento foi conduzido em delineamento inteiramente casualizado, com 39 tratamentos representados por cada espécie e quatro repetições. A unidade experimental foi composta por uma planta com um ponto de inoculação. Os dados da população de nematóides foram transformados em ( x +1); aplicada a análise de variância e o teste de médias (Tukey, a 5 %).
2.3. Verificação da infectividade de Ditylenchus gallaeformans a Miconia calvescens
do biótipo do Havaí em condições de campo
3. RESULTADOS
3.1. Determinação indireta da gama de hospedeiros de Ditylenchus gallaeformans
Dentre as 58 famílias representadas neste estudo, D. gallaeformans foi encontrado parasitando apenas plantas pertencentes à família Melastomataceae. Nos três locais de estudo, as espécies hospedeiras pertenciam aos gêneros Miconia (cinco espécies) e Clidemia (uma espécie). Em Viçosa, no bairro Cristais, o nematóide foi encontrado infectando as espécies M. albicans e M. ibaguensis; na Dendrologia, foi encontrado nas espécies M. albicans, M. ibaguensis e M. Latecrenata; e no Município de Guaraciaba, foi encontrado infectando as espécies C. capitellata, M. ibaguensis,
M. calvescens e M. albicans.
3.2. Determinação da gama de hospedeiro por meio de inoculações artificiais
Quadro 2 – Lista de plantas inoculadas com D. galaeformans e os respectivos resultados
Ordem Família Espécie S P
Lythraceae Punica granatum L. - -
Psidium guajava L. - -
Syzygium sp. - -
Myrciaria sp. - -
Marlierea sp. - -
Eugenia sp. - -
Myrtaceae
Eucaliptus sp. - -
Combretaceae Terminalia catappa L. - -
Clidemia hirta D. Don - -
Clidemia capitellata D.Don + +
Clidemia sp. - -
Leandra lacunosa Cogn. + +
Leandra sp. (1) + -
Leandra sp. (2) - -
Miconia albicans (SW.) Steud. + -
Miconia corallina Spring. + -
Miconia prasina DC. + +
Miconia ibaguensis Triana + +
Miconia latecrenata Naudin + -
Miconia cinnamomifolia Naudin + -
Miconia calvescens DC. + +
Miconia sp. (1) - -
Miconia sp. (2) + +
Tibouchina moricandiana Baill. - -
Tibouchina granulosa Cogn. - -
Tibouchina sp - -
Schizocentrum sp. - -
Myrtales
Melastomataceae
Dodonaea sp. - -
Fabales Fabaceae Phaseolus vulgaris L. - -
Rosales Rosaceae Fragaria sp. - -
Violales Cucurbitaceae Cucurbita sp. - -
Euphorbiales Euphorbiaceae Euphorbia heterophylla L. - -
Brassicales Brassicaceae Brassica oleracea L. - -
Malvales Malvaceae Hibiscus sp. - -
Geraniales Geraniaceae Geranium sp. - -
Anacardiaceae Schinus terebinthifolius Raddi - -
Sapindales
Rutaceae Citrus sp. - -
Rubiales Rubiaceae Coffea arábica L. - -
Solanales Solanaceae Capsicum sp. - -
Desenvolvimento de sintomas (S) e presença de Ditylenchus gallaeformans (P) em plantas inoculadas artificialmente em casa de vegetação.
Figura 1 – Sintomas causados por Ditylenchus gallaeformans em plantas coletadas no campo. A = Miconia albicans; B = Miconia latecrenata; C = Miconia
calvescens; e D = Miconia ibaguensis. Nas fotos A e B, as setas indicam
galhas em folhas e inflorescências; e nas fotos C e D, as setas indicam galhas folhares.
3.2.1. Descrição dos sintomas por espécie
Clidemia capitellata: apresentou período latente de 35 dias, desenvolveu sintomas em
duas das quatro plantas inoculadas, mas não apresentou a formação típica de galhas. Desenvolveu deformações no limbo foliar, que provocaram a fusão das nervuras. As regiões deformadas eram planas, de coloração verde-clara, que se desatacavam do restante do limbo foliar.
