UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DEPARTAMENTO DE ECOLOGIA
Vanessa Rodrigues de Morais
FINDING THE WAY HOME: MOVEMENT OF BUTTERFLIES IN NON-FAMILIAR HABITATS
2
VANESSA RODRIGUES DE MORAIS
FINDING THE WAY HOME: MOVEMENT OF BUTTERFLIES IN NON-FAMILIAR HABITATS
Dissertação apresentada à Coordenação do curso de Pós-graduação em Ecologia, da Universidade Federal do Rio Grande do Norte em cumprimento as exigências para obtenção de Grau de Mestre
Orientador: Prof. Dr. Marcio Zikán Cardoso
3 Seção de Informação e Referência
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede
Morais, Vanessa Rodrigues de.
Finding the way home: movement of butterflies in non-familiar habitats / Vanessa Rodrigues de Morais. – Natal, RN, 2015.
24 f.
Orientador: Márcio Zikán Cardoso.
Dissertação (Mestrado em Ecologia) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Ecologia.
4
O e a annot do right in one department of life whilst he is occupied in doing wrong in any other
depart e t. Life is o e i di isi le hole
5 Sumário
Resumo ... 6
Abstract ... 7
Introduction ... 8
Methods ... 10
Study area ... 10
Study species ... 11
Raising of experimental subjects ... 11
Experimental procedure ... 11
Data analysis ... 13
Results ... 14
Discussion ... 18
Perceptual ranges ... 18
Implications for conservation ... 20
References ... 21
Agradecimentos ... 26
6 Resumo
7 Abstract
Natural landscapes have been dramatically affected by habitat loss and fragmentation, which transform continuous forest in habitat patches imbedded in areas of non-habitat (matrices). These matrices, inhospitable or not, affect countless ecological process, like dispersal. One of the ways to understand this effect of matrix on dispersal is studying animal’s perceptual range, which is a range at which an animal perceive landscape elements. This perception is directly connected to the success to reach a new habitat patch while animals navigate through matrix. We evaluate the habitat perception of Heliconius erato butterfly. However, we were also interest in evaluate the effect of butterflies age and matrix type on its perception. Consequently, we raised butterflies on laboratory and matched with butterflies from forest during a release experiment. To determinate perceptual range, we did releases in two different matrices at three distances from forest (0, 30 and 100 meters) and measured the final angle reached for butterflies. We found that (I) butterflies released in edge were strongly oriented to forest; (II) than higher the release distance the lower perceptual ability and (III) there is an interaction between age and matrix type. Naïve butterflies oriented better on open field (perceptual range: 30-100 meters) and experienced oriented better at coconut plantation (perceptual range: 30-100 meters).
Key-words: Habitat fragmentation, Naïve, Butterfly, Heliconius erato, Perceptual
8 Introduction
Habitat fragmentation is the norm in landscapes dominated by humans. Because of that natural populations tend to persist in subdivided patches (Schelhas & Greenberg 1996). The combined effect of habitat loss and fragmentation is a net increase in distances between remnant habitat patches (Baguette et al. 2013) which ultimately decreases the probability of migrants between patches. In this context, the viability of populations in patchy landscapes depends on the flux of individuals between patches through dispersal or migration (Hanski 1998; Haddad 1999).
Dispersal in animals can be the result of routine movements of an individual, such as foraging, or can derive from special movements such as the establishment of individuals in a new place away from their natal site (Van Dyck & Baguette 2005). In heterogeneous landscapes, this process consists of leaving the natal patch, crossing non-suitable matrix habitat and settling in a new habitat patch (Baguette & Dyck 2007). Success in a dispersal event depends strongly on the ability to cross the matrix. Experimental studies have shown that different types of matrix offer different resistances to movement of organisms and these can vary between species (Haddad 1999; Ries & Debinski 2001; Ricketts 2001; Prevedello & Vieira 2010).
The ability to locate a suitable habitat patch is fundamental for a successful dispersal (Lima & Zollner 1996; Zollner 2000) and can be estimated as perceptual range. The spatial extension of habitat perception is a crude measure of an organism’s capacity to perceive its surroundings. It is considered as the maximum distance in which an animal can perceive the presence of landscape elements (Lima & Zollner 1996; Zollner & Lima 1997). Perceptual range is important because it influences the probability of an individual successfully finding another habitat patch while navigating in inhospitable matrix (Zollner 2000) and is also used in estimates of functional connectivity in landscape ecology (Eycott et al. 2012).
