Effects
of domain connection and
disconnection
on
the
yields
of
in-plane bimolecular reactions in membranes
Eurico C. C. Melo, * Isabel M. G. Lourtie,* Mantripragada B. Sankaram,§Thomas E.Thompson,§ andWinchil L. C.
VazI
*Centro deTecnologia
Quimica
eBiol6gica-l.S.T.,
Oeiras, Portugal;tCentrodeAn6liseeProcessamentodeSinais,Complexo1, I.S.T., Lisboa, Portugal; §Department of Biochemistry, University ofVirginia,Charlottesville,Virginia22908, USA;andlUnidadedeCidnciasExactaseHumanas,Universidade doAlgarve,Faro, Portugal
ABSTRACT It hasrecently been shown(Vaz,W. L.C.,E.C.C. Melo,and T. E.Thompson.1989.Biophys.J.56:869-875;1990.Biophys. J. 58:273-275) that in lipid bilayer membranes in which ordered and disordered phases coexist, the ordered phase can form a two-dimensional reticular structure that subdivides the coexisting disordered phaseintoa disconnected domain structure. Herewe
considertheoreticallytheyields ofbimolecular reactions between membrane-localized reactants, whenboth thereactantsandproducts areconfinedtothedisorderedphase.Itis shown thatcompartmentalization ofreactantsin disconnecteddomainscanleadtosignificant reductions in reaction yields. The reduction in yield was calculated for classical bimolecular processesand for enzyme-catalyzed reactions. Theseideas can be used to explain certain experimental observations.
INTRODUCTION
Biological membranes are made up ofa lipid bilayer, andintegralandsurface-attachedproteins.The
lipid
bi-layer is viewed as a quasi-two-dimensional fluid-likesheet inwhich lateral diffusion and redistribution
ofcom-ponents occurs
(Singer
andNicholson,
1972). Thislat-eral diffusion permits molecular reactions and interac-tions ofphysiological importancetooccurintheplane of
amembranesuchasthemitochondrial innermembrane
(Lenaz, 1988; Chazotte and Hackenbrock, 1989;
Rajar-athnam etal., 1989).
The lipidbilayer is a complex mixture of
lipids
thatmay varysignificantly from eachotherintheir chemical
and
physical
properties. Insuchmixtures,
phasesepara-tionsleadingtodomain formationare
possible
(WuandMcConnell, 1975; Lee, 1977; Tocanne et al., 1989). Phase separation is dependentupon temperature, pres-sure, chemical
composition,
and for membranes with chargedlipids,
alsouponthe pHandionic strength ofthe aqueous medium. Phaseseparation implies
nonhomo-geneous componentdistributionsnot only of thelipids but also of the membrane
proteins,
which mayprefer
onephaseoveranother.Forthe caseofreacting protein
or
lipid
species, nonhomogeneous distributionscan sig-nificantly affect reaction yields.Wehaverecently shown (Vaz etal., 1989, 1990; Bult-mann etal., 1991) that membranes in which ordered and disordered phases coexisthave adomain structure
in whichasmallmassfractionof ordered phase formsa
reticulum that subdividesthedisorderedphaseinto dis-connecteddomains.Ithas beenshown(Vazetal., 1989; Bultmannetal., 1991)that the massfraction ofthe or-deredphaserequiredtoachieve this subdivisioncanbe
aslowas20% of thesystem.
Address correspondenceto Dr.Thomas E.Thompson, Department of
Biochemistry,Box440MedicalCenter,UniversityofVirginia, Charlottesville, VA22908.
Here weconsiderspecificallytheeffects of reticulation
uponthe reactionyieldof bimolecular reactions
involv-ing membrane-boundreactants.Fromthestandpoint of biologicalsystems the effects of reticulation are twofold:
(a) if the existence of reticulation is unknown to the
outside observer, measurements ofthe reaction system can lead toincorrectthermodynamicparameters;(b) if the cell can by metabolic means movethe membrane domain structure back and forth across thepercolation threshold to connect or disconnect a particular phase,
the cell can control the extent of membrane-localized reactionsconfined to that phase. The general problem
concerns reactions occurring in a compartmentalized
spaceandis,inprinciple, alsoapplicabletoreactions in systems with phase separations involving disordered
phases only, butwithanabsolutesolubility preference of the reactants for one phase overthe other(Hatlee and Kozack, 1980).
