Biological profile of nanohydroxyapatite particles on isolated
and co-cultured osteoblastic and endothelial cells
Raquel Marinho Rodrigues Oliveira
MESTRADO EM BIOLOGIA CLÍNICA LABORATORIAL
VILA REAL, 2011
Dissertação orientada por: Professor Doutor Bruno Jorge Antunes Colaço Professora Doutora Maria Helena Raposo Fernandes
i
ACKNOWLEDGMENTS
Este espaço é dedicado a todos aqueles que contribuíram para a realização desta dissertação. A todos eles deixo aqui o meu sincero agradecimento.
Começo por agradecer à Coordenação de Curso e Comissão de Estágios, pela aceitação deste projecto de dissertação de mestrado.
Agradeço também ao Prof. Doutor Bruno Colaço, pela forma como orientou o meu trabalho. As recomendações e o permanente acompanhamento, aliados às palavras de encorajamento nos momentos mais complicados, foram primordiais para a realização deste trabalho.
Gostaria também de fazer um agradecimento especial à Prof. Doutora Maria Helena Fernandes por me ter orientado durante este ano e por me ter apresentado ao imenso mundo dos biomateriais. Agradeço os seus ensinamentos e conselhos, que em muito contribuíram para estimular a minha capacidade crítica face à investigação científica.
Gostaria ainda de agradecer a toda a equipa do Laboratório de Farmacologia e Biocompatibilidade Celular, da Faculdade de Medicina Dentária da Universidade do Porto, especialmente à Mestra Mónica Garcia pelas horas de orientação, disponibilidade e atenção.
Deixo também uma palavra de apreço ao Professor Doutor José Duarte, do Laboratório de Bioquímica e Morfologia Experimental da Faculdade de Desporto da Universidade do Porto, pela obtenção das imagens de microscopia electrónica de transmissão.
Agradeço também ao Professor Doutor Jorge Colaço, da Universidade de Trás-os-Montes e Alto Douro, pelo tratamento estatístico dos resultados finais.
Obrigada a todos os colegas que frequentaram este Mestrado, pelos momentos divertidos que me proporcionaram ao longo de todo o ano curricular, bem como por toda a ajuda que me deram para que, na UTAD me sentisse em casa.
ii
Não posso deixar de agradecer aos meus amigos, em particular à Tati e à Maria, que apesar da distância estão sempre disponíveis para oferecer bons conselhos e palavras amigas, e também à Viviana e à Raquel por me fazerem acreditar em novos começos.
Finalmente, agradeço aos meus pais e ao meu irmão pelo amor e apoio incondicionais e, pelo constante incentivo ao conhecimento, ao sonho e à aventura. A vós dedico este trabalho.
iii
ABSTRACT
Nano-hydroxyapatite (nanoHA) has a variety of proposed applications in bone repair/regeneration strategies. Most of the published work regarding the biological profile of nanoHA relies on films, composite scaffolds, dense or porous bulks or cements manufactured with HA nanoparticles. A clear deficit exists on studies addressing the biological assessment of individual nanosized HA particles. Considering the close association of angiogenesis and osteogenesis in bone formation events, this study aims to address the biological profile of nanoHA particles on isolated and co-cultured osteoblast and endothelial cells.
In the present study, nanoHA particles, sinthesized by a hydrothermal method, were characterized by X-ray diffraction analysis (XRD), Fourier transmission infrared spectroscopy (FTIR) and tansmission eletron microscopy (TEM). In vitro biological studies were performed using MG-63 osteoblast-like cells (103 cell cm-2) and human dermal microvascular endothelial cells (HDMEC, 2x104 cell cm-2) cultured isolated or co-cultured in endothelium medium. Cultures were maintained for 14 days in the absence (control) and in the presence of different concentrations of nanoparticles. NanoHA particles (1-100 μg ml-1) were added to the seeded cells at two different moments: day 1 and at day 7. Nanoparticles added at day 1: cultures were characterized at days 2, 7 and 14; nanoparticles added at day 7: cultures were evaluated at days 8(7) and 14(7), for cell viability/proliferation (MTT assay), pattern of cell growth (confocal layser scanning microscopy; CLSM), particle internalization and kinetics within cell compartiments (TEM).
Results showed that monocultures of MG-63 cells and HDMEC, maintained in the absence of nanoparticles, presented high viability and the expected pattern of cell growth. Also, control co-cultures exhibited high viability and the formation of clusters of osteoblastic cells surrounded by endothelial cells. The presence of nanoHA caused a dose-dependent increase in the viability/proliferation of osteoblast cells, but opposite results were found in HDMEC. Co-cultures showed a slight increase in the cell viability/proliferation. Particles were readily internalized by both MG-63 cells and HDMEC, but endothelial cells presented the higher percentage of intracellular vesicles loaded with nanoparticles. Co-cultured osteoblastic and endothelial cells exhibited less
iv
internalization of nanoparticles, compared to the respective monocultures. The described effects, caused by the presence of nanoHA particles, were more significant when those were added at the beginning of the culture.
Results suggest that nanoHA particles modulate the behaviour of osteoblastic and endothelial cells and also affects the interaction between these two cell types, representing a promissing biomaterial for new strategies in bone tissue engineering.
v
RESUMO
A nano-hidroxiapatite (nanoHA) tem inúmeras aplicações em estratégias de reparação e regeneração do tecido ósseo. A maioria dos trabalhos publicados, a propósito do perfil biológico da nanoHA, baseia-se em filmes, compósitos, estruturas densas ou porosas, ou cimentos compostos por nanopartículas de HA. No entanto, existe um défice evidente em relação à avaliação biológica de nanopartículas isoladas de HA. Considerando a estreita associação entre os processos de angiogénese e osteogénese nos eventos de formação óssea, este estudo pretende abordar o perfil biológico das nanopartículas de HA em monoculturas e co-culturas de células osteoblásticas e endoteliais.
No presente estudo, as nanopartículas de HA, sintetizadas por método hidrotermal, foram caracterizadas por XRD, análise FTIR e TEM. Os estudos biológicos
in vitro foram realizados utilizando culturas isoladas e co-culturas de células
osteoblásticas humanas MG-63 (103 cell cm-2) e células endoteliais microvasculares dérmicas humanas (HDMEC, 2x104 cell cm-2), em meio de cultura endotelial. As culturas foram mantidas por 14 dias na ausência (controlo) e na presença de nanopartículas em diferentes concentrações. As nanopartículas de HA (1-100 μg ml-1) foram adicionadas às células em cultura em dois momentos diferentes: ao dia 1 e ao dia 7. Para as partículas adicionadas ao dia 1: as culturas foram caracterizadas nos dias 2, 7 e 14; para as partículas adicionadas ao dia 7: as culturas foram avaliadas aos dias 8(7) e 14(7), para os padrões de viabilidade/proliferação celular (MTT) e de crescimento celular (CLSM), assim como para a internalização das partículas e a sua cinética dentro dos compartimentos celulares (TEM).
Os resultados obtidos demonstraram que as monoculturas de células MG-63 e HDMEC, na ausência de nanopartículas, apresentaram alta viabilidade celular e o esperado padrão de crescimento celular. Para além disso, as co-culturas controlo apresentaram alta viabilidade celular e a formação de aglomerados de células osteoblásticas rodeadas por células endoteliais. A presença de nanopartículas de HA promoveu um aumento dose-dependente na viabilidade/proliferação celular dos osteoblastos, no entanto resultados opostos foram encontrados nas culturas de HDMEC. Já nas co-culturas o padrão de viabilidade/proliferação celular apresentou um
vi
ligeiro aumento. As partículas eram facilmente internalizadas por ambos os tipos celulares, contudo as células endoteliais apresentaram uma maior percentagem de vesículas intracelulares carregadas com nanopartículas. As co-culturas de células osteoblásticas e endoteliais apresentaram menor internalização de partículas, em comparação com as respectivas monoculturas. Os efeitos descritos, causados pela presença de nanopartículas de HA, foram mais significativos quando estas eram adicionadas no início da cultura.
