Universidade de Trás-os-Montes e Alto Douro
Efeito do enriquecimento ambiental no comportamento de tigres (Panthera tigris) em cativeiro
Dissertação de Mestrado em Engenharia Zootécnica
Marta Sofia Leocádio Teixeira
Orientador: Professor José Júlio Gonçalves Barros Martins
iv
Universidade de Trás-os-Montes e Alto Douro
Efeito do enriquecimento ambiental no comportamento de tigres (Panthera tigris) em cativeiro
Dissertação de Mestrado em Engenharia Zootécnica
Marta Sofia Leocádio Teixeira
Orientador: Professor José Júlio Gonçalves Barros Martins
Composição do Júri:
Professor Victor Manuel de Carvalho Pinheiro Professor Severiano José Cruz da Rocha e Silva Professor José Júlio Gonçalves Barros Martins
ii Declaro que esta dissertação de mestrado é resultado da minha pesquisa e trabalho pessoal e das indicações do meu orientador. Todo o conteúdo apresentado nesta dissertação é da exclusiva responsabilidade da autora.
iii Agradecimentos
Ao meu orientador Prof. Doutor José Júlio Gonçalves Barros Martins, por toda a sua disponibilidade, apoio e auxilio prestado na realização desta dissertação.
Ao Zoo da Maia por me ter aceite de braços abertos e por me ter dado a liberdade em todas as tarefas idealizadas. Em especial à Eng. Andreia Silva e à Paula Telinhos que me ajudaram na integração e realização deste estudo no Jardim Zoológico da Maia. A toda a restante equipa técnica e a todos os funcionários do Jardim Zoológico da Maia que me receberam carinhosamente e com toda a hospitalidade.
Aos meus companheiros neste estudo, Asha, Jasmim e Ankur por me acompanharem em todas as horas de observação.
Ao Prof. Doutor Severiano Silva pela ajuda prestada para a análise dos dados.
A todos os meus amigos e colegas de Biologia e Engenharia Zootécnica, com quem partilhei momentos de grande aprendizagem mas também de muita farra e diversão.
Às quatro melhores amigas que a vida académica me poderia ter dado, Francisca, Ju, Renata e Sara e ao Tiago, obrigada por estarem sempre ao meu lado, são sem dúvida muito importantes para mim e nada teria sido tão maravilhoso sem vocês.
Ao meu namorado, Vasco, que sempre me acompanhou em todas as aventuras e sonhos. Sempre acreditou em mim e me deu força para continuar a lutar pelo que acredito.
Por fim, resta-me agradecer aos meus pilares, a minha família. Aos meus pais que sempre acreditaram em mim e sempre me deram força para construir o meu caminho. À minha mãe que mesmo quando as coisas corriam menos bem, estava ao meu lado com todo o amor, compreensão e dedicação.
v Resumo
O enriquecimento permite aumentar a complexidade do ambiente em cativeiro e possivelmente aumentar o bem-estar animal, através da estimulação de comportamentos ativos e específicos da espécie, permitindo também reduzir os comportamentos estereotipados comuns em felinos alojados em zoos.
O presente estudo analisou o efeito do enriquecimento ambiental nos padrões de comportamento em três tigres (Panthera tigris) alojados em cativeiro no Zoo da Maia. Neste estudo foram adicionados dois períodos diferentes de enriquecimento: sensorial olfativo (E1) e ocupacional (E2). Em cada um dos enriquecimentos, 2 itens foram colocados no recinto exterior dos felinos com o objetivo de avaliar os efeitos nos níveis de atividade, diversidade comportamental e estereotipias. Cada um dos enriquecimentos teve a duração de 10 dias, sendo apresentado aos animais como enriquecimento canela em pó, fezes de ungulados (E1); blocos de peixe congelado e um poste de madeira com um saco de sarapilheira pendurado (E2).
Foram registados 38 comportamentos e agrupados em categorias 9; a frequência média de cada comportamento para o geral dos animais foi comparada antes (baseline), durante (E1, E2) e após as fases enriquecimentos (após E).
O enriquecimento resultou num aumento significativo na frequência das categorias exploração, alimentar e locomoção. Não se observaram diferenças significativas na estereotipia pacing perante o efeito enriquecimento, talvez devido ao reduzido tamanho da amostra ou reduzida frequência inicial deste comportamento. No entanto, o enriquecimento diminuiu a frequência de comportamentos relativos a interações agonísticas.
Em conclusão, a canela, as fezes de ungulados, os blocos de gelo com peixe e o poste com saco de sarapilheira no topo são itens de enriquecimentos baratos e fáceis de introduzir nos recintos, que podem aumentar os comportamentos ativos e melhorar o bem-estar de tigres em cativeiro.
vi Abstract
Enrichment can increase the complexity of the captive environment and possibly enhance captive animal’s welfare by stimulating active and species specific’s behaviours and also reducing stereotypical behaviours commonly seen in zoo felids.
The presente study examined the effect of environmental enrichment on behavior patterns of three captive tigers (Panthera tigris) housed in Zoo da Maia. In this study, two different enrichment time periods: olfactory (E1) and occupational (E2). To each enrichment 2 items were added to outdoor enclosures of felids to test their effects on activity levels, behavior’s diversity and stereotypic pacing. The cinnamon, ungulate feces (E1) and frozen fish balls and a wooden pole with burlap sack on the top (E2) were presented over a 10 days period to the animals.
The 38 behaviours were grouped into 9 categories; the mean frequency that the animals present to each behavior was compared before (baseline), during (E1, E2) and after enrichment treatment (após E).
The enrichment resulted in a significant increase of frequency in exploratory, feeding and locomotor categories. A significant difference in stereotypic pacing was not observed across treatment, perhaps due to the small sample number or the diminished frequency of this behavior. However, the enrichment decrease the frequency of agonistic behaviors.
In conclusion, cinnamon, ungulate feces, frozen fish balls and wooden pole with burlap sack on the top are inexpensive and easy to administer enrichment items that may be used to increase active behaviors and improve the welfare of captive tigers.
vii
Sumário
Agradecimentos ... iii Resumo ... v Abstract ... vi Índice de figuras... ix Índice de tabelas ... x Índice de quadros ... x Índice de gráficos ... x Lista de abreviaturas ... xi 1. Introdução ... 1 2. Revisão Bibliográfica ... 3 2.1 O tigre ... 3 2.1.1 Taxonomia e evolução ... 32.1.2 Distribuição geográfica e habitat ... 4
2.1.3 Subespécies ... 6 2.1.4 Estatuto de conservação ... 11 2.1.5 Morfologia ... 12 2.2 Biologia do tigre ... 14 2.2.1 Longevidade ... 14 2.2.2 Reprodução ... 14
2.2.3 Predação e hábitos alimentares ... 15
2.2.4 Comportamentos e comunicação ... 16
2.3 Conservação Ex-situ ... 17
2.3.1 Papel dos zoos... 17
2.3.2 Alojamento ... 19
2.3.3 Alimentação em cativeiro ... 21
2.3.4 Bem-estar animal ... 22
2.3.5 Comportamento de tigres em cativeiro ... 23
viii 3. Material e métodos ... 29 3.1 Local do estudo ... 29 3.2 Animais em estudo... 29 3.3 Habitat e maneio ... 31 3.4 Delineamento do estudo ... 33
3.4.1 Antes do enriquecimento (Baseline) ... 34
3.4.2 Enriquecimento (E) ... 34
3.4.3 Após enriquecimento (Após E) ... 35
3.5 Observações sistemáticas ... 36
3.6 Análise de dados... 37
3.6.1 Etograma e análise de comportamentos ... 37
3.6.2 Análise estatística ... 41 4. Resultados ... 43 4.1 Comportamentos lúdicos ... 43 4.2 Comportamentos de locomoção ... 46 4.3 Comportamentos de manutenção ... 47 4.4 Comportamentos alimentares ... 50 4.5 Comportamentos de marcação ... 52
4.6 Comportamentos de interação afiliativa ... 54
4.7 Comportamentos de interação agonística ... 55
4.8 Comportamentos de inatividade ... 57
4.9 Comportamentos estereotipados ... 58
4.10 Categorias, efeito geral do enriquecimento... 59
5. Discussão ... 61
6. Conclusão ... 67
7. Considerações finais ... 69
8. Referências bibliográficas ... 71
ANEXOS ... 77
ix ANEXO B - Alimentação trimestral ... 80 ANEXO C - Folha de registo ... 81 ANEXO D - Comportamentos ... 82
Índice de figuras
Figura 1 As sequências de DNA nas 37 espécies de felinos foram comparadas com objetivo de
determinar a sua árvore genealógica. Os fósseis forneceram as datas que indicam quando ocorreu a grande ramificação. Adaptado de O’Brien 2007. ... 3
Figura 2 Escala de tempo geológica. Grande variação do nível do mar ao longo do Pleistoceno
contribuiu para a distribuição geográfica dos tigres. Adaptado de O’Brien 2007. ... 4
Figura 3 Distribuição geográfica histórica e atual das nove subespécies de tigre. As linhas a
tracejado constituem os limites aproximados entre subespécies. O istmo de Kra divide a subespécie Indochinesa e o tigre Malaio. Adaptado de Luo et al., 2010. ... 5
Figura 4 Classificação taxonómica da espécie tigre. ... 6 Figura 5 Distribuição geográfica das populações existentes em meio selvagem. A amarelo estão
representadas as áreas onde atualmente se encontram tigres e a vermelhos a as áreas onde estes estão possivelmente extintos. Os dados referentes a cada país surgem da última estimativa global da população de tigres. Fonte: (Goodrich et al., 2015);(Harley, 2016). ... 11
Figura 6 Crânio de leão (cima) e de tigre (baixo). Os crânios são muito semelhantes em tamanho
e especto geral, contudo existem algumas diferenças que os distinguem quando vistos de cima como as suturas apresentadas. Adaptado de Turner A., 1997. ... 12
Figura 7 Diferenças na cor e padrão na pelagem de tigre. Através da localização destes
exemplares induz-se que (A) Sumatra, tigre de Sumatra; (B) Índia, tigre de Bengala; (C) Malaia, tigre Malaio; (D) Bangladesh, tigre de Bengala. Fonte: Luo et al., 2010. ... 14
Figura 8 Reflexo de flehmen. Fonte: autora ... 17 Figura 9 Aparelho hioide. O ligamento (indicado pela seta) não é ossificado nos grandes felinos,
garantindo ao tigre a capacidade de rugir. Fonte: Turner, A. e Antón, M., 1997. ... 17
Figura 10 Ilustração de um reptilário no Zoo de Londres, no período Vitoriano onde os animais
eram exibidos muito próximo uns dos outros Fonte: http://www.victorianlondon.org/ ... 18
Figura 11 Formas de estereotipias locomotoras - pacing comuns em animais carnívoros. Estas
envolvem caminhar numa rota fixa – para trás e frente, em círculo ou em forma de oito. Fonte: Clubb, 2006. ... 24
Figura 12 Planta do zoo. ... 29 Figura 13 Itens utilizados no enriquecimento 1. Canela (E1.1) e fezes de ungulados (E1.2). ... 34 Figura 14 Itens utilizados no enriquecimento 2. Poste com saco (E2.1) e bloco de gelo (E2.2).