Recentlywehaveexamined fromatheoretical
stand-point
the effect ofcompartmentalization
on amem-brane-confined homodimerizationreaction(Thompson
et al., 1992). The ideas developed in this paper have beenappliedinanexperimental analysis ofthe
concen-tration dependence of electron spin resonance (ESR) line shapes ofaphosphatidylcholine spin labelin two-component, two-phase phosphatidylcholine bilayers (Sankarametal., 1992). Inthe present
manuscript,
weextend thetheoretical
analysis
toincludemorecomplex in-plane reaction systems.THEORY
In general when reactants are randomly distributed over a compart-mentalized system, the product yieldwillbe lower than when the
reac-tants aredispersed in a continuous system. The yield decreases as the average numberofreactantmolecules per compartment decreases. As
ameasureof theextenttowhich a reaction is hindered by reticulation of the membrane, let us define the expected relative yield,
(1)
1506 0006-3495/92/12/1506/07 $2.00 Biophys. J. ©BiophysicalSociety
Volume63 December 1992 1506-1512
(
(P>
= (Iklet>(,tcont
>
astheratio oftheyield expectedtobe achieved in the reticulated
mem-brane,(t,,t>,totheyield expectedtobe achieved inacontinuousone,
<D,t>.Theyieldornetyield,
<(>
is definedasthefinal number of product moleculespercompartment/the initial number ofreactantmoleculespercompartment.
Wemodelthe reticulated membraneasapatternoftwo-dimensional fluid domains that,regardless ofshape,have thesame area.Moreover,
weconsider that the barriers between domainscannotbecrossedbythe
reactantmoleculessothat thereisnoexchangeofreactantsbetween
reactioncompartments. Forthe sake ofsimplicity,weassumepoint molecules. We makeafurthersimplifying assumptionthat the
reac-tionsarenotreversible sothat the chemicalprocesseswithina
com-partmentproceeduntilatleastoneof thereactantsisfullyconsumed.
Itmustbe stressed thatwedonotconsider the kineticsof the reactions
butonlytheiryield,sothat neither the dimensionnortheshapeof the
domains (two-dimensional vessels) needstobe taken into account.
The lifetime of disconnecteddomains is assumedtobe muchlonger than the timerequiredtoattaincompletereaction.
Inpractice,wedeal withdispersions of cellsorlipid vesiclesin which
the reaction spaceis the lipid bilayermembrane. Both are apriori
compartmentalizedreaction systems. Reticulation of the membrane
superimposesasecondarycompartment pattern onthisalready
dis-cretereactionspace.Weareconcerned hereonly with the secondary compartmentalizationthat makes thesingle vesicleorcell,whichwe
shallrefertoasthe"unit,"ourpointof reference. We consider that
encountersbetween allreactantmolecules inasinglenonreticulated unitarepossible,whereas in the reticulated unitonlyreactionswithin
singledomains will takeplace.Inbothcases,thenumber,i,ofproduct molecules, P, formedinagivenreactioncompartmentisafunction of
thenumberofreactantmolecules present in that compartment. Itmust
bestressed that all the unitsareassumedequivalent and,intheabsence
ofreticulation,reaction compartments(domains)and unitsare
indis-tinguishable. Since,inourmodel,the reticulum divides each unit into domains ofequalarea, uponreticulation all reactioncompartments
remainequivalent.Inbothcasesthe availablereactantmolecules,R
(R=Rl,R2, * -),aredistributed randomlyinthe reaction
compart-ments.Asaconsequence,theobservedyieldis themeanvalue taken
overthe entiresetofcompartments.
