Hã}r §{riacr,4rr§n*{
lnstituto Superior
deAgronomia
Universidade Técnica de Lisboa
§V
lmpacto dos predadores introduzidos
na
ilha do Corvo
no sucesso reprodutor das populações
de
cagarro
(Calonectri
s diomedea
boreal
is)
Ana
Catarina Marcos Henriques
DissertaÇão
para obtenção do Grau
de
lVestre
em
Gestão e
Conservação
dos Recursos Naturais
Orientador:
Doutor Mark BoltonCo-Orientador:
Doutora [VariaTeresa
Ferreirada Cunha
CardosoPedro Luís Geraldes
)rs
1)9--Júri:
Presidente
-
Doutor António lt/anuel Dorotêa Fabião, Professor Associado do lnstituto Superior de Agronomia da Universidade Técnica de LisboaVogais
-
Doutora lrlaria Teresa Ferreira da Cunha Cardoso, Professora Associada do lnstituto Superior de Agronomia da Universidade Técnica de Lisboa;-
Doutor [Vark Bolton, da Royal Society Íor the Protection of Birds;-
Doutor Luís Ít/iguel Nunes Valente Afonso Reino.Lisboa (2010)
do projecto LIFE+ NAT/P/00649 llhas santuário para as aves marinhas financiado ao abrigo
do
programa LIFEe
coordenado pela Sociedade Portuguesado
Estudo das Aves/
'apea
AGRADECI]VIENTOS
"...Na verdade eu não podia viver como estes homens, mas na hora da morte queria ser um deles..."
in "A llha do Corvo"; Medeiros 1987
Este trabalho
naa
teria sido possível
sem
o
apoio,
disponibilidade, ajuda, amizade, ideias, revisões, bibliografia, fornecidos por inúmeras pessoas. Para todos omeu mais sincero
e
reconhecido obrigado.A
todos os que passaramou
estão pelo Corvo, entre outros, Pedro G, Thijs V., JB, Pedro D., Sr.Zé, Artur 5., Carlos 5., SandraH., Sandra M., Nuno lttl., Steffen O., hrlark
8.,
Joel8.,
ao pessoal do staff da SPEA, Jaime R., Vítor P. e, claro, meus amigose
família,já
se sabe.Um agradecimento às seguintes organizações que permitiram que este projecto se realizasse.
Royal Society for the Protection of Birds 'fJ' 'i.,
ar
Câmara Mlunicipal de Corvo
á
coivoE
Governo dos Açores
Secretaria Regional do Ambiente e do lvlar dos Açores
e em especiala:
Sociedade Portuguesa para o Estudo das Aves
a
?
ffi&
ffiffi
i,r&
ln the
Corvo island, Azores,there
are
important speciesoÍ
seabirds classified asprioritary
by the
Annex
I
of
the
Bird
Directive,for
example Cory's
ShearwaterCalonectris diomedea borealis,
is
one of the
speciesthat
representsin
terms
of number and density the biggest expression in this archipelago inan
European level.One
of the
major threatsfor the
birdsthat
reproducein
islandsis the
presence of predators, like rats, so, in this context, we analysed the variables that could explain the distributionof
rodents, identified the accessible places of nesting of Cory's, monitoredthe reproductive season and tried to understand the causes of fail in the egg and chick
phase.
The
results point
out
a
positive relation between
rat
abundance
and reproductive successof
Cory's.The
characteristicsof the
reproductive season are similar to those describedfor
Berlengas and Selvagens, but they differ in the hatching success and chick success. We verified that in the Íirst daysof
life of the chicks thepredation
is
very
significantand other
variables shouldbe taken into
account, for example the predation by cats.Keywords: Cory's shearwater, Corvo, reproductive sucess, invasive species, rodents, cats
The Corvo lsland has unique features, in terms of endemic fauna and flora, numbers oÍ human population, introduced predators, size of the island, among others, that gave to this island the status of Biosphere Reserve. Additionally, there are important species of
seabirds
that only
reproducein
restrictedareas
oÍ
Europe,and
are
classiÍied asprioritary
by
AnnexI
of the
Birds Directive. Nevertheless,the
populationsof
somespecies
are
stillto
confirm(e.g
Pterodroma feae, Oceanodroma castro) and others need more research in terms oÍ numbers and localization of the colonies (e.g Puffinus puffinus, Puffinus assimilis).The more conspicuous of the seabirds here present is Cory's Shearwater, Calonectris diomedea borealis and has the biggest expression in terms of numbers and densities in the Azores archipelago which represents 6O-700/" of the world population. The Corvo population is one of the major ones but the nidification areas and the actual size of the population is still not well known.
One of the major causes of the decline of the seabirds, besides the fisheries, specially
those
oneswho
reproducein
cavitieson the
soil, are
the
introduced predators in islands, places that originally yield millions oÍ seabirds. The biggest problem is that they evolved in environments that were predator free and lack defense mechanisms to facethe
predationof
eggs and
chicksby
rats and cats. This
causesthe
reproductive collapse in some species and could force the nesting places to sub-optimal conditions,that can compromise the viability of the populations. Last year, SPEA started
a
LIFE+ project "Safe islands for seabirds"to
assess the impacts of the predators on seabirds and prepare the restoration of habitat, by ways of predator and invasive plants controland
reforestationwith
endemic species.This
project
has an
important ecological relevance as it can represent a important step in the developing oÍ protocols for exotic species eradicationin
islands with human population, with both cats and rats as the major predators.The
objectives
of
this
thesis were
to
quantify
the
influence
of
rodents
in
thereproductive success
of
Cory's
Shearwater.We
analysethe
distributionof
threespecies
of
rodents (Rattus
rattus,R.
noruegicusand Mus
musculus)in
different habitats and their dependence with some environmentalvariables. The method chosenwas
by
means of tracking tunnels in two different seasons, one in April-May and theother
in
September-October,that
wantedto
express differentÍood
availability anddifferent stages
of the
reproductive periodof
Cory'sin
Corvo. Forthis
analysis weto village. Also
we
noticed some
differences betweenthe
house mice
and the rats in
terms
ofabundance and selection of habitat.
We
were
ableto
identify different accessible coloniesof
Cory's Shearwaterin
the island, and monitor almost 200 nests, once a week, in eight different places of Corvo. We analysed and compared times of hatching, fledging, incubation period and breeding success, with other islands (Berlenga, Selvagem Grande and others from Azores, suchas Graciosa) where Cory's shearwater reproduces, and finally we tried
to
understandwhich are
the
main causesof
failure usingthe
model Nest Survivalfor the
program MARK. This program is an important ecological tool for testing different factors and the correlation between them that could influence the weekly nest survival. Also give us the weight of the modelfor explaining the data.Our results point out the synchronization of times of hatching and fledging among the
different places
in the
Atlantic,but there's
a
major differencein the
hatching and Íledging successes. Surprisingly the causes oÍ failure don't seem to be related with rat abundance,as the
majorityof the
studies pointout, ant the time with
less survival probability seems to coincide with the first days after hatching when the chick is more vulnerable. Our assumption is that there is a third variable that wasn't account for, thatis
influencingthe
abundanceof
rats
andthe
reproductive successof
Cory's. This variable is the presence of cats.The
analyseof the
Life
Table demonstratethe
big
resiliencein
fluctuationsof
the population of Cory's Shearwater in Corvo lsland, a characteristic of long lived species.We
recommend thatfor the
planningof
the
eradication protocol both cats and rats should be target of control at the same time, incorporate prioritary areas of control for rodents andto
study more accuratelythe
causes offail
that influencesthe
seabirds success.lrurnoouçÃo ______-_
Relevância ecológica- _
Projecto LIFE+ llhas Santuário para as aves marinhas Área de estudo
Biologia dos predadores
introduzidos---Aves marinhas nidiÍicantes
-
cagarro----lmpacto dos predadores introduzidos nas aves
marinhas.----1 1 2 2 4 7
I
Objectivos-
---
11 Merooor-ocrA_ _ _-___ __ 12 12 13 18 18 21 22 25 25 34 40l. Distribuição e índices de abundância de roedores Análise dos dados
ll.
