MARCIA CRISTINA CALIXTO
Tese apresentada à Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, para obtenção do título de Doutor em Agronomia, Área de Concentração: Fitotecnia
P I R A C I C A B A Estado de São Paulo - Brasil
MARCIA CRISTINA CALIXTO
Engenheiro Agrônomo
Orientador: Prof. Dr. FRANCISCO DE ASSIS ALVES MOURÃO FILHO
Tese apresentada à Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, para obtenção do título de Doutor em Agronomia, Área de Concentração: Fitotecnia.
P I R A C I C A B A Estado de São Paulo - Brasil
DadosInternacionais de Catalogação na Publicação (CIP) DIVISÃO DE BIBLIOTECA E DOCUMENTAÇÃO - ESALQ/ USP
C a lixto , Ma rc ia C ristina
Hib rid a ç ã o so má tic a e ntre C itrus sine nsis e C .g ra nd is / Ma rc ia C ristina C a lixto . - - Pira c ic a b a , 2003.
99 p . : il.
Te se (d o uto ra d o ) - Esc o la Sup e rio r d e Ag ric ultura Luiz d e Q ue iro z, 2003. Bib lio g ra fia .
1. C itric ultura 2. G e né tic a ve g e ta l 3. Fusã o d e p ro to p la sto 4. Hib rid a ç ã o ve g e ta l 5. La ra nja 6. Me lho ra me nto g e né tic o ve g e ta l 7. Po rta -e nxe rto s I. Título
C DD 634.3
AGRADEÇO
A Leonor e Benedito (in memorian)
Ana Paula e Marcos Henrique,
Ana Lúcia, Ana Cecília, Amanda Letícia, Vera Lúcia Uilson José
OFEREÇO
Marcelo Anna Alissa
A Deus, pela oportunidade desta vida.
A Universidade de São Paulo, ao Conselho de Pós-Graduação e ao Departamento de Produção Vegetal, pela acolhida.
A todos os professores do Departamento de Produção Vegetal, pelos ensinamentos.
A CAPES, pela concessão da bolsa de estudo.
Ao Prof. Dr. Francisco de Assis Alves Mourão Filho, pela dedicada orientação, pelo apoio incondicional, pelo incentivo e pelo exemplo.
A Profa. Dra. Beatriz Madalena Januzzi Mendes, do Laboratório de
Biotecnologia Vegetal do CENA/USP, por todo apoio e pela atenção durante a realização do trabalho.
Ao Centro APTA Citros “Sylvio Moreira”, na pessoa do Prof. Dr. Jorgino Pompeu Jr., pelo fornecimento do material vegetal utilizado na pesquisa.
A Profa. Dra. Maria Lúcia Carneiro Vieira, do Laboratório de Genética da ESALQ/USP, pela oportunidade de aprendizado, pela disponibilidade e pela atenção.
A Profa. Dra. Adriana Pinheiro Martinelli Rodriguez, pelo apoio e estímulo.
A Carlos Alberto de Oliveira, pela extrema paciência e pelo auxílio nas análises moleculares e contagem de cromossomos.
Ao Dr. Jude W. Grosser, pelos preciosos ensinamentos e pela atenção. A Maria Célia Rodrigues, Elisabete Aparecida Sarkis São João, Luciane Aparecida Lopes, Ivete Monteiro de Almeida, Helena Rodrigues Camargo e Thiago dos Santos Silveira, pelo apoio, convívio e solidariedade.
Aos funcionários David Ulrich, Aparecido Donizete Serrano, José Antônio Volpato e Éder de Araújo Cintra, por todo trabalho dispensado.
Ao Departamento de Fitopatologia, pela análise do solo.
A Alexandra Pavan, em especial, pela cumplicidade e companheirismo. Aos colegas de curso, Sally Ferreira Blat, Maria Cecília de Arruda, Bianca Moschetta de Morais Sarmento, Léa Araújo de Carvalho, Cristiane Gonçalves de Mendonça, Fernando Alves Azevedo, Eduardo Suguino, Norma Virgínia Migone Segovia, por todas as alegrias compartilhadas.
A amiga Lilian Ellen Pino, pela amizade sincera.
Aos colegas Janaynna, Hérica, Joanne, Raquel, Nailton, Alice, Márcio, Vagner, Fernanda, Girlene, Mariza, Gustavo, Carol, pelo convívio.
As amigas Ana Carolina Bruno e Marília Gabriela Salvatti Della Vechia, pelo prazer da nossa amizade.
A estagiária Cristiane Fumagalli de Lima, pelo apoio.
Aos amigos Weliton Antônio de Almeida Bastos e Maria Angélica Pereira de Carvalho Costa, que tanto me ensinaram e me fizeram crescer, num convívio diário de amizade, respeito, solidariedade e incentivo.
SUMÁRIO
Página
LISTA DE FIGURAS ... viii
LISTA DE QUADROS ... ix
LISTA DE TABELAS ... x
RESUMO ... xi
SUMMARY ... xiii
1 INTRODUÇÃO ... 1
2 REVISÃO DE LITERATURA ... 3
2.1 Importância da citricultura no Brasil e no mundo... 3
2.2 Origem e distribuição dos citros ... 5
2.3 Importância da utilização de porta-enxertos na citricultura ... 5
2.3.1 Importância da gomose de Phytophthora na cultura dos citros ... 10
2.4 Melhoramento de porta-enxertos de citros ... 12
2.5 Biotecnologia para o melhoramento genético de citros ... 17
2.6 Hibridação somática em citros ... 19
2.7 Considerações finais ... 32
3 MATERIAL E MÉTODOS ... 34
3.1 Estabelecimento e cultivo de calos embriogênicos... 34
3.2 Estabelecimento e cultivo de células em suspensão... 35
3.3 Seleção de plantas matrizes de C. grandis... 35
3.4 Hibridação somática ... 36
3.4.1 Isolamento de protoplastos ... 36
3.4.3 Fusão e plaqueamento dos protoplastos ... 38
3.5 Regeneração de plantas ... 39
3.5.1 Embriogênese somática ... 39
3.5.2 Germinação de embriões e enraizamento de plântulas ... 39
3.6 Aclimatização ... 40
3.7 Confirmação da hibridação somática... 40
3.7.1 Análise morfológica ... 40
3.7.2 Análise citológica ... 41
3.7.3 Análise molecular ... 41
3.7.3.1 Extração de DNA ... 42
3.7.3.2 Quantificação de DNA ... 42
3.7.3.3 Seleção de ‘primers’ para utilização nas reações de RAPD ... 43
3.7.3.4 Reação de amplificação do DNA ... 43
4 RESULTADOS E DISCUSSÃO ... 45
4.1 Manutenção de calos e culturas em suspensão... 45
4.2 Seleção de plantas matrizes de C. grandis... 46
4.3 Hibridação somática ... 51
4.3.1 Eficiência do protocolo utilizado para a hibridação somática ... 51
4.3.2 Resultados das fusões de protoplastos ... 54
4.3.3 Híbridos somáticos de C. sinensis + C. grandis ... 55
4.4 Confirmação da hibridação somática ... 59
4.4.1 Análise citológica ... 59
4.4.2 Análise morfológica ... 62
4.4.3 Análise molecular por RAPD... 63
5 CONCLUSÕES ... 66
ANEXO ... 67
LISTA DE FIGURAS
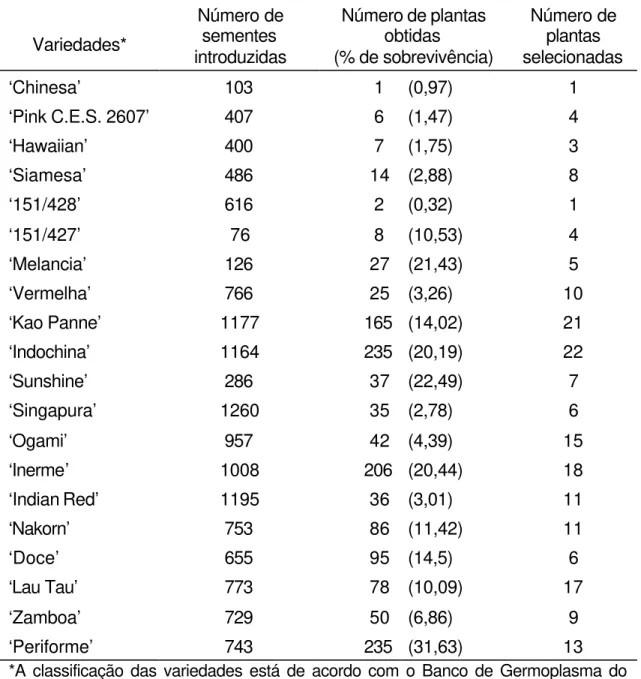
Página 1 Seleção de plantas de toranja tolerantes à Phytophthora sp. ... 50
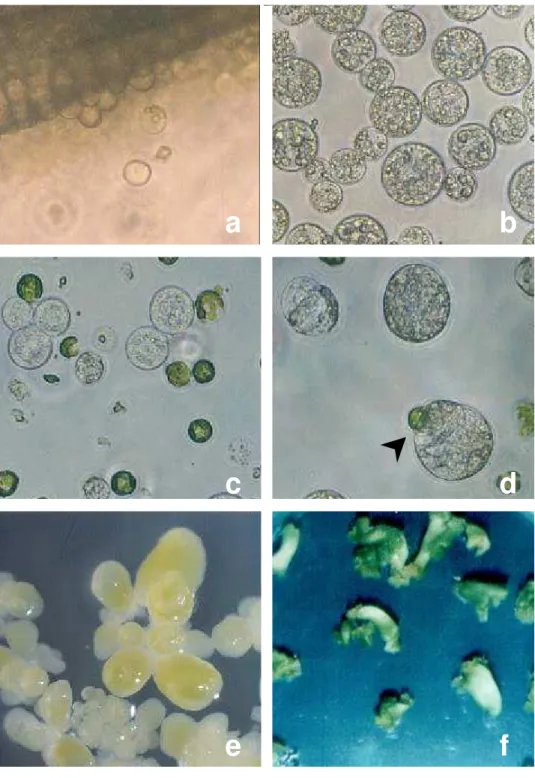
2 Isolamento de protoplastos, fusão e cultura de protoplastos e indução à embriogênese somática ... 52
3 Plantas regeneradas das fusões de protoplastos de C. sinensis com
C. grandis, aclimatizadas em casa-de-vegetação ... 56
4 Análises morfológica, citológica e molecular das plantas regeneradas da fusão de laranja ‘Hamlin’ + toranja enxertada ‘Indian Red’ ... 60