... 35
x Índice de tabelas
Tabela 1 Características e alguns parâmetros das nove subespécies de tigre. O peso e
comprimento são correspondentes a machos (M) e fêmeas (F). Fonte imagens:
http://www.animalesextincion.es/; http://www.worldwildlife.org/ ... 10
Tabela 2 Principais categorias de enriquecimento e subdivisões. Métodos de enriquecimento utilizados em tigres em cativeiro. Adaptado de Bloomsmith et al. (1991). ... 26
Tabela 3 Identificação e características dos tigres do Zoo da Maia... 31
Tabela 4 Exemplo de relatório trimestral da alimentação dos tigres do Zoo da Maia. ... 33
Tabela 5 Grupos e condições de enriquecimento ambiental utilizados no estudo. ... 36
Tabela 6 Etograma e terminologia explicativa dos comportamentos dos tigres do Zoo da Maia. ... 39
Índice de quadros Quadro 1 Frequência média dos comportamentos lúdicos (ocorrências por 70 minutos). ... 45
Quadro 2 Frequência média dos comportamentos de locomoção (ocorrências por 70 minutos). ... 47
Quadro 3 Frequência média de comportamentos de manutenção (ocorrências por 70 minutos). ... 49
Quadro 4 Frequência média de comportamentos alimentares (ocorrências por 70 minutos). .. 51
Quadro 5 Frequência média de comportamentos de marcação (ocorrências por 70 minutos). 53 Quadro 6 Frequência média de comportamentos afiliativos (ocorrências por 70 minutos). ... 54
Quadro 7 Frequência média de comportamentos agonísticos (ocorrências por 70 minutos). ... 56
Quadro 8 Frequência média de comportamentos de inatividade (ocorrências por 70 minutos). ... 57
Quadro 9 Frequência média de comportamentos estereotipados do tipo “pacing” (ocorrências por 70 minutos). ... 58
Índice de gráficos Gráfico 1 Frequência média e SEM dos comportamentos lúdicos com efeito do enriquecimento. ... 44
Gráfico 2 Frequência média e SEM do comportamento locomotor “escalar” no efeito enriquecimento. ... 46
Gráfico 3 Frequência média e SEM dos comportamentos alimentares com efeito do enriquecimento. ... 51
Gráfico 4 Frequência média e SEM dos comportamentos de interação agonística com efeito do enriquecimento. ... 56
xi Lista de abreviaturas
Após E – pós-enriquecimento Ca – cálcio
CITES – Convention on International Trade in Endangered Species of Wild Fauna and Flora
cm – centímetro
DNA - ácido desoxirribonucleico mtDNA – DNA mitocondrial E1 – enriquecimento 1 E2 – enriquecimento 2
IUCN – International Union for Conservation of Nature Kcal - quilocaloria kg – quilograma km - quilómetro km2 – quilómetro quadrado m – metro m2 – metro quadrado Ma – milhão de anos p – probabilidade PVC - policloreto de polivinila SEM – erro padrão da média
SSC - Species Survival Commission UI – unidades internacionais
vs - versus
1
1. Introdução
O tigre (Panthera tigris) é certamente um dos felinos mais facilmente reconhecíveis pela sua pelagem listrada única. Esta espécie inclui também o maior felino vivo atualmente. Contudo, os tigres enfrentam fortes pressões de caça furtiva e perda de habitat, sendo forçados a competir pelo território com as densas populações humanas (Kitchener & Yamaguchi, 2010).
Os grandes felinos no seu meio selvagem têm geralmente extensas áreas territoriais, onde realizam comportamentos típicos da espécie como perseguição e captura associados à caça. No entanto, apesar dos jardins zoológicos terem como fim a educação, conservação e investigação (Mench, 1996), os alojamentos dos animais frequentemente não permitem, em todo ou em parte, a expressão desses comportamentos naturais, na maior parte dos casos por limitações espaciais e pelas reações humanas negativas a comportamentos predatórios (Mellen et al., 1998). Possivelmente, como consequência dessas restrições, muitos animais desenvolvem comportamentos anormais (Mason et al., 2007).
Sensíveis a esta problemática, muitos parques zoológicos têm vindo a implementar alterações na gestão dos felinos e dos seus alojamentos (Carlstead & Shepherdson, 1994). Exemplo disso é a adoção de práticas de enriquecimento ambiental, em que são aplicadas alterações nos alojamentos e maneio dos animais com o objetivo de os estimular psicológica e fisicamente. Pretende-se assim que estes possam expressar os seus comportamentos naturais, reduzindo os comportamentos anormais e melhorando o seu bem-estar (Bashaw et al., 2003).
Este estudo teve como objetivo, estudar o efeito do enriquecimento ambiental no comportamento de tigres em cativeiro. Para isso, foram introduzidos novos elementos no recinto dos animais, os quais permitiram aumentar a complexidade do alojamento, criando novas interações entre os animais e o meio envolvente. Estas alterações visavam despoletar novos interesses e, consequentemente, proporcionar maior ocupação física e psicológica, aumentando a atividade e reduzindo a frequência de estereotipias que são os comportamentos anormais mais comuns nos felinos em cativeiro.
3
2. Revisão Bibliográfica
2.1 O tigre
2.1.1 Taxonomia e evolução
O aparecimento do que hoje conhecemos como felinos modernos começou com a divergência da linhagem Panthera (Oken, 1816) das espécies de felinos ancestrais, há cerca de 10.8 milhões de anos (Ma) (Fig. 1). Alguns milhões de anos mais tarde, esta linhagem divergiu em dois grupos. Um deles era composto por duas espécies de leopardo-nebuloso (Buckley-Beason et al., 2006; Kitchener et al., 2006), englobando o outro os grandes felinos do género Panthera: o leão (Panthera leo), jaguar (Panthera onca), leopardo-das-neves (Panthera uncia), leopardo (Panthera pardus) e o tigre (Panthera tigris) (Johnson et al., 2006; O'Brien & Johnson, 2007). A separação da linhagem Panthera foi seguida de rápidos eventos de divergência e migração, há cerca de 3.7 Ma, originando as cinco espécies Panthera atualmente existentes. Algumas espécies Panthera derivadas da Ásia ocuparam a América (jaguar e leão), África (leão e leopardo) e outras permaneceram no seu local de origem (tigre, leopardo-das-neves e leopardo-nebuloso) (Hemmer, 1987; Johnson et al., 2006).