Os resultados obtidos sugerem que as nanopartículas de HA têm capacidade de modular o comportamento das células osteoblásticas e endoteliais, afectando também a interacção entre estes dois tipos celulares, apresentando-se como um biomaterial promissor para novas estratégias utilizadas pela engenharia de tecido ósseo.
vii
TABLE OF CONTENTS
Acknowledgments……… i
Abstract……… iii
Resumo………. v
List of Contents……… vii
List of Figures……… ix
List of Tables……….. xii
List of Abbreviations ……… xiii
Chapter I Introduction I.1. Bone biology………. 2
I.1.1. Bone morphogenesis ……… 2
I.1.2. Bone structure……… 3
I.1.2.1. Periosteum and endosteum ……….. 5
I.1.2.2. Bone matrix………. 6
I.1.2.3. Bone cells ………... 8
I.1.3. Bone vascularization ……… 11
I.1.4. Bone tissue metabolic role ……… 12
I.1.5. Bone remodeling……… 14
I.2. Bone repair……….. 15
I.2.1. Spontaneous bone repair………. 15
I.2.2. Strategies for bone repair……… 17
I.2.2.1 Bone Tissue Engineering……….. 18
I.2.2.1.1. Biomaterials……… 19
I.2.2.1.2 Cells……… 22
I.3. Objectives………. 27
Chapter II Material and Methods II.1. Hydroxyapatite nanoparticles………. 30
II.1.1. Hydrothermal synthesis of HA nanoparticles ……… 30
II.1.2. Structural/chemical characterization ……….. 30
II.1.3. Preparation of different concentrations of HA nanoparticles……… 31
II.2. Cell culture……… 32
II.2.1. Monoculture of MG-63 cells ……… 32
II.2.2. Monoculture of Human Dermal Microvascular Endothelial Cells…. 32 II.2.3. Co-culture of MG-63 cells and HDMEC……… 33
II.3. Biological characterization……….. 33
II.3.1. Cell viability/proliferation………. 33
II.3.2. Morphologic evaluation……….. 34
viii
II.5. Statistical analysis……… 36
Chapter III Results III.1. Hydroxyapatite nanoparticles characterization ………. 38
III.2. In vitro biological studies of MG-63 osteoblast-like cells………. 41
III.3. In vitro biological studies of human dermal microvascular endothelial cells………. 46
III.4. In vitro biological studies of MG-63 cells and endothelial cells co-culture…. 50 Chapter IV Discussion and Conclusion IV.1 Discussion………. 57
IV.2 Conclusion………. 62
Chapter V References………..………. 64
ix
LIST OF FIGURES
Figure 1. Overview of a long bone constitution. Compact bone composed by osteons (detailed view) and cancellous bone with trabeculae. Periosteum (inner and outer layer) and endosteum representation……….. 4 Figure 2. The origins and location of bone cells: osteoblasts, osteoclasts,
osteocytes and bone lining cells………. 8
Figure 3. Image showing a osteoclasts digesting bone tissue. Osteoclast with multiple nuclei and a ruffled border in direct contact with the bone matrix. As a result of osteoclasts activity a Howship’s lacunae is formed………. 11 Figure 4. The stages of fracture repair. (a) Hematoma formation: following injury, disruption of blood vessels leads to formation of a hematoma; (b) Soft callus formation: this stage involves the formation of new blood vessels from pre-existing ones (angiogenesis), the external callus and the external callus; (c) Hard callus formation: the callus becomes mineralized; (d) Bone remodeling: the large fracture callus is replaced with secondary lamellar bone, and the vascular supply returns to normal……….
16
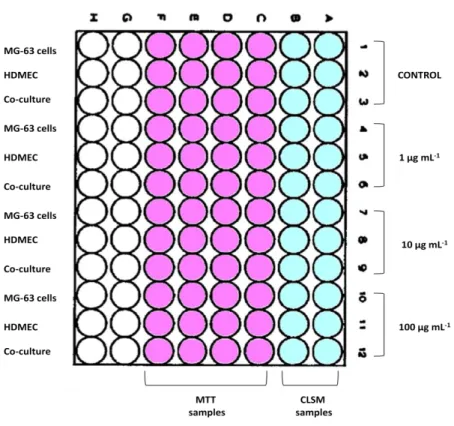
Figure 5. Representative diagram of the cultures organization in a 96-well
plate………... 35
Figure 6. TEM images of the synthesized nanoHA particles: (a) HS particles as prepared; (b) HSster particles. The nanoparticles presented a prismatic rod-like
shape, with a length of ≈ 75 nm and a width of ≈ 25 nm. The values of the scale
bars are (a, b) 100 nm……… 38
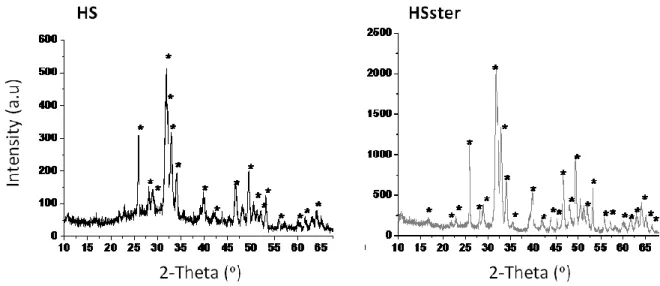
Figure 7. X-ray diffraction patterns of the synthesized nanoHap particles: HS, as prepared and HSster... 39
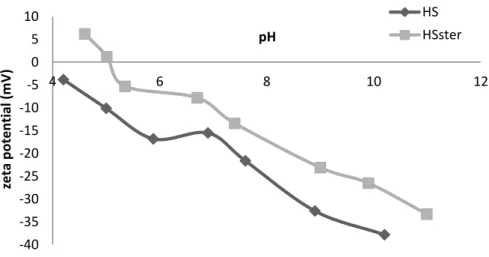
Figure 8. pH dependence of the synthesized particles zeta potential (ξ): HS non and HSster particles... 40
Figure 9. FTIR spectra of the synthesized particles: HS and HSster particles……….
41 Figure 10. Cell viability/proliferation of isolated MG-63 cells cultured for 14 days in the absence (control) and presence of nanoHA particles (1 μg mL-1 – 100 μg mL
-1
) estimated by MTT assay. (A) Data referring to the addition of nanoHA at day 1; culture evaluation at days 2, 7 and 14. (B) Data referring to the addition of nanoHA at day 7; culture evaluation at days 2, 8 and 14……….. 42 Figure 11. CLSM images of MG-63 cells monoculture. Cultures stained for F-actin (green) and nucleus (red); 200x. (A) MG-63 cells cultured for 2 days in the absence of nanoHA (control). (B) Images: a-d represent MG-63 cell cultures performed for 7 days in the absence (a) and presence of 1, 10 and 100 μg mL-1
x
nanoHA (b-d, respectively) added at day 1; e-h represent MG-63 cell cultures performed for 14 days in the absence (e) and presence of 1, 10 and 100 μg mL-1 nanoHA (f-h, respectively) added at day 1; i-k represent MG-63 cell cultures performed for 14 days in the presence of 1, 10 and 100 μg mL-1 nanoHA respectively, added at day 7. The value of the scale bars is: 100 μm……… 43 Figure 12. Representative TEM images of MG-63 cell cultures exposed to 10 μg mL-1 nanoHA particles. Cultures maintained for 2 (a), 7 (b) and 14 (c and d) days. Images: (a) emission of pseudopods surrounding the nanoparticles; (a, b and d) clusters of nanoparticles entrapped in intracellular vesicles; (c) apoptotic cell with rounded shape, condensed chromatin at nuclear periphery and cytoplasmic vacuolization. The values of the scale bars are: (a,b and d) 1 μm; (c) 2 μm………. 45 Figure 13. Cell viability/proliferation of isolated HDMEC cultured for 14 days in the absence (control) and presence of nanoHA particles (1 μg mL-1 – 100 μg mL-1) estimated by MTT assay. (A) Data referring to the addition of nanoHA at day 1; culture evaluation at days 2, 7 and 14. (B) Data referring to the addition of nanoHA at day 7; culture evaluation at days 2, 8 and 14……….. 46 Figure 14. CLSM images of HDMEC monocultures. Cultures stained for CD31 (green) and nucleus (red), 200x. Images: a-d represent HDMEC cultures performed for 7 days in the absence (a) and presence of 1, 10 and 100 μg mL-1 nanoHA (b-d, respectively), added at day 1; e-h represent HDMEC cultures performed for 14 days in the absence (e) and presence of 1, 10 and 100 μg mL-1 nanoHA (f-h, respectively), added at day 1; i-k represent HDMEC cultures performed for 14 days in the presence of 1, 10 and 100 μg mL-1 nanoHA respectively, added at day 7. The values of the scale bars are: (a-k) 100 μm…………. 48
Figure 15. Representative TEM images of HDMEC cultures exposed to 10 μg mL-1
nanoHA particles. Cultures maintained for 2 (a and d) and 8 (b and c) days. Images: (a) emission of pseudopods surrounding the nanoparticles; (b, c and d) clusters of nanoparticles entrapped in intracellular vesicles. The values of the
scale bars are: (a-d) 1 μm……… 49