9
The majority of studies on perceptual range use individuals captured at the location where the study is performed and are presumed to be familiar with the native habitat. On the other hand, there are few studies with dispersal behavior that uses naïve individuals (Ovaskainen et al. 2008; Öckinger & Van Dyck 2012; Bergerot et al. 2012). Even so, these studies did not specifically test the effect of age and experience in movement.Understanding how young individuals behave can be interesting because most part of the animals on this age category are more inclined to disperse (Clobert et al. 2001) and their behavior can be quite different from resident individuals that already have some experience with the environment.
In the tropical butterfly Heliconius dispersal is done by young individuals especially on their first week of life (Mallet 1986a). After dispersal, experienced adults tend to stay in areas that contain feeding and oviposition plants where they develop into a daily routine of visiting flowers and hostplants. Because of this, Heliconius butterflies are one of the few invertebrates that have home range behavior (Turner 1971; Mallet 1986b). This means that experienced adults are not likely to disperse after establishing their home range. It also suggests that there may exist distinct behaviors between experienced adults and young naïve butterflies. The consequences of these differences have not been adequately addressed in the literature. Consequently, this study was conducted in order to understand how habitat perception varies between naïve and experienced adults of Heliconius butterflies.
In order to do so we evaluated the influence of two different matrix types and distances from a patch in the butterflies’ perceptual ability. With respect to experience of butterflies, we expect that naïve butterflies will show a higher perceptual range than experienced butterflies. With respect to matrix type, we expect that butterflies in matrices with less visual obstruction will express a higher perceptual range because of easiness for visual orientation (Forero-Medina & Vieira 2009; Prevedello, Forero-Medina & Vieira 2011).
10 Methods
Study area
The study was carried from November 2013 to December 2014 in a landscape composed of a mid-sized 80 ha Atlantic Forest remnant and several agricultural matrix habitats that border the forest patch (5°56’S, 35°11’W) (Fig. 1).
We selected two matrix habitats to perform our experiment: a coconut plantation and open field (grassland). The coconut plantation habitat is characterized by widely spaced Cocos nucifera palms with sparse herbaceous vegetation growing under individual trees. The open field is an area under natural regeneration. It is dominated by grasses and contains a few scattered shrubs and trees. Although both landscape units may contain flowers that could be used by Heliconius, these butterflies are seldom seen in these habitats. Individual butterflies are easily seen flying at the edge of the forest that borders the plantation and the open field.
11
Study species
We chose the species Heliconius erato to test the ability of this butterfly on localizing habitat patches (Figure 2). This species is found in all the Neotropical region and are associated with forested habitats and their edges, rarely living in open areas (Brown 1981). Heliconius butterflies are model organisms in ecology and evolution, easy to recognize in the field and also easy to track because of its slow and predictable flight (Srygley & Chai 1990).
Figure 2. An adult of Heliconius erato phyllis (Source: Ananda Silva).
Raising of experimental subjects
In order to obtain naïve butterflies, females of Heliconius erato were collected from forest near the experimental site and brought to the lab, where they were housed in a large outdoor insectary. Females were provided with flowers of Lantana camara and a solution of water, sugar and amino acid. Fresh branches of the host plant Passiflora cincinnata, were used to obtain eggs and raise larvae. Freshly emerged individuals were sexed, marked on the wing with a non-toxic pen and kept in an insectary before being taken to the field for the experiments. The average number of days spent in the insectary was 5 (min = 1, max = 26).
Experimental procedure
12
release procedure simulates the events a dispersing individual would experience while moving through matrix habitat (Zollner & Lima 1997).
In order to evaluate whether animals orient themselves in the expected bearing, we measured the bearing of butterflies released at different distances from the forest edge into the matrix. In each matrix type (coconut plantation or open field) we released butterflies at three distances from the forest edge: 0, 30 and 100 meters. (Fig. 4). To avoid a potential location bias, we randomized the release “corridor”, an imaginary line containing the three release distances. We created five such “corridors” in the coconut plantation and three in the open field. The latter is smaller than the plantation, thus the smaller number of “corridors”.
Experiments were performed using a pair of butterflies, consisting of a naïve and an experienced butterfly. For each naïve butterfly, an experienced adult of H. erato was collected in situ and used as a control on the experiment. The experimental pairs were preferentially matched for species and sex. In some cases where individuals of H. erato were not available in the field we used H. melpomene. Previous work had shown that the responses of experienced adults do not differ between the two species (Falcão et al., in prep.) and we concluded this would not affect the experimental outcome.
The release procedure involved placing each individual butterfly into a black box (13 x 9 x 7 cm) (Figure 4), which were then stored in a cooler and transported to the release point. The black box was used to avoid individuals (especially naïve butterflies) using visual cues during the transport.