LISTA DE QUADROS
Página 1 Características dos principais porta-enxertos de citros ... 9
2 Híbridos somáticos interespecíficos de Citrus ... 26
3 Híbridos intergenéricos de citros, produzidos por hibridação somática a partir de pais sexualmente compatíveis ...
27
4 Híbridos intergenéricos de citros, produzidos por hibridação somática a partir de pais sexualmente incompatíveis ...
27
5 Cíbridos obtidos a partir de hibridação somática em citros ... 28
LISTA DE TABELAS
Página 1 Variedades de toranja selecionadas para resistência à Phytophthora
sp., utilizadas na hibridação somática como parental
não-embriogênico ... 47
HIBRIDAÇÃO SOMÁTICA ENTRE Citrus sinensis E C. grandis
Autora: MARCIA CRISTINA CALIXTO Orientador: Prof. Dr. FRANCISCO DE ASSIS ALVES MOURÃO FILHO
RESUMO
A hibridação somática de citros tem sido extensivamente aplicada, favorecendo o desenvolvimento de híbridos somáticos em programas de melhoramento genético, como fonte de germoplasma ou como variedades copa e porta-enxerto. Neste contexto, este trabalho foi desenvolvido com o objetivo de selecionar plantas de toranja (Citrus grandis L. Osbeck) tolerantes à Phytophthora
sp. e utilizá-las como parentais no processo de hibridação somática com outras espécies do gênero Citrus, a fim de produzir híbridos somáticos para o
melhoramento de porta-enxertos. Plantas de 20 variedades de toranja tolerantes à
Phytophthora spp. foram selecionadas, após serem cultivadas em solo infestado.
variedades de toranja selecionadas e plantas enxertadas de 12 variedades de toranja, como parentais não-embriogênicos, utilizando-se a técnica de fusão química, via polietilenoglicol (PEG). Microcolônias foram transferidas para meio de cultura MT semi-sólido, suplementado com 500 mg.l-1 de extrato de malte para
SOMATIC HYBRIDIZATION OF Citrus sinensis AND C. grandis
Author: MARCIA CRISTINA CALIXTO Adviser: Prof. Dr. FRANCISCO DE ASSIS ALVES MOURÃO FILHO
SUMMARY
Citrus somatic hybridization has been extensively applied assisting the development of the somatic hybrids which can be used in improvement programs, indirectly as germoplasm source or directly as scion and rootstock varieties. In this context, this research was developed with the objective of selecting plants of pummelo (C. grandis L. Osb) tolerant to Phytophthora sp. and use these plants as
parents in the somatic hybridization process with other species of Citrus. Plants of
20 pummelo varieties, tolerant to Phytophthora sp., were selected after being
1 INTRODUÇÃO
A cultura dos citros tem se destacado no âmbito nacional e internacional. Segundo a FAO (2003), a produção de citros foi estimada em mais de 103 milhões de toneladas métricas em 2002, tendo o Brasil participado com mais de 20 milhões de toneladas métricas. Atualmente, o Brasil é o maior produtor de citros, seguido pelos Estados Unidos, China e México. A citricultura brasileira tem se baseado na laranja, cuja produção ultrapassou 18 milhões de toneladas métricas. O Estado de São Paulo participou, na safra citada, com mais de 2/3 da produção nacional, o que correspondeu a 83% da produção total de laranja, constituindo-se o principal estado produtor e processador desta fruta no país, seguido pelos Estados de Sergipe, Bahia e Minas Gerais (FNP Consultoria & Comércio, 2003).
O Brasil é o maior produtor e exportador de suco cítrico concentrado do mundo, exportando quase 100% do que é produzido, o que corresponde a cerca de 1 milhão de toneladas métricas (FNP Consultoria & Comércio, 2003).
Programas de melhoramento genético, via hibridação sexual, seleção de mutações espontâneas e indução de mutações visando à seleção de novas variedades mais produtivas, resistentes a pragas e doenças, bem como melhor adaptadas às condições abióticas, têm sido conduzidos, porém, com pouco sucesso, devido a aspectos da biologia reprodutiva inerentes à espécie (Vardi, 1981; Grosser & Gmitter Junior, 1990a; Pompeu Júnior, 1991).
Utilizando-se diferentes técnicas biotecnológicas, tem-se conseguido ultrapassar barreiras naturais ao melhoramento dos citros, contribuindo desse modo com programas de melhoramento convencional e auxiliando no desenvolvimento de variedades com características superiores.
A produção de híbridos somáticos de citros e cíbridos, via fusão de protoplastos, tornou-se uma ferramenta importante nos programas de melhoramento de variedades de citros no mundo. Plantas híbridas somáticas de citros têm sido regeneradas de mais de 200 combinações parentais, e muitas combinações de cíbridos também têm sido produzidas (Grosser et al., 2000). Apesar disto, existe a necessidade do desenvolvimento e melhoramento das variedades copa e porta-enxerto brasileiras de interesse comercial, principalmente no que diz respeito a resistência a doenças, produtividade e qualidade dos frutos, para assim, ampliar a base genética dos pomares comerciais.
Visando colaborar com o melhoramento de citros via fusão de protoplastos, o principal objetivo deste trabalho foi a produção de híbridos
somáticos utilizando-se toranjas (C. grandis L. Osbeck)como parental, a fim de
2 REVISÃO DE LITERATURA
2.1 Importância da citricultura no Brasil e no mundo
Os citros são as frutas mais produzidas no mundo, num volume de 24% da produção total, superando as culturas da uva, banana e maçã. Para o ano agrícola 2002/2003, a produção nacional está estimada em mais de 370 milhões de caixas (40,8 Kg), e o Estado de São Paulo foi responsável por cerca de 80% deste total (ABECITRUS, 2003; FNP Consultoria & Comércio, 2003).
Segundo Neves & Boteon (1998), as indústrias de suco concentrado de laranja para exportação comercializam, anualmente, cerca de 85% da produção paulista. No mundo, a cada dez copos de suco de laranja consumidos, oito são de procedência brasileira, e o Estado de São Paulo é responsável por 98% da produção total.
Além da importância econômica, a citricultura paulista também tem uma grande função social, pois as atividades da cadeia produtiva de citros empregam cerca de 400 mil pessoas, direta ou indiretamente, no Brasil (Neves & Boteon, 1998).
concentrado e congelado são as maiores consumidoras dos frutos desta variedade em São Paulo (FNP Consultoria & Comércio, 2003).
A elevada concentração da produção de laranja e de suco concentrado no chamado ‘cinturão paulista”, levou Neves & Neves (1996) a denominá-lo de
commodity sui generis, em face dos localizados impactos alocativos e
distributivos de sua produção, incomuns em outras explorações agrícolas. Cerca de 9% de todo o agronegócio brasileiro vem da laranja, oitavo produto da pauta de exportações do país (FNP Consultoria & Comércio, 2003).
Brasil e Estados Unidos constituem duas das principais áreas de produção citrícola mundial. Os Estados de São Paulo e da Flórida cultivam mais de 1 milhão de hectares em citros, produzindo anualmente cerca de 85% do volume total de suco de laranja natural ou concentrado. Juntas, essas indústrias movimentam algo em torno de 17 bilhões de dólares em todos os elos da cadeia produtiva, consideradas estratégicas para a economia destes países (Neves & Boteon, 1998).
Não existem limitações climáticas para os citros, exceto em algumas áreas do Nordeste onde as chuvas são inferiores a 700 mm por ano, e outras na região Sul, onde podem ocorrer geadas fortes (Amaro et al., 1991). As restrições climáticas constituem-se em sério fator limitante da produção de laranjas para fins industriais, influenciando em várias características da laranja, tais como coloração, acidez, teor de sólidos solúveis, aroma, sabor, espessura da casca e período de maturação. Esses fatores limitam a especificidade da fruta adequada ao processamento para extração de suco (Neves, 1996).
2.2 Origem e distribuição dos citros
O gênero Citrus e outros gêneros relacionados à subfamília Aurantioideae,
família Rutaceae, são nativos da região sudeste do continente asiático, com ramos filogenéticos que se estendem do centro da China ao Japão, e do leste da Índia a Nova Guiné, Austrália e África Tropical (Swingle & Reece, 1967; Soost & Cameron, 1975). O norte da região Indo-Burma também é apontado como centro de origem da maioria das espécies cítricas (Tanaka, 1954). Estudos recentes indicam que a região de Yunnan, no centro sul da China, também pode ter sido importante para a origem e dispersão inicial de um grande número de espécies primitivas (Davies & Albrigo, 1994).