Inthehomogeneousmembrane,aconcentration[P]J,tofproduct molecules results after completion ofthe reaction. For the reticulated
casethe final concentration of theproductis[P],,t.Therefore, Eq. 1 maybe rewritten
=(P I r(2)
[P ]<ont(2)
Letusdenote the unit'Uand the domainbyD. Hence, the expected
concentrationofproduct moleculesisgiven by
[P] =
[O1]
z iPp(i), (3)i
wherePp(i) stands for the probability of obtainingiproduct molecules
inadomain.Recalling thatinthenonreticulatedsituationthe domain extendsoverthe entireunit,theexpectedrelativeyieldwhen each unit
is divided intoNdom domainsis
E iPp( i,Ndom)
= Ndom iPp(i,Ndom= 1l) Theprobability
Pp(i,Ndom) ...* 2 P(ilnRl, * nRm)
nRI nRm
X PRIo adRm(hRledis nRmi Ndom) (r)
dependsonboth thekind ofreaction and the distribution of the
reac-tantmolecules R(R=Rl,...,Rm)amongthe domains.InEq.5the
conditional probabilityP(iInR1, ..., nRm)accountsfor the
mecha-nism of thereaction,whilePRI .Rm(nRt ... nAm,Nd&m)represents
thejoint probabilityofhavingrespectivelynRI,...,nRmreactant
mole-cules, Rl,...,Rm, inadomain. Forabimolecularreaction,ifthe
distributions of thereactantmolecules(RlandR2),PRI(nR,,Nd,m)
andPR2(nlR2,Nd,m),areassumedtobemutually independent, Eq.5
becomes
sp dom)
= z
flP(ilnR1,
nR2)PR(l(nR,Ndom)PR2(nR2,
Ndom). (6)nRI nR2
Provided thatalargenumberofreactantmoleculesaredistributedover alargenumber of units withoutcrowding effects,thedistributionof reactantmoleculesoverthe different units followsaPoisson distribu-tion law
An(n)=-e (7)
Themeannumber ofreactantmoleculesperreactioncompartment,U,
is either g = [R]/['U] for the nonreticulated case or ,u = [RI/
(['U]Ndom)when there is reticulation.
Inthenextsection, theaboveformalism is appliedtofourimportant
reactionmechanisms.
CASE STUDIES
We confine ourselves to the study of four cases: (a) a
Michaelis-Menten type enzymatic reaction, (b) a
con-ventionalbimolecularreaction, (c) adimerization
reac-tion,and
(d)
consecutivereactionsconsisting oftwo se-quential Michaelis-Menten type steps.Enzymatic
reaction
Weconsiderareaction inwhichanenzyme Ecatalyzes
thetransformation ofasubstrate R intoa
product
P:E+R -E+P.
While inahomogeneous mediumaminuteamountofE is enoughtocause acomplete conversion of RtoP,ina compartmentalizedmedium the yieldwilldependupon
the probability of having atleast one molecule of E in every compartment. Under certain conditions thiscan lead to a drastic decrease in the yield of P. The condi-tional probability that reaction takes place in a given domain withanetyield of i molecules ofP, given thatnE
molecules ofenzyme and nR molecules ofreactantare
present, is given by i=0 I =nR. (8) nEl=ew
P(ijnE,
nR)= 1 nE* °, O elsewhereSubstitution ofEq. 7 and Eq. 8 in Eq. 6 leads to the
relativeyield exp- [E]N dom, ~~(P> = [E]I I-exp -IVU IJ (9)
Meloetal. Reactionsin Reticulated Membranes 1507W]
It should be notedthat, in this case, the relativeyield,
p>,
doesnotdepend
onthe concentration ofR,
[R].
Inparticular, itrepresentsthe ratio between the probabili-ties offindingatleastonemolecule of Einagiven reac-tion compartment for both the reticulated and the nonreticulatedsituations. Since, in thesecompartments,
all Rmolecules thatcancomeincontactwithanE mole-cule are converted, the expected net yield is, in both cases,proportionaltothemeannumberof Rmolecules percompartment.