13
In order to release the experimental subject, the box was placed on the release point on a 1.3 m platform. A line, attached to the box lid was gently pulled and the butterfly moved out of the box. Observations started as the butterfly flew from the box. Two observers followed the butterfly at a safe distance (at least 5 m) and noted their behavior. At the end of the observation period the final angle and the flight time were recorded. The observation lasted until the butterfly (1) reached the edge, (2) was lost from sight or (3) remained inactive (e.g. landed on a shrub or tree in the matrix) for more than 10 minutes.
Important for our work is the assumption that once animals face an inhospitable matrix, they will move toward their habitat (patch). Therefore, we assume the directional movement will reveal their capability of perceiving the habitat patch (Zollner & Lima 1997; Prevedello et al. 2010; Sozio et al. 2013).
We standardized our release procedure as much as possible. The sequence of release points and order of release were randomized. Similarly, the same was done regarding the direction of lid opening (0º, 90º, 180º e 270º) using the magnetic north as reference. Releases were performed under similar weather conditions (sunny days) and between 8:00 – 11:00 a.m.
Data analysis
14 Results
We released 190 butterflies, half of it lab-raised naïve individuals matched with field-caught, experienced adults. Forty-five pairs were tested in the coconut plantation and 50 pairs were tested in the open field. Heliconius melpomene was used in 24 releases (always experienced controls).
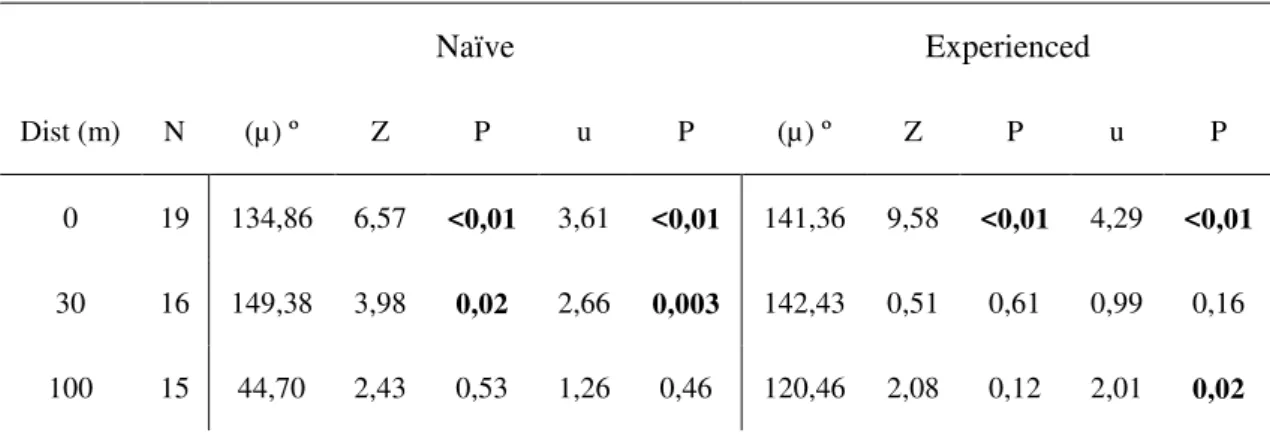
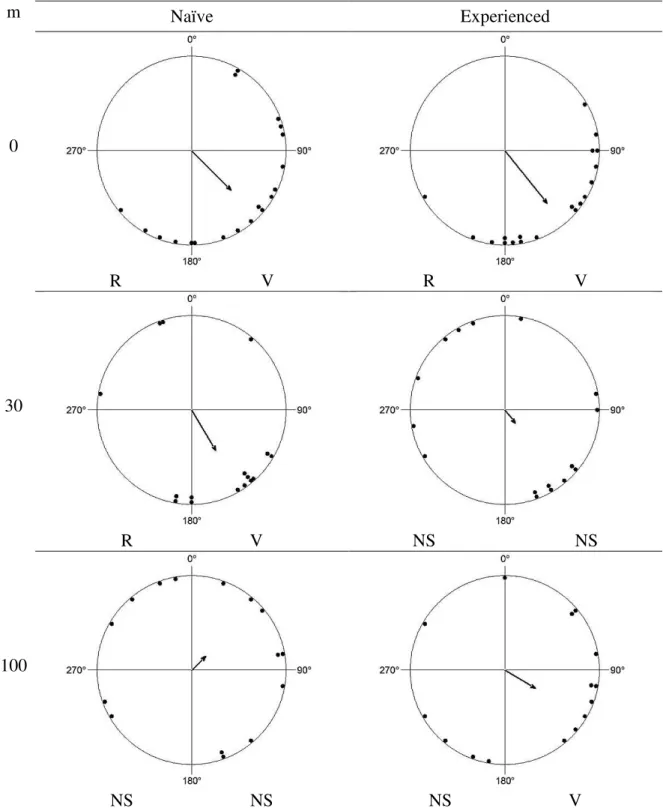
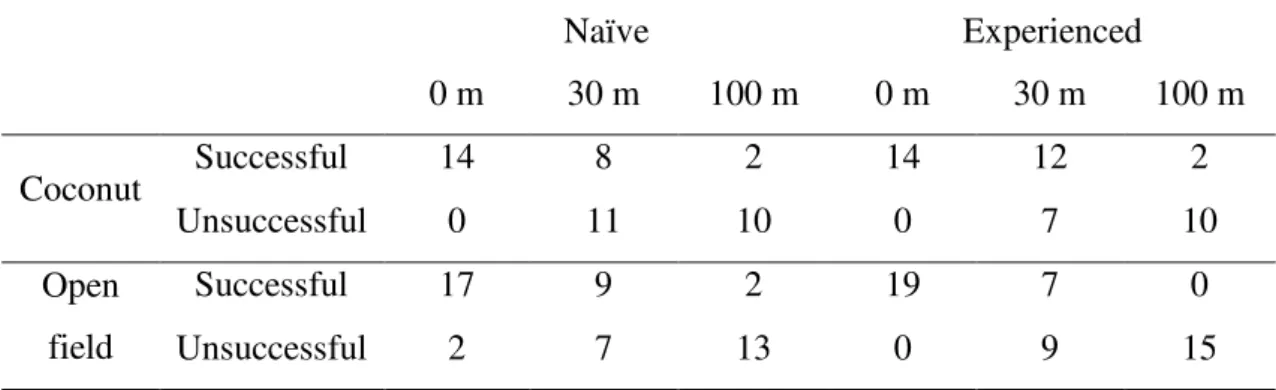
Distance played a significant role in shaping flight directionality in naïve and experienced butterflies in both landscape units (Tables 1 and 2). Naïve butterflies tested in the coconut plantation successfully oriented towards the habitat patch when released at the border (d = 0 m, V-test: u = 3,591, P <<0.01) (Fig. 5). Yet, at d = 30 