Figura 1 As sequências de DNA nas 37 espécies de felinos foram comparadas com objetivo de
determinar a sua árvore genealógica. Os fósseis forneceram as datas que indicam quando ocorreu a grande ramificação. Adaptado de O’Brien 2007.
4 Até o final do Plioceno e início do Pleistoceno, o tigre foi amplamente distribuído pelo leste Asiático (Hemmer, 1987; Mazák, 1996). A alternância entre os períodos frios (glacial) e quentes (inter-glacial) resultou na mudança do nível do mar ao longo do Pleistoceno (Fig. 2) o que provavelmente causou repetidas restrições e expansões na distribuição geográfica e abundância de tigres (Hemmer, 1987; J. et al., 1999; Kitchener & Dugmore, 2000). Observações em algumas espécies Felidae modernas (O'Brien & Johnson, 2005) mostram que o tigre tem uma diversidade genética populacional relativamente baixa, na sequência das reduções demográficas relativamente recentes e / ou eventos fundadores (Luo et al., 2004)
Figura 2 Escala de tempo geológica. Grande variação do nível do mar ao longo do Pleistoceno
contribuiu para a distribuição geográfica dos tigres. Adaptado de O’Brien 2007.
Através da análise do DNA mitocondrial (mtDNA) foi possível estabelecer que o ancestral comum mais recente do tigre teve origem há cerca de 72.000-108.000 anos (Luo et al., 2004). Pensa-se, por isso, que o tigre surgiu há pouco tempo, em comparação com os leopardos modernos que se pensa terem origem em África há entre 470.000 e 825.000 anos (Uphyrkina et al., 2001).
O tempo de coalescência do mtDNA do tigre moderno (ou seja, a fusão de linhagens anteriores) deverá ter ocorrido há cerca de 73.500 anos atrás, coincidindo com uma catastrófica erupção vulcânica em Toba, Sumatra, o maior evento vulcânico explosivo conhecido na terra (Rampino & Self, 1992). Esta erupção devastadora durante o Pleistoceno Superior pode estar também associada a fenómenos de bottleneck na evolução humana (Ambrose, 1998), ao grande evento de dispersão para norte do elefante asiático (Fleischer et al., 2001) e talvez também tenha contribuído para uma enorme redução de tigres na pré-história (Luo et al., 2010b).
2.1.2 Distribuição geográfica e habitat
Estudos paleontológicos indicam que o tigre evoluiu numa área mais ou menos restrita do leste da Ásia e posteriormente colonizou a Ásia ocidental, através dos Himalaias, da Ásia Central e das pontes de terra para as Ilhas de Sunda, em consequência da descida do nível do mar durante a formação dos glaciares. Quando as temperaturas globais aumentaram, as ilhas ficaram novamente reduzidas pela elevação das marés, as florestas aumentaram e os
5 desertos e prados diminuíram. Portanto, o período glacial restringiu a distribuição geográfica do tigre, mas impulsionou o fluxo de genes para as Ilhas de Sunda, enquanto que o período interglacial impediu o fluxo de genes para as ilhas, mas incentivou a vasta colonização do continente (Kitchener & Yamaguchi, 2010).
Parte da capacidade do tigre para persistir a alterações da paisagem deve-se á sua flexibilidade adaptativa, pois estes animais toleram uma ampla gama de condições de habitat (Fig.3).
Figura 3 Distribuição geográfica histórica e atual das nove subespécies de tigre. As linhas a
tracejado constituem os limites aproximados entre subespécies. O istmo de Kra divide a subespécie Indochinesa e o tigre Malaio. Adaptado de Luo et al., 2010.
Estes animais podem viver em florestas tropicais, ou florestas de pinheiro, carvalho e bétula, no Extremo Oriente russo, sobrevivendo a temperaturas de −34ºC. Esta adaptação é resultante de um estilo de vida com gastos energéticos relativamente baixos, o que se traduz em elevadas taxas de sucesso na captura de presas, perseguições curtas e longos intervalos entre a morte de grandes presas (Miquelle et al., 2005).
O tamanho da área territorial dos tigres varia de acordo com a densidade de presas. Fêmeas de tigre de Bengala têm áreas de 200 a 1000 Km2, sendo que a área média ocupada
6 esconderijos, muitas vezes entre a vegetação densa ou em cavernas e em cavidades debaixo de árvores. Estes grandes felinos podem percorrer 16 a 32 km numa única noite. Estes geralmente defendem o seu território mas também há conhecimento de o compartilharem pacificamente (Mazák, 1981; Sunquist & Sunquist, 2002).
2.1.3 Subespécies
O reconhecimento de uma subespécie requer a descrição de caracteres hereditários objetivos que cada indivíduo da subespécie carrega e que estão realmente identificados para a mesma. Isto é, eles encontram-se apenas nessas subespécies e não em outras populações da mesma espécie (Fig. 4) (Avise & Ball, 1990).
Os membros de uma subespécie compartilham uma área geográfica ou habitat, um grupo de caracteres fenotípicos concordantes filogeneticamente e uma história natural única em relação a outras subdivisões da espécie (O'Brien & Mayr, 1991).
Reino: Animalia Filo: Chordata Classe: Mammalia Ordem: Carnivora Família: Felidae Subfamília: Pantherinae Género: Panthera
Espécie: Panthera tigris (Linnaeus, 1758)
Figura 4 Classificação taxonómica da espécie tigre.
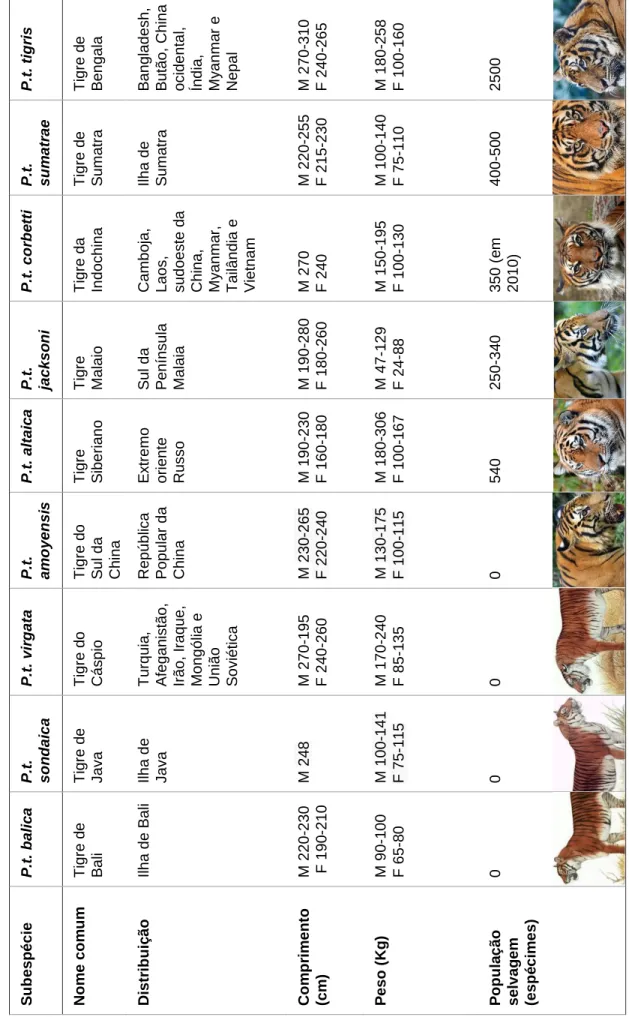
Conforme ilustrado na tabela 1, atualmente são reconhecidas nove subespécies de tigre (Goodrich et al., 2015). Destas, três já se encontram extintas e uma quarta provavelmente só sobrevive em cativeiro (Seidensticker, 2010).
Tradicionalmente, estas subespécies foram definidas devido á sua distribuição geográfica combinada com características morfológicas, tais como o tamanho do corpo, particularidades do crânio, cor da pelagem e padrão das listras (J. et al., 1999).
Das subespécies já extintas fazem parte: (1) P.t. virgata (Illiger, 1815), o tigre do Cáspio, descrito com base em caracteres de pelagem e um número desconhecido de espécimes
7 (Illiger, 1815); (2) P.t. sondaica (Temminck, 1844), o tigre de Java (Temminck, 1844); e (3) P.t. balica (Schwarz, 1912), o tigre de Bali, também classificado com base num número limitado de espécimes (Hilzheimer, 1905; Pocock, 1929; Schwarz, 1912).
Nos tempos modernos as análises genéticas demonstraram uma história genealógica única e independente para a distinção e reconhecimento de cinco e possivelmente, seis subespécies de tigre atuais: (1) P.t. altaica (Temminck, 1844), o tigre Siberiano; (2) P.t. amoyensis (Hilzheimer, 1905), o tigre do Sul da China sobrevive apenas em cativeiro; (3) P.t. corbetti (Mazák, 1968), o tigre da Indochina; (4) P.t. jacksoni (Luo et al. 2005), o tigre malaio; (5) P.t. sumatrae (Pocock, 1929), o tigre de Sumatra; e (6) P.t. tigris (Linnaeus, 1758), o tigre de Bengala.