Figure 16. Cell viability/proliferation of MG-63 cells and HDMEC co-cultured for 14 days in the absence (control) and presence of nanoHA particles (1 μg mL-1 – 100 μg mL-1) estimated by MTT assay. (A) Data referring to the addition of nanoHA at day 1; culture evaluation at days 2, 7 and 14. (B) Data referring to the addition of nanoHA at day 7; culture evaluation at days 2, 8 and 14……….. 51 Figure 17. CLSM images of MG-63 cells and HDMEC co-cultures. Cultures stained for CD31 (green) and nucleus (red), 100x. Images: a-d represent co-cultures performed for 7 days in the absence (a) and presence of presence of 1, 10 and 100 μg mL-1 nanoHA (b-d, respectively), added at day 1; e-h represent co-cultures performed for 14 days in the absence (e) and presence of 1, 10 and 100 μg mL-1 nanoHA (f-h, respectively), added at day 1; i-k represent co-cultures performed for 14 days in the presence of 1, 10 and 100 μg mL-1 nanoHA, respectively, added
xi
at day 7. The values of the scale bars are: (a-k) 250 μm……… 52 Figure 18. Representative TEM images of MG-63 cells and HDMEC co-cultures exposed to 10 μg mL-1 nanoHA particles. Cultures maintained for 2 (a and c) and 7 (b and d) days. Images: (a) emission of pseudopods surrounding the nanoparticles; (a,b and d) clusters of nanoparticles entrapped in intracellular vesicles; (c) cellular debris in the extracellular space, resulting from the degeneration of an apoptotic cell. The values of the scale bars are: (a and b) 1
μm; (c) 2 μm; (d) 3 μm……… 54
Figure 19. Representative TEM images of MG-63 cells and HDMEC monocultures and co-cultures exposed to 10 μg mL-1 nanoHA particles. Cultures maintained for 2 days, show the differences in the amount of vesicles loaded with nanoparticles between the co-culture system (a), the MG-63 cells monoculture (b) and the HDMEC monoculture (c). The values of the scale bars are: (a-c) 1 μm……… 55
xii
LIST OF TABLES
Table 1. Bone matrix main non-collagenous proteins……… 7
Table 2. Studies that used nanoHA, in vitro and in vivo, for BTE………. 21
Table 3. Different cell types used in BTE and their characteristics……… 23
Table 4: Properties of nanoHA particles obtained by hydrothermal synthesis: HS
and HSster………. 38
Table 5. Semi-quantitative evaluation of the process of 10 μg mL-1 nanoHA internalization and kinetics within MG-63 cells……….. 45 Table 6. Semi-quantitative evaluation of the process of 10 μg mL-1 nanoHA internalization and kinetics within HDMEC……… 50 Table 7. Semi-quantitative evaluation of the process of 10 μg mL-1 nanoHA internalization and kinetics within co-cultures……… 54
xiii
LIST OF ABBREVIATIONS
3D three Dimensional
ALP Alkaline Phosphatase
Ang-1 Angiopoietin 1
ATCC American Type Culture Collection
BET multipoint Brunauer-Emmett-Teller isotherm
bFGF basic Fibroblast Growth Factor
BMP Bone Morphogenic Protein
BMUs Basic Multicellular Units
BSA Bovine Serum Albumin
BSP Bone Sialoprotein
BTE Bone Tissue Engineering
cAMP cyclic Adenosine Monophosphate
cGMP cyclic Guanosine Monophosphate
CLSM Confocal Laser Scanning Microscopy
COL I Type I Collagen
DPBS Dulbecco’s Phosphate Buffered Saline
DMSO Dimethylsulfoxide
ECGS Endothelial Cell Growth Suplement
ECM Endothelial Culture basal Medium
EDTA Ethylenediamine tetraacetic acid
ESC Embrionic Stem Cell
FBS Foetal Bovine Serum
FTIR Fourier Transform Infrared Spectroscopy
HA Hydroxyapatite
HDMEC Human Dermal Microvascular Endothelial Cell
HIF Hypoxia-inducible Transcription Factor
HOBs Human Osteoblasts
HS Hydrothermal Synthesis
IGF Insuline-like Growth Factor
M-CSF Macrophage Colony Stimulating Factor
MMPs Metalloproteinases
MSC Mesenchymal Stem Cell
MTT
xiv
OC Osteocalcin
ON Osteonectin
OPG Osteoprotegerin
OPN Osteopontin
PECAM-1 Platelet-Endothelial Cell Adhesion Molecule
PBS Phosphate Buffered Saline
PDS Polydioxanone
PGA Polyglycolic acid
PLA Polylactide acid
PLC Poly(ε-caprolaction)
PTH Parathyroid Hormone
RANK Receptor Activator of Nuclear Factor κ B
RANKL RANK Ligant
RNA Ribonucleic Acid
SPSS Statistical Package for the Social Sciences
SSA Specific Surface Area
TEM Transmission Electron Microscopy
TGF-β Transforming Growth Factor beta
VEGF Vascular Endothelial Growth Factor
XRD X-ray diffraction
α-MEM Minimum Essential Medium, alpha modification
CHAPTER I
1
I.1. Bone biology
Bone is a dynamic connective tissue that performs several key functions within higher vertebrates. It is the material from which the skeleton is made, providing support and protection to the internal organs (Saladin, 2004). Besides, it provides attachment for muscles, facilitating the locomotion process; it is involved in the synthesis of blood cells and in the metabolism of mineral ions such as calcium and phosphorous (Morgan and Bouxsein, 2008). Throughout life, bone is continually changing to adapt its shape and structure to perform different tasks. This process of renewal happens by laying down new bone tissue and resorbing old (Einhorn, 1996; Hing, 2004).
During this work, we will review the main aspects of bone biology, such as, bone morphogenesis and structure, bone vascularization, bone metabolic role and finally bone remodeling and repair.
I.1.1. Bone morphogenesis
Osteogenesis is the process through which mature bone is formed. Accordingly, the ossification process occurs by two different methods: intramembranous ossification and endochondral ossification (Seeley et al., 2004).
The formation of flat bones like the vault of the skull, the pelvis, and the majority of face bones occurs by intramembranous ossification. Thus, during embryonic development, mesenchymal cells condensate and differentiate into osteoprogenitor cells, and then into osteoblasts, which line up around blood vessels (Marks and Hermey, 1996). At this point, all the necessary conditions to start to produce and deposit new bone matrix are present (Buckwalter et al., 1996a). The first structure to be formed is a network of trabeculae composing the cancellous bone, which undergoes continuous mineralization. When the osteoblasts and osteoclasts coupled actions begin, the trabeculae are remodeled. At the end of this process, the
2
newly formed bones present a typical arrangement: the cancellous bone is flanked by compact bone (Saladin, 2004; Seeley et al., 2004).
Endochondral ossification occurs in long bones, like the bones of the vertebral column, the base of the skull, part of the mandible and in the appendicular skeleton, formed from a cartilage model (Meghji, 1992; Buckwalter et al., 1996a). Thus, embryonic mesenchymal cells differentiate into chondrocytes, which proliferate and produce a collagenous matrix, leading to the formation of a hyaline cartilage model (Ortega et al., 2004).
In the interior of the cartilage model, chondrocytes undergo a progressive hypertrophy and the cartilage matrix becomes mineralized, leaving behind interconnected spaces (Marks and Hermey, 1996). Simultaneously, at the surface of the developing endochondral bone, the perichondrium is responsible for laying down a thin osteoblastic layer, surrounding the diaphysis, named periosteal collar (Marks and Hermey, 1996).
From this moment on, primary ossification takes place. Therefore, blood vessels invade the central region of the mineralized and hypertrophied cartilage, taking with it osteoprogenitor cells and osteoblastic precursors, responsible for osteoblastic differentiation, and therefore bone formation (Marks and Hermey, 1996; Scotti et al., 2010). At the same time, osteoclastic invading cells resorb the central part of the cartilage leading to the formation of a marrow cavity (Buckwalter et al., 1996a).