O primeiro fruto cítrico a ser conhecido pelas civilizações européias foi a cidra (C. medica L.), seguida pela laranja azeda (C. aurantium L.), limão (C.
limon Burmann) e laranja doce (Webber et al., 1967). A introdução dos citros no
restante do globo, incluindo as Américas, Sul da África e Austrália, foi realizada pela expansão colonial européia. Todas as espécies de Citrus economicamente
importantes foram originadas no continente asiático, à exceção do pomelo (C.
paradisi Macfadyen), que provavelmente surgiu como um híbrido natural em
Barbados, após a introdução da toranja e da laranja doce pelos colonizadores europeus (Scora et al., 1982).
Atualmente, as plantas cítricas são cultivadas em uma ampla faixa ao redor do globo, compreendida entre os paralelos 35º N e 35º S. Entretanto, as principais áreas produtoras concentram-se em regiões subtropicais, em latitudes superiores a 20º N ou 20º S (Davies & Albrigo, 1994).
Desde a sua introdução no Brasil, feita pelos colonizadores portugueses por volta de 1540, até o final do século 19, as plantas cítricas foram propagadas por sementes (Pompeu Junior, 1991).
Quando a indústria cítrica tornou-se importante no âmbito comercial, no início do século passado, o uso de árvores enxertadas foi iniciado e a laranja ‘Caipira’ (C. sinensis L. Osbeck) era o porta-enxerto mais utilizado. A baixa
resistência a seca e a gomose determinaram o uso da laranja azeda, tendo sido o mais importante porta-enxerto até a década de 40 (Pompeu Junior, 2001). Segundo o autor, nesta época, 12 milhões de árvores representavam a indústria cítrica em São Paulo, da qual 90% estava em porta-enxerto de laranja azeda e outros 10%, em limão ‘Cravo’ (C. limonia), laranja ‘Caipira’, limão Rugoso (C.
jambhiri Lush.) e lima doce da Pérsia (C. limettioides Tan.).
A grande preferência brasileira pelo porta-enxerto laranja azeda se dava devido às condições favoráveis ao desenvolvimento desse cavalo, boa afinidade com a maioria das variedades comerciais, além da qualidade das frutas produzidas sobre esse porta-enxerto (Moreira, 1941).
A introdução do vírus da tristeza dos citros (CTV) em São Paulo, em 1937, sua rápida disseminação pelo pulgão preto (Toxoptera citricidus Kirk.) causou a
morte de árvores com porta-enxertos de laranja azeda e lima da Pérsia, que não são tolerantes a este vírus. Experimentos conduzidos pelo IAC em colaboração com o Departamento de Agricultura dos EUA (USDA) com cerca de 400 citros e gêneros relacionados para selecionar porta-enxertos resistentes foram feitos. Esta pesquisa permitiu a renovação da citricultura brasileira com os porta-enxertos limão ‘Cravo’, tangerina ‘Cleópatra’ (C. reshni Hort. ex Tanaka), limão
‘Rugoso’ e citrange ‘Troyer’ (Poncirus trifoliata L. Rafinesque x C. sinensis L.
Osbeck) (Chapot, 1975; Pompeu Junior, 1991; 2001).
‘Cravo’ às doenças exocorte e xiloporose (Pompeu Junior, 2001), cuja presença nos pomares eram mascaradas pela tolerância da laranja azeda, se tornou um sério problema, desde que estas doenças podiam reduzir em até 70% a produção de frutos. As pesquisas levaram, então, ao uso de clones nucelares a partir de 1955 (Pompeu Junior, 2001). Segundo este autor, de 1961 à 1970, a utilização do limão ‘Cravo’ aumentou de 77 para 99%.
A partir da década de 70, uma doença de etiologia desconhecida, o declínio dos citros, se tornou um problema tanto na Flórida quanto no Brasil, sendo os porta-enxertos mais suscetíveis P. trifoliata, limão ‘Rugoso’ e limão
‘Cravo’ (Grosser & Gmitter Junior, 1990a). O declínio vem causando a morte de milhões de plantas enxertadas sobre estes cavalos, com perdas estimadas em cerca de 200 milhões de dólares anuais (FUNDECITRUS, 2003).
Segundo Pompeu Junior (2001), a intolerância do limão ‘Cravo’ ao declínio causou uma pequena diversificação de porta-enxertos usados no Brasil. Tangerina ‘Cleópatra’, limão ‘Volkameriano’ (C. volkameriana Tan. e Pasq.),
tangerina ‘Sunki’ (C. sunki Hort.) e, no início dos anos 90, citrumelo Swingle (C.
paradisi Mac. x Poncirus trifoliata L. Raf.) se mostraram boas alternativas para a
diversificação no uso de porta-enxertos.
A história da citricultura vem mostrando a importância desta diversificação na utilização de porta-enxertos, além da necessidade de variedades porta-enxerto melhoradas. A utilização generalizada de um único porta-enxerto para todas as variedades copa provavelmente não atende às características peculiares de cada cultivar, impedindo que a planta, mesmo recebendo os tratos culturais adequados, manifeste todo o seu potencial produtivo (Pompeu Junior et al., 1986).
O conhecimento das características dos porta-enxertos e a racional utilização delas permite maior produtividade, quer pelo aumento da produção, quer pelo adiantamento ou atraso na maturação dos frutos (Pompeu Junior, 1991). Segundo o autor, o porta-enxerto induz à variedade copa alterações no seu crescimento, tamanho, precocidade de produção, produtividade, época de maturação e massa dos frutos, permanência dos frutos na planta, conservação da fruta após a colheita, transpiração das folhas, fertilidade do pólen, composição química das folhas, capacidade de absorção, síntese e utilização de nutrientes, tolerância a salinidade, resistência a seca e ao frio, resistência e tolerância à doenças e pragas, e resposta a produtos de abscisão.
A escolha do porta-enxerto a ser utilizado é uma fase de grande importância quando no planejamento de um pomar. Essa escolha deverá levar em conta, também, o local de instalação do pomar, o clima, o solo, a copa e o manejo a ser adotado, já que não existe um porta-enxerto ideal. Em função dessas características, será escolhido o porta-enxerto que melhor se adapta à situação. O Quadro 1 apresenta características dos principais porta-enxertos utilizados no Brasil.
viveiro Resistência à gomose à tristeza à seca média grande grande média grande grande baixa grande baixa média grande média média grande média média grande média alta grande média alta pequena grande alta grande média alta média média alta média média Início de produção
precoce precoce médio médio médio médio precoce médio médio médio médio
Produção boa boa boa boa boa boa boa boa boa boa boa
Qualidade dos frutos
média média boa boa boa boa ótima boa boa boa boa
Tamanho das plantas
médio médio grande grande grande grande pequeno grande grande grande grande
Solo indicado arenoso ou argiloso arenoso ou argiloso arenoso ou argiloso
argiloso argiloso arenoso ou argiloso
úmido arenoso ou argiloso
arenoso ou argiloso
argiloso argiloso
Nematóide4, 5
suscetível suscetível suscetível suscetível suscetível suscetível resistente suscetível resistente resistente resistente
Indicado para citros em geral citros em geral3 citros em geral citros em geral citros em geral citros em geral citros em geral2
limões citros em geral2
citros em geral2
citros em geral2
Declínio suscetível suscetível tolerante tolerante tolerante tolerante suscetível tolerante tolerante suscetível suscetível
1 As características representam informações coletadas em experimentos e pomares comerciais; 2 Exceto Laranja Pera e Tangor Murcote; 3
Exceto Laranja Pera; 4
Tylenchulus semipenetrans; 5 Fonte: Willits & New Comb. Inc. California - EUA. Adaptado de Pompeu Junior (1991) e Citrograf Mudas (2003).
2.3.1 Importância da gomose de Phytophthora na cultura dos citros
Além das influências sobre as características da copa, um bom porta-enxerto deve apresentar características de resistência ou tolerância a doenças, especialmente aquelas que afetam diretamente o porta-enxerto, como as doenças de solo.
De acordo com Feichtenberger (2001), muitas doenças importantes prejudicam a indústria citrícola brasileira. Duas doenças bacterianas são muito importantes: o cancro cítrico (Xanthomonas axonopodis Vanterin et al.) e a
clorose variegada dos citros - CVC (Xylella fastidiosa Wells et al.). As maiores
doenças viróticas são leprose e tristeza, que se disseminam facilmente, causando grandes perdas. Declínio ou ‘Citrus Blight’, de etiologia desconhecida, também é facilmente disseminado e muito importante. As mais graves doenças causadas por fungos são mancha preta dos citros (Guignardia citricarpa Kiely), verrugose
(Elsinoe australis Bitancourt & Jenk, e Elsinoe fawcetti Bitancourt & Jenk),
melanose (Diaporthe citri Wolf), rubelose [Erythricium salmonicolor (Berk. &
Broome) Burdsall (sin. Corticium salmonicolor Berk. & Broome)], e podridão floral
ou queda prematura de frutos (Colletotrichum acutatum Simmons). As doenças
induzidas por Phytophthora [Phytophthora parasitica Dastur (sin. P. nicotianae
Breda de Haan), P. citrophthora (R. E. Sm. & E. H. Sm.) Leonian e P. palmivora
(E.J.Butler) E. J. Butler], principalmente podridão do colo e da raiz, são as mais sérias doenças de solo de citros (Feichtenberger, 2000). Dois nematóides são importantes: o nematóide dos citros (Tylenchulus semipenetrans Cobb) e o
nematóide de lesão (Pratylenchus jaehni sp n. (sin. P. coffeae (Zimm.) Filip. &
Schuur. Stek).
doenças, principalmente no caso do cancro cítrico, CVC, e das doenças induzidas por Phytophthora, nematóides, exocorte e linhagens severas do vírus
da tristeza - CTV (Feichtenberger, 2000; 2001).