Leandra lacunosa: apresentou período latente de 30 dias e desenvolveu sintomas em
Miconia prasina: apresentou período latente de 25 dias e desenvolveu os sintomas nas
quatro repetições. As galhas desenvolveram-se por todo limbo foliar, distribuídas pelas duas superfícies. Inicialmente, as galhas apresentaram-se como pequenos pontos de colorações mais claras que o restante do limbo foliar e, em seu entorno as folhas desenvolveram coloração marrom, que permaneceu com o desenvolvimento da galha. Os sintomas foram muito severos neste hospedeiro, provocando fortes deformações no limbo foliar e a paralisando completamente o desenvolvimento da gema apical (Figura 2).
Figura 2 – Sintomas de galhas causadas por Ditylenchus gallaeformans em Miconia
prasina. A = detalhe da coloração marrom no entorno das galhas; e B =
detalhe da deformação total da região apical do ramo aos 120 dias após a inoculação.
Miconia ibaguensis: apresentou período latente de 28 dias e desenvolveu os sintomas
nas quatro repetições. As galhas desenvolveram-se por todo limbo foliar distribuídos pelas duas superfícies. Inicialmente, as galhas surgiam na região das nervuras das folhas em expansão como pequenos pontos de coloração clara, devido ao acumulo de tricomas hialinos no local (Figura 3A). Com o desenvolvimento da folha, as galhas iam surgindo por toda a sua extensão e aumentavam consideravelmente de volume. Os sintomas neste hospedeiro eram mais severos do que nos outros e resultaram na deformação total da região apical (Figura 3B).
Figura 3 – Sintomas das galhas causadas por Ditylenchus gallaeformans em Miconia
ibaguensis. A = detalhe dos sintomas iniciais com pilosidade densa de
coloração branca no início dos sintomas aos 28 dias após a inoculação; e B = detalhe dos sintomas severos com deformação completa da região apical do ramo aos 120 dias após a inoculação.
Miconia calvescens: apresentou período latente de 32 dias. Os sintomas
manifestaram-se em três das quatro repetições. As galhas demanifestaram-senvolveram-manifestaram-se por todo limbo foliar, porém apenas na superfície adaxial da folha. Neste hospedeiro, nos momentos iniciais do desenvolvimento dos sintomas, as galhas eram de coloração clara, como observado nos outros hospedeiros, mas com a evolução dos sintomas, elas adquiriam tons avermelhados. Foi comum neste hospedeiro ocorrer distorção generalizada do limbo foliar com a formação de expansões filiformes de até 3 cm de altura, porém a galhas eram pequenas e encontradas nas reentrâncias formadas por estas expansões (Figura 4).
Miconia sp. (2): o período latente foi de 28 dias. Os sintomas manifestaram-se nas
quatro repetições. As galhas desenvolveram-se semelhantes às da M. calvescens, porém, não houve formação de expansões filiformes.
Figura 4 – Sintomas das galhas causadas por Ditylenchus gallaeformans em Miconia
calvescens. A = detalhe da coloração clara na fase inicial dos sintomas; B =
detalhe das galhas aos 120 dias após a inoculação; e C = detalhe das expansões filiformes formadas no limbo foliar.
Tabela 1 – Número médio de nematóides por planta inoculada com Ditylenchus
gallaeformans e fator de reprodução (FR)
Espécie No Nematóide FR
Miconia ibaguensis 6.840 a 2,15
Miconia prasina 4.360 ab 1,45
Miconia calvescens 2.490 b 0,83
Miconia sp. (2) 2.120 bc 0,71
Clidemia capitellata 460 cd 0,15
Leandra lacunosa 230 d 0,08
3.3. Verificação da infectividade de Ditylenchus gallaeformans a Miconia calvescens
do biótipo do Havaí em condições de campo
As plantas de M. calvescens, biótipo do Havaí, foram infectadas naturalmente
por D. gallaeformans e desenvolveram os sintomas de galhas semelhantes aos observados no biótipo do Brasil (Figura 5). Os sintomas foram observados nas plantas cultivadas nos Municípios de Viçosa e Guaraciaba, MG, e a presença do nematóide foi confirmada nas plantas com sintomas das duas localidades, pela extração e observação dos nematóides em microscópio de luz.