2.1.3.1 Panthera tigris tigris – Tigre de Bengala
O território do tigre de Bengala abrange o Bangladesh, Butão, China ocidental, Índia, Myanmar ocidental e o Nepal (Nowell & Jackson, 1996), onde habita em florestas subtropicais e tropicais, florestas húmidas e secas. Este é o símbolo nacional da Índia e de Bangladesh, sendo a mais numerosa e a segunda maior subespécie de tigre. Os machos atingem um comprimento total entre 270 a 310 cm e um peso compreendido entre 180 e 258 kg, enquanto as fêmeas variam de 240 a 265 cm e possuem entre 100 a 160 kg de peso vivo (Mazák, 1981).
2.1.3.2 Panthera tigris sumatrae – Tigre de Sumatra
Encontrado em estado selvagem somente na ilha de Sumatra, na Indonésia, esta é a menor das subespécies atualmente existentes. O tamanho dos machos varia entre 220 a 255 cm e o seu peso de 100 a 140 kg, enquanto as fêmeas medem entre 215 e 230 cm, apresentando um peso de 75 a 110 kg (Mazák, 1981). O seu tamanho reduzido revela a adaptação às densas florestas e presas menores que existem no seu habitat nativo. Possui uma zona nasal larga e curta, um plano occipital largo e uma pelagem com inúmeras listras relativamente amplas (Hemmer, 1987; Mazák, 1981).
2.1.3.3 Panthera tigris corbetti – Tigre da Indochina
Pode ser encontrado no Camboja, Laos, sudoeste da China, Myanmar, Tailândia e Vietnam. Habita florestas tropicais e florestas secas, preferencialmente em regiões montanhosas. Possui uma coloração mais escura comparativamente a outras subespécies, com listras duplas, curtas e raras (Hemmer, 1987; Mazák, 1981; Nowell & Jackson, 1996).
Os machos têm em média 270 cm de comprimento e pesam entre 150 e 195 kg, enquanto as fêmeas têm em média 240 cm e 100 a 130 kg (Mazák, 1981).
8 2.1.3.4 Panthera tigris jacksoni – Tigre Malaio
Presente exclusivamente no sul da Península Malaia. Não era considerada uma subespécie até 2004 e era tida como parte da subespécie indochinesa. Tal classificação mudou em função de um estudo genético, sendo a única subespécie identificada com base em marcadores moleculares, que mostrou possuírem mtDNA e sequências de micro-satélite distintas da subespécie da Indochina (Luo et al., 2004). No entanto, Mazák e Groves (2006) não encontraram diferenças morfológicas claras (em medições cranianas ou características da pelagem) entre os tigres da Península Malásia e os que habitam noutros lugares na Indochina (Mazák & Groves, 2006).
A divisão da subespécie tradicional, tigre da Indochina, em dois grupos tão distintos, tem implicações significativas para a compreensão da geografia regional do Sudeste Asiático. Os resultados apoiam a hipótese de que o Istmo de Kra tem sido uma barreira ecológica, restringindo o fluxo génico entre populações de tigres na península da Malásia e a Indochina. Na verdade, o Istmo de Kra é considerado uma transição biogeográfica significativa (Whitmore, 1984).
A sua descrição científica formal, de acordo com o Código Internacional de Nomenclatura Zoológica, é ainda inexistente, tornando este taxon um nomen nudum (ICZN, 1999). Ainda assim, o nome de subespécie proposto "Panthera tigris jacksoni" tem sido amplamente utilizado, por exemplo, pela IUCN / SSC Lista Vermelha de Espécies Ameaçadas (Goodrich et al., 2015). Nesta subespécie os machos apresentam um comprimento total de 190 a 280 cm e pesam entre 47 a 129 kg, enquanto as fêmeas variam de 180 a 260 cm com um peso de 24 a 88 kg (Khan, 1986).
2.1.3.5 Panthera tigris altaica – Tigre Siberiano
O tigre Siberiano, ao contrário das outras subespécies, vive numa área de clima frio, sendo o habitat caracterizado pela existência de florestas boreais (de carvalhos e coníferas) e está confinado quase inteiramente ao Extremo oriente Russo e fronteira da China com a Coreia do Norte (Luo et al., 2010a).
É a maior das subespécies de tigre e o maior felino existente atualmente. Os machos têm um comprimento de 190 a 230 cm e pesam entre 180 e 306 kg. Já as fêmeas têm em média 160 a 180 cm e um peso de 100 a 167 kg (Mazák, 1981). Estes tigres apresentam reduzida diversidade genética em comparação com outras subespécies, possivelmente como resultado da era pós-gelo ou da intensa perseguição humana no início do século XX, o que levou a que os efetivos desta subespécie tenham atingido números na casa dos 20 a 30 animais (Luo et al., 2004).
9 2.1.3.6 Panthera tigris amoyensis – Tigre do Sul da China
Entre todas as subespécies, o tigre do Sul da China é a mais controversa, classificada como criticamente ameaçada e funcionalmente extinta do meio selvagem (Nyhus, 2008).
Na década de 1950, existiam aproximadamente 4.000 tigres na República Popular da China. No entanto, a realização de campanhas de erradicação do tigre em grande escala combinadas com a extensa perda de habitat reduziram a população desta subespécie. Estima-se que em 1982 já só sobrevivessem 150-200 indivíduos na natureza (Lu & Sheng, 1986). Em 2007 existiam apenas 72 exemplares em cativeiro (Luo et al., 2008), descendentes de somente 6 exemplares com herança genética indeterminada (Tilson et al., 1997).
Os machos variam de 230 a 265 cm de comprimento e tem um peso compreendido entre 130 a 175 kg. As fêmeas têm entre 220 e 240 cm de comprimento e de 100 a 115 kg de peso (Mazák, 1981).
2.1.3.7 Panthera tigris virgata – Tigre do Cáspio
Entre 1920 e 1970, a população de tigres na Ásia Central diminuiu e desapareceu por razões comuns a outras subespécies como a caça, a conversão do seu habitat limitado pelo cultivo e concomitante declínio de presas (Mazák, 1996, 1981). O tigre do Cáspio tornou-se extinto em fevereiro de 1970, quando o último sobrevivente foi morto a tiro na Turquia (Mazák, 1996). Esta subespécie era também encontrada no Afeganistão, Irão, Iraque, Mongólia e União Soviética (Jackson, 2011). O tamanho total para os machos varia de 270 a 195 cm e o seu peso de 170 a 240 kg, já as fêmeas tinham de 240 a 260 cm e pesavam 85 a 135 kg (Mazák, 1981).
2.1.3.8 Panthera tigris sondaica – Tigre de Java
Esta subespécie apenas existia na ilha de Java, foi extinta na década de 1970 em resultado da caça e destruição do seu habitat. Os tigres machos pesavam 100 a 141 kg com cerca de 248 cm de comprimento, as fêmeas pesavam 75 a 115 kg (Mazák, 1981).
2.1.3.9 Panthera tigris balica – Tigre de Bali
A sua existência era limitada à ilha de Bali, na Indonésia. Estes tigres foram caçados até a extinção. O último exemplar desta subespécie foi uma fêmea adulta, provavelmente morta em 1937 em Sumbar Kima, oeste de Bali (Seidensticker, 1987).
Esta era a menor de todas as subespécies com tamanho comparável ao leopardo; os machos tinham 220 a 230 cm de comprimento e pesavam 90 a 100 kg e as fêmeas mediam 190 a 210 cm com peso entre 65 e 80 kg (Mazák, 1981).