Fig. 1 presentsaplot of<
p>
versusNdom. As expected,a sharp decrease in the relative yield of the reaction is
observeduponreticulation when the number ofenzyme moleculesperreactioncompartmentis small. Infact,as a result ofcompartmentalization a large fraction ofR moleculesarenotaccessibletotheenzyme, E.
In additiontotherelative reaction yield, it is also
im-portant toevaluate how thenetyield of Pchanges with [E]inthe reticulated reactionspace. Inthiscasethenet
yield ofPisgiven by (numerator of Eq. 9)
<
(k,,
>
= 1 - exp{-[EUINdom
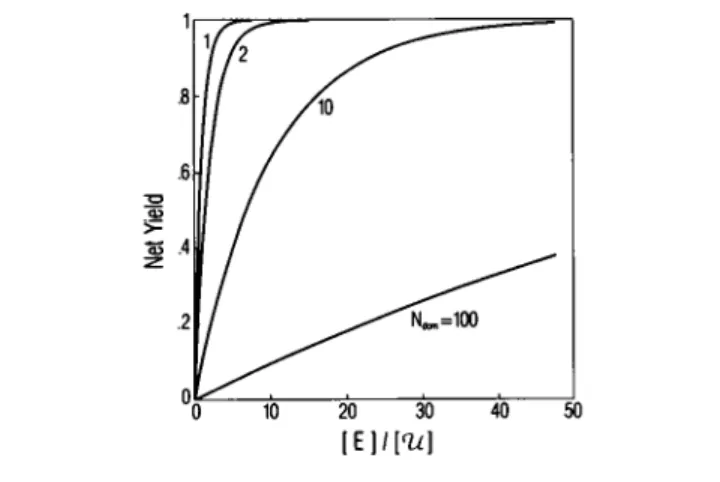
(10)Fig.2presents a plotof
<
¢t>
versus[E]
/
['U]
forseveral statesofreticulation.Thedifferencebetween the curvesis simply a consequence ofthe change in the average valueof[E]in the reaction compartment resulting from
differences in the degree of compartmentalization.
Hence thecurves in Fig. 2 actually represent the same curveunderdifferent concentration scaling.
-a 75
[EI [ilJ
FIGURE 2 Netyield ofacatalyzed reactionas afunction ofthecatalyst concentrationfor values ofNdom= 1, 2, 10, and 100.
P)1 nRInR2l< nRsIee i
P(ij|nRI,snR2)=41 nRI > nR2, nR2 =i.
LO
elsewhere(11)
Asinthe previouscase,the relative yield is obtainedas the ratiobetweentwoprobabilities, i.e.,
1 {
[Rl]
[R2] ((p> = [llNdom [U]Ndom I _f [Rl ] [R2] \[W] 'Vu]
(12) whereCONVENTIONAL BIMOLECULAR REACTION
Inthecaseofthe bimolecular reaction
Rl + R2-P
theconditionalprobability P( i nR1,nR2) that describes
the reaction mechanism isnowgivenby
.8
.4
f(ORI, AR2)= exp[-(ARI +AR2)A
oo nRrl nR2 AnRI-I
RI.
X (nR1 -nR2) (13)
nR1o nR2 0 nR2! nRl.
In Eq.
13,f(ARl,
/'R2) stands for theprobability thatno reactiontakes place in the wholesystem.Fig. 3 shows that fora fixed value of [R2]/[U], the
tXii=20
.8 20 -100~~~~~ra A[~~~~~~~3 NdfOFIGURE 3 Relativeyieldofaconventionalbimolecularreactionasa
function ofcompartmentalization forafixedconcentration ofoneof thereactants.[RlI/[l]issetas20, while [R2]/[lJ]= 1, 10, 20,30,
50,and 100.