Figura 5 – Sintomas de galhas provocadas pela infecção natural de Ditylenchus
gallaeformans em plantas de Miconia calvescens biótipo do Havaí,
4. DISCUSSÃO
A introdução de agentes exóticos para o controle biológico requer muita atenção, pois este estará livre e poderá causar impacto em plantas de importância agronômica e na flora nativa (PARKER, 1991). Os estudos de gama de hospedeiros são fundamentais para obtenção de agentes de biocontrole seguros e para introdução em áreas onde não ocorre naturalmente. Mais de 500 angiospermas, entre mono e dicotiledôneas, são conhecidas como hospedeiras de nematóides do genêro Ditylenchus, porém algumas espécies do gênero apresentam alto grau de especificidade com seus hospedeiros, como pode-se observar com D. drepanocercus e D. phyllobius (ROBINSON et al., 1979; SEIXAS et al., 2004).
Além dos hospedeiros encontrados no teste indireto de hospedeiros, outros cinco foram identificados: Leandra lacunosa Cogn. e Miconia corallina Spring. procedentes da Estação Ecológica do Tripuí, em Ouro Preto, MG; Miconia waltherii Naudin, procedente da Reserva Ecológica do Caraça em Santa Bárbara, MG; e Leandra aurea Cogn. e Leandra sp., procedentes do Município de Macaé de Cima, RJ. Todos hospedeiros apresentaram os sintomas característicos da infecção causada por
D. gallaeformas em folhas e inflorescências e os nematóides obtidos das extrações não
apresentaram diferenças morfológicas em relação aos encontrados nos outros hospedeiros.
Entre os seus hospedeiros está a planta-alvo, M. calvescens. Seixas et al. (2004) verificaram que D. drepanocercus foi capaz de infectar apenas plantas de M. calvescens do biotipo existente no Brasil e não infectou as do biótipo existente no Havaí, sugerindo a existência especialização fisiológica no patossistema. Isto não foi observado para
D. gallaeformans, que demonstrou capacidade de reproduzir-se em plantas dos dois
biótipos, tanto em inoculações artificiais como em condições naturais, em plantas sentinelas cultivadas no campo. Esta observação é reforçada pelo fato do inóculo utilizado nos testes em condições controladas ser obtido de plantas de M. ibaguensis, e este se mostrou capaz de reproduzir-se nos diferentes hospedeiros. O desenvolvimento dos sintomas no campo em plantas do biótipo do Havaí mostra que D. gallaeformans é capaz de reproduzir-se neste hospedeiro e poderá disseminar-se espontaneamente por áreas infestadas por M. calvescens, no caso de uma possível introdução para o controle biológico desta invasora.
Há evidências de que a disseminação de D. gallaeformans, de plantas naturalmente infectadas para as plantas de M. calvescens do biótipo do Havaí, tenha ocorrido pela água de chuva. Em recipientes instalados sob a projeção da copa dos hospedeiros naturalmente infectados, para coleta da água de chuva, foi verificada a presença do nematóide na água recolhida, o que indica que a chuva tem papel importante na disseminação e redistribuição do nematóide entre os ramos de uma mesma planta.
D. gallaeformans encontra-se amplamente disseminado pela região sudeste do
et al. (1978) verificaram que D. phyllobius, potencial agente de biocontrole de Solanum elaeagnifolium, apresentava-se amplamente distribuído na América do Norte e somente
parasitando este hospedeiro, D. drepanocercus foi relatado no Brasil, na América Central e na Índia (SEIXAS et al., 2004) e D. dipsaci está mundialmente distribuído principalmente em regiões temperadas e subtropicais (STURHAN e BRZESKI, 1991). A distribuição de D. gallaeformans por ambientes com características ecológicas e climáticas bem diferentes indica que o nematóide possui ampla capacidade de adaptação as diferentes condições de ambiente, o que aumenta as chances de sucesso no caso de uma futura introdução no Havaí. As condições climáticas observadas no Havaí são semelhantes às observadas em regiões tropicais do Brasil (SEIXAS, 2002).