10 P .t . tig ris T igr e d e B e n g a la B a n g lad e sh , B u tã o , Chin a o cide n ta l, Ín d ia, M ya n m a r e Nep a l M 2 7 0 -310 F 2 4 0 -265 M 1 8 0 -258 F 1 0 0 -160 2500 P .t . s u m a tr a e T igr e d e S u m a tr a Ilh a de S u m a tr a M 2 2 0 -255 F 2 1 5 -230 M 1 0 0 -140 F 7 5 -1 1 0 400 -5 0 0 P .t . c o rb e tt i T igr e d a In d o ch ina Cam b o ja, L a o s, su d o e ste d a Chin a , My a n m a r, T a ilâ n d ia e V iet n a m M 2 7 0 F 2 4 0 M 1 5 0 -195 F 1 0 0 -130 3 5 0 ( e m 2010) P .t . ja c k s o n i T igr e M a laio S u l d a P e n ín su la M a laia M 1 9 0 -280 F 1 8 0 -260 M 4 7 -1 2 9 F 2 4 -88 250 -3 4 0 P .t . a lt a ic a T igr e S ibe rian o E xtr e m o o rien te Russo M 1 9 0 -230 F 1 6 0 -180 M 1 8 0 -306 F 1 0 0 -167 540 P .t . a m o y e n s is T igr e d o S u l d a Chin a Rep ú b lica P o p u lar d a Chin a M 230 -265 F 2 2 0 -240 M 130 -175 F 100 -115 0 P .t . v ir g a ta T igr e d o Cásp io T u rq u ia , A fe g a n istã o , Ir ã o , Ir a q u e , M o n g ó lia e Uniã o S o viét ica M 2 7 0 -195 F 2 4 0 -260 M 1 7 0 -240 F 8 5 -1 3 5 0 P .t . s o n d a ic a T igr e d e Ja va Ilh a d e Ja va M 248 M 100 -141 F 7 5 -1 1 5 0 P .t . b a li c a T igr e d e B a li Ilh a d e B a li M 2 2 0 -230 F 1 9 0 -210 M 9 0 -1 0 0 F 6 5 -8 0 0 S u b e s p é c ie No m e c o m u m Dis tr ib u iç ã o Co m p rim e n to (c m ) P e s o ( Kg ) P o p u la ç ã o s e lv a g e m (e s p é c im e s ) T a b e la 1 Car a cte rís tica s e a lgu n s p a râ m e tr o s d a s n o ve su b e sp é cies d e tig re . O p e so e c o m p rim e n to sã o co rr e sp o n d e n te s a m a ch o s ( M ) e fê m e a s ( F ). F o n te i m a g e n s: h tt p :/ /www .a n im a lese x tin cion .e s/; h tt p :/ /www .wo rldwil d life .o rg /
11 2.1.4 Estatuto de conservação
Existem diversos fatores que colocam os tigres no sensível estatuto em que se encontram atualmente. Apesar dos constantes esforços aplicados na conservação e manutenção desta espécie, é inegável a importância económica para os parques zoológicos onde estes animais são exibidos.
Por outro lado, os tigres são mortos ilegalmente pela sua pele, servindo para tapetes e peças decorativas. Além disso, a medicina tradicional chinesa utiliza partes de tigre para tratar doenças e lesões. Embora não haja evidências de efeito sobre a doença, o úmero deste animal continua a ser prescrito para tratar o reumatismo (Sunquist & Sunquist, 2002).
Por norma, os tigres evitam o contacto humano e muito raramente estes se tornam "caçadores humanos". Geralmente os tigres que atacam homens são aqueles que não podem efetivamente caçar grandes presas porque se tornaram aleijados, velhos ou já não têm um habitat adequado e presas disponíveis. O aumento das populações humanas e a competição pelos recursos naturais tem vindo a aumentar a pressão sobre os tigres e o seu habitat, aumentando a probabilidade de interações negativas entre o Homem e o tigre (Mazák, 1981; Sunquist & Sunquist, 2002).
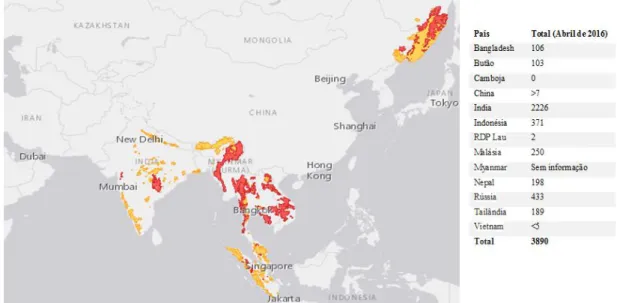
As ameaças específicas aos tigres variam regionalmente, mas a perseguição humana, a caça e destruição do habitat são fatores universais na diminuição das populações de tigre. Apesar do perigo de extinção que ameaça a espécie há vários anos, os dados mais recentes mostram que o número de tigres selvagens está em ascensão. Após um século de declínio constante, em 2016 foram contabilizados mais 700 tigres, totalizando 3.890 no censo global mais recente (Fig. 5) (WWF, 2016).
Figura 5 Distribuição geográfica das populações existentes em meio selvagem. A amarelo estão
representadas as áreas onde atualmente se encontram tigres e a vermelhos a as áreas onde estes estão possivelmente extintos. Os dados referentes a cada país surgem da última estimativa global da população de tigres. Fonte: (Goodrich et al., 2015);(Harley, 2016).
12 Atualmente o tigre é classificado como em perigo (EN), na Lista Vermelha de Espécies Ameaçadas da IUCN (Goodrich et al., 2015) e está listado no Anexo I do CITES (CITES, 2016).
2.1.5 Morfologia
Como a maioria dos felinos, o tigre mantém o plano corporal primário do seu ancestral remoto. Esta espécie representa uma das duas que atualmente atingiram o tamanho máximo para este plano corporal nos felinos vivos atualmente. Se usarmos o comprimento do crânio como um indicador do tamanho corporal, é mais uma vez claro que os maiores leões e tigres são aproximadamente do mesmo tamanho (Fig. 6). Há muito debate sobre se os tigres são os maiores dos felinos, mas estudos mostram que o tigre selvagem moderno, provavelmente, tem uma média de peso corporal (peso corporal estimado, sem o conteúdo estomacal) de 160 kg para os machos adultos e 115 kg para a fêmea adulta, enquanto a média para o leão macho adulto selvagem é 175 kg e para a fêmea adulta de 120 kg. Por esse facto, o leão é muitas vezes reconhecido como o maior felino vivo. No entanto, como a variação de tamanho intraespecífica parece ser maior no tigre que no leão, ao considerarmos cada subespécie destes dois grandes felinos e o tamanho dos seus indivíduos podemos afirmar que o tigre possui os maiores felinos vivos atualmente (Kitchener & Yamaguchi, 2010).
Figura 6 Crânio de leão (cima) e de tigre (baixo). Os crânios são muito semelhantes em tamanho
e especto geral, contudo existem algumas diferenças que os distinguem quando vistos de cima como as suturas apresentadas. Adaptado de Turner A., 1997.
13 2.1.5.1 Variação do tamanho corporal do tigre
Existem diversos fatores evidentes para que a variação geográfica possa influenciar o tamanho corporal dos tigres (Kitchener, 1999) e que tem por base os seguintes pressupostos: Regra de Bergmann (Geist, 1987): o tamanho corporal aumenta com a latitude e consequente descida da temperatura, assim a relação superfície/volume decresce; em latitudes a norte os tigres maiores perdem menos calor e necessitam de relativamente menos energia do que os tigres menores.
Regra de Guthrie Geist (Geist, 1987): o tamanho do corpo aumenta com o aumento da latitude, devido a um aumento da duração e amplitude do pico da produtividade sazonal. Por outras palavras, quando os mamíferos estão a crescer durante a primavera e verão, há mais alimento disponível para que atinjam melhor o seu potencial de crescimento. Porém, a altas latitudes, a produtividade sazonal diminui e uma redução no tamanho do corpo seria de esperar, mas os tigres não se encontram tão a norte para que isso aconteça.
Island Dwarfing (Lister, 1994): grandes mamíferos muitas vezes apresentam um tamanho muito menor nas ilhas. O tigre de Bali foi provavelmente o menor dos tigres contemporâneos. Possivelmente, o reduzido tamanho corporal resultaria da limitada disponibilidade de presas de grande porte.
Podemos assim constatar que o tamanho corporal varia consideravelmente entre subespécie de tigres, existindo animais de grandes dimensões no extremo leste da Rússia até animais mais pequenos nas Ilhas de Sunda (Mazák, 1996, 1981). No continente as fêmeas não variam muito em tamanho, contudo há muita variação entre machos de diferentes subespécies (Mazák, 1981). Isso parece sugerir que o dimorfismo sexual se revela no aumento do tamanho do corpo de tigres machos.
2.1.5.2 Coloração e marcas
A coloração base nos tigres varia de amarelo claro até ao vermelho escuro (Fig. 7); (Mazák, 1996, 1981; Pocock, 1929) os tigres mais escuros são encontrados em florestas tropicais húmidas, enquanto pelagens mais claras são encontradas em florestas temperadas menos húmidas (Ortolani, 1996). No entanto, a variação dentro das populações pode ser considerável e pode variar com a estação e exposição de horas luz no verão (Kitchener, 1999). Os tigres têm sobre a pelagem listras pretas verticais ao longo dos flancos e ombros que variam em tamanho, comprimento e espaçamento. A parte inferior das pernas, barriga, peito, garganta e focinho são brancos ou claros. O branco é também encontrado acima dos olhos e estende-se para as bochechas. Uma mancha branca está presente na parte de trás de cada orelha. As linhas escuras sobre os olhos tendem a ser simétricas, mas as marcas em
14 cada lado da face são frequentemente assimétricas. A cauda é laranja-avermelhado e rodeada com várias faixas escuras (Mazák, 1981; Sunquist & Sunquist, 2002).