The epiphysis of a long bone and, the extremities of irregular and flat bones, are formed by secondary ossification. Separating the epiphysis from the diaphysis is a newly formed structure named epiphysial or growth plate. This specific area is responsible for chondrocytes segregation, proliferation and hypertrophy, resulting in longitudinal bone growth, until skeletal maturity, when the diaphysis and epiphysis fuse with each other (Marks and Hermey, 1996; Ortega et al., 2004).
A proper vascular network is also important to ensure the transportation of oxygen, nutrients, several cells and soluble factors, necessary to the skeletal homeostasis. The modification of this homeostasis leads to several bone pathologies,
3
such as osteomyelitis, osteonecrosis and osteoporosis, caused by abnormal blood supply (Carmeliet, 2003).
Angiogenesis is a biological process through which new vessels sprout from pre-existing ones, bridge and growth until a new functional structure is formed (Vander et
al., 2001a). During bone formation, the angiogenic process is important to the
intramembranous and endochondral ossification. Thus, the vascular invasion into the mesenchymal zone during intramembranous ossification and the vascular supply to the growth plate during endochondral ossification, are essential to a complete process (Saladin, 2004). Moreover, during fracture repair neoangiogenesis occurs until vascular supply returns to normal (Carano and Filvaroff, 2003).
The normal angiogenic process is regulated by several factors, like vascular endothelial growth factor (VEGF); basic fibroblast growth factor (bFGF); various members of the transforming growth factor beta (TGF-β) family; hypoxia-inducible transcription factor (HIF) and angiopoietins 1 (Ang-1) (Kanczler and Oreffo, 2008). The VEGFs are important growth factors since they are involved in the regulation of vasculogenesis and angiogenesis, and also play an important role in bone growth and repair (Carano and Filvaroff, 2003). The main effects of VEGFs are on endothelial cells, but the VEGF receptors are also expressed on monocytes, chondrocytes and osteoblasts. Moreover, besides the angiogenic effects, VEGFs can induce haematopoietic stem cell recruitment from bone marrow, monocyte chemoattraction, neural protection and bone formation (Ferrara and Davis-Smyth, 1997).
I.1.2.
Bone structure
The overall structure of the bone is composed of two different types of organization: compact or cortical bone and, cancellous or trabecular bone. While the first type is relatively dense and is found at the bone surface, functioning as a protection in mechanically demanding regions, cancellous bone consists in a network of trabeculae surrounding interconnected spaces and it is found in the interior of bones (Fig. 1) (Tartaglia and Waugh, 2002a).
4
Compact bone represents 80% of the mature skeleton, distributed by the long bones diaphysis and surrounding the majority of bones (Pratt, 1991). It is constituted by quasi-cilindrical elements called osteons or Harvesian systems, which are formed by densely packed collagen fibrils called lamellae (Rossert and Crombrugghe, 1996). Each ofthese structures presents a central lumen containing a blood vessel responsible for the vascular supply along the axis of a long bone. Moreover, compact bone also has transversal blood vessels, perpendicular to the axis of a long bone, located in Volkmann canals, as well as capillaries and venules from microcirculation and, occasionally, a nerve fiber (Seeley et al., 2004).
Figure 1. Overview of a long bone constitution. Compact bone composed by osteons (detailed view) and cancellous bone with trabeculae. Periosteum (inner and outer layer) and endosteum representation (Adapted from Seeley et al., 2004).
Cancellous bone is also called spongy bone, because of the porous appearance created by the network of trabeculae. It exists within the cortical bone, in the central region of flat and irregular bones such as vertebrae, skull and pelvic bones but also in the long bones mataphysis and epiphysis (Pratt, 1991). Despite its porous appearance, cancellous bone has a structural support function, since its trabeculae are oriented by stress and tension lines in the interior of the bone. As well as compact bone, cancellous bone matrix is organized in lamellae, without a complete osteon
5
organization (Saladin, 2004). Furthermore, this type of bone filled by blood vessels, bone marrow cells and adipocytes is related with cellular functions presenting a high metabolic rate (Buckwalter et al., 1996b).
Whether it is compact or cancellous, bone has layers of connective tissue and bone forming cells covering its external and internal surfaces, namely the periosteum and the endosteum. Furthermore, as a connective tissue, bone is constituted by the extracellular matrix, with organic and inorganic components, functioning as a scaffold for the different types of bone cells (Seeley et al., 2004).
I.1.2.1. Periosteum and endosteum
Periosteum is the external coating of the majority of bone surfaces, present in locations of high risk of fracture and, absent from tendon, articular surfaces and, ligament insertions (Allen et al., 2004). The periosteum is attached to the bone by collagen fibers, also called Sharpey’s fibers oriented by tension forces that penetrate into the bone promoting a tight junction between bone tissue and the periosteum (Saladin, 2004; Junqueira and Carneiro, 2005).
Periosteum is described as a fibro-vascular membrane that is composed by an external layer, named fibrous layer and, an inner layer called osteogenic layer (Fig. 1) (Allen et al., 2004). The fibrous layer of the periosteum is composed by numerous fibroblasts with collagen and elastin fibers, associated in a microvascular network. The inner osteogenic layer is formed by osteogenic progenitor cells which differentiate into osteoblasts. This part of the periosteum is responsible for the diaphysis diameter increase and for fracture regeneration, since cellular growth occurs by layers (Dwek, 2010).
Endosteum is a single-celled membrane formed by osteoprogenitor cells and hematopoietic stem cells (Balduino et al., 2005). It is the inner coating of all bone cavities, such as, the medullary cavity of long bones diaphysis and the trabecullae in cancellous bone (Fig. 1) (Seeley et al., 2004; Mescher, 2010).
6
These two types of connective tissue, intimately connected with bone, are directly involved in bone growth and regeneration during fracture repair, acting as a continuous source of bone cells (Junqueira and Carneiro, 2005).
I.1.2.2. Bone matrix
The bone extracellular matrix has two main components: the organic matrix and the inorganic matrix. The organic matrix, composed by collagen and non-collagenous proteins, accounts for 35% of the total weight of bone tissue compared with 65% for the inorganic components (Ca2+ and PO43-) (Buckwalter et al., 1996b).
The organic extracellular matrix major component is collagen type I (COL I), a bone structural protein responsible for bone’s wide range of mechanical properties (Rossert and Crombrugghe, 1996).
Composing one collagen molecule structure are three polypeptide chains organized in a triple helix. Each chain is made by a sequence of aminoacids with high content in glycin, prolin and hydroxyproline. COL I is composed of two α1 chains and one α2 chain, being each type of chain coded by two different genes (Van Der Rest and Garrone, 1991; Rossert and Crombrugghe, 1996; Cowles et al., 1998).
Procollagen is the synthesized form of collagen, formed by triple helix α chains, which will have their N- and C-propeptides removed by specific proteases. These changes promote the molecules assembling into a fibril form, displayed in a overlapping manner with spaces between adjacent fibrils. The intermolecular crosslinks formed have a very high tensile strength functioning as a scaffold for bone growth (Eyer, 1996; Rossert and Crombrugghe, 1996).The fibrillar collagens group is composed by: type I, type II, type III, type V, type XI, type XXIV and type XXVII collagen forms (Van Der Rest and Garrone, 1991; Bou-Gharios and Crombrugghe, 2008).
The remainder part of the organic matrix is composed by non-collagenous proteins such as glycoproteins, proteoglycans and several other exogenous proteins with important biological activity in the bone (Cowles et al., 1998). Table 1 gives an overview of bone matrix main non-collagenous components, as well as their function in bone tissue.
7
Table 1: Bone matrix main non-collagenous proteins.
Bone matrix
non-collagenous proteins Function References
Glycoproteins
Alkaline phosphatase (ALP)
Mediates mineralization.
May be involved in phosphate transport. Sugawara et al., 2002. Whyte, 2008. Gade et al., 2011. Osteonectin (ON)
Positive regulator of bone
formation and remodeling.
Related with hydroxyapatite
deposition. Boskey et al., 2003. Delaney and Hankenson, 2009. Osteopontin (OPN/ BSP-I)
May promote osteoblasts and osteoclasts attachment to the bone matrix. Standal et al., 2004. Noda and Denhardt, 2008. Bone sialoprotein (BSP-II) Initiates mineralization. Controls osteoblast
differentiation and activity.