D. gallaeformans foi mais severo tanto no campo como em casa de vegetação e
em plantas do gênero Miconia provocando galhas foliares que paralisaram completamente o desenvolvimento da região apical dos ramos. Isto foi observado pelos resultados obtidos no teste de gama de hospedeiros entre as plantas da família Melastomataceae. As que desenvolveram os sintomas severos e apresentaram maior taxa de reprodução do nematóide foram as do gênero Miconia. No campo foi comum observar a ocorrência de galhas em inflorescências, o que reduziu drasticamente a frutificação. Orr et al. (1975) observaram que D. phyllobius reduziu em 75% a taxa de frutificação de S. elaeagnifolium. Este é um fato relevante, pois a alta taxa de produção de sementes é uma das principais características responsáveis pela dificuldade de controle das melastomatáceas em áreas onde elas são invasoras (MEYER, 1994; MEDEIROS et al., 1997).
caso da introdução para não promover a entrada de outra espécie-problema nas ilhas do Pacífico.
Estes resultados permitem concluir que a gama de hospedeiros de
D. gallaeformans foi restrita a plantas da família Melastomataceae e aos gêneros Miconia, Leandra e Clidemia. O gênero Miconia mostrou-se como o mais suscetível. D. gallaeformans foi capaz de infectar e desenvolver os sintomas em plantas de M. calvescens tanto no biótipo encontrado no Brasil como no do Havaí. O
REFERÊNCIAS BIBLIOGRÁFICAS
DIETRICH, A. Evaluation of pathogens from Costa Rica for biological control of
Miconia calvescens in Hawaii. 2006. 115 f. Diplomarbeit im Studiengang Biologie
Leibniz Universität Hannover, Universidad de Costa Rica, Escuela de Biología, San Pedro, San José, Costa Rica.
KAISER, B. A. Economic impacts of non-indigenous species: Miconia and the Hawaiian economy. Euphytica, v. 148, p. 135-150, 2006.
MEDEIROS, A. C.; LOOPE, L. L.; MCELVANEY, S. Status, ecology, and management of the invasive plant, Miconia calvescens DC (Melastomataceae) in the Hawaiian Islands. Records of the Hawaii Biological Survey for 1996. Bishop Museum
Occasional Papers, v. 48, p. 23-36, 1997.
MEYER, J. Y. Mecanismes d'invasion de Miconia calvescens en Polynesie Français. Ph.D. Thesis. L'Université de Montpellier II Sciences et Techniques du Languedoc. Montpellier, France, 1994.
MEYER, J. Y. Observations on the reproductive biology of Miconia calvescens DC. (Melastomataceae), an alien invasive tree on the islands of Tahiti (South Pacific Ocean).
Biotropica, v. 30, n. 4, p. 609-624, 1998.
OLIVEIRA, R. D. L.; BARRETO, R. W.; SANTIN, A. M. A new species of
Ditylenchus causing galls on the invasive weed Miconia calvescens. In: ANNUAL
MEETING OF THE ORGANIZATION OF NEMATOLOGIST OF TROPICAL AMERICA (ONTA), 37., 2005. Viña Del Mar, Chile. Resumos, Viña Del Mar, Chile, 2005. 90 p.
ORR, C. C.; ABERNATHY, J .R.; HUDSPETH, E. B. Nothanguina phyllobia, a nematode parasite of silver-leaf nightshade. Plant Disease Reporter, v. 59, p. 416-418, 1975.
PARKER, P. E. Nematodes as biological control agents of weeds. New York: Chapman Hall, 1991. p. 58-68.
ROBINSON, A. F.; ORR, C. C.; ABERNATHY, J. R. Distribution of Nothanguina
phyllobia and its potential as a biological control agent for silver-leaf nightshade. Journal of Nematology, v. 10, n. 4, p. 362-367, 1978.
ROBINSON, A. F.; ORR, C. C.; ABERNATHY, J. R. Behavioral response of
Nothonguina phyllobia to selected plant species. Journal of Nematology, v. 11, n. 1,
p. 73-77, 1979.
SEIXAS, C. D. S. Controle biológico de Miconia calvescens (Melastomataceae) com
fitopatógenos. 2002. 123 f. Tese (Doutorado em Fitopatologia) – Universidade Federal
de Viçosa, Viçosa, 2002.
SEIXAS, C. D. S.; BARRETO, R. W.; FREITAS, L. G. Ditylenchus drepanocercus (Nematoda), a potential biological control agent for Miconia calvescens (Melastomataceae): host-specificity and epidemiology. Biological Control, v. 31, n. 1 p. 29-37, 2004.
SIMBERLOFF, D.; STILING, P. Risks of species introduced for biological control.