A gomose causada por Phytophthora sp. geralmente se manifesta no colo
da planta, provocando podridão e exsudação de goma, podendo expandir-se para as raízes principais até 20 ou 30 cm abaixo do solo e para cima do tronco. Portanto, quando toda a periferia do tronco é atingida a planta morre por estrangulamento devido ao ataque do câmbio ou floema, o que interrompe o fluxo descendente de seiva. Na sementeira, estes fungos causam o “tombamento” ou
damping-off, atacando as plântulas e afetando os tecidos da região do colo, onde
surgem lesões deprimidas e de coloração escura, que crescem e provocam a morte das plantas (Fundecitrus, 2003; Prates & Pelegrinetti, 1995).
Entre os porta-enxertos, Poncirus trifoliata, citrumelo ‘Swingle’, citranges e
a laranja azeda são altamente resistentes à Phytophthora. As tangerinas, tangelos
e limões são medianamente suscetíveis ou tolerantes, enquanto as laranjas doces, incluindo a laranja ‘Caipira’, são altamente suscetíveis à doença (Amorim, 1997; Davies & Albrigo, 1994).
Diversas espécies de Phytophthora já foram registradas causando
podridões de colo e de raízes de plantas cítricas no Brasil. P. parasitica está
largamente disseminada nas regiões produtoras de citros e é a principal espécie responsável por elevadas perdas na cultura. P. citrophthora ocorre principalmente
na região sul do país, onde as temperaturas geralmente são inferiores às das outras regiões (Feichtenberger, 1996; Feichtenberger et al., 1997). P. palmivora
ocorre em muitos poucos casos em plantios comerciais tendo sido detectada, nos últimos anos, também em viveiros (Feichtenberger, 2001).
Nos Estados Unidos, as doenças causadas por Phytophthora vêm
espécies deste fungo podem se tornar epidêmicas com uma produção exponencial de inóculo secundário.
A incidência das doenças induzidas por Phytophthora sp. é muito alta em
plantios novos devido, principalmente, ao uso de plantas produzidas em viveiros contaminados, os quais são ainda bastante freqüentes (Feichtenberger, 2001). Segundo o autor, um levantamento realizado pelo Instituto Biológico e pelo Fundecitrus, no Estado de São Paulo, Phytophthora foi detectada em 26% e
26,1% das amostras de solo e de substrato sem solo coletadas em abril/maio de 2000 dos 300 campos e 94 recipientes de mudas amostrados, respectivamente. No levantamento realizado em março/abril de 2001, realizado apenas nos viveiros selecionados, Phytophthora foi detectada em 14,7% das amostras coletadas dos
101 viveiros amostrados. As práticas inapropriadas utilizadas em muitos viveiros e o uso de mudas contaminadas com Phytophthora são as maiores causas da
alta incidência nos viveiros amostrados em São Paulo. Neste mesmo levantamento, o patógeno foi detectado em 18,7% das 48 sementeiras amostradas nos viveiros.
O uso de plantas livres de Phytophthora spp. durante o estabelecimento de
novos plantios ou durante o replantio é a medida mais importante no manejo da gomose em árvores jovens e adultas (Feichtenberger, 1996; 2000). A obtenção e uso de porta-enxertos resistentes pode ser, portanto, encarada como mais uma importante estratégia de manejo das doenças causadas por Phytophthora.
2.4 Melhoramento de porta-enxertos de citros
qualidade do suco. Entre as características patológicas, evidencia-se a tolerância ou a suscetibilidade dos diferentes porta-enxertos a doenças como gomose (Phytophthora sp.), tristeza, declínio e aos nematóides (Mourão Filho, 1996).
Também apresentam diferentes níveis de tolerância a fatores abióticos, tais como0 baixa temperatura, salinidade, seca e encharcamento.
Os primeiros programas de melhoramento de citros foram iniciados na Flórida, em 1893, com Swingle e Webber. Desde então, numerosos programas foram desenvolvidos com diversos objetivos (Davies & Albrigo, 1994).
O melhoramento genético tradicional tem apresentado limitações à obtenção de novas variedades porta-enxerto e copa, devido à descontinuidade nos programas estabelecidos e a fatores biológicos característicos da espécie como, por exemplo, longos ciclos de reprodução, juvenilidade, poliembrionia nucelar (apomixia), alta heterozigose, incompatibilidade sexual, esterilidade gametofítica e poliploidia (Cristofani, 1997; Grosser & Gmitter Junior., 1990a; Ling et al., 1989; Vardi, 1981; Vardi & Galun, 1989; Vardi & Spiegel-Roy, 1982; Vardi et al., 1974). Como conseqüência, as variedades desenvolvidas para a citricultura mundial, neste último século, têm sido originadas a partir de seleções naturais e mutações (Soost & Cameron, 1975). Cerca de 24% das variedades de laranjas doces são comprovadamente originadas de mutações de gemas, enquanto 23% o foram a partir de mutações em plântulas. O restante tem origem desconhecida (Hodgson, 1967). Como exemplo, temos a laranja ‘Baianinha’, que provavelmente é uma variação da laranja ‘Baia’ (Figueiredo, 1991) que, por sua vez, provém de mutação da laranja ‘Seleta’ (Moreira & Pio, 1991).
A apomixia ou embrionia nucelar representa um dos maiores obstáculos para programas de melhoramento via hibridação sexual em citros. Os embriões nucelares competem com o embrião zigótico por espaço e nutrientes nas sementes em desenvolvimento resultando, freqüentemente, na perda do embrião zigótico (Cristofani, 1997). Devido à poliembrionia, a produção de uma população de híbridos segregantes de tamanho suficiente para a seleção se torna difícil. Várias técnicas têm sido utilizadas para identificar plântulas zigóticas e plântulas nucelares como, por exemplo, isoenzimas ou marcadores do tipo RAPD.
A poliploidia é bastante comum em citros e é um dos fatores que interferem na já complexa transmissão de caracteres. O número básico de nove cromossomos aparece em algumas variedades, não somente multiplicado por dois, mas também em maior número, como ocorre com as formas tetraplóides e triplóides (Cristofani, 1997). Segundo Moreira & Pio (1991), as plantas tetraplóides (2n=4x=36 cromossomos) apresentam as folhas maiores na largura, mais espessas e a coloração tende a ser mais escura. O crescimento é mais lento nos tetraplóides e as brotações vigorosas são menos comuns. A planta é menor, menos ereta e mais compacta, demora mais a florescer e geralmente frutifica menos.
A partenocarpia, desenvolvimento de frutos sem fertilização levando à formação de frutos sem sementes, ocorre em citros devido à esterilidade gamética ou incompatibilidade. A esterilidade gametofítica e a auto-fecundação têm sido, juntamente com a triploidia, as principais causas do aparecimento de frutos sem sementes (Grosser & Gmitter Junior, 1990a; Vardi & Galun, 1989).
resistência ao vírus da tristeza dos citros, resistência aos nematóides dos citros, embrionia nucelar e o caráter folha trilobada de P. trifoliata (Cristofani, 1997).
Desse modo, cruzamentos entre parentais com características complementares, freqüentemente não produzem a progênie híbrida desejada. Progênies fracas são produzidas como resultado da depressão por endogamia (Barret & Rhodes, 1976; Swingle & Reece, 1967), onde genes deletérios antes mascarados por genes dominantes são expressos devido à segregação meiótica (Grosser & Gmitter Junior, 1990a). Devido a esse fato, dentre os muitos híbridos obtidos por fecundação controlada pelo homem, vários eram pouco vigorosos e por isso poucos apresentam valor comercial (Moreira & Pio, 1991). A autofecundação de híbridos F1 com maior heterozigosidade pode ser mais
vantajosa no melhoramento, mas não tem sido adequadamente investigada. Por outro lado, indivíduos mostrando características interessantes são geralmente usados como progenitores, mas não existe nenhuma garantia que tais características estarão presentes regularmente nos híbridos (Cameron & Frost, 1968; Grosser & Gmitter Junior, 1990a). A produção contínua de descendentes zigóticos fracos e não competitivos devido à depressão por endogamia pode ter selecionado para altos níveis de embrionia nucelar (Davies & Albrigo, 1994).
A maioria das espécies de citros tem longo período juvenil, levando cinco ou mais anos até a primeira floração (Grosser & Gmitter Junior, 1990a) e de 10 a 15 anos para que possam ser bem avaliadas as características de produção e qualidade dos frutos. Aliado a este fato, a juvenilidade dos citros está associada à presença de espinhos, árvores com hábito de crescimento vertical e grande vigor, resultando em árvores de grande tamanho. Isso torna o melhoramento de citros atividade de longo prazo, caro, além de exigir grande extensão de terra (Davies & Albrigo, 1994; Koller, 1994).
podem ser utilizados em programas de melhoramento (Grosser & Gmitter Junior, 1990a). Como exemplos de híbridos produzidos através de hibridação natural, citam-se o tangor ‘Murcote’ (C. reticulata Blanco x C. sinensis L. Osbeck),
citrange ‘Carrizo’ e ‘Troyer’ (Poncirus trifoliata Rafinesque x C. sinensis L.
