Differential trypanosome surface coat regulation by a CCCH protein that co-associates with procyclin mRNA cis-elements.
Texto
Imagem
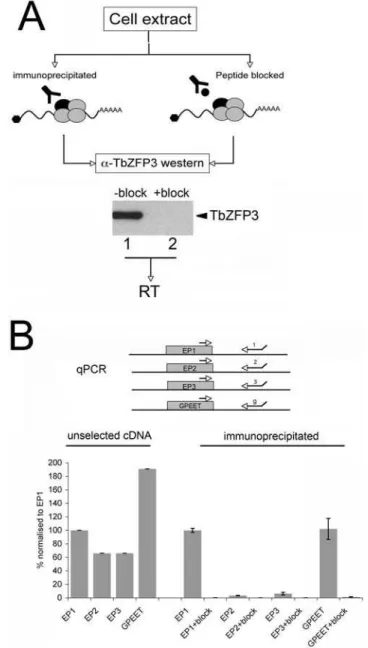
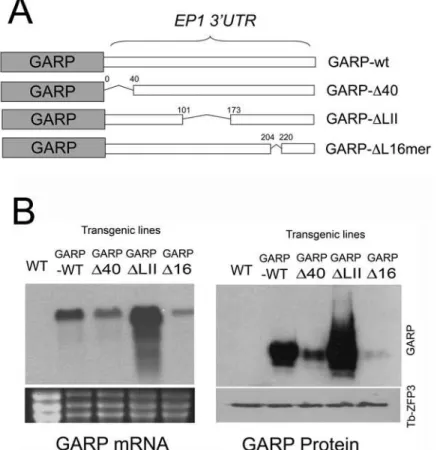
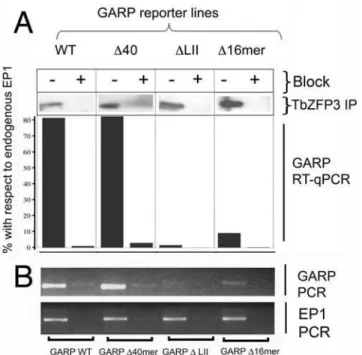
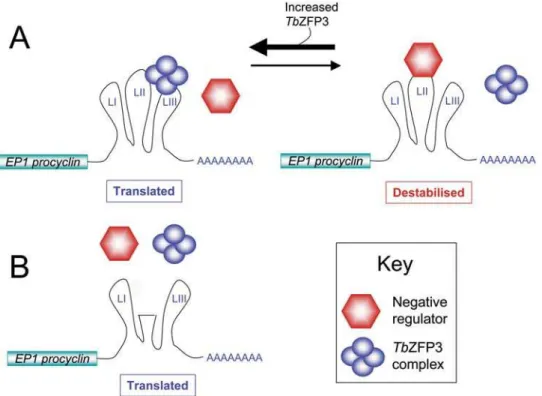
Documentos relacionados
Apesar de existir uma grande tendência tanto na doutrina como na jurisprudência em aceitar que é constitucionalmente admissível obrigar o arguido, em processo
Para tanto, argumentaremos que a demanda por gastos militares é uma propriedade estrutural do sistema internacional cujo comportamento é afetado por fatores presentes na
At a higher concentration of Pb treatment (1000 mg/L), an increased level of antioxidative enzymes activities in both leaf and root tissues suggests that enhanced levels of
Competence, warmth, experience level, competition (rivalry), problem solving, employee relations (manager-employee relationship), interpersonal skills, connectedness,
Following mass spectrometry identification of CSPG, ApoE and cystatin C, we showed by both inhibition of endogenous protein and reconstitution with exoge- nous protein that CSPG
Upon overexpression of miR-124 in Daoy cells, Nur77 mRNA and protein levels and the mRNA levels of Nur77 target genes decreased, showing that miR-124 affects not only Nur77
Interestingly, we found that co-expression of RASSF1C and IGFBP-5 reduced PIWIL1 mRNA and protein levels in lung cancer cell lines, while co-expression of RASSF1A- RASSF1C did not
Using K562 cells as a model, we found that overexpression of THAP11 suppressed hemin induced erythroid differentiation of K562 cells with fewer benzidine-positive cells and lower