Biological Conservation, v. 78, p. 185-192, 1996.
STURHAN, D.; BRZESKI, M. W. Stem and bulb nematodes, Ditylenchus spp. In NICKLE, W. R. (Ed.). Manual of agricultural Nematology. New York, 1991. p. 423-459.
WAPSHERE, A. J. A strategy for evaluating the safety of organisms for biological weed control. Ann. App. Biol., v. 77, p. 201-211, 1974.
WATSON A. K. Host range of, and plant reaction to, Subanguina picridis. Journal of
CAPÍTULO 2
HISTOPATOLOGIA DE GALHAS INDUZIDAS POR Ditylenchus
gallaeformans sp. n. EM FOLHAS DE Miconia ibaguensis
1. INTRODUÇÃO
Miconia ibaguensis Triana, membro da família Melastomataceae, é nativa do
Brasil e encontrada em regiões de Cerrado e Mata Atlântica. As folhas das Melastomataceae são geralmente pilosas, opostas ou opostas-cruzadas e pecioladas, com lâminas inteiras, lanceoladas, ovaladas ou oblongas e com margens lisas ou serradas. Os tricomas, de formas variadas e complexas, constituem importante auxílio na identificação dos gêneros e das espécies dessa família, sendo rara a ocorrência de tricomas simples. A epiderme, muitas vezes, é composta por células mucilaginosas e é freqüente a presença de hipoderme (REIS et al., 2005).
Nematóides membros da família Anguinidae, estudados como possíveis agentes de controle biológico de plantas apresentam como característica comum a capacidade de induzir em seus hospedeiros deformações e galhas nos tecidos aéreos (PARKER, 1991). Entre estes se destacam Anguina amsinckia (Steiner e Scott) Thorne, responsável pelo desenvolvimento de galhas em folhas e inflorescências de Amsinckia intermedia Fischer e Meyer (PANTONE, 1987), Subanguina picridis (Kirjanova) Brzeski, que forma galhas em folhas de membros da família Asteraceae (WATSON, 1986) e Ditylenchus
descritos acima, apresentam como características comuns a formação de uma cavidade central onde se alojam os nematóides; estas cavidades são rodeadas por células nutritivas da qual o nematóide obtém alimento, e é comum em todos os casos, o desenvolvimento vasto dos tecidos vasculares no entorno das cavidades e a formação das galhas em tecidos meristemáticos (GODFREY, 1940; ORR et al., 1975; WATSON, 1986). Galhas foliares, formadas por nematóides, são caracterizadas por apresentarem hipertrofia e proliferação desordenadas de células do mesofilo e apresentam cavidades centrais onde os nematóides se alojam (MANI, 1964).
A formação de galhas representa uma reação da planta diante da invasão de um agente biótico, caracterizando uma extrema especialização entre os dois organismos. No caso de galhas formadas por nematóides, o desenvolvimento destas traz certos benefícios para o parasita, pois servem de abrigo, fornecem alimento e, em alguns casos, auxiliam na dispersão para novas áreas (PARKER, 1991).
Em 2005 foi observada em Viçosa, MG, a ocorrência de galhas em folhas e inflorescências de melastomatáceas, com destaque para M. ibaguensis devido à severidade e abundância dos sintomas sobre este hospedeiro. Em estudos posteriores ficou determinado que Ditylenchus gallaeformans sp. n. era o agente etiológico destas galhas.
2. MATERIAL E MÉTODOS
2.1. Coleta e processamento do material coletado no campo
2.2. Histopatologia de folhas de M. ibaguensis inoculadas com Ditylenchus
gallaeformans
3. RESULTADOS
As características anatômicas das galhas provenientes de plantas coletadas no campo e das inoculadas em condições controladas foram semelhantes e, portanto, os resultados serão apresentados em conjunto.