B-p--a ounlVlm ee br19
FIGUREi Relativeyield of product formation for a catalyzed reaction
as afunction ofthe number ofdomains per reaction unit. The different
curvescorrespond tovarying catalystconcentrations, [E]/ [ l]= 1,5, 10,and 100.
decreasein reactionyieldofthesimplebimolecular
reac-tion considered here issomewhatlessthan thatobserved for the enzyme catalyzed reaction considered earlier
(Figs. 1 and2). Another result of the calculation is that
the relative reactionyield isnotamonotonicfunctionof [Rl
].
This effect ismoreclearly visiblewhenweanalyze the evolution of thenetreactionyieldI
[R1]
[R2] if[R] [R2][WUNdom [']Ndom' [U]
] 1 _[R2] [R]R] > [R2]
t \[W ] dom [ dom 1 [W] [e ]
(14)
withf(*, *)given in Eq. 13. In Fig. 4 we plot the net
yield,
<k,t>,
withrespect to[Rl1! ['U] foragivenvalue of[R2]
/ [Ml].Weseethatthenetyieldexhibitsa mini-mumat[Rl] =[R2]. The meaningof thisminimumis easytounderstand ifextremecases,andasimplified sce-nario,areconsidered. Ifthe number ofRl moleculesina unit is much smallerthan the number ofR2 molecules init, theprobability thatuponreticulationanRlmolecule is in thepresenceofatleastoneR2molecule in thesame
compartmentisrelatively high. Conversely,ifthe num-ber of Rl molecules per unit is much larger than the
number of R2molecules, uponreticulationalarge
frac-tionof theR2moleculesareexpectedto react.Ifthe unit
ispopulated withanequalnumber ofRl and R2
mole-cules,inthe absenceofreticulationreaction isexpected
togotocompletion.However,reticulationwillresult in
acertain fraction of reaction compartments (domains) which have unequal number of Rl andR2moleculesso that the net result will be a significant fraction of unreactedreactants. -o C) [Rli
[1,1
Dimerization reaction
Herewedealnotonlywiththesimple bimolecular reac-tion
2R R2
butwealsocompareittothemoregeneralcaseof poly-merformation reactions for thecasesof aggregation of3 and 4monomers
Rk (k=2,3,and4).
If,inhomogeneousmedia,adimerizationreaction pro-ceeds until total consumption of the monomer, upon
reticulationeveryunpaired molecule will be lost forthe process,withaconcomitantdecrease in yield. This drop inyieldis stillmoremarked in thecaseoftheformation of trimersortetramers.
The conditional probability that the reaction takes place inagivendomain withanetyield of i molecules of Rk, given thatnRmoleculesarepresent, isgiven by 'P(ilunR) 1 i=nR/k, 1 i=(nR- 1)/k, 1 i=(nR-k+ 1)/k, 0 elsewhere nR
multiple
of k (nR- 1) multipleofk (nR-k + 1) multiple of k (15)Taking into consideration that, for these reactions, all
theinvolved molecules pertaintothesamereactant,the relative yield is 1_ n( [R] ) (4P>= (16) where oo k-1 (kjl+j2-1 )
f(AR) = exp(-AR) . 12 kji+j2)! ijI-O j2-0 (kj1 +12)!
(17) In Fig. 5 wepresentplots ofthe relative yield of
forma-tion ofdimer, trimer andtetramerforasingle
concentra-tion ofR,
[R]/[UJ]
= 20,as afunction ofthe number of domains. Thedifferences in yield for dimers andhigher aggregation numbersareclearlyobserved.From the study of the relative yield we canfind the
extentofhindrance resulting from membrane reticula-tion. The primary reticulation effect, which is always
presentinvesiclesorcellsuspensions, is better dealt with through examination of thenetyield of the reaction
(18)
< t
>
= I f[Rt[]N&m
J)
ReactionsinReticulatedMembranes 1509
FIGURE 4 Netreactionyieldforaconventional bimolecular reaction asafunctionof[RI/[U],for[R2]/[l/J =20,andNd&m=1,5,10,20,
50, and 100.