and 100 m, naïve butterflies failed to orient in the expected direction (V-test: u= -0,778, NS and u= -0,88, NS, respectively) (Fig. 5). In the open field experiments naïve butterflies successfully oriented towards the habitat patch both at the edge and at 30 m away from the habitat patch (V-test: u = 3,611, P <<0.01 and u = 2,661, P = 0.003, respectively) (Fig. 6). Individuals released at 100 m failed to orient themselves in the expected direction (V-test: u = 1,26, NS) (Fig. 6).
Responses of the experienced butterflies were consistent with previous findings (Falcão et al, in prep.). In the coconut plantation, experienced adults successfully oriented towards the habitat patch at the border and at 30 m (V-test: u = 3,032, P << 0.01 and u= 2,087, P = 0.02, respectively) (Fig. 5) but failed to orient properly at 100 m (Vtest: u = -1,191, NS) (Fig. 5). Responses of experienced butterflies in the open field were less consistent. As before, releases at the border produced significant orientation towards the patch (V-test: u = 4,29, P << 0.01) (Figure 6). Individuals released at 30 m failed to orient themselves towards the patch (V-test: u = 0,99, NS) (Fig. 6), while individuals released at 100 m successfully oriented toward the forest (V-test, u = 2,01, P = 0.02) (Fig.6).
15
Table 1 – Flight directedness of butterflies released on coconut plantation. N= sample
size (number of individuals); µ = mean vector (mean angle). Rayleigh test (Z) and P-value associated. V-test (u) and P-P-value associated. Statistically significant results are in bold.
Naïve Experienced
Dist (m) N (µ) ° Z P u P (µ) ° Z P u P
0 14 161,92 8,95 <0,01 3,59 <0,01 177,22 9,96 <0,01 3,03 <0,01
30 19 267,85 0,55 0,58 -0,78 0,78 162,80 3,08 0,04 2,09 0,02
100 12 249,46° 1,61 0,20 -0,88 0,81 300,70 0,73 0,49 -1,19 0,88
Table 2 – Flight directedness of butterflies released on open field. N= sample size (number of individuals); µ = mean vector (mean angle). Rayleigh test (Z) and P-value associated. V-test (u) and P-value associated. Statistically significant results are in bold.