Osbeck), tangelo ‘Orlando’ e ‘Nova’ (C. reticulata Blanco x C. paradisi
Macfadyen), entre outros.
De acordo com Moreira & Pio (1991), a variação genética se origina de processos distintos. A maior parte da variação genética decorre de recombinação gênica, sendo que os métodos de melhoramento por cruzamento exploram estas recombinações. Ocasionalmente, podem ocorrer mutações de ponto, aberrações cromossômicas estruturais e numéricas, alterações estas que podem ser decorrentes de mutações induzidas ou espontâneas.
A seleção clonal baseia-se na ocorrência de mutações somáticas espontâneas em gemas ou ramos, que podem dar origem a plantas que mantêm as características originais da variedade, mas oferecem novas características relacionadas à produção, qualidade de fruto ou resistência a doenças. A seleção de clones de variedades comerciais, dentro de pomares, tem sido praticada em várias regiões, possibilitando a liberação de novas variedades a exemplo de tangerinas, laranjas e pomelos (Grosser & Gmitter Junior, 1990a; Grosser et al., 1992b). Como exemplos da seleção clonal de mutantes naturais, os mais importantes foram aqueles que deram origem à variedades comerciais como ‘Natal’, ‘Bahia’, ‘Baianinha’, ‘Westin’, ‘Piralima’, ‘Lima Verde’ e ‘Folha Murcha’, dentre outras (Teófilo Sobrinho et al., 1978; Domingues et al., 1995; Donadio et al., 1995).
Em geral, os citros e outros gêneros da subfamília Aurantioideae são diplóides, com 2n=2x=18 cromossomos (Cameron & Frost, 1968; Koller, 1994; Moreira & Pio, 1991; Spiegel-Roy & Vardi, 1984). Triplóides e tetraplóides podem ser utilizados como porta-enxertos com o objetivo de diminuir o porte da copa (Grosser & Gmitter Junior, 1990a).
Dentre os vários programas de melhoramento, a seleção de clones nucelares foi o que resultou em maiores ganhos econômicos para a citricultura (Salibe et al., 1984). A produção destes foi iniciada na Estação Experimental de Limeira, em torno de 1938 (Moreira, 1962), e foram propagados a partir de 1955 (Salibe et al., 1984; Rodriguez, 1956; Moreira, 1962). Até o momento é material de grande uso comercial no Brasil (Passos et al., 1973; Figueiredo, 1991).
Pode-se mencionar ainda, a obtenção de clones pré-imunizados de laranja ‘Pêra’, viabilizando, portanto, o uso comercial da mais importante variedade cítrica do país, pela relação econômica que resultou (Muller & Costa, 1991; Muller et al., 1999). Após os clones nucelares, é possível que este tenha sido um dos grandes avanço da citricultura brasileira.
2.5 Biotecnologia para o melhoramento genético de citros
A incorporação de técnicas de biotecnologia, a exemplo da cultura de tecidos, genética molecular, fusão de protoplastos, transformação genética, entre outras, em programas de melhoramento de citros, permite tanto facilitar e acelerar a utilização da variabilidade disponível (Grosser & Gmitter Junior, 1990a) como utilizar germoplasma ainda não explorado (Grosser et al., 1996b), criando novas combinações que poderão ser incluídas em programas de melhoramento convencional, ou resultar em variedades totalmente novas (Gmitter Junior et al., 1992; Ollitraut & Luro, 1995).
enfoques foi a obtenção de embriões nucelares in vitro a partir de espécies
monoembriônicas, com a finalidade de produzir plantas isentas de viroses. Posteriormente, intensificou-se a obtenção de calos nucelares embriogênicos, devido à possibilidade de se obter embriões somáticos em grande quantidade a partir destes calos (Cristofani, 1991).
A cultura de tecidos em citros foi iniciada por volta de 1940, quando calos de cidra foram induzidos in vitro a partir de brotos (Demétriadès, 1954, citado por
Mourão Filho, 1995). Ranga Swamy (1958; 1959) aproveitou a capacidade natural de proliferação das células de tecidos nucelares, e os estabeleceu in vitro, com a
finalidade de obter plântulas livres de vírus. Posteriormente, foram obtidos embrióides in vitro de diversas variedades poliembriônicas (Kochba et al., 1972;
Mitra & Chaturvedi, 1972) e monoembriônicas (Rangan et al., 1968; 1969; Bitters et al., 1970; Deidda, 1973; Esan, 1973; Juarez et al., 1976; Navarro & Juarez, 1977; Button & Kochba, 1977).
A regeneração de plantas, por organogênese ou embriogênese somática, a partir do cultivo de células e tecidos vegetais in vitro, é a base para a utilização
de técnicas de biotecnologia no melhoramento genético (Gmitter Junior et al., 1992).
Mais recentemente, com o desenvolvimento da biologia molecular e celular, novas técnicas têm sido aplicadas com o objetivo de sobrepor as barreiras enfrentadas pelo melhoramento tradicional, como a hibridação somática e a transformação genética. Em citros, protocolos de cultura de tecidos encontram-se descritos para um grande número de espécies e gêneros relacionados, permitindo-se hoje que a técnica de hibridação somática seja parte integrante de programas de melhoramento em vários países (Grosser & Gmitter Junior, 1990a).
interespecíficos, intergenéricos, de pais compatíveis ou não, ou ainda, cíbridos, cuja combinação genômica ocorre a nível do citoplasma.
A transformação genética, por sua vez, consiste na introdução de genes derivados de plantas relacionadas ou não, e mesmo de outros reinos (bactérias, fungos, animais) no genoma de plantas permitindo, assim, melhorar o valor agronômico de espécies cultivadas, que possam atuar como biorreatores vivos para a produção de proteínas e metabólitos de importância econômica ou, ainda, como meio para se estudar a ação de genes durante o desenvolvimento e outros processos biológicos (De Block, 1993; Farias, 2001).
Outra técnica que pode ser aplicada ao melhoramento de citros é a cultura de endosperma. O endosperma é um tecido triplóide que apresenta 2n 3x=27 cromossomos e pode ser cultivado in vitro, produzindo calos e formando
embriões (Louzada, 1990; Gmitter Junior et al., 1990).
2.6 Hibridação somática em citros
A hibridação somática de protoplastos de plantas é uma técnica através da qual as propriedades celulares podem ser modificadas mediante fusões nucleares e/ou citoplasmáticas (Carneiro et al., 1998).
O sistema de produção de híbridos somáticos pela fusão de protoplastos de citros é baseado na capacidade de regeneração de plantas via embriogênese somática a partir de calos (Motomura et al., 1995). O alto potencial de regeneração de plantas a partir de células embriogênicas de citros as torna material adequado para fusão de protoplastos, transformação genética e armazenamento in vitro de germoplasma (Kunitake & Mii, 1995).
Ohgawara & Kobayashi, 1991). Este é um processo que combina os genomas nucleares completos de ambos os parentais podendo, entretanto, nas combinações entre espécies mais distantes, ocorrer a eliminação cromossômica. A heterozigose potencial dos híbridos somáticos é grande, dependendo das diferenças dos alelos dos parentais. Em contraste à hibridação sexual, em que a herança citoplasmática é predominantemente maternal, os híbridos somáticos permitem a contribuição de ambos os parentais (Grosser & Gmitter Junior, 1990b). Esta combinação pode gerar uma variabilidade genética adicional, que pode surgir a partir da combinação de genes citoplasmáticos e nucleares (Kumar & Cocking, 1987).
Os híbridos somáticos apresentam a grande vantagem de serem alotetraplóides, mantendo os genes dos parentais por não haver segregação meiótica. Portanto, os genes deletérios recessivos existentes nos parentais que não se expressam, por uma condição heterozigota, provavelmente, permanecerão não expressos nos híbridos somáticos, e características controladas por genes dominantes ou codominantes presentes em um dos parentais podem se expressar nos híbridos somáticos (Grosser & Gmitter Junior, 1990a). Uma outra vantagem da hibridação somática, segundo estes autores, seria a possibilidade do uso direto dos híbridos obtidos como porta-enxertos, sem a necessidade de realizar retrocruzamentos.
Esta técnica de melhoramento genético, entretanto, também apresenta algumas limitações. De acordo com Grosser & Gmitter Junior (1990a), uma das duas espécies parentais tem de possuir capacidade morfogênica in vitro para ser
Em programas de melhoramento genético de citros, a hibridação somática tem sido extensivamente aplicada (Grosser & Gmitter Junior, 1990a; 1990b; Grosser et al., 1990), favorecendo o desenvolvimento de híbridos interespecíficos (Tusa et al., 1990); intergenéricos de parentais sexualmente compatíveis (Grosser et al., 1988a; Deng et al., 1992), intergenéricos de parentais sexualmente incompatíveis (Grosser et al., 1988b; Ling & Iwamasa, 1994) e cíbridos (Saito et al., 1993; Grosser et al., 1996a).