Malaval et al., 2008.
Robey, 2008. Osteocalcin
(OC)
Associated with bone turnover.
Regulator of osteoblastic function. Ducy and Karsenty, 1996. Lee et al., 2000. Proteoglycans Decorin and Biglycan
Most abundant proteoglycans in bone matrix.
Related with osteoblast
differentiation and consequent matrix mineralization.
Modulate collagen fibrillogenesis and organization. Parisuthiman et al., 2005. Iozzo, 1998. Mochida et al., 2009. Lamoureux et al., 2007. Other proteins Transforming Growth Factors β (TGF-β1; -β2; -β3) Promote osteoblastic
differentiation, migration and proliferation.
Stimulate COL I production by osteoblasts.
Inhibit collagenase synthesis.
Janssens et al., 2005. Leask and Abraham, 2004. Feres-Filho et al., 1995. Insulin-like Growth Factors (IGF I, IGF II)
Stimulate osteoblastic
proliferation.
Stimulate COL I synthesis.
Maintain collagen integrity by decreasing collagenase synthesis.
Devescovi et al., 2008. D’Avis et al., 1997. Salgado et al., 2004a. Bone Morphogenetic Proteins 2/7 (BMPs-2/7) Involved in osteoblastic differentiation.
Induce ectopic cartilage and bone formation.
Key role during bone and cartilage repair.
Xiao et al., 2007. Yu et al., 2010.
Badlani et al.,
8
The inorganic phase of the bone is mainly composed by calcium and phosphorous ions, organized in a calcium phosphate compound, named hydroxyapatite (HA, Ca10(PO4)6(OH)2) (Marks and Hermey, 1996). These mineral
crystals, with plate-like shape, are deposited by cellular control into the extracellular matrix, between adjacent collagen fibrils, originating the strength and stiffness of bone (Olszta et al., 2007; Orimo, 2010). In addition to calcium and phosphorous there are other ions, found in minor percentage in bone, such as bicarbonate, potassium, sodium, magnesium and citrate (Buckwalter et al.,1996b).
I.1.2.3. Bone cells
Throughout life, bone cells play an important role in bone maintenance, through their capability to synthesize and resorb the extracellular matrix. The cells responsible for these events are: osteoblasts, osteocytes, bone linning cells and osteoclasts (Fig. 2).
Figure 2. The origins and location of bone cells: osteoblasts, osteoclasts, osteocytes and bone lining cells (Adapted from Marks and Hermey, 1996).
Bone cells have different origins (Fig. 2). Osteocytes, osteoblasts and bone linning cells are derived from undifferentiated pluripotent mesenchymal cells, located in bone marrow, endosteum and periosteum (Buckwalter et al., 1996b). Proliferation and differentiation of mesenchymal cells are under influence of hormones and growth factors, determining the type of cells formed during differentiation. Thus,
9
mesenchymal cells can differentiate into osteoblasts, chondroblasts, adipocytes, myoblasts and fibroblasts (Triffitt, 1996). Osteoclasts belong to the monocyte-macrophage lineage, being the final product of hemopoietic lineage (Suda et al., 1996).
Located at the formation and remodeling zones of bone, are osteoblasts – bone forming cells – responsible for synthesize and secret unmineralized bone matrix, called osteoid (Cowin and Doty, 2007). Moreover, osteoblasts are also involved in the mineralization process of bone matrix (Aubin and Lui, 1996; Orimo, 2010).
Active osteoblasts are mononucleated, cuboidal cells tightly packed along bone surface, with a prominent Golgi apparatus, well-developed rough endoplasmatic reticulum, and large quantities of mitochondria (Marks and Hermey, 1996; Junqueira and Carneiro, 2005). This type of bone cell is responsible for the synthesis of bone matrix proteins, such as COL I and noncollagenous proteins, that are stored in vesicles and then released to the extracellular space, by exocytosis. Additionally, bone matrix become calcified when osteoblasts secrete vesicles filled with calcium ions (Ca2+) and phosphate ions (PO42-), promoting HA crystals formation and deposition throughout
the osteoid (Gay et al., 2000; Seeley, 2004; Orimo, 2010). By the time that the matrix synthesis process is over, inactive osteoblasts flatten, acting as a protective layer of the newly formed bone (Junqueira and Carneiro, 2005).
During bone formation, certain osteoblasts become surrounded by bone matrix and differentiate into osteocytes (Cowin and Doty, 2007). Thus, osteocytes are smaller than osteoblasts presenting dendritic shape, oval nucleus and few cytoplasm, less prominent endoplasmatic reticulum and smaller Golgi apparatus (Marks and Hermey, 1996). Mature osteocyte occupies the matrix lacunae, and communicates with neighboring osteocytes, as well as, bone surface and blood vessels, through filipodia connections, ensuring the access to oxygen and nutrients (Nijweide et al., 1996). Besides, osteocytes function as strain detectors and consequently coordinate osteoblasts and osteoclasts activity, when bone remodeling is needed (Saladin, 2004).
Bone lining cells are inactive osteoblasts, located at bone surface, at zones where bone is not under formation or resorption (Marks and Hermey, 1996). These cells are flat and elongated, presenting a spindle-shaped nucleus, and fewer
10
cytoplasmatic organelles (Cowin and Doty, 2007). Additionally, Everts and colleagues have proposed that bone lining cells in vitro are responsible for: 1) “cleaning” bone surface from nonmineralized collagen fibrils before osteoclastic attachment; 2) for digesting the collagen left by osteoclasts after resorption ; 3) form a cement line and deposit a thin layer of fibrillar collagen on the cleaned surfaces (Everts et al., 2002).
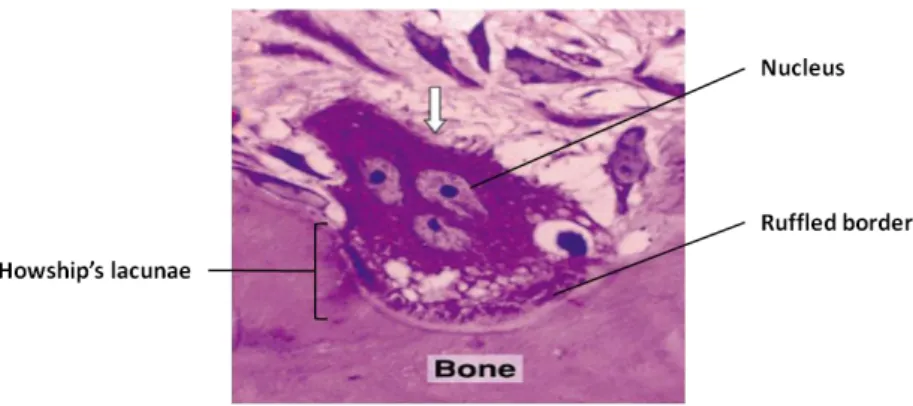
The fourth cellular component of bone is the osteoclasts (Fig. 3), a large (20-100 µm), multinucleated (2 to 50 nucleus) cell. The main function of this type of cell is to resorb bone, dentin and calcified cartilage (Marks and Hermey, 1996).
Osteoclasts have their polarity changed during the resorption cycle. Thus resorbing osteoclast is highly polarized, when compared with nonresorbing osteoclast (Väänänen, 1996). When resorption sites are determined, osteoclasts attach to bone surface, through a specific membrane domain named sealing zone, which delimits other membrane domain - the ruffled border. This specific zone, with its finger-like projections, is where the bone matrix demineralization and degradation process take place (Saladin, 2004; Mescher, 2010). The ruffled border creates an acidic environment between the osteoclasts and the matrix causing HA crystals dissolution, consequently forming resorption lacunae, named Howship’s lacunae (Väänänen and Zhao, 2008). After matrix demineralization, degradation of collagenous matrix happens with the ruffled border secreting several proteinases, such as metalloproteinases (MMPs) and lysossomal cysteine proteinases (Kusano et al., 1998; Delaisse et al., 2000).Additionally to those mentioned membrane domains, osteoclasts have a homogeneous membrane, known as basolateral membrane, and a secretory domain that function as a site for the resorbed products exocytose (Väänänen et al., 2000).