Anatomia de folhas sadias: a epiderme é formada por uma única camada de células em ambas as faces, sendo as células da face adaxial maiores que as da face abaxial e variavam de quadradas a retangulares, nos cortes transversais. A cutícula era, em geral, mais espessa na face adaxial. Foi observada a presença de tricomas ramificados classificados como “tricomas estrelados pedunculados” e emergências formadas, por células epidérmicas e parenquimáticas. O parênquima lacunoso apresentava arranjo compacto com poucos espaços intercelulares e ocupava aproximadamente 70 % da espessura do mesofilo; já o parênquima paliçádico ocupava aproximadamente 30 % da espessura do mesofilo. Constantemente era observada a presença de drusas nas células do mesofilo. O sistema vascular, na nervura central, apresentava-se em forma de arco constituído por uma unidade maior, em forma de arco aberto e por duas ou três unidades menores, formando feixes bicolaterais (Figura 1), enquanto os feixes vasculares de menor porte eram colaterais.
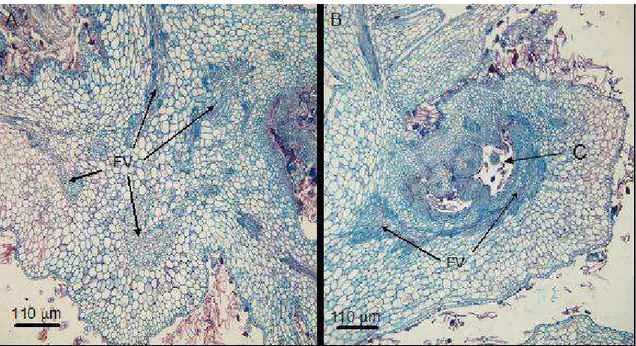
Figura 1 – Seções transversais de folhas adultas sadias de Miconia ibaguensis. A = região mediana da folha com destaque para nervura central; B = lâmina foliar; EP = epiderme; Eb = epiderme abaxial; Ed = epiderme adaxial; TR = tricoma; FL = floema; XL = xilema; EM = emergência; e M = mesofilo.
As galhas apresentavam vasto desenvolvimento do sistema vascular, que se encontrava desorganizado quando comparado ao de folhas sadias, sofrendo redirecionamento para o centro das galhas (Figura 2(A e B)). As galhas apresentavam desenvolvimento contínuo, e em uma mesma galha eram observadas regiões de tecido velho e novo.
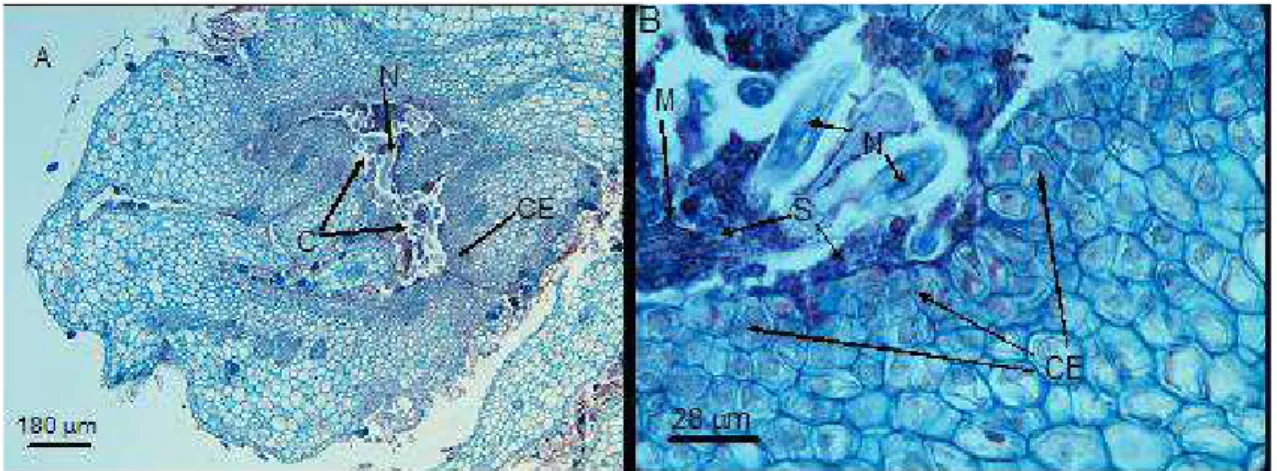
Em uma única galha eram observadas várias criptas que possuem aberturas para o ambiente externo, e no seu entorno as células mostravam-se densas, com muito conteúdo celular e com aspecto meristemático; as criptas apresentam no seu interior substâncias coradas de azul, misturadas a nematóides e micélio fúngico (Figura 3).