A sua pelagem laranja listrada levaria a pensar que este se destaca num fundo verde tropical, mas combina com os padrões florestais de sol e sombra, sendo uma camuflagem perfeita para este grande predador.
Figura 7 Diferenças na cor e padrão na pelagem de tigre. Através da localização destes
exemplares induz-se que (A) Sumatra, tigre de Sumatra; (B) Índia, tigre de Bengala; (C) Malaia, tigre Malaio; (D) Bangladesh, tigre de Bengala. Fonte: Luo et al., 2010.
2.2 Biologia do tigre 2.2.1 Longevidade
Os tigres geralmente vivem 15 anos em estado selvagem, no entanto em cativeiro é bastante comum os tigres viverem até aos 20 anos, podendo chegar aos 26 anos de idade. Estima-se que a maioria dos tigres adultos morre como resultado da perseguição e caça humana, contudo também as suas presas de grande porte podem, ocasionalmente, feri-los fatalmente. Os jovens tigres enfrentam inúmeros perigos quando se dispersam da área de proteção da progenitora, incluindo ser atacados e comidos por tigres machos. A taxa de sobrevivência para as crias ronda os 50% (Mazák, 1981; Sunquist & Sunquist, 2002).
2.2.2 Reprodução
Os tigres são solitários e apenas procuram o sexo oposto para o acasalamento, neste período o macho e a fêmea podem ficar juntos durante 20 a 80 dias. Após esse período os tigres adultos machos não fornecem cuidados parentais (Sunquist & Sunquist, 2002).
15 As fêmeas entram em estro a cada 3 a 9 semanas e estão recetivas durante 3 a 6 dias, revelando comportamentos de cio como vocalizações e marcas de urina. A cópula é rápida e ocorre várias vezes ao longo do dia. O período de gestação dura aproximadamente 103 dias e a ninhada tem em média 3 crias. A progenitora passa a maior parte do tempo a cuidar das crias durante esta fase vulnerável. O desmame ocorre entre os 90 a 100 dias de idade mas os jovens tigres começam a seguir a mãe com cerca de 2 meses de idade e a ingerir a primeira comida sólida. Dos 5 aos 6 meses de idade começam a participar nas caçadas, permanecendo com a mãe até terem entre 18 meses e 3 anos de idade. Os jovens tigres atingem a maturidade sexual por volta dos 3 a 4 anos de idade, no caso das fêmeas, e 4 a 5 anos nos machos (Karanth, 2001; Sunquist & Sunquist, 2002).
2.2.3 Predação e hábitos alimentares
Os tigres preferem caçar à noite, quando as suas presas são mais ativas. Eles utilizam a audição e visão para localizar as suas presas, recorrendo menos ao olfato (Schaller, 1967). Usam uma abordagem furtiva, tirando partido de cada rocha, árvore e mato como cobertura e raramente perseguem presas durante longas distâncias (> 150 m). Como outros felinos, os tigres são digitígrados, ou seja, eles andam sobre os dedos, e as macias almofadas do pé distribuem o seu peso, dando aos tigres não só um movimento de andar fluido mas também silencioso. Estes agem através de passos cautelosos e permanecem junto ao chão para que não possam ser vistos ou ouvidos. Geralmente matam por emboscada, deixando a presa sem equilíbrio assim que saltam para ela. Os tigres são excelentes caçadores, no entanto apenas 1 em cada 10 a 20 ataques é bem-sucedido e resulta na morte da presa (Mazák, 1981; Schaller, 1967; Sunquist & Sunquist, 2002). São utilizadas duas possíveis táticas, consoante o tamanho da presa. Os animais pequenos, de peso inferior a metade do peso do corpo do tigre, são mortos por uma mordida na parte de trás do pescoço. Os caninos são inseridos entre as vértebras do pescoço, forçando-as a afastar e seccionando a medula. Nos animais maiores, uma mordida na garganta é usada para esmagar a traqueia do animal e sufocá-lo. Os tigres raramente se alimentam no local de captura e arrastam presas de até 700 kg para vegetação densa, antes de se começarem a alimentar (Karanth & Sunquist, 2000). Protegem o seu alimento de outros predadores e mantêm-se perto da carcaça (Karanth, 2001).
A sua língua é coberta com duras papilas que permitem retirar a carne dos ossos e os seus dentes caninos são os mais longos entre os felinos vivos, com 7,5 a 10 cm de comprimento (Sunquist & Sunquist, 2002).
Os tigres são carnívoros e como tal a sua da dieta é maioritariamente composta por várias espécies de grandes ungulados, como o chital, o veado, o búfalo e ungulados domésticos, como bovinos, cavalos e caprinos. Em casos raros os tigres podem atacar
16 elefantes e rinocerontes. Quando ocorre escassez de alimento, capturam aves, peixes, crocodilos, ratos e sapos (McGregor, 2010).
Os tigres podem ingerir entre 18 e 40 kg de carne quando caçam grandes presas, sendo o restante consumido ao longo de vários dias (Mazák, 1981; Schaller, 1967; Sunquist & Sunquist, 2002). A quantidade máxima que um tigre pode comer em 24 horas é cerca de um quinto do seu próprio peso corporal, o que para um grande macho se traduz em 45 kg (McDougal, 1997).
Estes animais não têm predadores naturais, sendo o ser humano o seu único predador conhecido. No entanto, os tigres adultos machos podem ser potenciais predadores de crias e juvenis da sua espécie (Schaller, 1967; Sunquist & Sunquist, 2002).
2.2.4 Comportamentos e comunicação
Os tigres são animais solitários, assim a única relação a longo prazo pode ser observada entre a mãe e a sua descendência. Alguns comportamentos que estes animais exibem, demostra que são seres admiráveis. Estes felinos têm uma grande capacidade de saltar, sendo capazes de o fazer até distâncias de 8 a 10 metros. Outra característica de destaque é o fato de os tigres serem excelentes nadadores. A água não costuma ser um obstáculo à sua movimentação, sendo que os tigres podem facilmente atravessar rios com 6-8 km de largura. Estes felinos são também bons trepadores, fazendo uso das suas garras retráteis e pernas poderosas (Mazák, 1981; Sunquist & Sunquist, 2002).
A comunicação entre os tigres é mantida por marcas olfativas, sinais visuais e vocalização. As marcações olfativas são depositadas sob a forma de um líquido de odor almiscarado que é misturado com a urina e pulverizado sobre as árvores, erva ou rochas. Estas servem para os tigres marcarem o seu território e identificarem outros indivíduos. A deteção de odores é associada a um comportamento típico denominado flehmen. Durante o reflexo de flehmen, a língua paira sobre os incisivos, o nariz está enrugado, e os caninos superiores estão descobertos (Fig. 8). Esta reação é comumente vista em machos após cheirarem urina, marcações odoríferas, fêmeas em estro ou crias da sua espécie. Ao esfregar o seu corpo ou cabeça numa superfície ou objeto o tigre deixa também a sua marca olfativa (Karanth, 2001; Schaller, 1967; Sunquist & Sunquist, 2002).
17 Figura 8 Reflexo de flehmen.
Os sinais visuais incluem a interpretação da postura corporal e marcas de garras deixadas em árvores ou outros objetos. As manchas brancas na parte de trás das orelhas, e o seu padrão listrado podem também ser usados na comunicação intraespecífica (Mazák, 1981; Schaller, 1967; Sunquist & Sunquist, 2002). Os tigres podem comunicar vocalmente de várias formas como: rugir, rosnar, miar, assobiar, soprar. Cada som tem seu próprio propósito e parece refletir a intenção ou o humor do tigre. Por exemplo, o rugido de um tigre é geralmente um sinal de posição dominante; transmite a outros indivíduos o quão grande ele é e a sua localização. Um gemido demonstra submissão, já o sopro é associado a interações afiliativas. A capacidade do tigre rugir, característica dos grandes felinos, vem da presença de um ligamento elástico no aparelho hioide da garganta, representado na figura 9 (Karanth, 2001; Schaller, 1967; Sunquist & Sunquist, 2002).
Figura 9 Aparelho hioide. O ligamento (indicado pela seta) não é ossificado nos grandes
felinos, garantindo ao tigre a capacidade de rugir. Fonte: Turner, A. e Antón, M., 1997.
2.3 Conservação Ex-situ 2.3.1 Papel dos zoos
A origem dos jardins zoológicos surge por volta de 3000 a.C. no antigo Egipto, onde eram mantidas grandes coleções de animais exóticos. A posse de animais surgiu por duas
18 grandes razões: (1) muitas das espécies tinha significado religioso; (2) os animais exóticos eram considerados um símbolo de poder (Bostock, 1993).