Monitorizaçáo dos ninhos de Cagarro e sucesso reprodutor-Análise dos dadoslll.
lmpacto dos roedores nas aves marinhas-Análise dos dadosRESULTADoS
l. Distribuição e índices de abundância de roedores
ll.
Monitorização dos ninhos de Cagarro e sucessoreprodutor-lll.
lmpacto dos roedores nas avesmarinhas---DrscussÃo
CorusroenRÇoES FrNArs .53
45
METoDoLOGIA
Tabelas
1: Habitats e tracking tunnels
associados-Pá9.
2: Variáveis ambientais
3: Modelos para o programa
MARK---Fiouras
1: Esquema de um tracking
tunnel----REsulrRoos
Tabelas
4: índices de abundância para ratos e murganhos nas duas épocas por
linha-14 15
23
12
Pág
5: Locais de nidificação dos cagarros na ilha do
Corvo---6:
Características do período reprodutor dos cagarros na ilha do Corvo. 7: Modelos testados noMARK---8: Resultados dos modelos testados no MARK
25 34 36 41 44 Fiouras
2: Abundância de ratos e murganhos porlocalem Abril-Maio
3: Abundância de ratos e murganhos por local
Setembro-Outubro---4:
lmportância das variáveis ambientais paraa
presença de ratos emAbril-Maio e Setembro-Outubro_ _ __
5: Dependência parcial das variáveis ambientais na presença de ratos na
época Abril-Maio__
6: : Dependência parcial das variáveis ambientais na presença de ratos na época Setembro-Outubro__ ___
7: lmportância das variáveis ambientais para a presença de murganhos
em Abril-Maio e Setembro-Outubro Pág 26 26 28 29 30 31
murganhos na época de
Abril-Maio_________
____________g29:
Dependência parcialdas
variáveis ambientaisna
presença demurganhos na época Setembro-Outubro 33
10: Dados totais das monitorizações aos ninhos de cagarro__
11: Fenologia das características do período reprodutor dos cagarro na ilha do Corvo
12: Número total de posturas por semana_
13: Número total de crias que eclodiram por semanâ.
14: Sucesso reprodutor e número de ninhos monitorizados por local 15: Ciclo de vida da populações de cagarro na ilha do Corvo___ 16: Função polinomial entre sucesso reprodutor e abundância dos ratos
35 36 37 38 39 39 42
17: Sucesso de eclosão e sucesso de saída do ninho de acordo com a
abundância de ratos por
local___
____________ 4918: Sucesso reprodutor ponderado
semanal___
___________43ANEXoS
Tabelas
pág.9: ldentiÍicação das linhas e habitats dos tracking
tunnels
___Anexo 2(1) e 2(z)10: Valores do "Mean decease accurancy para as variáveis ambientais
que aÍectam a distribuição dos
ratos_
_____Anexo 511: Valores do "Mean decrease accurancy" para as variáveis ambientais
que aÍectam a distribuição dos
ratos_
_____Anexo 612:
Comparaçõesentre
as
Íenologiasdo
período reprodutor nasSelvagens, Berlengas e Açores (Klamp & Furness
1992)______
____Anexo 1113: Modefos testados no MARK, tabela
completa___
___Anexo 1214:Malrizde
Leslie
_______Anexo 13Fiouras
í9: Localização dos tracking tunnels na ilha do Corvo
Pág. _Anexo 1
. Anexo 3
26: Localização dos ninhos de Cagarro na ilha do
Corvo---27: Ninhos de
28: Adulto de cagarro dentro do ninho.
31,32 e 33: Predação de adulto, ovo e cria,
respectivamente---34: Esquema exemplificativo do eÍeito da variável gatos
_Anexo 7
. Anexo 8
_Anexo 8 29 e 30:Adulto e cria, cria no ninho respectivamente
---
---Anexo
9 _Anexo 10Esta
tese
de
mestradoÍoi
desenvolvida como parte integrantede
algumas das acções do projecto LIFE+ "llhas santuário para as aves marinhas", nomeadamente aonível
da
análiseda
distribuiçãoe
abundânciados
roedores introduzidos(ratos
emurganhos) e monitorização da época reprodutora das populações de cagarro na llha do Corvo.
Relevâncla eco!ógica
As comunidades insulares começaram
a
despertar interesse paraos
naturalistasdesde
Darwin,como
evidênciaspara
a
origem
derivativadas
espécies.As
ilhasoceânicas
são
habitats
naturais
de
comunidadesde
plantas
e
animais
comrelativamente
pequena
diversiÍicação,redes tróficas
simples
e
altas taxas
deendemismo (Chapuis
eÍ
a/.
1995). Em ecossistemas insulares, mais susceptíveis àentrada
e
naturalizaçãode
espécies invasoras, quando uma espécieé
introduzida, acidental ou intencionalmente, as consequências podem ser devastadoras, sendo um dos principais factores de ameaça à Biodiversidade do planeta. De facto, a maior parte das extinções a nível mundialocorreram ou estão a ocorrer em sistemas insulares. Por exemplo, desdeo
séculoXVll,93%
das
176 espéciesou
subespéciesde
aves queforam
consideradasextintas
ocorreramem
ilhas (King
1985). Neste sentido,
a viabilidade das populaçôes de aves marinhas nidificantese
coloniais encontra-se em riscoa
longo prazo,já
queé
apenasem
ilhase
ilhéus, onde a faunae
flora nativasevoluiu
sem
a
pressãode
predadorese
competidores naturais,guê
estas
avesconseguem encontrar combinações
de
habitatóptimo
e
ausênciade
predadores(Ricklefs 1990) para
se
reproduzirem.A
introdução
de
espécies exóticas
é considerada umasdas
maiores causasde
perturbaçãodo
ecossistemae
perda de biodiversidade em ilhas (Moors & Atkinson 1984; Atkinson 1985; Williamson 1996) etem
causado enormes impactos paraa
agricultura, saúde pública, turismoe
outras actividades económicas, actuando sinergisticamente comas
alterações climáticas e redução das áreas geográficas de distribuição dos habitats naturais (Silva et a/. 2008).No
entanto entre todasas
espécies exóticasé
necessário averiguar quaisas
que adoptaram carácter invasor, quais as que serão possíveis de erradicar ou controlar equais são as espécies consideradas prioritárias em termos
de
acçõese
medidas decontrolo. Este tipo de estudos, nomeadamente ao nível
da
prevençãoe
erradicação,devem
ser
preÍerencialmente realizadosem
ilhas isoladase
de
pequena dimensão (Angel et al.2006).O
arquipélago dos Açores, sendo um grupo de nove ilhas especialmente isolado,jovem
e
fruto
de
acontecimentos pré-históricos (glaciações, vulcõese
sismos) apresenta uma baixa diversidade biológica, nomeadamente
a
nível da flora, quandocomparado
com
os
outros
arquipélagosda
macaronésia,sendo
por isso
muito susceptível à entradae
naturalização de espécies exóticas (Santos-Reislggg).