40 60
Nd,,
FIGURE5 Relativeyieldofadimerizationreaction(k=2)for [R]/ [U]=20 as a functionof the number of domains per unit, Ndom. Plots arealsopresented for thecaseof trimer(k=3) and tetramer(k=4) formation.
withf(*) given in Eq. 17. The netyieldasfunction of
[R]/['U]
for k=2, and 4, is plotted in Figs. 6, A and B.Consecutive reactions
We consider two cases ofsequential enzyme-catalyzed reactions:(a) the action ofanenzyme on apro-enzyme
to produce a second enzyme that catalyzes the subse-quentreaction inthe sequence, and(b) the product of
thefirstenzymaticreaction is thesubstrateof the second
enzymatic reaction in the sequence. We consider the
formercase first:
El + Rl El + E2 E2+ R2 E2 + P
where El and E2arethe catalysts. Itcanbe shown that, whether thesystemis reticulatedornot,thenetyield of Pis thejoint probability of havingatleast one El and oneRl moleculeinthe smallest reactioncompartment.
Therefore, if the distributions of all reactants are as-sumedtobeindependent, from Eq. 1, the relative yield is
(1-exp{- [ex]N-})(l P{ [R]
(
[El>
1 exp [Rl](19)
Notingthatthe concentrations of Rl and E2 areequal whenthe first reactionstepis performed withprobability
one, the above expression is theproduct of the relative yields for both reactions.
In Fig. 7, we comparethe singlestep catalyzed
reac-tion with the consecutive mechanism, for the case of
[El]/[U
] = 10.The consecutive reaction is performedatthreedifferentmean occupancynumbers for Rl. As expected, when the concentrations of the two starting
reactantsareverydifferent,oneof thestepsbecomes the
bottleneck of theglobal mechanism. But when the
con-centrationsofEl andRlarecomparable, the decrease in yield dependsonthe concentration of both theenzyme and thereactant.
Forthe secondcaseof the consecutive reactions men-tionedabove,weconsiderthefollowingsequence
El + Rl El + P1 E2+ P1 E2+ P2.
Inthiscasethenetyield ofP2is thejoint probability of havingatleastoneElandoneE2 molecule in the
small-estreactioncompartment. Again, recalling that the dis-tributions of all reactantsare independent, from Eq. 1, the relativeyield is given by
[El] I[E2]
1 exp-[- ]Ndom }) 1 exp-[- ]NdO )
-P(>
-expj- [EU]l)(1 eP [E2] 1)(20)
Both Eqs. 19 and 20areformally identical. The relative yield ofthe final product is, for bothcases,theproduct of
-o a) -o 5: [R1 [RI E1,7
FIGURE 6 Representationof the net yield as a function of the mean numberofRmolecules perunit for different number ofcompartments perunit. (A) thecaseofdimerformation. (B) the case of tetramer formation.
151- ipyia ora oue6 eebr19
.N
-a)
cs
NdO,
FIGURE7 Relative yields for the finalproduct oftwoconsecutive
cata-lyzedreactionsas afunctionof Ndom with [El]/[ l(]= 10.Theyields
shownareidentical fortwodifferent types of reaction sequences(see
textof Eqs. 14and 15).XrepresentsRlinthefirstcase(Eq. 14)and E2in the secondcase(Eq. 15). Curvesaregivenfor[X]/['1]= 1,10,
and100. Forreference,thecorrespondingcurvesforasinglestep
cata-lyzed reactionwith [E]/['U] = 10 and 100 are also shown(broken lines).
two relative yields. However, theyrepresent two physi-cally distinct situations. The relative yield of the final productas afunction of number of domains forafixed meanvalue of Elperdomain is identical for Eqs. 19 and 20, and given inFig. 7.
DISCUSSION
We have demonstrated that the reaction yield of four typesofbimolecular reactions confinedto a membrane
surface is decreased by compartmentalization ofthe
sur-face forarandomdistributionofreactants over the com-partments. Theyield is highlydependent upon both the reactionmechanismand the reactantconcentrations.