Naïve Experienced
Dist (m) N (µ) º Z P u P (µ) º Z P u P
0 19 134,86 6,57 <0,01 3,61 <0,01 141,36 9,58 <0,01 4,29 <0,01
30 16 149,38 3,98 0,02 2,66 0,003 142,43 0,51 0,61 0,99 0,16
16 Figure 5 – Angular orientations of Heliconius butterflies released in the coconut plantation at 0, 30 and 100 meters from border of habitat patch. Each point around the circle represents the final direction of an individual butterfly. The arrow represents the mean vector. The expected bearing of a butterfly successfully orienting to the habitat patch is approximately 130º. Under each graph letter R represent significant results for Rayleigh test, and V represent significant results for V test. NS means No Significant result.
m Naïve Experienced
0
R V R V
30
NS NS R V
100
17 Figure 6 – Angular orientations of Heliconius butterflies released in the open field at 0, 30 and 100 meters from border of habitat patch. Each point around the circle represents the final direction of an individual butterfly. The arrow represents the mean vector. The expected bearing of a butterfly successfully orienting to the habitat patch is approximately 130º. Under each graph letter R represent significant results for Rayleigh test, and V represent significant results for V test. NS means No Significant result.
m Naïve Experienced
0
R V R V
30
R V NS NS
100
18 Discussion
Some important patterns emerged from our release experiments. Butterflies released at the edge invariably chose the correct path to the patch. At intermediate distances, perception is influenced by matrix type. Naïve individuals showed more orientation in open field than in the coconut plantation, while experienced individuals show the opposite behavior. Finally, there is a clear effect of distance on the ability to choose the correct direction, i.e., habitat perception is distance dependent.
On edge releases the bearing pattern to the habitat repeated in all moments, independent from matrix type. This result validate our assumption that when butterflies perceive the habitat they move towards it. From 66 releases on edge, only two naïve butterflies, when released on field under regeneration, have not oriented to habitat. This preference from patch on edge releases was already demonstrated and used to confirm the same assumption in butterflies (Merckx & Van Dyck 2007), marsupials (Forero-Medina & Vieira 2009; Prevedello et al. 2010), mice’s (Sozio et al. 2013) and lizards (Auburn et al. 2009).
Perceptual ranges
With this experiment we found that matrix influences the range of habitat perception in accordance with previous studies (Haddad 1999; Ries & Debinski 2001; Ricketts 2001; Prevedello & Vieira 2010; Prevedello et al. 2011; Sozio et al. 2013). Our experiment also shows that matrix type interact with butterfly’s experience. Accordingly, naïve butterflies oriented better at open field while experienced butterflies oriented better on coconut plantation. This differed from our initial expectations that naïve butterflies would have better perceptual ranges than experienced butterflies.
Naïve individuals have an estimated perceptual range within 0 - 30 m on coconut plantation and 30 - 100 m on open field. Experienced individuals have a perceptual range within 30 - 100 m on coconut plantation and within 0 - 30 m on open field. But with an unexpected directedness to patch at 100 meters. Further studies should focus on this interval between 30 and 100 meters, to a better determination of perceptual range.
19
2013). On open field, they can easily see where the patch is, while on coconut plantation the visual obstruction is higher. In this case. the matrix with less vegetation height revealed the higher perceptual range, as found for marsupials (Forero-Medina & Vieira 2009; Prevedello et al. 2011). This suggests that visual elements play an important part in the orientation process.
We could hastily conclude that coconut plantation is worse than open field to the movement of naïve butterflies. However, we can also speculate that the lack of orientation on coconut plantation is because naïve butterflies faced more amenable environmental conditions that could be a positive motivation to explore the matrix rather than to adopt an escape behavior.
For dispersing organisms realizing short movements, matrices with less vegetation obstruction will facilitate habitat location. However, it is not realistic to think that organisms realizing long dispersal movements will prefer this type of matrix, which offer more mortality risks. When they became exhausted and starved, while look for a suitable habitat, they will probably look for a matrix more structurally similar to their habitat (Prevedello & Vieira 2010; Prevedello et al. 2011).
For experienced butterflies, the matrix with higher structural complexity allowed the expression of a higher perceptual range. We suspect that this is a consequence of home range behavior (Turner 1971), which likely makes the butterfly more familiar not only with its natal patch but also with the surrounding habitats. The fact that we found marked butterflies in the forest suggests that there is a mechanism used by the butterflies to navigate. It is also possible that in highly fragmented landscapes composed by small patches individuals find the edge that separate patch from matrix more often (Ries & Debinski 2001) and that they enter the matrix during routine movements. Since the coconut plantation is more shaded than the open field and may be more amenable to movement, perhaps exploitative movements concentrate in this kind of matrix rather than in the open field. In this case, the matrix most similar to habitat would give the chance for more a successful dispersal event (Ricketts 2001; Eycott et al. 2012). This could be tested by observing the frequency of spontaneous crossing between the habitat and the two matrix habitats.