Figura 3 – Seções transversais de galhas adultas em folhas de Miconia ibaguensis, infectadas por Ditylenchus gallaeformans. A = aspecto geral da galha mostrando as células ao redor das criptas com muito conteúdo celular; B = detalhe das células do entorno das criptas e da substância encontrada no interior das criptas com a presença de micélio; N = nematóide; C = cripta; CE = células do entorno das criptas; M = micélio fúngico; e S = substâncias coradas de azul no interior das criptas.
Nas galhas de folhas adultas havia aumento no número de tricomas multicelulares, quando comparado com tecidos sadios. Também foi observada a formação de tricomas glandulares pluricelulares, não identificados em folhas sadias (Figura 4).
Figura 4 – Seções transversais de galhas adultas em folhas de Miconia ibaguensis, infectadas por Ditylenchus gallaeformans. A = detalhe do tricoma secretor encontrado no interior das criptas; B = detalhe de células não coloridas na borda das criptas, indicando a formação do meristema de cicatrização; TR = tricoma; C = cripta; e CA = camada de abscisão.
3.1. Cronologia do desenvolvimento das galhas em tecidos inoculados artificialmente
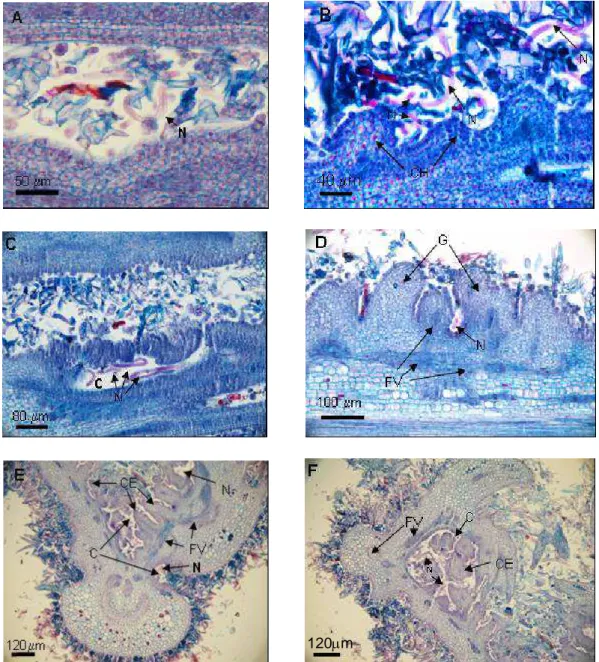
No quinto e décimo dias após a inoculação os nematóides eram observados dispersos pela região apical do ramo, nos espaços existentes entre as folhas primordiais misturados aos tricomas. Não foi observada nenhuma alteração nos tecidos (Figura 5 (A)).
No 15o dia já foram observadas alterações no meristema fundamental dos primórdios foliares. As células começaram a se multiplicar desordenadamente formando invaginações. Foi observada grande concentração dos nematóides em locais próximos às alterações ocorridas no meristema apical (Figura 5(B) e Figura 6).
A formação primária das galhas com as criptas foi observada no 20o dia. Essas são originadas das invaginações provocadas pelo desenvolvimento irregular das células do meristema fundamental. Havia sempre grande quantidade de nematóides com presença também de ovos no interior das criptas. As células do entorno destas cavidades apresentavam conteúdo celular denso (Figura 5(C)).
Figura 5 – Desenvolvimento cronológico das galhas em folhas de Miconia ibaguensis parasitadas por Ditylenchus gallaeformans. A = seção longitudinal do primórdio foliar, mostrando o nematóide entre os tricomas aos cinco dias após a inoculação (DAI); B = seção longitudinal na região do primórdio foliar mostrando o mesofilo sofrendo hiperplasia no inicio do desenvolvimento das galhas, aos 15 DAI; C = seção longitudinal na região do primórdio foliar mostrando a formação típica da galha com cavidades centrais onde os nematóides se concentram aos 20 DAI; D = seção transversal das folhas em início de expansão, mostrando as galhas em estádio avançado de desenvolvimento com criptas habitadas pelos nematóides, e com desenvolvimento dos feixes vasculares no entorno das criptas, aos 25 DAI; E e F = seção transversal da folha completamente deformada pelo nematóide; as células do entorno das criptas apresentam muito conteúdo celular e feixes vasculares bem desenvolvidos, aos 30 e 35 DAI, respectivamente; C = criptas; N = nematóide; CE = células do entorno das criptas; FV = feixes vasculares; G = galha; e CH = células sofrendo hiperplasia.