Além disso, é um arquipélago que Íornece habitat de nidificação para muitas aves marinhas que se reproduzem em zonas muito restritas a nÍvel mundial.Projecto LIFE+
"llhas
Santuário para as Aves Marinhas,,O projecto LIFE+, cuja área de intervenção é a ilha do Corvo e ilhéu de Vila Franca
do
campo,
pretende,
a
longo ptazo,
um
aumento
substancialem
termos
dadistribuiçáo
e
da
densidadede
aves
marinhas consideradas prioritáriase
que
sê reproduzem em zonas muito restritas a nível Europeu. Por outro lado, contribuirá paraalcançar
o
objectivode
parar
a
perdade
biodiversidadepara 2010
e
cumprir os objectivosde
coesãoda
União Europeia noque
respeitaàs
regiões menos viáveis economicamente, como é o caso dos Açores, mais especificamente a ilha mais remota e menos habitada do arquipélago, a ilha do Corvo.Das acções deste projecto destacam-se a análise da distribuição e abundância dos mamíÍeros exóticos na ilha do Corvo
e
ilhéu de Vila Franca; identificação de espéciese
áreas utilizadas paraa
nidiÍicaçãode
aves marinhas bem comoos
impactos dos mamíÍeros introduzidos na faunae
flora nativas, iniciaçãoda
restauraçãode
habitatpara
as
aves
marinhas
em
algumas
áreas
seleccionadas, nomeadamente por reflorestação com espéciesde
flora endémicas bem como promoçãode
acções decontrolo
de
flora
exótica. Alémdisso,
pretende-se promover acçõesde
educação ambiental e empreendedorismo que visam a conseryação dos valores naturais da ilha.Um
dos
objectivos principais prende-secom
a
elaboraçãode um
protocolo para controloe
futura erradicação dos predadores introduzidosem
ilhas habitadas,o
que torna este projecto pioneiro a níveleuropeu.Área de estudo
A
llhado
Corvo, nos Açores está situada no extremo Noroestedo
arquipélago, possui uma área de 17 km2 e uma única povoação, a Vila do Corvo, com cerca de 400habitantes. A nível da flora terrestre registam-se 95 endemismos, dos quais Sg sâo do arquipélago dos Açores, 19 da macaronésia
e
17 da Europa. Em termos de artrópodesterrestres estão registados
24
endemismosdos
Açorese
seisda
macaronésia. Nogrupo
dos
Moluscos,estão
identificadoscinco
endemismosdos
Açorese
um
damacaronésia.
A
ilhado
Corvo possui 18tipos
de
habitat natural abrangidos pelo AnexoI
daDirectiva Habitats, sendo quatro considerados prioritários.
De entre estes
habitats destacam-se as charnecas macaronésicas endémicas (4050), turÍeiras (7110, 7130 e91D0), enseadas
e
baías pouco profundas (1160),grutas
marinhas submersas ou semi-submersas (8330), falésias com vegetaçãodas
costas macaronésicas (1250), vegetação vivaz das costas de calhaus rolados (1220) e recifes (1170)'Em termos
de
flora, salientam-seas
espéciesque
caraclerizamas
florestas de Laurissilva macaronésica como pau-branco Picconia azorica, faia Myrtca taya, urze Erica azorica, cedro-do-malo Juniperus brevifolia, vidália Azorina vidalli, entre outras.No que diz respeito ao uso do solo, maioritariamente é utilizado para pastoreio de
gado
bovinoo
que
restringiuas
principais zonas arbóreasàs
ribeirasque
abrigam espécies como o incenso Pittosporum undulatum (de carácter invasor) e a criptomériaCriptomeria
japonica.
Existem, actualmente,poucas zonas
de
vegetaçáo natural característica da Íloresta de Laurissilva e a maioria são provenientes de antigas zonade
pastagemem
regeneração natural. Nas zonasde
maior altitude (e.9. Caldeirão)ainda permanecem extensas áreas
de
turfeira,no
entanto encontram-seem
risco devido ao pastoreio e pisoteio do solo. As espécies do género Sphagnum sp., que aquise
encontram,são
importantes paraa
retençãode
águana
ilhae
manutenção delagos
que
abrigam algumas espéciesde
aves pouco comuns paraa
Europa (e.9.garças,
patos,
narcejas, galinholase
outras
limícolas migradoras provenientes do continente americano).Na ilha
do
Corvo,e
à
semelhança das outras ilhasdo
arquipélago, as principaisameaças
à
biodiversidadeÍlorística
e
biótopos naturais
relacionam-secom
asmonoculturas para pastagem.
Cerca
de
41o/oda
área emersada
ilha está classificada sob as designações desítio
de
importância comunitáriae
Zona Especialde
Protecçãono
âmbitoda
rede Natura 2000da
União Europeia. Comporta ainda três zonasde
protecção integral à apanha de lapas e uma reserva voluntária marinha, que embora não sendo de estatuto oÍicial, representa um bom exemplo do esforço da população para a conservação do património natural. No ano 2006foi
criadoo
Parque Natural Regional do Corvo, queinclui
o
Caldeirãoe
as
falésias em terra, delimitado pelas coordenadas geográficas estabelecidaspor
Decreto Legislativo Regional (DRL na56/2006122 Dezembro) que integra nos seus limites azona de reserva de Biosfera outorgado no ano 2007. A área total classificada como reserva da Biosfera da llha do Corvo é de 25.853ha.O Check
List"das
avesdo
arquipélago dos Açores (Le Grand 1983) refere 14espécies nidificantes para a ilha do Corvo, das quais, oito são subespécies endémicas
dos Açores e uma é subespécie endémica da macaronésia.
As
aves marinhasaté
agora identificadas que aquise
reproduzem são: cagarroCalonectris
diomedea
borealis,garajau-comum
Sterna paradisea, garajau-rosadoSterna
dougalli,
estapagado Puffinus puffinus e frulho Puffinis assimitis. A nidificaçãode
freira-do-bugio Pterodromafeae,
roquinho Oceanodroma castroe
roquinho-de-monteiro Oceanodroma roquinho-de-monteiroi (espécie recentemente descrita) encontra-se aindapor confirmar. Todas
as
avesda
ilhado
Corvo estão classiÍicadas como prioritárias pelo Anexo I da directiva Aves.Biologia dos
roedoresintroduzidos
Na ilha
do
Corvo estão identiÍicadas oito espéciesde
mamíferos (Mathiaset
al. 1998; ICN 1999)- Roedores: rato-preto Rattus rattus, ralazana-castanha Rattus norvegicuse murganho
Mus musculus (lCN 1999; Collares-Pereira et
al.2007)
- gato doméstico Felis catus
-
cabras, ovelhas e vacas-
morcego-dos-Açores Nyctalus azoreum (nãoestá
confirmadaser
esta
a
espécie residente no Corvo)Com excepção da espécie endémica morcego-dos-Açores classificada como rara,
todas
as
outras
apresentam estatutode
não
ameaçado. Sendo,na sua
maioria,comensais
em
território
continental,
estas
espécies
viram
aumentada
a
suaprobabilidade
de
colonizaçãoe
sucesso
em
meio
insular,
onde
a
ausência depredadores
de
médioe
grande porte
representaum factor
decisivo (Santos-Reis1ee8).
A introdução de roedores nas ilhas do Atlântico, nomeadamente no Corvo, foi mais
ou
menos recente, cercade
400
anospara
o
corvo,
quando comparadascom
oMediterrâneo
que
datamde
introduçõesde
2000 anos.
Neste pontoé
importanteentender qual
o
efeitoa
longo prazodas
espécies exóticas na comunidade nativa, tendo em conta as características da ilha bem como com o tempo de introdução dos roedores.As
três
espéciesde
roedores
cosmopolitas, rato-preto, ratazana-castanha emurganho, evoluíram provavelmente
no
continenteAsiático,
donde
dispersaramassociados
a
sistemasde
transportes,e
actualmente ocupamtodas
as
partes do mundo (Buckle & Fenn 1992). Grande parte do êxito evolutivo dos roedoresrelaciona-se
com as
reduzidas dimensõesda
maioriados
indivíduos, possibilitando-lhes oacesso
a
fontes alimentares pouco exploradaspor
outros animais,a
sua coloraçãomimética,
o
que
evita
possíveis predadores (Mathias
et al.
1998)
e
elevadaÍecundidade
e
posiçãonas
cadeias tróficas (Mathias 1998), contribuindo para que atinjam números consideráveis, ameaçando a biodiversidade dos ecossistemas.Rato-preto
Habitat: generalista, podendo
ser
encontrado desdeo
níveldo
mar até altitudes elevadas. Adapta-se facilmentea
climas rigorosos onde ocupa Íirmemente um nichorico em
recursos
alimentares,
com
poucos
predadores
e
com
competição interespecífica mínima (PyeeÍ
a/. 1999). Segundo Mathias (1998) encontra-se mais frequentemente em áreas florestais ou agrícolas e evita biótopos muito húmidos sem cobertura arbustiva ou arbórea,Áreavital:
variade
0.5a
tha
(Dowding & tvlurphy 1994). No lnverno, reduzindo-seo
número de esconderijos habitáveis,as
populações tendema
concentrar-se mais àvolta das habitações humanas (Pye eÍ a/. 1999).