Segundo Grosser (1993), duas estratégias podem ser utilizadas para a aplicação da tecnologia de hibridação somática nos programas de melhoramento de porta-enxerto. A primeira visa produzir híbridos somáticos que combinem genótipos complementares de variedades de porta-enxerto. Uma segunda estratégia é a hibridação de espécies sexualmente incompatíveis de gêneros relacionados que possuam características de interesse agronômico.
Quanto ao melhoramento de variedades copa, os produtos da hibridação somática não possuem potencial para serem utilizados diretamente, já que frutos tetraplóides possuem características não aceitáveis pelo mercado consumidor, como casca grossa e forma irregular (Grosser et al., 1992b), mas podem ser incorporados ao melhoramento como parental tetraplóide (2n=4x=36 cromossomos), em cruzamento com variedades diplóides monoembriônicas (2n=2x=18 cromossomos), gerando progênie triplóide (2n=3x=27 cromossomos) (Ohgawara et al., 1991; Grosser et al., 1992a; Mourão Filho et al., 1996; Grosser et al., 2000a), potencialmente sem sementes, devido à ausência de complementos cromossômicos não balanceados durante a segregação meiótica, como é o caso do cruzamento de tangelo ‘Nova’ com o híbrido somático tangelo ‘Nova’ + laranja ‘Succari’ (Grosser et al., 1998b). A hibridação somática também permite combinar variedades as quais o melhoramento convencional é inibido por esterilidade gametofítica e baixa produção de sementes, como laranja ‘Washington Navel’ e tangerina ‘Satsuma’ (C. unshiu Marc.) (Kobayashi et al.,
Os métodos mais usados para induzir as fusões de protoplastos incluem: aplicação de corrente elétrica (eletrofusão) (Zimmerman, 1982), e o tratamento com polietilenoglicol (PEG) (Kao & Michayluk, 1974; Negrutiu et al., 1986).
Estas duas técnicas têm como princípio favorecer a agregação de protoplastos, que normalmente se repelem por causa das cargas negativas da membrana plasmática, e induzir a instabilidade das membranas (Bengochea & Dodds, 1986). No entanto, o método mais difundido de produção de híbridos somáticos de citros, de acordo com Grosser (1994) e Grosser et al. (1996a; 1996b), é o tratamento com polietilenoglicol (PEG), que induz a fusão química de protoplastos isolados de calos embriogênicos friáveis de origem nucelar ou culturas de células em suspensão como um parental não embriogênico, com protoplastos isolados a partir de folhas de plântulas. Este método permite a fácil determinação da eficiência de fusão, pois os heterocários podem ser facilmente identificados pela presença de marcadores distintos de ambas fontes parentais (grãos de amido derivados de protoplastos de cultura embriogênica e cloroplastos derivados de protoplastos de folhas). Além disso, é eficiente, barato e não parece influenciar na viabilidade dos protoplastos (Grosser & Gmitter Junior, 1990a). Entretanto, Saito et al. (1991) comentam que este método consome muito tempo, é complicado e tóxico para os protoplastos e, que as técnicas de eletrofusão são muito mais simples e eficientes.
Saito et al., 1991; Zimmerman, 1982). Entretanto, estes autores comentam que esta técnica requer o uso de aparelhos de alto custo, o que limita a sua aplicação em larga escala.
Após a fusão, os protoplastos são cultivados em meio nutritivo, para estimular a regeneração da parede celular e divisão das células (Carneiro et al., 1998). Em cultura, as células se dividem formando colônias e, por meio de variações na composição do meio, densidade celular, freqüência de subcultura, condições adequadas de temperatura e luminosidade, pode-se regenerar plantas por embriogênese somática (Grambow et al., 1972). Entretanto, em alguns casos, protoplastos podem regenerar embriões somáticos diretamente sem passar pela fase de calo (Carneiro et al., 1998).
Calos nucelares de citros obtidos a partir de óvulos são talvez a única e a mais apropriada fonte de protoplastos embriogênicos (Vardi & Galun, 1988). Entretanto, Hidaka & Kajiura (1988) introduziram in vitro embriões de sementes
jovens de espécies de citros e obtiveram calos friáveis embriogênicos da região do hipocótilo do embrião. Protoplastos isolados desses calos deram origem a plantas. Ling et al. (1989) introduziram in vitro anteras de Calamondin (C.
madurensis Loureiro), das quais emergiram embriões. Alguns embriões
produziram calos embriogênicos na região do hipocótilo, a partir dos quais foram isolados protoplastos, que deram origem a plântulas.
A utilização de calos tem sido relacionada com altas taxas de variação somaclonal em diversas culturas. Entretanto, embriões de citros obtidos a partir de calos conservam a identidade genética da planta mãe e expressam pouca variabilidade (Jiménez, 1996).
O primeiro exemplo de hibridação somática em citros foi descrito por Ohgawara et al. (1985). O híbrido intergenérico foi obtido pela fusão de protoplastos induzida por PEG, isolados de calos embriogênicos de laranja ‘Trovita’ com protoplastos isolados de folhas de P. trifoliata. Posteriormente, o
‘Washington Navel’ + tangor ‘Murcote’ (Kobayashi et al., 1988a) e laranja ‘Washington Navel’ + tangerina ‘Satsuma’ (Kobayashi et al., 1988b), com o objetivo de superar barreiras impostas pela hibridação sexual. Grosser et al. (1989) relataram novo híbrido intergenérico a partir da cultura de protoplastos isolados de lima ‘Key’ (Citrus aurantifolia Swingle) e laranja ‘Valência’, o qual
deve combinar as características de tolerância ao frio e à tristeza, da laranja ‘Valência’, com o curto período de juvenilidade da lima ‘Key’.
A partir daí, muitos híbridos somáticos de citros já foram produzidos, como exemplificado resumidamente nos Quadros 2 a 6. Revisão bem mais completa também pode ser consultada em Grosser et al. (2000a). Vários grupos de pesquisa têm se dedicado à hibridação somática, em países como França, Japão, Israel, Espanha, além dos Estados Unidos e Brasil, onde a citricultura tem se destacado na economia local. A Universidade da Flórida, nos Estados Unidos, e o Brasil, através de pesquisas desenvolvidas nos laboratórios de Biotecnologia Vegetal do USP/CENA e de Biotecnologia de Plantas Hortícolas, na USP/ESALQ, no Estado de São Paulo, vêm realizando pesquisas com hibridação somática, com o intuito de contribuir para o melhoramento genético das espécies cítricas (Vardi et al., 1987; Grosser et al., 1988a; 1988b; 2000a; Mendes-da-Glória et al., 2000; Mendes et al., 2001). Os híbridos produzidos no Brasil estão descritos no Quadro 7.
Segundo Grosser et al. (1992b), muitos resultados já foram obtidos com relação a produção de híbridos somáticos em citros, pela fusão de protoplastos. Como exemplos, pode-se citar híbridos interespecíficos de Citrus, com parentais
sexualmente compatíveis, como laranja ‘Hamlin’ + tangerina ‘Ponkan’, laranja ‘Valência’ + tangor ‘Murcote’ (Grosser et al., 1998b); híbridos somáticos intergenéricos de Citrus, obtidos a partir de pais sexualmente compatíveis, como
laranja ‘Hamlin’ + Microcitrus papuana, laranja ‘Succari’ + kunquat ‘Meiwa’
sexualmente incompatíveis, como tangelo ‘Nova’ + C. ichangensis (Grosser et al.,
1996b), kunquat ‘Mame’ + Poncirus trifoliata (Miranda et al., 1997), tangelo ‘Page’
+ Murraya paniculata L. Jack (Guo & Deng, 1998).
Durante o processo de fusão de protoplastos, podem ocorrer vários resultados possíveis. Se ocorre a fusão dos dois núcleos, tem-se um híbrido verdadeiro. Quando ocorre perda cromossômica de um ou de ambos os núcleos, chama-se híbrido assimétrico. Se todo o núcleo de um dos parentais é perdido, resta apenas um núcleo na mistura dos dois citoplasmas, este produto é chamado de cíbrido (Bengochea & Dodds, 1986).
Segundo Grosser et al. (1996a), a grande maioria dos cíbridos descritos na literatura são provenientes de falhas na fusão nuclear e perda de material genético. Saito et al. (1991;1993) descrevem que as plantas cíbridas obtidas mostraram a morfologia do parental cujos protoplastos foram isolados de mesófilo foliar. Além disto, estas plantas possuíam o genoma nuclear das células do mesófilo combinado com o genoma mitocondrial das células dos calos nucelares, o que mostrou que células cíbridas com o genoma nuclear apenas do parental não-embriogênico também são capazes de regenerar plantas por embriogênese.
Híbridos somáticos potenciais são identificados com base na morfologia, e são comprovados pela determinação do número de cromossomos, além de análises isoenzimáticas e RAPD (DNA polimórfico amplificado ao acaso) (Grosser & Gmitter Junior, 1990a). Nestas avaliações, é esperado que híbridos somáticos alotetraplóides de citros tenham 36 cromossomos, sendo 18 cromossomos de cada um dos parentais diplóides. RAPD e isoenzimas são utilizados para mostrar a expressão de genes ou a presença de DNA, respectivamente, de cada parental no híbrido somático correspondente.
e selecionar híbridos superiores sob o ponto de vista genético e horticultural (Grosser & Gmitter Junior, 1990b).