Figura 6 – Corte longitudinal do meristema apical de Miconia ibaguensis parasitadas por Ditylenchus gallaeformans. Detalhe da concentração dos nematóides, no meristema apical, aos 15 dias após a inoculação. MA = meristema apical; N = nematóide; e PF = primórdio foliar.
regiões da galha distantes dos pontos onde estavam concentrados os nematóides (Figura 5(D)).
4. DISCUSSÃO
Os sintomas observados em folhas de M. ibaguensis seguem os mesmos padrões observados em outras espécies hospedeiras (Capítulo 1); portanto, é possível que as alterações ocorridas na anatomia foliar sejam similares em todos os hospedeiros conhecidos deste nematóide.
É comum em nematóides causadores de galhas aéreas que o desenvolvimento inicial da galha dependa da penetração dos nematóides nos tecidos do meristema fundamental, originando uma cavidade central que abrigará os nematóides por alterações no padrão de diferenciação das folhas. As células desta região sofrem hiperplasia e, em alguns casos, hipertrofia, resultando na formação das galhas (WATSON et al., 1979; SKINNER et al., 1980; WATSON, 1986).
D. gallaeformans não apresentou esta característica de penetração nos tecidos.
Ao que tudo indica ele se apresenta como um ectoparasita migrador, pois, em nenhum momento foi observada a sua penetração nos tecidos para induzir a formação das galhas. No momento inicial da infecção, os nematóides acumulavam-se na região entre os primórdios foliares, protegidos pelos tricomas aí existentes; nestes locais ocorre a formação de um microclima que, provavelmente, é ideal para migração e estabelecimento inicial, além de protegê-los do dessecamento (GODFREY, 1940).
meristema fundamental em plantas atacadas por nematóides de parte aérea, seja provocada pelo aumento da concentração de auxinas nos tecidos próximos aos locais infectados pelos nematóides, como uma forma de resposta enzimática da planta aos danos provocados por este ataque (VIGLIERCHIO, 1971; HARRIS e SHORTHOUSE, 1996).
Ao que tudo indica nos primórdios foliares D. gallaeformans encontra a proteção contra a dessecação por um período de tempo suficiente para formação das galhas, que é, de aproximadamente, 20 dias; além de os tecidos meristemáticos dos primórdios foliares serem mais propícios a este tipo de alterações (GOODEY, 1932; FAWCETT, 1938; GODFREY, 1940; NORTON e SASS, 1966). Estes fatores foram determinantes para o desenvolvimento da doença nos primórdios foliares.
Em galhas foliares induzidas por D. phyllobius, S. picridis e A. Amsinckia, as cavidades centrais onde se alojam os nematóides não apresentavam ligações com o ambiente externo (SKINNER et al., 1980; WATSON, 1986; PANTONE, 1987). Porém, as cavidades formadas em galhas incitadas por D. gallaeformans apresentavam ligação com o meio externo, o que possibilita rápida saída do nematóide destes locais, quando necessária sua migração para novos sítios de infecção.
Foi observado que as células do entorno das criptas onde estavam localizados os nematóides apresentavam grande quantidade de conteúdo celular, indicando intensa atividade celular nestes locais (SKINNER et al., 1980; HARRIS e SHORTHOUSE, 1996). Acredita-se, também, que esteja ocorrendo acúmulo de substâncias nutritivas, como observado por Watson (1986), em folhas de Acroptilon repens atacadas por
S. picridis; as células que circundavam as cavidades onde estavam localizados os
nematóides também exibiam conteúdo celular denso, resultante do acúmulo de fotoassimilados e da proliferação de mitocôndrias e ribossomos, sugerindo que estas eram as células nutritivas usadas para a transferência de alimentos entre a planta e o nematóide. Fenômeno semelhante foi observado em galhas foliares de Solanum
alaeagnifolium infectadas por D. phyllobius (SKINNER et al., 1980). Isso é comum
também nas galhas formadas por insetos (HARRIS e SHORTHOUSE, 1996).