11
Figure 3. Image showing an osteoclast digesting bone tissue. Osteoclasts with multiple nuclei and a ruffled border in direct contact with the bone matrix. As a result of osteoclasts activity a Howship’s lacunae is formed (Adapted from Junqueira and Carneiro, 2005).
I.1.3. Bone vascularization
The presence of blood vessels in bone tissue is very important in cases of bone growth, bone healing, fracture repair and osteogenesis. A proper blood supply is necessary to maintain bone vitality, and to sustain bone and bone marrow cells function, through oxygen and nutrients input and metabolic waste products output (Shim, 1968).
Blood supply in long bones is made by several arteries, which penetrate the bone cortexes in different locations: in the diaphysis, in the metaphysis and in the epiphysis. Hence, there are the nutrient artery, the metaphysial artery, the epiphysial artery, and periosteal vessels that carry arterial blood to the bone. This group of arteries is interconnected by longitudinal anastomoses creating a network around the diaphysis and metaphysis (Tartaglia and Waugh, 2002b).
Bone supply is made by centrifugal flow, meaning that arterial blood goes first to the medullary cavities and then flows to the outer cortex through the arteries that arise inside the Haver’s and Volkmann’s canals. This group of vessels is responsible for blood supply to the inner 2/3 of the cortex, while the periosteal vessels supply the outer third (Al-Motabagani, 2002; Travlos, 2006).
12
Along the diaphysis axis, there is a sinus that drains into veins: metaphysial vein, epiphysial vein and centromedullary veins. This group of veins runs in parallel with the arteries, and carries venous blood out of the bone (Laroche, 2002; Tartaglia and Waugh, 2002b).
I.1.4. Bone tissue metabolic role
Bone tissue contains 99% of total body calcium and, some of the most important functions of this tissue are the storage and regulation of its homeostasis (Buckwalter et al., 1996b; Marks and Hermey, 1996; Cowin and Doty, 2007). The balance of calcium in the body, through calcium diffusion in and out of the bone, is regulated by numerous factors. Local factors including cytokines, growth factors and prostaglandins, and calcitropic hormones, such as, parathyroid, vitamin D and calcitonin, are involved in bone-cell functions (Marks and Hermey, 1996; Buckwalter et
al., 1996a). However, calcium homeostasis depends not only on the bone and on the
referred factors, but also on kidneys and the gastrointestinal track (Vander et al., 2001b). Accordingly, bone mineralization occurs only when calcium and phosphorous levels in the blood are sufficient for bone normal deposition (Saladin, 2004).
Parathyroid hormone (PTH) has direct effect in bone tissue, since it stimulates osteoclastic function and therefore, the release of calcium from bone to the bloodstream (Buckwalter et al., 1996a; Fitzpatrick and Bilezikian, 1999). In addition, PTH controls calcium resorption by the kidneys and promotes vitamin D synthesis (Buckwalter et al., 1996a).
Vitamin D final form (1,25-didydroxyvitamin D3), is synthesized in the kidneys
depending on the levels of serum calcium (Buckwalter et al., 1996a; Rossert and Crombrugghe, 1996). Despite having several actions in different cells and tissues, vitamin D is primarily associated with bone function and mineral homeostasis (Fitzpatrick and Bilezikian, 1999). It has an inductive effect in bone resorption, by promoting the differentiation of osteoclastic precursors into active osteoclasts (Suda et
13
plays an important role in enhancing calcium absorption in the intestine (Fitzpatrick and Bilezikian, 1999; Goltzman et al., 2004).
Calcitonin, produced and secreted by the parafollicular cells of thyroid gland, is responsible for the decrease of calcium serum levels, since it has an inhibitory effect on osteoclasts (Chambers, 1991; Buckwalter et al., 1996a). The regulatory function of calcitonin is amplified during ‘’calcium stress’’ periods, such as, growth, pregnancy, and lactation, therefore protecting the skeleton from excessive resorption (Vander et al., 2001b). These important features have been taken into account in several studies, where the therapeutic utility of calcitonin in the treatment of metabolic bone diseases, like Paget disease, hypercalcemia and osteoporosis, is investigated (Buckwalter et al., 1996a; Vander et al., 2001b).
Bone cell function can be influenced by other hormones, such as, glucocorticoids, thyroid hormone and estrogens. An important function of glucocorticoids in bone metabolism is its inhibitory effect on bone matrix production by the osteoblasts. Thyroid hormones, thyroxine and tri-iodothyronine, enhance bone osteoclastic resorption. Finally, estrogens have a major role in bone maintenance since inhibit osteoclastic function. Thus, clinically, it is known that bone loss in postmenopausal females is associated with decreased estrogen levels (Buckwalter et
al., 1996a; Suda et al., 1996).
Local factors, including cytokines, growth factors and prostaglandins affect cell-to-cell and cell-to-extracellular matrix interactions. Cytokines and growth factors mediate cell-to-cell interaction within bone, regulating several features of bone cells during skeletal development and in the remodeling process of the mature skeleton. Prostaglandins regulate a diverse group of biological processes, including inflammation, blood flow and, ion transport across cell membranes. Additionally, prostaglandins promote formation and proliferation of osteoclasts and, subsequently have a positive regulation on bone resorption (Rossert and Crombrugghe, 1996; Buckwalter et al., 1996a; Lorenzo and Raisz, 1999).
14
I.1.5. Bone remodeling
Bone is a specialized tissue responsible for body protection and locomotion. To play these functions, bone has to be capable to respond and adapt to several mechanical stresses (Einhorn, 1996; Hill, 1998).
Bone remodeling is a process of continuous bone shaping through replacement of old bone by new tissue, involving the coupled actions of bone resorbing and bone forming cells (Buckwalter et al., 1996a; Lemaire et al., 2004). This adaptive phenomenon also occurs to repair microdamage caused by repeated physical stresses and loads (Lemaire et al., 2004). Therefore, during bone remodeling, differentiated osteoclasts are induced to resorb bone on a particular surface, being replaced by osteoblasts responsible for bone formation. This process is played by an association of bone cells and bone extracellular matrix, designated basic multicellular units (BMUs), which act geographically and chronologically separated from other BMUs keeping bone morphology unchanged (Hill, 1998; Raggatt and Partridge, 2010).
The triggering signal that attracts osteoclastic precursors to the microdamage site, to start the remodeling process, is not completely understood, but seems to be associated with osteocyte signalization through their canaliculi network along the bone after a structural damage or with bone cell response to an unusual hormonal level (Hill, 1998; Seeman, 2008). When mature osteoclasts are at the resorption site, osteoclastogenesis begin with the renewal of the damage, creating a resorption bay in the bone tissue. After the maximum resorption pick, osteoclasts undergo apoptosis and osteoblastic precursors are attracted to the lacunae by TGF-β and COL I released from the digested matrix. Then the osteoblastic precursors differentiate into osteoblasts that start bone formation until the lacunae is totally filled (Hill, 1998; Raggatt and Partridge, 2010).
Moreover, the two bone remodeling events – osteoblastogenesis and osteoclastogenesis – are dependent on several mechanisms involving cytokines, growth factors, hormones and vitamins. A group of different cytokines namely macrophage colony stimulating factors (M-CSF) and receptor activator of nuclear
15
factor κ B ligand (RANKL), are directly related with osteoclasts differentiation, survival and expansion and, thus, with bone resorption. M-CSF and RANKL are both expressed by osteoblasts, and have their receptors on the surface of osteoclastic precursor cells, namely c-fms and receptor activator of nuclear factor κ B (RANK). The binding of these cytokines to its receptors is the signalizing mechanism necessary to osteoclasts activation and differentiation (Udagawa et al., 2000; Proff and Römer, 2009). Additionally, osteoblasts also express a soluble decoy receptor for RANKL named osteoprotegerin (OPG), which prevents RANK – RANKL connection and inhibits osteoclasts differentiation, blocking bone resorption. The OPG – RANKL – RANK balance is so important to bone homeostasis, that several alterations on this system are related to human metabolic bone diseases (Yavropoulou and Yovos, 2008; Kohli and Kohli, 2011).
I.2. Bone repair
When exposed to an injury, usually a fracture, bone tissue reacts spontaneously by remodeling itself, through several biological events, until the normal function and morphology are completely restored. However, there are special cases, such as large fractures, bone defects, or even pathological conditions, where this natural faculty is not enough to regenerate the injured tissue, and so several alternative strategies should be taken in order to repair bone tissue (Kumar et al., 2007a; Dimitriou et al., 2011).