Inthe case ofthe enzyme catalyzedreactionit iseasy tounderstandthat, since onlyvery small amountsofthe enzymeneed be present,compartmentalizationcanlead
to a very low average enzyme concentration per com-partment, even ifthesize ofthe compartments is
rela-tively large. This is the situation depicted onthe
right-handsideofthe curvesinFig. 1.Thus,the consequence
ofcompartmentalization foranenzymatically catalyzed reaction inabiologicalmembraneisamarkedinhibition ofthe processregardless ofcompartmentsize.When the average numberofenzyme molecules per compartment
isless thanfivethereactionisessentiallyshutoff. Similar
but more dramatic effectsare seen for consecutive en-zymecatalyzed reactions shown inFig. 7.
For a bimolecular reaction involving two different
reactantstheeffect of compartmentalizationonthe rela-tiveyield,asshown inFig. 3, isnot asgreat asit is foran
enzyme catalyzed reaction. The yield is, however, a
rathercomplexfunctionoftheconcentrationsofthe two reactants, as shown in Fig. 4. In the special case of a
single
homodimerizationreaction,
the decrease in therelative yield observeduponcompartmentalization
origi-natesonlyinanincrease ofthe number of molecules left over in domains with an odd number of molecules. When theformation ofeither trimersortetramersis con-sidered, themean number of molecules leftoverper
do-mainincreases, leadingtoafurther decrease in the rela-tive yield (see Fig. 5). A similar argument can also be
appliedtotheanalysis ofthenetyield,asdepicted inFig.
6. The fraction of unreacted molecules decreases as a consequence of the increase in the
[R]
/ ['U]. Thisbe-comes moreevident in thesigmoidal shapeofcurvesin Fig. 6 B. In fact, forverysmall values of
[R]
/ [U],
the probability of havingatleast k molecules inasingle do-main isverylow, being much smaller for k=4 than fork=2.Therefore,anearlyzeronetyield is observed when [R]/['U]isverylow.
The arguments presented in this paper have been given in thecontextofcompartmentalizationas aresult ofchanges of the phasestructureofalipidbilayer
mem-brane in which the reactants are confined. It is clear, however, that the conclusions are independent ofthe means of compartmentalization, which in biological membranes could include compartmentalization by meansof proteincomponents(Freire andSnyder, 1982) andcytoskeletalstructures(Saxton, 1990)aswellasfor hypothetical situations in which proteins are separated from each other by preferential solubilizationin phase-separated domainsinthesamemembrane. We havealso limitedourconsiderationstoreactions confinedto
two-dimensionalsystems.Thesametreatmentalsoappliesto
compartmentalized three-dimensional systems and the conclusionsare,in everyaspect,similar.
Recently the basic ideas developed in thispaperhave been usedto understand experimental results obtained in two unrelated studies. In the first of these, the line broadeningofaspin labeledphosphatidylcholine dueto
spin-spin interactions between labels wasexamined by ESRspectroscopy in bilayers formed from mixtures of dimyristoyl phosphatidylcholine and distearoyl phos-phatidylcholine as afunction oftemperatureand com-position(Sankarametal., 1992). The detailednatureof theline broadeningintheregion of coexistence of fluid andgel phasescanbeunderstoodquantitatively only by taking into account the compartmentalization of the spin-labeled phospholipidincertainportions of the
two-phase region. Thisstudy providesadirect experimental
testof thebasicideas outlinedabove. Inthe second, the difference between the apparentequilibrium poise ofa reactionindisconnected, ascomparedtoconnected
sys-tems,hasprovidedanexplanation forananomalous situ-ation observedinphotosyntheticsystems(Lavergne and Joliot, 1991). The anomaly concerns the difference in equilibrium constantdetermined under lowlight inten-sity and theconstants determined using artificial redox
carriers. Lavergne and co-workers explain the difference
as being due to the attainment ofglobal equilibrium
among the subsystems of each chloroplast through the
Melo et al. Reactionsin ReticulatedMembranes
useofartificialredoxmediators,whereasatlow
illumina-tionintensity each globalequilibrium subsystem attains
its own equilibrium. The ideas that lead to thesuccessful resolution of the anomalous behavior ofphotosystems
areconceptuallyanalogous to theideas developedin this paperconcerning the effects of in-plane connectionand
disconnection onmembrane-localized reactions.
