Lethal concentration of methyl methanesulfonate in a new potential invertebrate model for ecotoxicology
Texto
Imagem
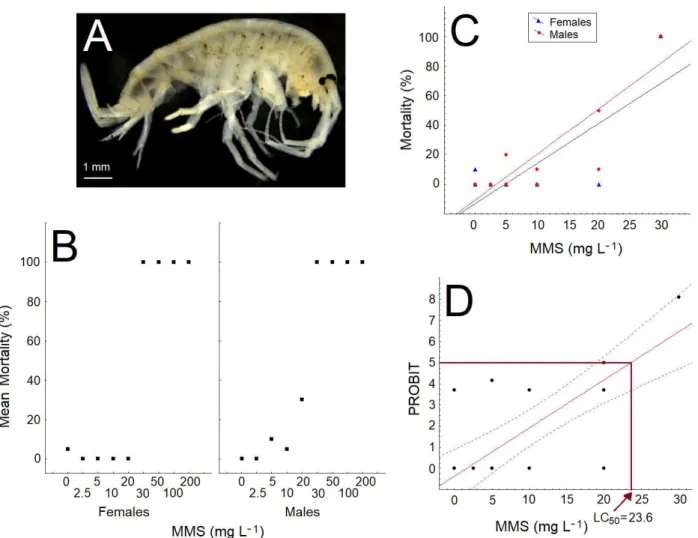
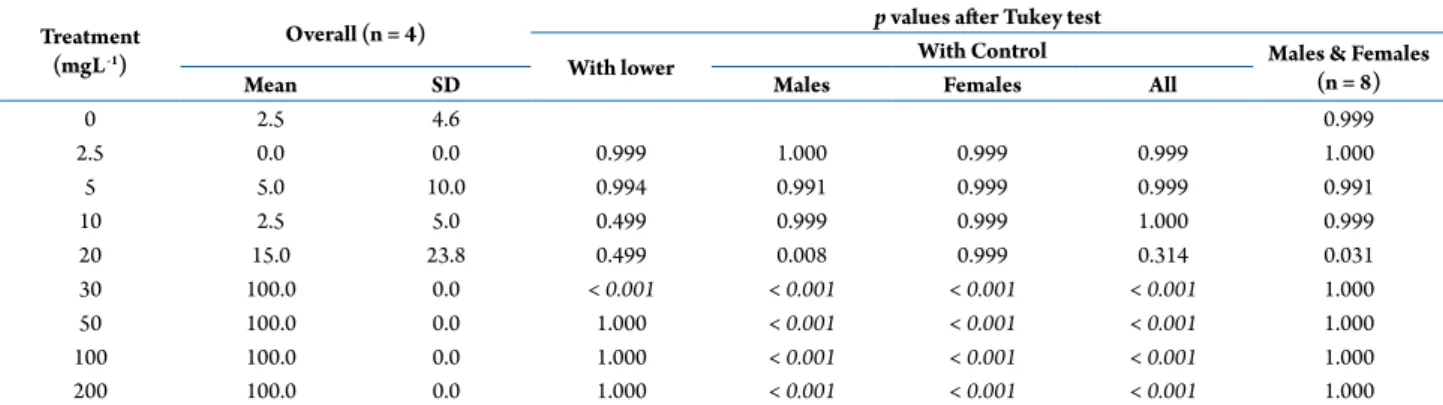
Documentos relacionados
Effects of polydatin on cell apoptosis in liver cancer cell lines treated with different concentrations of polydatin for 48 h (A), determined by fl ow cytometry assay; B, effect of
En la conciencia intrasilábica, se pudo observar que los niños se encuentran por debajo del 50% de desarrollo de este nivel, esto se debe a que requiere mayor
Considerando que os teores de macronutrientes no solo dos três sítios não foram diferentes e que as plantas de erva-mate eram da mesma procedência, conclui-se que a luminosidade foi
Strain C-10 and all selenate resistant mutants were poured into MMS - containing 10 mg.L -1 of D.L–methionine and different concentrations of chromate, in order to detect
At this concentration of LAS-C12, the metabolism tended to increase with the time of exposure, and after 48 h the increase was statistically significant in relation to the
Comparison between 共a兲 steady-state PL upon He–Cd excitation and 共b兲 time-resolved spectra performed with above-band-gap excitation for a time delay of 0.02 ms and time window
90 Figura 4-14 Resultados de variação de massa em envelhecimento húmido em estufa – provetes de escala piloto alta pressão (PE_D27) .... 90 Figura 4-15 Resultados do coeficiente
There are three main contributions of this study to the literature: (i) a business re- lationship network model; (ii) a business community detection algorithm; and (iii) a