As
populaçõesde
rato
pretoque
vivemÍora das
zonas urbanas, normalmente apresentam ciclos de reprodução e abundância de população de acordo com os ciclosde
colheitas. Foi verificadoque
consegue subsistir mais facilmentena
ausência da ratazana-castanha revelando-se, por isso, fortes competidoras.Ratazana-castanha
Habitat: ocupa preferencialmente zonas urbanas e normalmente encontra-se junto à água, onde se alimenta de restos de comida, sendo menos comum em áreas cultivadas.
Árealvital: bastante variável. Um estudo nas ilhas Kapiti indicou áreas vitais na média dos 5.8ha (Bramley 1999).
A
ratazana-castanhaé
uma espécie terrestre, escavadorae
com pouca aptidáopara trepar.
É
potencialmente capazde
se
reproduzirtodo
o
ano
em
climas temperados quando há comida disponível, mas normalmente reproduz-sede
Maio aOutubro (lnnes et
al.
2001).Murqanho
Habitat:
ocorre com
elevadas densidadesem
biótoposde
marcada influência humana (tvlathias 1998, lvloreira 1987). No caso das populaçÕes que ocupam ilhas,devido
à
diminuiçãode
potenciais predadorese
competidores,o
murganho coloniza também áreas naturais, como a floresta indígena, podendo atingir números igualmente consideráveis e propiciar impactos negativos a estes ecossistemas (Moreira 1987).Área vital: constrói
os
seus ninhos perto das fontes alimentarese
raramente se afasta mais de 10 metros das mesmas (King eÍ a/. 1gg6).O murganho está listado como uma das cem piores espécies invasoras de todo o mundo
("one
Hundredof the
world's worst
lnvasive Alien'species") pelo grupo de especialistas em espécies invasoras do IUCN (União [rlundial de Conservação) (Loweet al.
2000).O
seu modode
reprodução estáem
muito relacionado como
elevadosucesso
na
colonizaçãode
ilhas. Num estudo
na ilha
lt/acQuarieem
1993, Pyeverificou
que
a
biologia reprodutorado
murganho tinha uma base sazonal, na qual correspondia um período enastro de apenas 3 meses no lnverno. A reprodução inicia-se no início da Primavera e ocorre continuamente até finais do Outono, os machos são potencialmente capazes de se reproduzirem todo o ano. Nas populações selvagens areprodução atinge
o
seu pico no Verão (Fitzgerald etal.
1981; Wilson et a/. 2006) etende a diminuir acentuadamente no fim do Outono
e
lnverno (Fitzgerald et a/. 1g81;Jtíoreira 1 987; Wilson et al. 2006).
O efeito destas três espécies
de
roedores nos biotas insulares tem sido severo eabrange diferentes níveis
no
ecossistema.Todos
sâo
omnívoros, predando aves,pequenos mamíferos,
répteis,
insectos,anfíbios
e
moluscose
consumindo umagrande variedade
de
espéciesde Ílora. No
entanto estas espécies diferem no querespeita
a
caracteríslicas comportamentaise
portanto
no
seu
potencial impactoecológico.
Apesar
de
pequeno,
o
murganho apresenta
grandes
densidadespopulacionais,
a
ratazana-castanha é grande e fossorial, com implicações a nível dasaves que nidificam no chão ou em buracos, ao passo que o rato preto é mais pequeno mas um trepador nato com implicações a nível dos ninhos em árvores.
Aves marinhas
nidiÍicantes
-
CagarroOs estudos que afectam o sucesso reprodutor nos Procelariformes têm produzido
um
elevado númerode
publicações mas cujas conclusões são surpreendentemente diversas para um grupo tão homogéneo, variando a nível da espécie e mesmo para amesma espécie entre os diferentes estudos (Mougin 1998).
Uma
das
mais
abundantesaves
marinhas
da
ilha
do
Corvo
é
o
cagarroCalonectris
diomedea borealis,
um dos
maiores
ProcelariÍormes nidificantes do Atlântico Nordesle, essencialmente pelágico, que tolera ventos fortes e águas agitadas(Cramp
&
Simmons 1977\. Existemtrês
subespécies conhecidas (Warham 1990) aC.d borealis
das
ilhasda
Macaronésia (Canárias, Madeirae
Açores), C.d. edwarsii (Cabo Verde)e
C.d.diomedea (Mediterrâneo) (Monteiro&
Furness 1996). Nidifica em ilhase
ilhéus, originalmente isoladose
livres de predadores terrestres, em fendas de escarpas, cavidades naturais no solo ou tocas de coelho. Em relação ao cagarro, as ilhas dos Açores abrigam 65-70/" da população mundial (Monteiroetal.
1996) e umadas
maiores populaçõesdo
arquipélago (cercade
30
mil
indivíduos)é a
do
Corvo(Ramirez
et at.
2OO8),no
entantonesta ilha
as
populaçõesestão mal
estudadas (Ramos et a\.1997).A
subespécieC.
d.
borealis
está
incluída
no
Anexo
I
da
DirectivaAves
eConvenção
de
Berna,e
apresenta estatutode
conservação considerado vulnerável pelo IUCN.Caoarro
Os cagarros têm uma esperança média de vida de 50-60
anos, passam 90%
da
sua vidano
mare
só
aos oito anos regressam para as colónias de nidificação onde permanecem9
meses, desde finaisde
Fevereiro até Íinais de Outubro. Asua principal área pelágica durante
o
lnvernoé a
zona este da América do Sul (Monteiro & Furness 1996). Entre as avesmarinhas
que
se
reproduzem
em
colónias,
o
cagarrodistingue-se
pelo
elevadograu
de
sincronizaçãode
certas actividades nos locais de nidificação (Hammer & Read 1987; Thibault et a\.1997).As colónias mais estudadas pertencem às da Selvagem Grande, (Zino 1971;Zino
et al.
1987) Berlenga (Granadeiro 1991),ilhéu
da
Praia,
na
Graciosa,onde
estáapenas referido
o
períodode
postura (Monteiroet al.
1996)e
ilhéuda
Vila, SantaMaria.
As
fenologias
apresentadas
para
os
diferentes locais denotam
umasincronização entre e dentro de colónias, com as datas médias de postura entre final
de
lMaioe o
início de Junho,a
eclosão em finais de Julhoe
a
saídado
ninho entre finais de Outubroe
inícios de Novembro (Zinoet
al. 1987; Granadeiro 1991; Monteiro eÍ a/. 1996).Após a chegada das aves aos locais de nidificação, a sua actividade reparte-se por
um período diurno de alimentação no mar ou em terra
a
incubaro
ovoe
um períodonocturno,
onde
estão
em terra nos
locais
de
cria
e
desenvolvemuma
intensa actividade aéreae de
vocalização, Ao entardecer reúnem-se em grande número nomar, perto
das
colónias. Sendo uma espécie gregária,as
zonasonde
nidifica são densamente povoadase
cada fêmea adulta põe um único ovo, aparentemente sem capacidade de reposição nos casos de perda acidental (Cramp & Simmons1gl7).