20
the expected direction, they failed to reach the proper patch. This would be more in line with our previous findings (Falcão et al, in prep.), namely that individual at 100 m fail to orient properly to the patch.
Estimates of perceptual range show that it varies widely among taxa. This is mostly because species have peculiar characteristics and different life histories. In butterflies, studies estimating perceptual ability have shown ranges that go from less than 50 m to as much as 150 m, depending on the species (Harrison 1989; Conradt et al. 2000; Cant et al. 2005; Merckx & Van Dyck 2007; Öckinger & Van Dyck 2012). Different population may also express different perceptual abilities. In the case of the European butterfly Pararge aergeria it was found that differences depend on landscape of origin. Individuals from fragmented landscape had an estimated perceptual range higher (100 m) than those from continuous landscape (50 m) (Merckx & Van Dyck 2007). Our study shows that experience and habitat play roles in perceptual ranges.
Implications for conservation
Current research in landscape ecology tries to elucidate how the matrix affects the dynamics of populations and individual organisms. Dispersal between patches depends on matrix quality, once that matrix can positively or negatively affect the existing effects of isolation (Turner, Gardner & O’Neill 2001).
Brazil’s Atlantic forest is a very fragmented biome, mostly composed of relatively small patches (Ribeiro et al. 2009). Thus, animal populations are threatened with isolation problems that include environmental and genetic stochastic effects. Furthermore, the matrix between these patches is a barrier to movement for most of the organisms. Although previous studies had shown the importance of the matrix for butterfly movement few studies have included this element in their experimental design (Sweaney, Lindenmayer & Driscoll 2014).
The determination of perceptual range of species is one step to improve this gap because it strongly influences the success of a dispersant organism while moving through the matrix. In theory, the higher the perceptual range of an organism the lower the risk of mortality, because it increases the chances of finding a suitable habitat and consequently spend less energy looking for it (Lima & Zollner 1996; Zollner & Lima 1997, 2005).
21
will be useful to determine the distance that should exist between patches or stepping-stones, in a range that facilitate disperser’s movement. Also, it can be used in models of dispersal that uses organisms perspective in a more realistic way (Pe’er & Kramer-Schadt 2008; Bergerot et al. 2012; Severns, McIntire & Schultz 2013).
Our work was the first to estimate the perceptual range of a tropical butterfly. It was a glimpse of how Heliconius erato butterflies are oriented on some of the matrix types that composes the landscape in our study area. However, it raises new questions about the insect background: what is the navigational mechanisms that they use? How the cognition works on orientation? (Collett & Graham 2004; Wystrach & Graham 2012). These mechanisms of animal navigation are well known specially for insects like honeybees (Menzel et al. 2000, 2005; Najera, McCullough & Jander 2015) and ants (Zeil, Narendra & Stürzl 2014), and have recently been studied in invasive snakes (Pittman et al. 2014). Nevertheless, this avenue can be better explored for butterflies, which is an opportunity for further studies.
References
Auburn, Z.M., Bull, C.M. & Kerr, G.D. (2009) The visual perceptual range of a lizard, Tiliqua rugosa. Journal of Ethology, 27, 75–81.
Baguette, M., Blanchet, S., Legrand, D., Stevens, V.M. & Turlure, C. (2013) Individual dispersal, landscape connectivity and ecological networks. Biological Reviews, 88, 310–326.
Baguette, M. & Dyck, H.V. (2007) Landscape connectivity and animal behavior: functional grain as a key determinant for dispersal. Landscape Ecology, 22, 1117– 1129.
Batschelet, E. (1981) Circular Statistics in Biology. Academic Press Inc.
22
Bridgman, L.J., Benitez, V.V., Grana Grilli, M., Mufato, N., Acosta, D. & Laura Guichon, M. (2012) Short perceptual range and yet successful invasion of a fragmented landscape: the case of the red-bellied tree squirrel (Callosciurus erythraeus) in Argentina. Landscape Ecology, 27, 633–640.
Brown, K.S. (1981) The Biology of Heliconius and Related Genera. Annual Review of Entomology, 26, 427–457.
Cant, E.T., Smith, A.D., Reynolds, D.R. & Osborne, J.L. (2005) Tracking butterfly flight paths across the landscape with harmonic radar. Proceedings of the Royal Society B-Biological Sciences, 272, 785–790.
Carlsson, M.A., Schäpers, A., Nässel, D.R. & Janz, N. (2013) Organization of the olfactory system of nymphalidae butterflies. Chemical Senses, 38, 355–367. Clobert, J., Danchin, E., Dhondt, A.A. & Nichols, J.D. (eds). (2001) Dispersal. Also
available as: Hardback.