I.2.1. Spontaneous bone repair
As a living tissue, bone has the ability to regenerate in response to fractures, although it is unique in its scarless response (Carano and Filvaroff, 2003). The repair process involves several cell types, including macrophages, polymorphonuclear leukocytes, fibroblasts, endothelial cells and osteoblasts (Marsh and Li, 1999).
16
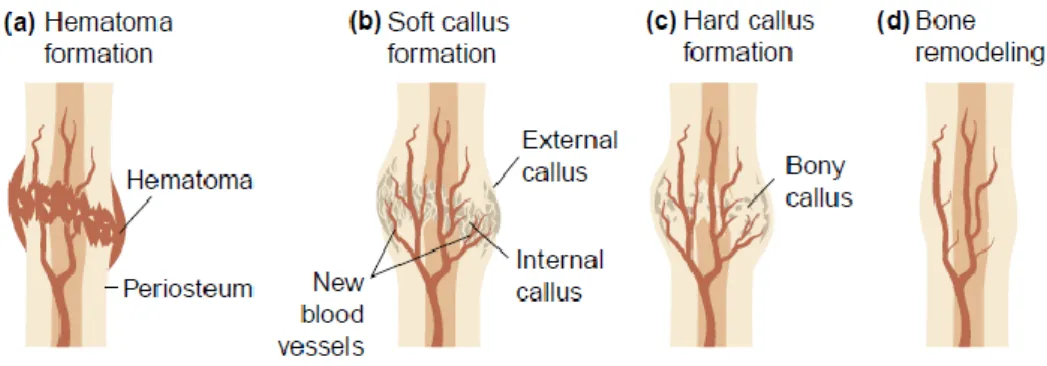
In response to fracture, there is a cell and tissue reaction, namely an inflammation phase preceding proliferative and remodeling phases (Carano and Filvaroff, 2003). When bone tissue is fractured, there is a bleeding and initially a blood clot is formed. Blood vessels permeability and volume change as a response to inflammation and as a consequence of vasodilatation several proteins, growth factors and cytokines arrive to the fracture site triggering the influx of inflammatory cells. At the fracture gap, bone cells and bone matrix are phagocyted and substituted by granulation tissue. This provisory tissue is continuously replaced by fibro-cartilage leading to a soft callus formation around the fracture site. Subsequently, the periosteum osteogenic potential allows new bone cell formation and, as a result, bone tissue is formed in the internal and external layer of the callus, creating a hard callus (Marsh and Li, 1999; Seeley, 2004). The remodeling phase of bone regeneration is characterized by fracture callus replacement by stronger lamellar bone, until normal bone morphology and total blood supply are restored (Fig. 4) (Marsh and Li, 1999; Carano and Filvaroff, 2003).
However, in cases of large bone fractures or defects, in pathological conditions (e.g. tumors, bacterial infections, osteonecrosis), in age related disorders like osteoporosis, and in other cases of large bone loss, bone tissue is not capable to restore its shape and function spontaneously (Seeley et al., 2004; Kumar et al., 2007a). To restore bone health, shape and function, surgical reconstruction is needed. Therefore, the proper strategy and bone substitute must be chosen.
Figure 4. The stages of fracture repair. (a) Hematoma formation: following injury, disruption of blood vessels leads to formation of a hematoma; (b) Soft callus formation: this stage involves the formation of new blood vessels from pre-existing ones (angiogenesis), the external callus and the internal callus; (c)
17
Hard callus formation: the callus becomes mineralized; (d) Bone remodeling: the large fracture callus is replaced with secondary lamellar bone, and the vascular supply returns to normal (Adapted from Carano and Filvaroff, 2003).
I.2.2. Strategies for bone repair
Bone grafting is the current treatment used to enhance bone repair process, in cases that bone cannot do it spontaneously. Thus, it can be made by replacing missing bone for natural material, either from the patient’s own skeletal (autografting) or that of a donor (allografting), or for man-made synthetic materials (Finkemeier, 2002; Salgado et al., 2004a).
To restore bone architecture and functionality two different strategies can be taken: 1) transplantation or 2) implantation (Hench, 1998).
Bone transplantation is the surgical procedure through which healthy and functional bone tissue is harvested to fill the skeletal defect. While bone autografting is the process used when bone tissue that fills the defect is from the patient’s own skeletal, bone allografting is used to fill the defect with bone tissue from living donors or cadavers (Porter et al., 2009; Dimitriou et al., 2011).
Despite being currently used to treat bone defects, autografting has some limitations. The amount of harvested tissue and the donor site pain and morbidity are the main barriers to this procedure (Laurencin et al., 1999). Also, bone allografting, is less used than autografting because, it has some risk concerning the possibility to induce host immune response or disease transmission (Salgado et al., 2004a ; Porter et
al., 2009).
To overcome the difficulties of bone autografting and allografting, implantation of man-made biomaterials appears to be the alternative therapy to treat bone defects (Hench, 1998). Either way, the material used to adequately treat bone tissue loss, should have certain properties, such as: biocompatibility, osteoconductivity and osteoinductivity. Hence, bone substitute should not incite an inflammatory response (biocompatibility), it should provide the adequate matrix to support bone cell
18
attachment and growth (osteoconductivity), and it also should be capable of stimulate cells to become bone forming cells (osteoinductivity) (Middleton and Tripton, 2000; Albrektsson and Johansson, 2001; Finkemeier, 2002; Porter et al., 2009).
In order to achieve an ideal bone graft substitute, capable of restore bone functionality and structure, Bone Tissue Engineering (BTE) arises as a promise to the future of regenerative medicine (Salgado et al., 2004a; Schieker et al., 2006).
I.2.2.1 Bone Tissue Engineering
BTE is presented as an alternative to bone transplantation. It relies on the association of osteoconductive scaffolds, made of biocompatible materials, with cells and osteoinductive growth factors, attempting to mimic natural bone (Schieker et al., 2006). Research groups try to combine knowledge from biology, physics, engineering and materials science, in order to create a dynamic structure capable of restore, maintain or improve tissue function, structure and biochemical behavior (Hench, 1998; Salgado et al., 2004b).
One of the major challenges in BTE concerns the creation of vascularized grafts. Indeed, if the main goal is to create a construct capable of mimicking natural bone, vasculature to provide oxygen and nutrients to the engineered tissue is of the most importance (Rivron et al., 2008). After implantation of the scaffold the spontaneous invasion of new blood vessels is to slow and insufficient, and the deeper tissues became deprived of oxygen and nutrients compromising the survival of the engineered construct.
Accordingly, several attempts to provide proper blood supply to the construct are in course. These strategies include: 1) scaffold design, which allows to choose the ideal pore size and pores interconnectivity essential to the correct host cell migration and assembling in a microvascular network for the entire implanted construct; 2) a technique that combines the angiogenic effects of growth factors, such as, VEGF and bFGF to induce the ingrowth of host vessels into the construct, named angiogenic factor delivery; 3) in vivo prevascularization based on prior implantation of the
19
engineered construct in a region with an artery and after total vascularization, the construct is harvested and reimplanted at the defect site; 4) in vitro prevascularization that combines the construct cells with endothelial cells to create a new vascular network. After formation of a prevascular network in vitro the construct is implanted and the host vasculature anastomoses with it (Rouwkema et al., 2008; Rivron et al., 2008).
Ultimately, the combined results of each technique will provide the necessary information to create a successful construct with functional and mature blood vessels during the construct life time.
I.2.2.1.1. Biomaterials
In 2009, Williams proposed a new definition of biomaterial as being “a substance that has been engineered to take a form which, alone or as part of a complex system, is used to direct, by control of interactions with components of living systems, the course of any therapeutic or diagnostic procedure, in human or veterinary medicine”.
In BTE, bone substitutes used to treat bone defects and help bone healing process have in their composition biomaterials. Furthermore, and as mentioned earlier, the ideal biomaterial that composes the engineered construct should be biocompatible, osteoconductive and osteoinductive. It also should be able to support mechanical loads and be bioactive and bioresorbable. Meaning that, it should promote neotissue formation while it degrades, allowing the substitution of the scaffold for newly formed bone (Navarro et al., 2008; Dorozhkin, 2010).
Metals, polymers and ceramics are the usual biomaterials that can be used as a scaffold for BTE.