Parentais Referências
laranja ‘Washington Navel’+ tangor ‘Murcote’ Kobayashi et al. (1988a) laranja ‘Washington Navel’+ tangerina ‘Satsuma’ Kobayashi et al. (1988b) lima ácida ‘Key’ + laranja ‘Valência’ Grosser et al. (1989) laranja ‘Bahia’+ pomelo ‘Marsh seedless’ Ohgawara et al. (1989) laranja ‘Valência’+ limão ‘Femminello’ Tusa et al. (1990) laranja ‘Washington Navel’ + tangor ‘Murcote’ Kobayashi et al. (1991)
C. sudachi + lima ácida Saito et al. (1991)
laranja ‘Hamlin’+ limão ‘Rugoso’
pomelo ‘Thompson’ + tangor ‘Murcote’ Grosser et al. (1992b) laranja azeda + limão ‘Volkameriano’
laranja ‘Valência’ + citrange ‘Carrizo’ Louzada et al. (1992) limão Milam híbrido + limão ‘Femminello’ Tusa et al. (1992) laranja azeda + limão ‘Cravo’ Grosser et al. (1994) tangerina ‘Ponkan’ + limão ‘Rugoso’ Moriguchi et al. (1996) laranja ‘Succari’ + tangerina ‘Dancy’
laranja ‘Succari’ + tangor ‘Murcote’ laranja ‘Succari’ + tangerina ‘Ponkan’
Mourão Filho et al. (1996)
laranja ‘Valência’ + tangelo ‘Minneola’ laranja ‘Valência’ + tangor ‘Murcote’ laranja ‘Valência’ + tangelo ‘Page’
laranja ‘Valência’ Rhode Red + tangerina ‘Dancy’ laranja ‘Succari’ + toranja ‘Hirado Buntan’
Grosser et al. (1998a)
tangelo ‘Nova’ + toranja ‘Hirado Buntan’
laranja ‘Hamlin’ + tangerina ‘Ponkan’ Grosser et al. (1998a) limão ‘Milam’ híbrido + citrumelo ‘Swingle’
tangerina ‘Cleópatra’ + citrange ‘Carrizo’ tangelo ‘Nova’ + toranja ‘Hirado Buntan’
Grosser et al. (1998b)
tangerina ‘Hongju’ + limão ‘Rugoso’
tangerina ‘Kinnow’ + tangerina ‘Bendizao’ Deng et al. (2000) pomelo ‘Star Ruby’ + cidra ‘Corse’
Quadro 2 - Híbridos somáticos interespecíficos de Citrus.
Parentais Referências
laranja ‘Trovita’ + P. trifoliata Ohgawara et al. (1985)
laranja ‘Hamlin’ + P. trifoliata ‘Flying Dragon’ Grosser et al. (1988a)
tangerina ‘Cleópatra’ + citrumelo ‘Swingle’ Grosser et al. (1992b) laranja ‘Valência’ + kunquat ‘Meiwa’ Deng et al. (1992) laranja ‘Valência’ + citrange ‘Carrizo’ Louzada et al. (1992) laranja ‘Succari’ + P. trifoliata ‘Argentine’
laranja azeda + P. trifoliata ‘Flying Dragon’ Grosser et al. (1994)
kunquat ‘Meiwa’ + tangerina ‘Dancy’
laranja ‘Hamlin’ + Microcitrus papuana Grosser et al. (1996b)
Quadro 3 - Híbridos intergenéricos de citros, produzidos por hibridação somática a partir de pais sexualmente compatíveis.
Parentais Referências
laranja ‘Hamlin’ + Severinia disticha Grosser et al. (1988b)
laranja ‘Hamlin’ + Citropsis gilletiana Grosser & Gmitter Junior (1990b)
tangerina ‘Cleópatra’ + Citropsis gilletiana Grosser et al. (1990)
laranja ‘Hamlin’ + Severinia buxifolia Grosser et al. (1992b)
tangerina ‘Ponkan’ + Citropsis gabunensis Ling & Iwamasa (1994)
tangelo ‘Seminole’ + Atalantia monophylla
tangelo ‘Seminole’ + Severinia buxifolia Motomura et al. (1995)
laranja ‘Succari’ + Severinia disticha
tangelo ‘Nova’ + Citropsis gilletiana Grosser et al. (1998)
tangerina ‘Cleópatra’ + Severinia disticha
tangelo ‘Nova’ + C. ichangensis Grosser et al. (1996b)
kunquat ‘Mame’ + P. trifoliata Miranda et al. (1997)
tangelo ‘Page’ + Murraya paniculata Guo & Deng (1998)
Parentais Referências
Microcitrus sp. + laranja azeda Vardi et al. (1989)
laranja ‘Navel’ + pomelo Ohgawara et al. (1989) laranja ‘Valência’ + limão ‘Feminello’ Tusa et al. (1990)
C. sudachi + limão ‘Galego’
C. sudachi + limão ‘Eureka’ Saito et al. (1993)
C. unshiu + laranja ‘Washington Navel’ Yamamoto & Kobayashi
(1995)
C. microcarpa + laranja azeda ‘Keen’
tangerina ‘Cleópatra’ + laranja azeda laranja ‘Valência’ + limão ‘Femminello’
Grosser et al. (1996a)
tangelo ‘Seminole’ + limão ‘Lisboa’ Moriguchi et al. (1996) tangelo ‘Seminole’ + limão ‘Rugoso’ Moriguchi et al. (1997) tangor ‘Murcote’ + laranja ‘Bonanza’ Li & Deng (1997) tangelo ‘Page’ + limão ‘Rugoso’ Guo et al. (1998) pomelo ‘Star Ruby’ + limão cv. LAC Ollitrault et al. (2000b)
aclimatizadas
laranja ‘Caipira’ limão ‘Cravo’ 11 Mendes-da-Glória et al. (2000);
Farias (2001)
limão ‘Cravo’ tangerina ‘Cleópatra’ 5 Latado (1998)
laranja ‘Valência Rohde Red’ limão ‘Volkameriano’ 2 laranja ‘Rubi Blood’ limão ‘Volkameriano‘ 13
tangerina ‘Cleópatra’ limão ‘Volkameriano‘ 12 Costa (2001)
limão ‘Cravo’ tangerina ‘Sunki’ 11
laranja ‘Valência’ cv. 68 Fortunella obovata 3
tangerina ‘Cleópatra’ laranja Azeda 23
limão ‘Cravo’ laranja Azeda 7
laranja ‘Caipira’ tangerina ‘Cleópatra’ 9 Mendes et al. (2001)
laranja ‘Caipira’ limão ‘Volkameriano’ 4
laranja ‘Caipira’ limão ‘Rugoso da Flórida’ 4
De acordo com Grosser et al. (1995) e Grosser & Chandler (2000), híbridos somáticos têm potencial para reter muitos dos atributos positivos de seus parentais. Porta-enxertos autotetraplóides têm mostrado grande potencial para controlar o tamanho das plantas (Lee et al., 1990, citados por Grosser et al., 1995), e observações preliminares de híbridos alotetraplóides também sugerem isto.
Com relação à análise do comportamento dos híbridos somáticos em condições de campo, Kobayashi et al. (1991) observaram que as plantas apresentaram bom desenvolvimento, com fertilidade de óvulo e pólen. Observações preliminares dos híbridos somáticos de porta-enxertos, contendo
Poncirus trifoliata como parental, indicam o desenvolvimento de árvores com
tamanho reduzido (Grosser et al., 1998a), o que permite plantios mais adensados (Grosser et al., 1995).
Outros resultados mostram que plantas de 4 anos de idade, enxertadas sobre o híbrido somático de laranja ‘Hamlin’ + limão ‘Rugoso’, plantadas sob alta pressão de seleção de resistência ao declínio, não apresentaram sintomas desta doença e obtiveram teste negativo para proteínas específicas do declínio (Grosser et al., 1998c).
Segundo Grosser et al. (2000b), plantas enxertadas no híbrido somático tangerina ‘Cleópatra’ + citrumelo ‘Swingle’ (e muitos outros híbridos somáticos similares) são menores do que as árvores de qualquer um dos seus parentais. Os híbridos somáticos que combinam trifoliata ‘Flying Dragon’ com laranja doce, pomelo, tangerina ‘Cleópatra’ e laranja azeda são mais adaptáveis ao viveiro do que ‘Flying Dragon’ (Grosser et al., 1998a).
Plantas de laranja ‘Valência Rohde Red’, sobre porta-enxerto híbrido somático de laranja azeda + lima doce ‘Palestina’, produziram um pouco a mais de frutos do que esta variedade sobre laranja azeda, ainda que as plantas com híbridos tivessem metade do tamanho das árvores com laranja azeda (Grosser & Chandler, 2000).
Muitos híbridos somáticos têm sido selecionados para resistência à
Phytophthora, podendo-se citar híbridos intergenéricos sexualmente compatíveis,
como laranja ‘Hamlin’ + P. trifoliata cv. ‘Flying Dragon’, sexualmente
incompatíveis, como laranja ‘Succari’ + Citropsis gilletiana e híbridos
interespecíficos, sexualmente compatíveis, como laranja ‘Succari’ + limão ‘Rugoso 8166’, todos produzidos no Citrus Research and Education Center (CREC) e citados por Grosser et al. (1995). Híbridos produzidos com os parentais de laranja doce + limão, têm apresentado uma resistência considerável, o que não era esperado pois ambos parentais são considerados e testados como sendo suscetíveis a esta doença (Grosser et al., 1998b). Híbridos somáticos com laranja trifoliata 50-7 foram selecionados por apresentarem elevada resistência à
Phytophthora nicotiana em testes realizados em casa-de-vegetação (Widmer et
al., 1998).