As coleções de animais selvagens são conhecidas ao longo da história, como a que era mantida pela nobreza nos tempos medievais na Torre de Londres. Os zoos modernos ganharam popularidade no século XIX, como um lugar que proporcionava prazer e entretenimento aos seus visitantes (Fig. 10) (Brown, 2014).
A Associação Americana de Zoos e Aquários (AZA) desenvolveu uma definição de jardins zoológicos e aquários com base nos seus padrões de acreditação:
Uma instituição cultural permanente que possui e preserva animais selvagens em cativeiro que representam mais que uma coleção e, sob a direção de uma equipa profissional oferece o cuidado adequado, apresentando-os de um modo estético e regular ao público. Deve também ser definido que tem como principal atividade a exposição, conservação e preservação da fauna da Terra, de forma pedagógica e científica (AZA, 2016a).
Outras definições também podem existir segundo outras associações nacionais e regionais.
Os três jardins zoológicos mais antigos conhecidos estão em conformidade com o reconhecimento geral de um zoológico moderno, e são o de Viena (1752), Madrid (1772) e Paris (1793). São caracterizados por serem ao ar livre, instalados num jardim ou parque (pelo menos em parte) e abertos ao público (Brown, 2014). Atualmente, muitos desses zoos mais antigos não tem um papel educacional, científico ou de conservação, contudo o Jardin des Plantes, em Paris, e o London Zoo têm um forte papel científico. A história dos zoos mostra o contexto evolutivo em que os animais têm sido mantidos e reflete as mudanças até à sociedade atual.
Figura 10 Ilustração de um reptilário no Zoo de Londres, no período Vitoriano onde os animais
19 Atualmente, o número de tigres em cativeiro é superior ao que existe em estado selvagem (Nyhus et al., 2010). Por esse motivo, estudar a influência deste ambiente no comportamento dos animais poderá fornecer respostas para compreender e melhorar a sua vida (Kirkwood, 2003; Mason et al., 2007). As linhagens cativo-adaptadas apresentam problemas, sendo assim difícil estudar o comportamento natural da espécie, utilizar programas de reprodução ou reintrodução no meio selvagem (Kirkwood, 2003; Mason et al., 2007). Concomitantemente, o bem-estar psicológico dos animais deve ser garantido (Hutchins & Smith, 2003), uma vez que estes possuem consciência de sensações agradáveis e / ou desagradáveis, o que os leva a sofrer quando o bem-estar é reduzido (Kirkwood, 2003). O comportamento torna-se assim uma ferramenta útil para identificar estes sentimentos e avaliar a condição de bem-estar.
Apesar do objetivo principal dos zoos e outras instituições similares ser a conservação, estes devem ter aliadas as áreas educativa, de pesquisa e lazer (McPhee, 2004; Mench, 1996). Para tal, os animais devem poder expressar os seus padrões normais de comportamento, garantindo o seu valor educacional. Resumidamente, as instituições zoológicas devem também garantir todos os aspetos do bem-estar animal, incluindo a saúde básica, bem-estar psicológico e simular as condições de vida naturais (Fraser, 2009).
2.3.2 Alojamento
Não existem critérios europeus definidos quanto às práticas de manutenção de tigres. A sua exposição irá depender dos recursos, clima e interesses da instituição. No entanto, a AZA reconhece as boas instituições com base em alguns critérios:
1) espaço ao ar livre relativamente grande;
2) presença de piscinas, fossos ou águas correntes; 3) vegetação natural; e
4) reduzir ou evitar barreiras entre os tigres e o público (Bush et al., 1994).
Na escolha do local para o recinto dos tigres é importante garantir que este não está posicionado muito perto de presumíveis presas. Embora esse seja um bom estímulo para o tigre, seria provavelmente um fator de stress para as presas. No entanto, se as condições ambientais e a topografia o permitir, o recinto pode ser projetado num local a favor do vento, proporcionando enriquecimento para os tigres, desde que não submeta os outros animais a um stress constante (Baker, 2006).
Os recintos devem sempre dispor de uma zona de recolha dos animais em que estes ficam fora do alcance do público. Isto é importante para garantir a segurança e facilitar a
20 manutenção, além de permitir aos tratadores acompanhar de perto o estado do animal. Eles podem passar a noite nesta área que deve ter pelo menos 2,4 metros por 2,4 metros (AZA, 2016b). Cada tigre deve ter o seu próprio espaço, equipado com uma fonte de água e uma plataforma onde poderá dormir (Baker, 2006).
O tamanho, natureza e capacidades físicas do tigre requerem várias exigências e meios de segurança. A jaula para um único animal deve medir pelo menos 12 m de largura e 12 m de profundidade (144 m²) (AZA, 2016b); as jaulas devem ser aumentadas em 50% por cada animal adicionado (Shoemaker et al., 1997). Nos recintos exteriores as paredes/vedações devem ter pelo menos 5 m de altura, onde no topo 1 metro deverá ter um ângulo de 45 graus para o interior. Estas devem ser inescaláveis e assentar numa base de cimento para evitar que os animais possam cavar por baixo. Outra maneira de impedir a fuga por baixo da cerca é enterrá-la pelo menos 1 m, com um ligeiro ângulo para o interior do recinto (Bush et al., 1994).
Os recintos exteriores mais recentes apresentam uma exposição mais naturalista com vegetação e solo natural, contudo não devem existir grandes árvores perto da vedação que possam permitir o acesso dos tigres ao topo. As plantas escolhidas devem ser cuidadosamente selecionadas para evitar espécies tóxicas. As piscinas devem ser projetadas de forma a manter a qualidade da água através de filtração para facilitar a limpeza, pois os tigres tendem a defecar na água (Bush et al., 1994).
O uso de fios elétricos em volta do recinto tem como fim manter os animais a uma certa distância de algumas áreas. A utilização de vidro é esteticamente agradável, permite uma melhor visualização dos animais, mas exige maior manutenção como limpeza e pode ser vulnerável a fraturas (Bush et al., 1994).
Nas áreas onde o pavimento não é natural (como a zona de recolha) o material mais comum é o cimento, o que por si não é uma superfície ótima devido á porosidade, abrasividade e dureza. Contudo, pode ser facilmente limpo, desinfetado, seca rapidamente e o facto de não ser poroso evita a acumulação de detritos orgânicos e contaminação. Para uma limpeza efetiva deve ser utilizada água quente e detergente para remover os detritos, juntamente com um desinfetante. Os agentes de desinfeção deverão ser selecionados com base na eficácia e baixa toxicidade para os tigres. Os compostos fenólicos devem ser evitados devido à suscetibilidade dos felinos a este produto químico. O recinto deve ter escoamento capaz de drenar a chuva ou a água de lavagem. Geralmente boas instalações também oferecem a possibilidade de conduzir os tigres de uma área para outra. (Bush et al., 1994)
21 2.3.3 Alimentação em cativeiro
Em estado selvagem, os animais carnívoros geralmente comem na totalidade, ou quase, as presas que capturam, incluindo os ossos, gordura, vísceras e outras peças. Contudo o uso de carne como única dieta para os animais carnívoros é ainda muito utilizado. O músculo embora tenha boas fontes de aminoácidos, alguns minerais (p. ex. sódio, potássio, ferro, selénio e zinco) e vitaminas B (B6 e B12), apresenta baixos níveis de cálcio, magnésio e vitaminas lipossolúveis (vitamina D, E e, em muitos casos, vitamina A) (Baer et al., 2010), o que frequentemente resulta em doenças ósseas, desequilíbrios nutricionais e perda de capacidade reprodutiva (Baer et al., 2010).
Face a este quadro, os zoos, para alimentar os tigres, devem optar por dietas comercialmente preparadas ou carcaças suplementadas. As dietas comerciais têm a vantagem de estarem facilmente disponíveis, requererem pouco ou nenhum trabalho de preparação e são formuladas com uma base nutricional adequada à espécie, no entanto não se assemelham à dieta natural de um tigre (Bush et al., 1987).
Com o objetivo de estimular também o comportamento de caça destes felinos sem descurar a componente nutricional, pode ser fornecido um pedaço da carne com suplementos. Se a dieta é preparada no zoo, esta deve ser suplementada de modo a fornecer 1% de cálcio (na matéria seca), particularmente se os ossos não são consumidos. Isto equivale a cerca de 7g de Ca/kg de carne. Se a carne for magra, as vitaminas A e E podem ter de ser reforçadas até aos níveis recomendados. Não deve ser feita suplementação com vitamina A se o fígado for consumido. A vitamina E pode ser necessária em dietas à base de carnes que contenham gorduras polinsaturadas (qualquer não-ruminantes), em níveis que fornecem 50-150 UI/kg de matéria seca (Dierenfield et al., 1994).
Os tigres adultos são alimentados de modo a manter a sua condição corporal. No geral, as necessidades básicas de manutenção destes animais traduzem-se da seguinte forma: 140 Kcal x (massa corporal em kg) 0.75 . Isto significa que um macho com 160 kg necessita de
6.300 kcal por dia. A maioria dos programas de maneio em tigres concluiu que o apetite e condição corporal aumentam quando estes realizam jejum durante um ou dois dias por semana.