Collett, T.S. & Graham, P. (2004) Animal Navigation: Path Integration, Visual Landmarks and Cognitive Maps. Current Biology, 14, R475–R477.
Conradt, L., Bodsworth, E.J., Roper, T.J. & Thomas, C.D. (2000) Non-Random Dispersal in the Butterfly Maniola jurtina: Implications for Metapopulation Models. Proceedings: Biological Sciences, 267, 1505–1510.
Van Dyck, H. & Baguette, M. (2005) Dispersal behaviour in fragmented landscapes: Routine or special movements? Basic and Applied Ecology, 6, 535–545.
Eycott, A.E., Stewart, G.B., Buyung-Ali, L.M., Bowler, D.E., Watts, K. & Pullin, A.S. (2012) A meta-analysis on the impact of different matrix structures on species movement rates. Landscape Ecology, 27, 1263–1278.
Forero-Medina, G. & Vieira, M.V. (2009) Perception of a fragmented landscape by neotropical marsupials: effects of body mass and environmental variables. Journal of Tropical Ecology, 25, 53–62.
23
Hanski, I. (1998) Metapopulation dynamics. Nature, 396, 41–49.
Harrison, S. (1989) Long-Distance Dispersal and Colonization in the Bay Checkerspot Butterfly, Euphydryas Editha Bayensis. Ecology, 70, 1236–1243.
Kovach, W.L. (2011) Circular Staistics for Windows.
Lima, S.L. & Zollner, P.A. (1996) Towards a behavioral ecology of ecological landscapes. Trends in Ecology & Evolution, 11, 131–135.
Mallet, J. (1986a) Gregarious roosting and home range in Heliconius butterflies. National Geographic Research, 2, 198–215.
Mallet, J. (1986b) Dispersal and gene flow in a butterfly with home range behavior: Heliconius erato (Lepidoptera: Nymphalidae). Oecologia, 68, 210–217.
Menzel, R., Brandt, R., Gumbert, A., Komischke, B. & Kunze, J. (2000) Two spatial memories for honeybee navigation. Proceedings of the Royal Society of London B: Biological Sciences, 267, 961–968.
Menzel, R., Greggers, U., Smith, A., Berger, S., Brandt, R., Brunke, S., Bundrock, G., Hülse, S., Plümpe, T., Schaupp, F., Schüttler, E., Stach, S., Stindt, J., Stollhoff, N. & Watzl, S. (2005) Honey bees navigate according to a map-like spatial memory. Proceedings of the National Academy of Sciences of the United States of America,
102, 3040–3045.
Merckx, T. & Van Dyck, H. (2007) Habitat fragmentation affects habitat-finding ability of the speckled wood butterfly, Pararge aegeria L. Animal Behaviour, 74, 1029– 1037.
Najera, D.A., McCullough, E.L. & Jander, R. (2015) Honeybees Use Celestial and/or Terrestrial Compass Cues for Inter-Patch Navigation. Ethology, 121, 94–102. Öckinger, E. & Van Dyck, H. (2012) Landscape Structure Shapes Habitat Finding Ability
in a Butterfly. PLoS ONE, 7, e41517.
24
radar reveals an effect of population age on movement distance. Proceedings of the National Academy of Sciences, 105, 19090–19095.
Pe’er, G. & Kramer-Schadt, S. (2008) Incorporating the perceptual range of animals into connectivity models. Ecological Modelling, 213, 73–85.
Pittman, S.E., Hart, K.M., Cherkiss, M.S., Snow, R.W., Fujisaki, I., Smith, B.J., Mazzotti, F.J. & Dorcas, M.E. (2014) Homing of invasive Burmese pythons in South Florida: evidence for map and compass senses in snakes. Biology Letters, 10, 20140040.
Prevedello, J.A., Forero-Medina, G. & Vieira, M.V. (2010) Movement behaviour within and beyond perceptual ranges in three small mammals: effects of matrix type and body mass. Journal of Animal Ecology, 79, 1315–1323.
Prevedello, J.A., Forero-Medina, G. & Vieira, M.V. (2011) Does land use affect perceptual range? Evidence from two marsupials of the Atlantic Forest. Journal of Zoology, 284, 53–59.
Prevedello, J.A. & Vieira, M.V. (2010) Does the type of matrix matter? A quantitative review of the evidence. Biodiversity and Conservation, 19, 1205–1223.
Ribeiro, M.C., Metzger, J.P., Martensen, A.C., Ponzoni, F.J. & Hirota, M.M. (2009) The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation, 142, 1141– 1153.