A hibridação somática têm se mostrado adequada também para a produção de porta-enxertos resistentes à nematóides. Híbridos somáticos com
Atalantia, Citropsis, Fortunella, Microcitrus e Citrus ichangensis têm mostrado
boa resistência ao nematóide Rodopholus citrophilus (Grosser et al., 2000b).
‘Kinkoji’ e tangerina ‘Cleópatra’ + limão ‘Rugoso’, se mostraram resistentes a este mesmo nematóide, e também ao nematóide do café (Pratylenchus coffeae).
2.7 Considerações finais
A predominância do uso do limão ‘Cravo’ como porta-enxerto na citricultura parece seriamente ameaçada devido a sua suscetibilidade ao declínio e, mais recentemente, à Morte Súbita dos Citros. Dados apresentados por Salva (2001) mostram que o declínio vem sendo a causa de uma súbita substituição, nos últimos anos, deste porta-enxerto por outros que apresentam alta tolerância à doença, como tangerina ‘Cleópatra’ e citrumelo ‘Swingle’.
Por ser pouco tolerante à tristeza, a laranja azeda, que chegou a representar 90% dos porta-enxertos utilizados na citricultura brasileira, teve de ser substituída. Seu uso têm sido insignificante, uma vez que é indicada, atualmente, apenas como porta-enxerto de limões.
Nas últimas décadas, muitos porta-enxertos foram selecionados como promissores, como é o caso da laranja ‘Gou-tou’, provável híbrido da laranja Azeda, que tem se mostrado tolerante à tristeza e à seca e resistente à
Phytophthora, e C. pennivesiculata, fenotipicamente similar ao limão ‘Cravo’, que
tem apresentado desempenho no campo superior ao do limão ‘Cravo’ e da laranja azeda (Pompeu Junior, 2001).
O limão ‘Cravo’ e a laranja azeda têm sido utilizados como parentais em fusões de protoplastos a fim de combinar características complementares no híbrido somático, especialmente em relação à resistência a doenças, além de características agronômicas desejáveis.
dos citros é muito estreita (Nicolosi et al., 2000). Estes autores concluíram que o limão ‘Cravo’ pode ser um híbrido entre cidra e laranja Azeda e que a laranja Azeda pode ser considerada um híbrido entre toranja e tangerina. Há tempos, cidras, toranjas e tangerinas têm sido consideradas espécies verdadeiras de citros e, as demais, seriam derivadas de hibridações entre estas três espécies verdadeiras (Scora, 1975 e Barrett & Rhodes, 1976, citados por Nicolosi et al., 2000).
3 MATERIAL E MÉTODOS
O presente trabalho foi desenvolvido no Laboratório de Biotecnologia de Plantas Hortícolas do Departamento de Produção Vegetal da Escola Superior de Agricultura “Luiz de Queiroz” (USP/ESALQ).
3.1 Estabelecimento e cultivo de calos embriogênicos
Óvulos abortados foram extraídos de frutos maduros de laranjas doces ‘Valência’ cv. 63, ‘Pêra’ cv. 158, ‘Natal’ e ‘Hamlin’ (C. sinensis L. Osbeck);
tangerinas ‘Cleópatra’ (C. reshi Hort. ex Tanaka), ‘Sunki’ (C. sunki Hort. ex
Tanaka), ‘Ponkan’ e ‘Changsha’ (C. reticulata Blanco); tangor ‘Murcote’ (C.
reticulata Blanco x C. sinensis L. Osbeck) coletados de plantas matrizes do
pomar do Departamento de Produção Vegetal, da USP/ESALQ, e do Banco de Matrizes do Centro APTA Citros “Sylvio Moreira”, em Cordeirópolis.
Após a desinfestação com solução de hipoclorito de sódio na concentração de 3:1 (v/v), por 20 minutos, seguida de três lavagens em água deionizada e autoclavada, introduziram-se 20 óvulos por placa de Petri (15 x 100 mm) contendo o meio de cultura MT semi-sólido (Murashige & Tucker, 1969) modificado pela adição de 500 mg.l-1 de extrato de malte (EME 0,146 M),
acrescido de 5 mg.l-1 de 6-benzilaminopurina (BAP), para indução à formação de calos embriogênicos, tendo sido mantidos no escuro, a 27 ºC.
mg.l-1 de carvão ativado, sem regulador vegetal. Para manutenção, os calos foram subcultivados em meio fresco a cada 4 semanas.
3.2 Estabelecimento e cultivo de células em suspensão
Pequenas porções dos calos de cada variedade (cerca de 500 mg) foram introduzidas em Erlenmeyers contendo os meios líquidos EME e H+H, modificados pela adição de 80 g.l-1 de sacarose, mantidos no escuro, à temperatura média de 27 ºC, sob agitação de 100 rpm.
Para a manutenção das células em suspensão, as culturas foram subcultivadas a cada 2 semanas, tendo sido utilizadas como fonte de protoplastos somente após 8 semanas de cultivo.
3.3 Seleção de plantas matrizes de C. grandis
Frutos maduros de C. grandis, das variedades ‘Chinesa’, ‘Pink C.E.S.
2607’, ‘Hawaiian’, ‘Siamesa’, ‘151/427’, ‘151/428’, ‘Melancia’, ‘Vermelha’, ‘Kao Panne’, ‘Indochina’, ‘Sunshine’, ‘Singapura’, ‘Ogami’, ‘Inerme’, ‘Indian Red’, ‘Nakorn’, ‘Doce’, ‘Lau Tau’, ‘Zamboa’ e ‘Periforme’foram coletados do Banco de Matrizes do Centro APTA Citros “Sylvio Moreira”, em Cordeirópolis, no Estado de São Paulo, em setembro de 2000, tendo sido devidamente higienizados, antes da retirada de suas sementes.
Fitopatologia da USP/ESALQ, tendo sido comprovada a presença de
Phytophthora sp. As bandejas foram mantidas em estufa telada coberta com
sombrite 50%, onde receberam apenas regas freqüentes.
Em março de 2001, foi feita a contagem do número de plantas germinadas em relação ao número total de sementes introduzidas. Para cada variedade, foram selecionadas as plantas mais vigorosas, tendo sido considerados os parâmetros altura da planta, diâmetro do caule e número de folhas. As plantas selecionadas foram transplantadas para sacolas plásticas contendo o mesmo solo utilizado para a germinação das sementes, a fim de manter a condição de estresse estabelecida pela presença do fungo. Nesta etapa, as plantas receberam adubações freqüentes e pulverizações preventivas com sulfato de cobre.
Em junho de 2001, as plantas foram podadas a fim de induzir brotações laterais, a partir do que já foi possível coletar folhas jovens e tenras, que foram utilizadas na hibridação somática como fonte de protoplastos de mesófilo foliar.
Para o isolamento de protoplastos foram utilizadas, ainda, plantas de toranja enxertadas sobre limão ‘Cravo’, das variedades ‘Doce’, ‘151/428’, ‘Indian Red’, ‘151/427’, ‘Kao Panne’, ‘Zamboa’, ‘Vermelha’, ‘Melancia’, ‘Pink C.E.S. 2607’, ‘Lemelo 150/926’, ‘Satsumelo’ e ‘Chinesa’. Estas plantas foram cultivadas em substrato Plantmax (Eucatex), sem a presença de Phytophthora spp., e
mantidas em casa-de-vegetação, onde receberam os mesmos cuidados dispensados aos ‘seedlings’.
3.4 Hibridação somática
Para o isolamento de protoplastos de calos embriogênicos e células em suspensão, cerca de 500 mg de calos foram colocados em cada placa de Petri (15 x 58 mm), contendo 2 ml do meio BH3 0,7 M (Grosser & Gmitter Junior,
1990a). Sobre os calos, foram adicionados, gota a gota, 2 ml da solução enzimática de Grosser & Chandler (1987), composta de 1% de Cellulase Onozuka R.S. (Yakult Pharmaceutical Ind. Co. Ltda.), 1% de Macerozyme R-10 (Yakult Pharmaceutical Ind. Co. Ltda.) e 0,2 % de Pectolyase Y-23 (Seishin), dissolvida em 8 ml do meio BH3 0,7 M (Grosser & Gmitter Junior, 1990a). As placas foram
vedadas com Parafilm.
Para o isolamento de protoplastos de mesófilo foliar, folhas jovens e tenras das plantas enxertadas e dos ‘seedlings’ selecionados de toranja, foram coletadas e desinfestadas em solução de hipoclorito de sódio na proporção de 2:1 (v/v), por 20 minutos, seguido de três lavagens em água deionizada e autoclavada. Com o auxílio de um bisturi, retirou-se a nervura central e foram feitos finos cortes no sentido transversal, ao longo da folha, para facilitar a penetração da solução enzimática e a liberação dos protoplastos. As folhas foram transferidas para Erlenmeyer de 250 ml, contendo 3 ml da solução enzimática de Grosser & Chandler (1987), que foram vedados e, juntamente com as placas, mantidas no escuro, sob agitação de 40 rpm, por 14-16 horas.
3.4.2 Purificação dos protoplastos
Os protoplastos isolados foram purificados, inicialmente, pela passagem da solução de isolamento contendo os protoplastos por peneira de nylon com malha de 50 µm e centrifugação do líquido resultante por 5 minutos a 700 rpm (100 g).