Metals and metal alloys such as, stainless steel, titanium, titanium alloys, aluminium, among others are used as permanent implants in large load bearing
20
defects. However, metal is a bioinert material and that characteristic excludes its direct use as a scaffold in BTE (Navarro et al., 2008).
As an alternative, there are bioactive and bioresorbable polymers from synthetic or natural sources. Some of the currently used polymers are: polyglycolic acid (PGA), Poly (ε-caprolaction) (PLC), polydioxanone (PDS), polylactide acid (PLA), hyaluronic acid, chitosan, alginats, collagen and others (Navarro et al., 2008). The use of these materials to do scaffolds in BTE relies in the fact that the polymer degradation rate can be controlled assigning high versatility to this biomaterial (Schieker et al., 2006).
Ceramics such as glass ceramics and calcium phosphates are also adequate biomaterials used to compose scaffolds due to their biocompatibility, osteoconductivity, osteoinductivity and high similarities with bone mineralized matrix. Furthermore, ceramics are bioactive and bioresorbable materials. One of the most used ceramics in BTE is HA (Dorozhkin, 2010; Nandi et al., 2010).
HA ceramics have different clinical applications in various forms: particles, injectable compositions, coatings on metal implants, dense or porous blocks, composites with polymers, among others. HA is often selected as biomaterial for BTE because it acts as stable phase, with lower solubility and therefore slower resorption rate, under normal physiological conditions. Additionally, it seems that HA is the ideal biomaterial to attract bone forming cells to seed, differentiate and assemble in the scaffold. However, scaffold properties, as surface area, roughness and porosity that greatly influence bone forming cells behavior, along with a poor fatigue resistance, which inhibits the use of HA scaffolds to load bearing applications, are problems to overcome (Cai and Tang, 2008; Dorozhkin, 2010).
Thus, taking into account the current progress in the nanotechnology field, there have been advances in the synthesis and study of nanoHA applied to BTE (Table 2). In fact, several methods have been used to synthesize HA nanoparticles, including hydrothermal synthesis, co-precipitation, microemulsion and sol-gel, in order to mimic bone natural crystals, in shape, dimensions and composition (Zhang and Gonçalves, 1997; Kweh et al., 1999; Kim et al., 2005; Wang et al., 2006).
21
NanoHA resemblance with natural bone minerals enhances the bioceramics biological and mechanical role. It is also known that nanosized materials (<100 nm) have large surface-to-volume ratio, which means that the smaller the particle the greater the surface area. Additionally, compared to microstructured particles, nanoHA particles present higher surface roughness and smaller porosity, which positively influence osteoinduction, osteoblastic functions and the interactions between the surface and serum proteins (Dorozhkin, 2009; Zhou and Lee, 2011).
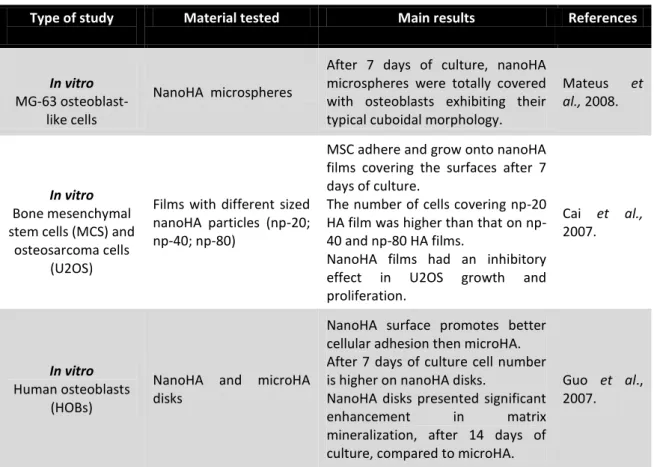
Several studies performed on osteoblastic cells with microHA and nanoHA have demonstrated the advantages in the use of nanoparticles. Thus, it has been proved that nanoHA is an adequate material to work in BTE, because it enhances the cellular response, promoting higher cellular adhesion, proliferation, migration, as well as, matrix mineralization around the implant, known as osseointegration, when compared to microHA (Guo et al., 2007; Shi et al., 2009; Zhu et al., 2010; Laranjeira et al., 2010; El-Fattah et al., 2010). The incoming information about the role of nanoHA reveals the importance of this biomaterial in future clinical application.
Table 2: Studies that used nanoHA, in vitro and in vivo, for BTE.
Type of study Material tested Main results References
In vitro
MG-63 osteoblast-like cells
NanoHA microspheres
After 7 days of culture, nanoHA microspheres were totally covered with osteoblasts exhibiting their typical cuboidal morphology.
Mateus et
al., 2008.
In vitro
Bone mesenchymal stem cells (MCS) and
osteosarcoma cells (U2OS)
Films with different sized nanoHA particles (np-20; np-40; np-80)
MSC adhere and grow onto nanoHA films covering the surfaces after 7 days of culture.
The number of cells covering np-20 HA film was higher than that on np-40 and np-80 HA films.
NanoHA films had an inhibitory effect in U2OS growth and proliferation. Cai et al., 2007. In vitro Human osteoblasts (HOBs)
NanoHA and microHA disks
NanoHA surface promotes better cellular adhesion then microHA. After 7 days of culture cell number is higher on nanoHA disks.
NanoHA disks presented significant
enhancement in matrix
mineralization, after 14 days of culture, compared to microHA.
Guo et al., 2007.
22
Table 2 (continued)
In vitro
MG-63 osteoblast-like cells
Granules of nanoHA and microHA agglomerates
Higher cellular adhesion and
proliferation achieved in nanoHA granules, than in microHA, after 6 days of culture.
Cells in culture expressed the typical osteoblastic markers (ALP; COL I; BMP-2; OPG; M-CSF). Laranjeira et al., 2010. In vitro MG-63 osteoblast-like cells
Films with different sized nanoHA (np-20; np-80)
and microHA
Compared to microHA, nanoHA films
promote superior osteoblastic
proliferation.
After 5 days of culture, cell number was higher on nanoHA than in microHA and, np-20 shows to be the most effective on cellular growth and differentiation. Shi et al., 2009. In vivo Rabbits with induced bone defects
NanoHA artificial bone with different pore sizes
Great osteogenic potential of
nanoHA combined with the adequate pore size enhances the ability of bone repair.
After 12 weeks, the implanted material was completed degraded
and bone morphology and
functionality was restored.
Zhu et al., 2009.
In vivo
Rats with induced bone defects
NanoHA and microHA pastes
NanoHA paste revealed to be a great
surface to cell attachment,
differentiation and proliferation. After 3 weeks of study, the amount of the newly formed bone around the biomaterial surface was higher in nanoHA than in microHA.
El-Fattah et al., 2010. In vivo Rabbits with induced bone defects
Artificial bones made of nanoHA powder (<100
μm) and HA
After 12 weeks, the defect filled with nanoHA was completely repaired. Thus, the biomaterial was degraded and bone morphology and function was restored.
Compared to nanoHA, HA artificial bone presented slower degradation, and promoted only partial bone repair.
Zhu et al., 2010.
I.2.2.1.2 Cells
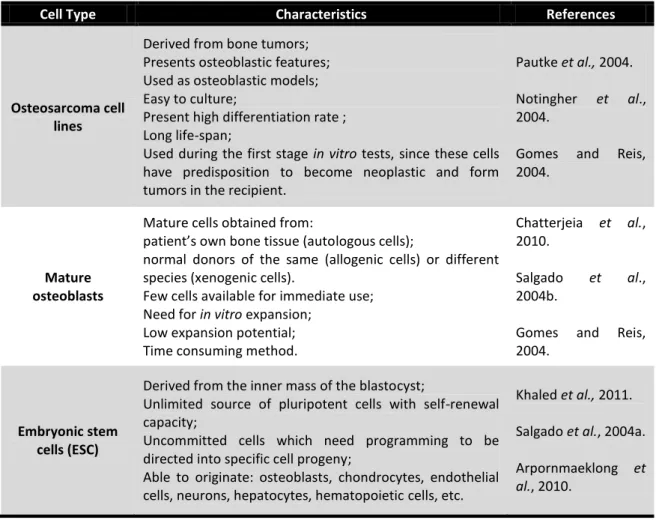
Cell type is another important element in tissue engineering. Accordingly, in
vitro studies are sensitive and rapid methods to evaluate cell morphology and