Ricketts, T.H. (2001) The Matrix Matters: Effective Isolation in Fragmented Landscapes. The American Naturalist, 158, 87–99.
Ries, L. & Debinski, D.M. (2001) Butterfly responses to habitat edges in the highly fragmented prairies of Central Iowa. Journal of Animal Ecology, 70, 840–852. Schelhas, J. & Greenberg, R. (1996) Forest Patches in Tropical Landscapes. Island Press,
25
Severns, P.M., McIntire, E.J.B. & Schultz, C.B. (2013) Evaluating functional connectivity with matrix behavior uncertainty for an endangered butterfly. Landscape Ecology, 28, 559–569.
Sozio, G., Mortelliti, A. & Boitani, L. (2013) Mice on the move: Wheat rows as a means to increase permeability in agricultural landscapes. Biological Conservation, 165, 198–202.
Srygley, R.B. & Chai, P. (1990) Flight morphology of Neotropical butterflies: palatability and distribution of mass to the thorax and abdomen. Oecologia, 84, 491–499. Sweaney, N., Lindenmayer, D.B. & Driscoll, D.A. (2014) Is the matrix important to
butterflies in fragmented landscapes? Journal of Insect Conservation, 18, 283– 294.
Turner, J.R.G. (1971) Experiments on the Demography of Tropical Butterflies. II. Longevity and Home-Range Behaviour in Heliconius erato. Biotropica, 3, 21. Turner, M.G., Gardner, R.H. & O’Neill, R.V. (2001) Landscape Ecology in Theory and
Practice: Pattern and Process.
Wystrach, A. & Graham, P. (2012) What can we learn from studies of insect navigation? Animal Behaviour, 84, 13–20.
Zeil, J., Narendra, A. & Stürzl, W. (2014) Looking and homing: how displaced ants decide where to go. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 369, 20130034.
Zollner, P.A. (2000) Comparing the landscape level perceptual abilities of forest sciurids in fragmented agricultural landscapes. Landscape Ecology, 15, 523–533.
Zollner, P.A. & Lima, S.L. (1997) Landscape-level perceptual abilities in white-footed mice: perceptual range and the detection of forested habitat. Oikos, 80, 51–60. Zollner, P.A. & Lima, S.L. (2005) Behavioral tradeoffs when dispersing across a patchy
26 Agradecimentos
Eu gostaria de agradecer os professores da Pós-Graduação em Ecologia da Universidade Federal do Rio Grande do Norte por todos os ensinamentos compartilhados. Ao CNPQ por fomentar a pesquisa cientifica e por minha bolsa de mestrado. Gostaria de agradecer imensamente a todos que ajudaram na realização deste trabalho. Meu orientador Márcio Zikán Cardoso, pela paciência e incentivo. Aos amigos e colegas Ananda Silva, Leonardo Machado, Giesta George, Larissa Santos, João Gabriel e Victor Hugo Gurgel por toda ajuda em campo e laboratório, pois sem vocês esses resultados não existiriam. Aos meus familiares e amigos do peito (Roseli (Mãe), Odete (Vó), Anne (irmã), Ananda Silva, Natália Ross, Talynne Lopes e Vandir Vilar) agradeço pelo amor e amizade. Sou imensamente grata a todas essas pessoas mencionadas aqui e também aquelas tantas que não foram mencionadas mas sabem que afetam positivamente minha vida.
27
Appendix (supplementary material)
Table 3 –Number of individuals used on release experiments
Coconut
Distance Naïve Experienced
0 H. erato Male H. erato Female 10 4
H. erato Male 5
H. melpomene Male 3
H. erato Female 2
H. melpomene Female 4
30 H. erato Male H. erato Female 11 8
H. erato Male 14
H. melpomene Male 1
H. erato Female 4
H. melpomene Female 0
100 H. erato Male
H. erato Female
5 7
H. erato Male 4
H. melpomene Male 3
H. erato Female 4
H. melpomene Female 1
Open Field
Distance Naïve Experienced
0 H. erato Male H. erato Female 11 8
H. erato Male 10
H. melpomene Male 2
H. erato Female 5
H. melpomene Female 2
30 H. erato Male H. erato Female 7 9
H. erato Male 9
H. melpomene Male 0
H. erato Female 6
H. melpomene Female 2
100 H. erato Male
H. erato Female
4 11
H. erato Male 5
H. melpomene Male 4
H. erato Female 4
H. melpomene Female 2
Table 4 – Number of individuals that successfully reached the target patch
Naïve Experienced
0 m 30 m 100 m 0 m 30 m 100 m
Coconut Successful 14 8 2 14 12 2
Unsuccessful 0 11 10 0 7 10
Open field
Successful 17 9 2 19 7 0