É.uma espécie monogâmica em que evidências da estrutura genética das populações de
cagarro
demonstraramque não
existe
fertilização extra-par, confirmandoque
osistema genético coincide com o socialque tem sido observado na espécie (Rabouam et a|.1999). Os principais factores de ameaça relacionam-se com poluição, nas zonas
de
alimentaçãono
mar
e
locaisde
nidificação,e
predadores,por
competição poralimento perturbação
e/ou
predaçãode
ovos
e
crias.A
informaçãoque
existe dalocalização
e
abundância de cagarroé
limitadae
encontra-se desactualizada, sendoque
o
Últimocenso
realizadoem
1996 contabilizoucerca
de
403
g20
indivíduos (Almeida et al.2005).lmpactos dos predadores
introduzidos
nas aves marinhasOs
impactos dos predadores introduzidos nas aves marinhas estão relacionadoscom
alteraçõesno seu
usodo
habitate
sucesso reprodutorque por
suavez
está parcialmente dependente da distribuiçãoe
comportamento do predador (Lahti 2001). Os ratos têm sido dos predadores introduzidos com maior sucesso nas ilhas oceânicaspelos humanos
e o
seu
impacto, especialmenteao
nívelde
avescom
estratégias reprodutivas do tipo colonial, tem sido enorme (ttloors & Atkinson 1gg4).Antes de ser colonizada, a ilha do Corvo, abrigava milhões de aves marinhas que actualmente
viram
reduzidoo
seu
habitatde
nidificaçãoe
aumentodo
insucesso reprodutor. As espécies introduzidas há cerca de 400-5OO anos, como os gatos, ratos e murganhos podem ter influência na viabilidade das populações naturais e endémicasda ilha do Corvo, biodiversidade de aves e a sua restrição para habitats sub-óptimos.
A
predaçãode
aves por mamíferos introduzidosé
um fenómeno que levoujá
váriasespécies
à
extinçáo
e
ao
declínio
populacional
de
outras (Atkinson
1985), nomeadamente as espécies antropogénicas representam uma das maiores causas de extinções em ilhas (Clout & Veitch 2003).Segundo
a
BirdliÍe Internacional (2004), 25o/" dasaves
ameaçadasem todo
o mundo são afectadas por predadores introduzidos. Diversos estudos começaram a ser desenvolvidos nos anos80
atéà
actualidadel,e
comprovaramjá
a
forte influênciaque
a
predação de ninhos exerce no sucesso reprodutivo das comunidades de aves.Por
exemplo,na
Nova
Zelândia,o
rato preto
causou graves danos
a
algumas populações de aves florestais, como foi o caso dos passeriformes Anthornis melanura,Petroica
australise
Philesturnus carunculatus,que
sofreram
um
rápido
declíniopopulacional
(Atkinson 1985).
Wilson
et
al.
(2006) afirmam
que
os
predadoresintroduzidos
tiveram
um
papel
determinanteno
declínio
populacionalda
espécie Nestor meridionalis, papagaio endémico da Nova Zelândia, havendoo
risco de viremmesmo
a
causara
sua extinção, caso não haja controlodas
populações. TamtÉmlnnes eÍ al. (2001) verificou que a predação é a causa primária do declínio continuado
da
populaçãodo
passeriforme Callaeas cinerea wilsoniem
florestas primárias deNorth lsland, na Nova Zelândia. Os resultados
de
um estudo acerca da predação de ninhosde
Apus pallidus, realizado em ilhas mediterrânicas, sugerem igualmente queos
ratos influenciam fortementea
diversidade localdas
comunidadesde
avese
adistribuição
de
muitas espécies.
Em
diferentes
ordens
de
aves
(anseriformes, ciconiiformes, galliÍormes, passeriformes, tinamiformes, etc.)o
sucesso reprodutivo é Íortemente influenciado pela predação e temo
potencial de afectar Íortemente a taxa de crescimento populacional(Sather
and Bakke, 2OO0 in Stephens et a\.2003). Outro exemplo,a
Halobaena caerulae (criticamente em perigo pelo IUCN) desapareceu dallha Macquarie, no Oceano Pacífico, devido principalmente
à
predação por ratose
asua zona de nidiÍicação está actualmente reduzida
a
ilhéus ao largo da ilha, onde os ratos não chegam. Outras aves marinhas são também afectadas pelos ratos, devido à sua perturbação nas zonas de nidificação causando o abandono prematuro dos ninhos (Angel et al. 2006) e consequentemente o colapso reprodutor das populações de aves marinhas.O
carácter generalista associadoà
facilidadeem
acedera
ninhos muitas vezes inacessíveisa
gatos, permitiram aos ratos, apósa
sua introdução, explorar as aves marinhas como um novo recurso alimentar, ovos, criase
mesmo adultos. Outra das'Wilcove
1985, Pierce 1986, Small & Hunter 1988, Major 1991, Andren 1992, Sieving1992, Vickery et al. 1992, Laurance el a/. 1993, Rudnicky & Hunter 1993, Major et al. 1994, Bosque & Bosque 1995, Haskell 1995, Penloup et al. 1 997, Wilson et al. 1997,
Hernandez eÍ a/. 1999, lnnes et a/. 1999, Martin & Joron 2003 e Thompson
lll
& Burhans 2003 in Sanches 2008razões relaciona-se com o facto de muitas espécies de aves marinhas terem evoluído
em
áreas
às
quais
os
mamíferostinham acesso
limitadoou
mesmo
nulo
(ilhasoceânicas,
falésias
inacessíveis)
daí
não
possuem quaisquer
adaptações comportamentaisou
ecológicasque
permitama
sua
coexistênciacom
predadoresintroduzidos
nem
com
a
perturbaçãodo seu
habitatde
nidificação, tornando-as especialmente vulneráveisa
introduçõesde
mamíferosem
ilhas.
Na
ilha
de
LordHowe,
em
1918,e
no
espaçode
uma década, maisde
4Oo/"de
espéciesde
avesterrestres Íoram extinguidas depois
da
invasão por rato-preto (Lack 1968; Burger & Gochfeld 1994).Na llha do Corvo os gatos Íoram introduzidos como animais de companhia e com o intuito de controlar as populações de ratos da ilha. Consequentemente, esta questão deve ser tida em consideração quando se aborda a diminuição do sucesso reprodutor das aves marinhas, podendo estas aves sofrerem um impacto substancialderivado da pressão exercida por parte dos gatos. Segundo Paton (1991
e
1994) os gatos podem influir directamente nos números de crias que eclodem anualmente; inclusivamente, na cidade de Adelaide, Austrália, os gatos Íoram responsáveis pela predação de cerca de50% das
populaçõesde
aves
nativas, nomeadamentenos juvenis
recentementeeclodidos
e
incidiu também
nos
roedores
introduzidoscomo
o
rato-pretoe
omurganho. Outro estudo realizado
por
Anon (1994)em sete
cidades Australianas, indicou que, para a maioria dos gatos, os roedores introduzidos constituíam a maioria das espécies predadas.Os estudos na Austrália são suportados e amplificados pelos trabalhos realizados
neste âmbito
na
Américado
Norte
e
Europa,em que
os
gatos
domésticos eassilvestrados usualmente
preÍerem pequenos mamíferos,
aves
e
lagartos,especialmente as presas que têm pesos inferiores a 1009. A preferência por presas de pequeno porte foi também confirmada num estudo de Childs (1986), na zona urbana
de
Baltimore.A
nível individual, alguns gatos assilvestrados introduzidosnas
ilhas especializaram-se em capturar espécies particularese
normalmente não habituais, amaioria destas constitui
aves
e
mamíferos
terrestres.
Na
ilha
Marion
os procelariformes constituema
maioriadas
presas,em
particularos
juvenisque
são directamente removidos dos ninhos (Bradshaw 1992).Objectivos
De acordo com as acções do projecto LIFE+ "lhas Santuário para as Aves
Marinhas" no Corvo e Vila Franca, e dada a relevância ecológica do estudo do impacto dos mamíferos introduzidos em ilhas na viabilidade das aves, os obiectivos propostos para esta tese relacionam-se com três capítulos gerais e as questões que se pretende responder para cada um são apresentadas em seguida:
l.
Distribuição
de roedores nosdiÍerentes
habitats e análise das variáveis que explicam a suadistribuição.
Quais os índices de abundância dos ratos e murganhos nas áreas de estudo?
Qualo efeito da época nos índices de abundância?
Que variáveis ambientais podem influir na distribuição dos ratos e murganhos na ilha do Corvo?
Nova identificação de habitats para a ilha do Corvo.
l!.