A quantidade da dieta deve aumentar em 10-20% em animais alojados no exterior durante os meses de inverno, decrescendo na mesma proporção durante o verão quando o apetite decresce (Dierenfield et al., 1994).
22 2.3.4 Bem-estar animal
As condições para promover o bem-estar animal foram descritas pela Comissão Brambell (1965). Esse documento, aplicado a animais de pecuária no Reino Unido, é conhecido como as Cinco Liberdades (Brambell, 1965):
1. Livre de sede, fome ou desnutrição; 2. Livre de desconforto;
3. Livre de dor, ferimentos e doenças;
4. Livre para exibir padrões normais de comportamento; 5. Livre de stress, medo e ansiedade.
A ausência de bem-estar é associada a indicadores fisiológicos negativos ou comportamentos anormais. Fisiologicamente, em situações adversas ocorre o aumento do ritmo cardíaco e respostas endócrinas, tais como a libertação de catecolaminas e cortisol. Simultaneamente há supressão de hormonas relacionadas com o anabolismo, o crescimento e a reprodução (Sapolsky, 2002).
Se no passado a ausência de comportamentos negativos era visto como um indicador de bem-estar (Swaisgood, 2007), atualmente o foco passa por identificar casos específicos de verdadeiro bem-estar (Yeates & Main, 2008). Desse modo, foram propostos vários princípios que incluem as mesmas abordagens estabelecidas nas cinco liberdades (Barnett & Hemsworth, 2009):
1. Minimizar as emoções negativas; 2. Maximizar as emoções positivas; 3. Garantir a adaptação;
4. Fornecer oportunidade para comportamentos naturais; 5. Fornecer ambientes naturais.
Os comportamentos naturais de um animal constituem um fator importante de muitas avaliações de bem-estar e podem ser definidos como:
Comportamento que os animais têm tendência a apresentar sob condições naturais, por estes comportamentos serem agradáveis e promoverem o funcionamento biológico (Bracke & Hopster, 2006).
Ao observar o comportamento animal é possível avaliar o seu estado de bem-estar através dos sinais auditivos, visuais, ou olfativos naturais da espécie. Os sinais específicos da espécie evoluíram para transmitir informações, como dor, angústia ou prazer a membros
23 da mesma espécie, deste modo os investigadores podem reunir informações sobre o estado de bem-estar de um animal (Mason & Veasey, 2010).
2.3.5 Comportamento de tigres em cativeiro
Em cativeiro o ambiente dos tigres é muito diferente do natural, onde estes vivem em grandes áreas (Sunquist, 2010). Nos jardins zoológicos são muitas vezes alojados com ou próximo de outros tigres (bem como de outras espécies) e as restrições espaciais e financeiras limitam o tamanho do recinto, resultando em habitats significativamente menores.
A ausência de presas vivas na sua alimentação e o fornecimento de dietas pré-preparadas não permite o desempenho dos seus comportamentos de caça naturais e consequente gasto energético. Do mesmo modo, Duckler (1998) descobriu que os crânios de tigres em cativeiro tinham protuberâncias que não são encontradas em espécimes selvagens. Este fenómeno tem origem na excessiva limpeza do pelo nos tigres em cativeiro e numa redução dos músculos da mandíbula devido à alimentação com alimentos processados.
Frequentemente é esperado que os tigres sejam ativos e visíveis durante o horário de funcionamento do jardim zoológico quando os visitantes estão presentes, o que entra em conflito com o seu padrão de atividade natural. Dadas estas diferenças, não é de estranhar que esta espécie exiba altos níveis de comportamentos anormais em cativeiro pois o seu ambiente difere radicalmente do que estes animais deveriam experienciar (Morgan & Tromborg, 2007). Os principais comportamentos anormais resultantes das limitações de um ambiente cativo são: automutilação, agressividade, comportamento alimentar anormal, comportamento maternal anormal, hiperatividade, apatia e estereotipias (Mason, 1991). Fatores como a personalidade e história de cada animal também podem influenciar estes comportamentos (Szokalski et al.).
2.3.5.1 Estereotipias
As estereotipias são definidas como movimentos morfologicamente semelhantes ou sequências de comportamento executadas repetidamente, sem qualquer função óbvia (Ődberg, 1987). As estereotipias desenvolvem-se como resposta à contenção física, falta de estimulação, medo ou frustração (Mason, 1991) e incluem ações como morder barras, brincar com a língua, baloiçar e limpeza do pelo excessiva (Rees, 2009).
A limitação do espaço é das causas mais referidas para o aparecimento do comportamento estereotipado. Na maioria dos casos, quanto menor o recinto, maior é a probabilidade de o animal exibir estereotipias (Carlstead, 1996). Lyons et al. (1997) estudaram
24 o padrão de comportamento de 19 espécies de felinos em cativeiro e descobriram que os animais alojados em recintos relativamente grandes, apresentavam níveis mais elevados de comportamentos exploratórios.
A reduzida diversidade de estímulos é outro fator que influencia o comportamento estereotipado. Em ambientes estéreis, os animais muitas vezes parecem aborrecidos devido à uma falta de novos elementos (Carlstead, 1996).
Nos tigres em cativeiro, as estereotipias mais comuns são as locomotoras, conhecidas como pacing. Esta estereotipia consiste em percorrer várias vezes o mesmo percurso (Fig. 11), em torno ou em parte de um recinto fechado ou nadar no mesmo percurso em torno de um tanque. Em muitos casos, a rota inclui caminhar ao longo da vedação do recinto repetidamente. Em situações extremas o animal caminha sobrepondo as suas próprias pegadas (Rees, 2015). Esta estereotipia pode causar dano físico direto, como abrasões, feridas ou abscessos (Mason, 1991), bem como reduzir a interação social e inibir o crescimento das crias (Carlstead & Shepherdson, 1994).
Figura 11 Formas de estereotipias locomotoras - pacing comuns em animais carnívoros. Estas
envolvem caminhar numa rota fixa – para trás e frente, em círculo ou em forma de oito. Fonte: Clubb, 2006.
Para os carnívoros, a hipótese mais citada como motivação e consequente estereotipia associada á locomoção surge a partir da forte motivação natural pela procura de alimento - foraging (Mason, 1993).
No meio selvagem, comportamentos como a procura de presas, perseguição, captura e morte das mesmas constituem comportamentos naturais para os tigres. Contudo num ambiente artificial, como um jardim zoológico esses comportamentos são anulados (Mason & Rushen, 2008). Assim, o facto de a locomoção ocorrer na fase mais estimulante da caça pode estar relacionado com o aparecimento de pacing (McFarland, 1987).
25 Concomitantemente, grandes felinos em cativeiro mostraram maior evidência de estereotipias nos dias em que não são alimentados – dias de jejum (Lyons et al, 1997; Ames, 2000).
2.3.6 Enriquecimento ambiental
O enriquecimento ambiental é geralmente associado à adição de estímulos sensoriais e escolhas no ambiente (Azevedo et al., 2007). Este propõe os seguintes objetivos:
(1) aumentar a diversidade comportamental;
(2) reduzir as frequências de comportamento anormal; (3) aumentar os comportamentos normais (selvagens); (4) aumentar a utilização positiva do ambiente; e
(5) aumentar a capacidade de lidar com os desafios de uma forma mais natural (Young, 2007a).
Apesar do grande contributo para o bem-estar animal, o enriquecimento pode trazer alguns inconvenientes, isto se não houver um bom delineamento. O tamanho impróprio dos objetos, estrangulamentos em cordas, envenenamentos ou contaminações alimentares e através de dejetos de outros animais, são fatores que se deve ter em conta no planeamento de qualquer tipo de enriquecimento. Do mesmo modo, o aumento dos custos de manutenção, trabalho adicional para os tratadores e falta de dados científicos que comprovem a consequente melhoria no bem-estar animal são também argumentos utilizados contra a prática de enriquecimento (Young, 2007b). Ainda assim, o comportamento estereotipado nos tigres em cativeiro não foi ignorado e nas últimas décadas diversas técnicas têm sido aplicadas (Hutchins & Smith, 2003). A maioria destes esforços centra-se em enriquecimentos que visam comportamentos alimentares e de caça naturais, também abordando o estímulo olfativo e o tamanho do território. Isto tem sido conseguido através da introdução de novos itens alimentares, alterações nas rotinas de alimentação e tipo de apresentação dos alimentos (Szokalski et al.).
Em última análise, o enriquecimento pode beneficiar tanto o animal (fisicamente/mentalmente) como o público visitante, indiretamente. O aumento da visibilidade e diminuição de comportamentos estereotipados contribuem para melhorar a experiência dos visitantes (Bashaw et al., 2003).