Monitorização da época reprodutora dos cagarros e análise dosucesso
reprodutor
Serão as datas de postura, eclosão e o sucesso reprodutor dos cagarros na ilha do Corvo semelhantes às das restantes ilhas da macaronésia?
Será o sucesso reprodutor concomitante com a viabilidade das populações de cagarros na ilha do Corvo?
!ll.
ldentiÍicar
causas deÍalha
nas Íases de ovo e cria e perceber qualo impacto
dos
roedores na época reprodutora.Será que uma redução na população de ratos resultará numa tendência populacionalpositiva ou o sucesso reprodutor está relacionado com outras razões ?
Há alguma relação entre taxas de sobrevivência de ovos e crias e a
abundância de roedores?
Quais os estádios que mais influenciam o sucesso reprodutor?
Haverá alguma relaçáo entre as medidas do ninho e o sucesso reprodutor do mesmo?
Terão os gatos impacto no sucesso reprodutor dos ninhos?
A metodologia que se segue é apresentada segundo a mesma linha dos capítulos e objectivos específicos para cada um.
l.
Distribuição
de
roedores
nos
diÍerentes
habitats
e
análise
das
variáveis que explicam
a
sua distribuição
O desenho experimental para o cálculo da abundância de roedores baseou-se na instalação
de22linhas
de tracking tunnels, num totalde
110 armadilhas, distribuídas por toda a ilha, de acordo com a identificaçáo dos habitats realizada no início de Abril(Tabela 1
e
Anexo 1). Este tipode
metodologia tem sido largamente utilizado (llhas Falkland, Maui no Hawaiie
em alguns lugares na Nova Zelândia) para monitorizar aabundância
de
pequenos mamíferos, sendo umaforma
bastante eÍicaza
nível de custo e facilidade.Cada linha
utilizadafoi
compostapor
cinco tracking tunnels, espaçados50
m(adaptada
de
Cooper&
Crossland 2007) entresi
e
100m
no
mínimoentre
linhas (Brownet al.
1996)(Anexo2.1 e
2.2). As armadilhas foram colocadas7
dias in situpara habituaçáo dos animais
e
um diade
iscagem após este período. Para isco foiutilizado manteiga
de
amendoim, considerada alimentíciae
olfactivamente atractiva(Mira
1990in
Marques 1997).Os
tracking tunnels utilizados foram essencialmentecompostos
por túneis
de
polietilieno rectangulares desenhadospara
permitir
apassagem
dos
animaissem
qualquerforma
de
captura (Anexo3). No
centro dotracking
tunnel
foi
colocadauma
esponja embebidaem
corante alimentar,e
nas extremidades duas folhasde
papel branco presas com elásticos nas extremidades (Figura 1). No dia seguinte as folhas foram recolhidase
identificadas as pegadas dos murganhos e dos ratos.lsco Placa em forma de tubo Esponja embebida com corante Folha de papel Elásticos Tábua de madeira
Figura
1:
Esquema exempliÍicativoda
montagemde um
trackingtunnel
para iscagem.Foram
realizadasduas
amostragensuma
em
Abril-Í\íaioe
outra
Setembro-Outubro. A primeira amostragem pretende reÍlectir uma baixa disponibilidade alimentarjá
que é o Íinaldo
lnverno eo
início da época reprodutora dos cagarrose
a segunda amostragem, finaldo
Verão, maior disponibilidade alimentare
sobrepõe-se ainda aofinalda
época reprodutora dos cagarros.É de referir que, na monitorização de roedores e mustelídeos, o método dos túneis
é
apenas uma medidada
sua actividade, fornecendo-nosum
índicede
abundânciarelativa,
e
não uma
medida directada
densidade populacional (Gillies&
Williams 2002).Análise
dos
dadosA identificação das marcas das diferentes espécies Íoi auxiliada pelo guia: "A short guide Íor identifying footprints on tracking tunnel papers" de Gillies & Williams (2002).
Os dados para as duas espécies
de
ratos Íoram tratados em conjuntojá
que não é possível distinguir as pegadas.O
índicede
pegadas paraa
abundância relativade
roedoresfoi
expresso em percentagem média de túneis marcados por linha. Os dados foram inseridos e tratadosem
lt/icrosoft Excel, numa cópiada
folha
de
dados
usadapelo
Departamento de Conservaçãoda
Nova Zelândia (dme://LIAMRO-S,3370/), que calcula a taxa percentualmédia (TPM)
de
pegadaspor
linhae
para cada
espécie (ratose
murganhos) da seguinte forma:tr
somatório linhas (Túneis com peqadas x 100 / Túneis disponíveis)Nq total de linhas
Para o cálculo da média da espécie por linha:
o
Número de presencas na linha(rato oumurqanho)
x 1005 (total de túneis em cada linha)
Realizaram-se
estes
cálculospara
as
duas
épocasde
amostragem,uma
para Abril-ttlaio e outra em Setembro-Outubro.Após reconhecimento local
os
diferenles habitatsda
ilha foram identificados poranálise visual
e
divisãoem
unidades coerentese
a
cada um
atribuídoum
código identificativo (Tabela 1). Em cada habitat foi instalado um número variável de trackingtunnels, dependente
da
homogeneidadedo
locale
dimensãodas
áreasde
habitat contínuo (Anexo 2.1 e 2.2).Tabela 1. IdentiÍicação de habitats da ilha do Corvo, número de túneis associado e número da linhas correspondente (Total de 110 túneis, 22 linhas em 10 habitats).
Descrição Na de Tracking tunnels Habitat Linha Hortas muradas
(MHl)
Tamarix (MH2) Falã Rochosa (MH3) Ribeira (MH4) Pasto Murado (MH5) Urze (MH6) Pasto com Urze (MH7) Arbóreo (MH8) Canas (MHe) Caldeira (MH10)Localizam-se ao nível do mar e correspondem a campos murados de hortículas e cereais
Zonas abandonadas
com
espécies exóticas.São
zonas
de
terrenos
incultos
perto
da Íalésia e expostas ao marAo
níveldo
mar, zonade
calhau rolado comespécies
do
intertidal.
Perto
do
porto
de embarqueRibeiras onde
se
mantêm espécies arbóreasprincipalmente
de
criptomeriae
incenso. O caudal enche no invernoZonas
privadas delimitadas
por
muros
de pedra com erva de pasto Para vacasZonas de
vegetação
endémica maioritariamente constituídas por urzeÁrea de pasto abandonadas em regeneraçáo
natural
onde
ocorrem algumas
espécies endémicas como o cedro do mato e a urzeZonas
de
vegetação
arbórea
sem
águas interioresZona perto da estrada com Arundo donax
Zona de maior altitude da ilha, com zonas de turfeira, lagoas e erva de pasto utilizada como baldio
10
1e15
102e16
103e4
105e9
6,7,8e
1410
10e11
5 12 20 15 13,21 e 2210
17e18
10
19 e 20t4
O
cálculodos
índicesde
abundânciade
roedorespor
habitatÍoi
efectuado de acordo com a seguinte fórmula:tr
Nn de trackinq tunnels com presenca de roedores (ratos ou muroanho) xi00
Ne totalde tracking tunnels do habitat
Procedeu-se
à
representação gráfica,em
Microsoft Excel,da
percentagem depegadas de roedores para cada habitat nas duas épocas.
Foi testado se existiriam diferenças significativas entre
os
meses de Abril-Maio eSetembro-Outubro na abundância de ratos e murganhos, por meio do teste t, amostras
emparelhadas,
lC=95%.
Esta
análise
foi
realizadacom
recurso
ao R
z.g.z
(RDevelopment Core Team 2009).
Variáveis ambientais oue influem na distribuicão dos ratos e murqanhos
Além
dos tipos
de
habitat, consideraram-seoutros factores
que, em
algumamedida, poderiam influir na abundância dos roedores, registando-se
a
presença das seguintes variáveis ambientais, num raio de 10 m para cada túnel:Tabela 2: Lista de variáveis ambientais consideradas na análise
Variáveis ambientais Código
Jvluro Rochas Lixo Altitude até 100m Altitude até 300m Altitude até 500m Água nas imediações
Espécies de fruto comestíveis Espécies endémícas
Espécies Exóticas Distância à vila
Distância à estrada mais próxima M R L 4100 4300 4500 A
c
ED EX DISTVIL DISTROAD 15A
quantificaçãoda
importânciadas
variáveisfoi
feita
recorrendoao
software R2.g.2
(R
DevelopmentCore Team
2OOg)pela
análiseda
média
de
precisão do package Random Forest (adiante reÍerido como RF) (Liaw & Wiener 2002).O RF consiste num elevado número de "árvores" aleatoriamente construídas, cada
uma referente
a
uma classe (Breiman 2001). Cada árvoreé
construída usando umaamostra
bootstrape
variáveis aleatoriamente seleccionadaspara
dividirda
melhorforma cada
nó. Os
modelos algorítmicos, comoo
RF,
apresentam vantagenS emrelação
aos
modelosde
dados, pois fornecemuma
maior precisão,ou
seja menorerro,
e
melhor inÍormaçãoa
nível dos mecanismos subjacentes aos dados (Breiman2001), além
disso pode
incluir inúmeras variáveise
quantificara
sua
importância. Optou-se por este tipode
análise, em vez dos convencionais métodosde
regressão logística, poiso
númerode
variáveisem
questãoera
muito elevadoe
comotal
os métodos convencionais de análise estatística não teriam potencialidade para prever de forma segura a distribuição da presença de ratos (Oppel, comun.pess).Deste modo, fez-se uma matriz
de 23
(habitats+
variáveis ambientais)por
1 10(túneis), com a presença/ausência de ratos. Realizou-se
o
mesmo tipo de matriz com a presença/ausência de murganhos.Em
relaçãoà
análiseda
médiade
precisãopelo
métodoRF, para
aceder à importância de cada uma das variáveis preditivas, os valores da variável em questãosão permutados aleatoriamente com as observações ouÍlier,
e
depoisos
dados dosoutliers
são
corridos nas árvores para obter novas predições.A
diferença entre asduas
predições,ou seja
entre
as dos
dados outliers
modiÍicadose
os
originais, divididos pelo erro padrão, é a medida de importância da variável (Cutler et al.2007\.As
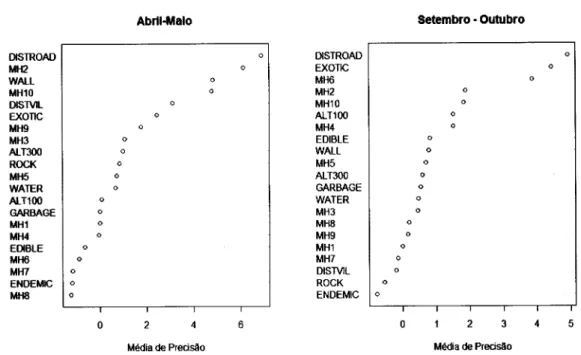
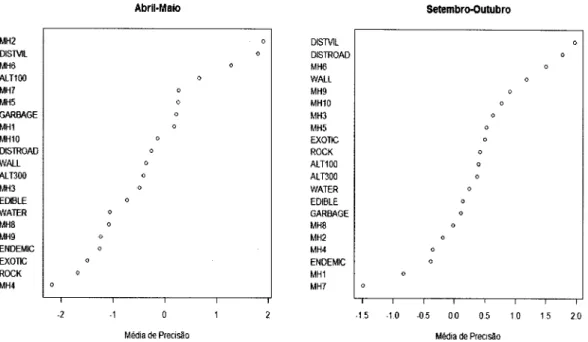
variáveis
ambientaismais
importantese
que foram
expressas graficamenterecorrendo
ao
R
2.9.2são
aquelas cujo valorda
divisãoda
médiade
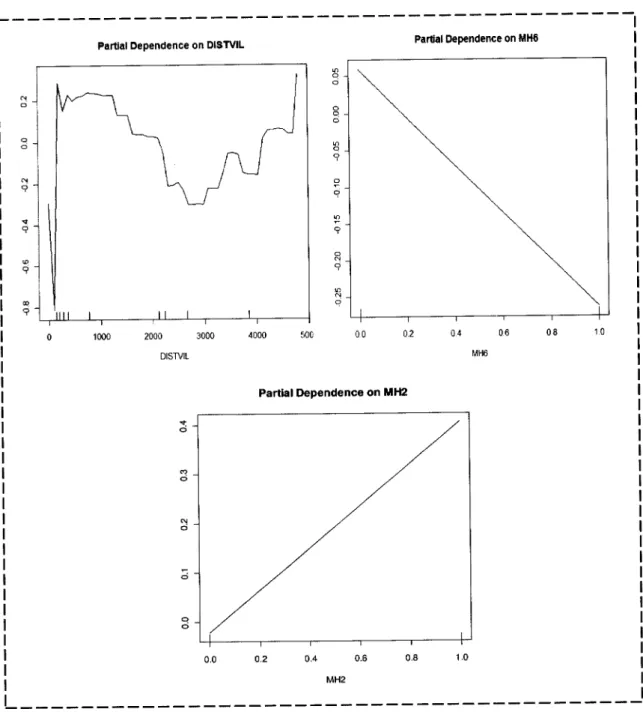
precisão da variável pela da variável mais elevada foram maiores que 0.7.Para a representação gráÍica utilizou-se gráficos de dependência parcial (Hastie et
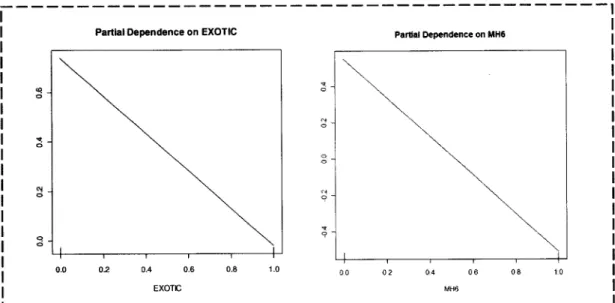
at.
2OO1) que podem ser utilizados para caraclerizar graÍicamenteas
relações entrevariáveis individuais ambientais
e
probabilidades previstasde
presençada
espécieobtidas
pelo
RF.
As
probabilidades (eixodo y)
foram
transformadasde
Íorma
aremover restrições
a
nívelda
amplitudefiá
que as
classiÍicaçõessão
binárias)e
atransformação
é
modelada como uma função linear das covariáveis: transÍormação logit. Deste modo quando as probabilidades se aproximam de zeÍo o logit aproxima-sede
(-)
e no outro extremo quando a probabilidade de aproxima de 1 o logit tende para (+) e quando a probabilidade é de 0.5 então o logit é 0'Nova identificacão de habitats
Apesar
de
nãoter
sido considerado no estudo fez-se uma análise de fotografiasaéreas
para uma
melhor definiçãodos tipos
de
habitat existentesno
Corvo, comrecurso
ao
programa ArcGis
e
carta
militar
do
Corvo
UTM
25N. Esta
análisepossibilitará mais tarde uma extrapolação dos dados obtidos, conferindo uma maior importância a este trabalho e resultando numa aplicação imediata do mesmo.
I
Monitorização
da
época
reprodutora
dos
cagarros
e
análise
do
sucesso reprodutor
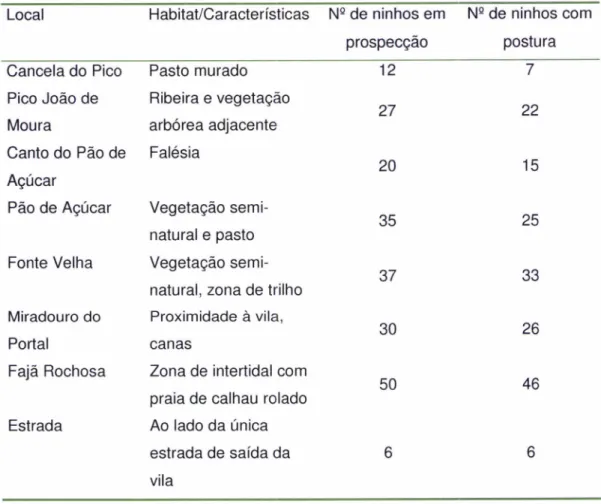
ldentiÍicacão e delimitacão dos locais de nidiÍicacão de caqarros acessíveis da ilha: Foram realizadas escutas nocturnas no período de (Abril-Maio) para identificação
de
áreas
de
prospecçãode
aves
adultas,
para
posterior procura
nos
Iugares acessíveis da ilha.Definiu-se a
piorique
a presença de adulto/casal ou, no caso de não ser visível, oseu canto característico no ninho
e
penas e/ou dejectosà
entrada de uma cavidade adequada constituiria um ninho potencial a ser utilizado na época reprodutora de 2009. Todos os ninhos encontrados foram marcados com GPS e numerados de Íorma visível perto da entrada do ninho com spray acrílico para facilitar a sua identificação. Tentou obter-se uma amostra de cerca de 150 a 200 ninhos (Ver Anexo 7).Depois de numerados os ninhos e divididos em áreas de amostragem, cada zona foi veriÍicada uma vez por semana, desde 25 de Maio
a
16 de Outubro. Neste períodoforam
efectuadas21
monitorizações completasa
todas
as
áreas.
Nos
8
locais identificados, em cada monitorização semanal foi registada a presença de adulto, ovoou cria,
e
no caso de falha, sempre que possível, foram apuradas as causas atravésdos
sinais presentesno
local. Foi classificado como predaçáo:-
ovo
-
pedaços decasca dentro
ou
à
porta
do
ninho;
-
cria
-
desaparecimentoda cria,
ou
morta dentro/porta do ninho (lgual et al.2006). O número de causas naturais foi consideradoinsignificante, apenas
dois
ninhos
com ovos
abandonados.Apesar
de
se
terem instalado duas câmaras com infravermelhos e detecção de movimento em dois ninhos escolhidos aleatoriamente, não foram obtidas imagens de predação.Análise dos dados
Para os ninhos ocupados em que se veriÍicou postura, foram registados os seguintes parâmetros e os cálculos realizados numa folha de cálculo do Microsoft Excel.
Data de oostura
-
data média em que foi colocado o ovoPeríodo médio
de
incubacão-
número médio de dias desde queÍoi
colocado oovo até à eclosão da cria
Data de eclosão
-
data média do nascimento da cria Data de saída do ninho-
data média de saída do ninhoSucesso reorodutor
-O sucesso reprodutor que se considera o estudo da sobrevivência do ninho ("nest
survival)
é
uma componente crítica no estudoda
época reprodutoradas
avese
édefinido como
a
proporçãode
ninhos que obtiveram sucesso numa amostra. Muitasvezes está positivamente enviesada relativamente à verdadeira "nest survival" devido
às
perdas iniciais nas Íases iniciaisda
incubação (Mayfield 1961). Assim para uma correcta obtenção do "nest survival" é necessário seguir o ninho logo desde o primeiro dia de incubaçáo.Para o cálculo do sucesso reprodutor, utilizou-se o modelo de Mayfield modificado de acordo com a fórmula:
tr
Ne de ninhos cujo iuvenilfoicriado com sucessoNe
totalde
ninhos com posturaque pode ser expresso em percentagem.
No caso dos cagarros o sucesso reprodutor está dependente de dois períodos: o
de incubação
e
o de alimentaçáo da cria. Um dos problemas com este cálculo é que parte-se do pressuposto que a "nest survival" diária, neste caso semanal, é constante no tempo e que a data de falha (ovo/cria) é conhecida com exactidão (Dinsmore eÍ a/. 2002), daí, para ultrapassar esta limitação, no capítulolll,
utilizámos outra forma para o cálculo do "nest survival", através do programa MARK.Analisou-se também o sucesso de eclosão, que corresponde à proporção de ovos
que eclodíram com sucesso, e sucesso de saída de crias dos ninhos
-
proporção de crias que saíram do ninho em relação ao número de ovos que eclodiram.Tendência pooulacional das populacões de caoarros
Para
a
análiseda
tendência populacionalpara
os
cagarrosna
ilha
do
Corvo utilizaram-se os seguintes parâmetros populacionais, em que o sucesso reprodutor é overificado
na
ilha do
Corvo
e a
sobrevivênciado
adulto
da
espécie
Calonectris diomedea borealisna
Selvagem Grande,já
que
é
das
colóniasde
cagarrosque
éseguida
há
mais tempoe
da qual se
dispõede
dados demográficos correctos. Osoutros
parâmetrossão
da
subespécie
Calonectrisdiomedea
diomedea, obtidos através do contacto com um investigador do RSPB (Royal Society for the Protectionof
Birds)
que
Íoram
considerados válidose
bastante aproximadosaos
reais para
asubespécie Atlântica em estudo.
r
Sucesso reprodutor : método de Mayfield (Corvo 2009)t
Sobrevivência do adulto: 0.92 (Mougin 1998)r
Sobrevivência do Juvenil (0-3 anos): 0.75 (GeofÍ, RSPB)I
Sobrevivência de imaturo (3-6 anos): 0.905 (Geoff, RSPB)r
Proporção de reprodutores com 4 anos: 0.04 (Geoff, RSPBr
Proporção de reprodutores com 5 anos: 0.08 (Geoff, RSPB)r
Proporção de reprodutores com 6 anos:O.4 (Geoff, RSPB)r
Proporçáo de reprodutores com 7 anos: 0.68 (Geoff, RSPB)r
Proporçáo de reprodutores comI
anos: 0.88 (Geoff, RSPB)I
Proporção de reprodutores com +8 anos: 0.96 (Geoff, RSPB)A
tabela de vidafoi
realizada recorrendoao
pacote poptoolsdo
Microsoft Excel, com base na matriz de Leslie construída com os dados anteriores.lll. lmpacto dos roedores
na
época reprodutora
Dinsmore
et
al.
(2002) desenvolveuum
modelopara
o
"nest
survival,,para
oprograma MARK (White & Burnham 1999), que é basicamente uma generalização do
estimador
de
Bart&
Robson (1982), que permiteao
utilizador desenvolver modelossobre
os
dados originaise
rigorosamente avaliaro
seu
suporte usando "Akaike's information criterion" (AlC). Este programa permite ao utilizador avaliara
importânciade factores ambientais que influenciam
o
"nest survival", podendo incluir covariantes individuaise
de
grupo,
específicasdo
tempo.Aqui,
a
sobrevivênciade um
ninho refere-seà
probabilidadede
um ninho sobreviverum
intervalo de tempo especifico, normalmente um dia, mas para o nosso caso uma semana.Os pressupostos do modelo "nest survival" do MARK são: (1) aos ninhos pode-se
atribuir
uma
idade correcta quandosão
encontrados;(2)
o
destinodos
ninhos écorrectamente determinado, 1=sucêsso/0=insucesso; (3)
a
descobertado
ninhoe
as monitorizaçÕesnão
influenciam
a
sobrevivência;(4)
o
destino
dos
ninhos
éindependente;
e
(5) as
taxas diáriasda
sobrevivênciado
ninhosão
homogéneas. (Dinsmore et al.2OO2)O
tipo
de
ficheiroque
vai ser
analisadotem
de
começarcom: "Nest
SurvivalGroup=1", seguido
dos
dados
dos
ninhospara
este grupo.
A
cada
ninho
estão associados 5 valores: (1) o dia em que o ninho foi encontrado, (2) o último dia em queo ninho foi verificado vivo, (3) o último dia em que o ninho foi verificado, (4) o destino
do
ninho (0=suc€sso /'l=predado)e
(5)o
número (frequência)de
ninhos com esta história. Depois destas informação podem adicionar-se covariáveis individuais.O programa, utiliza estas informaçôes para gerar uma história de encontros para
cada ninho