Although not directly relevanttotheproblem of
do-mainstructures inbiological membranes,much very
in-teresting workon thestructureofphases has beendone ontwo-componentmonolayersat anair/waterinterface
(for a review, seeMcConnell, 1991). The large dipole
momentassociated with thesemonolayers
gives
rise torelatively large domains, whichareusually arranged ina regular two-dimensional super lattice. Frequently,
do-main shapes exhibit symmetry properties arising from
the
chirality
ofcomponentmolecules. Structuresof thistypeandsizeareabsent in
bilayers,
presumably becausethereisalmostcompletecompensation ofthedipole mo-mentofoneofthe monolayers bythedipole ofthe other monolayercomprisingthebilayer.Thediscussion ofthe effectsof connection and disconnectionon
bilayer-con-fined reactions outlined above can, of course, be readily
appliedtoreactionsconfinedtomultiphasic monolayers
attheair/water interface.
This research was partially supported by the Junta Nacional de
Ivesti-ga,aoCientifica eTechnologica, Portugal, under contract no. PMCT/ CEN658/90, and by the National Institutes of Health, USA, under
grantsGM-14628andGM-23573.
Receivedfor publication 18 May 1992 and in revisedform 25August 1992.
REFERENCES
Bultmann, T., W. L. C. Vaz, E. C. C. Melo, R. B. Sisk, and T. E. Thompson. 1991. Fluid-phase connectivity and translational diffu-sion in aeutectic, two-component, two-phasephosphatidylcholine bilayer. Biochemistry. 30:5573-5579.
Chazotte, B., and C. R. Hackenbrock. 1989. Lateral diffusion asa
rate-limiting step in ubiquinone-mediated electron transport. J.
Biol. Chem. 264:4978-4985.
Freire, E., and B. Snyder. 1982. Quantitative characterization of the lateraldistribution of membraneproteinswithin thelipidbilayer. Biophys. J. 37:617-624.
Hatlee, M. D., and J. J. Kozak. 1980. Astochastic approachtothe theory ofintramicellar kinetics. I. Masterequationfor irreversible processes. J.Chem.Phys.72:4358-4367.
Lavergne, J., and P. Joliot. 1991.Restricted diffusion in photosynthetic membranes. TrendsBiochem.Sci. 16:129-134.
Lee, A. G. 1977.Lipid phase transitions andphasediagramsI.Lipid
phasetransitions. Biochim.Biophys.Acta.472:237-281.
Lenaz,G. 1988. Roleof mobilityof redox components in the inner mitochondrial membrane.J.Membr. Biol. 104:193-209.
McConnell, H. M. 1991.Structures andtransitions inlipidmonolayers attheair-water interface. Annu. Rev.Phys. Chem. 42:171-195. Rajarathnam, K., J. Hochman, M.Schindler,and S. Ferguson-Miller.
1989. Synthesis, location and lateral mobility offluorescently la-beledubiquinonelOinmitochondrial and artificial membrane.
Bio-chemistry.28:3168-3176.
Sankaram, M.B., D.Marsh, and T.E.Thompson. 1992. Determina-tionof fluid and gel domain sizes in two-component, two-phase lipid
bilayers:AnESRspin label study.Biophys.J.63:340-349. Saxton, M. J. 1990. The membrane skeleton oferythrocytes.A
perco-lation model.Biophys.J.57:1167-1177.
Singer,S. J., and G. L.Nicholson. 1972. Thefluidmosaic modelofthe
structureof cell membranes. Science(Wash. DC). 175:720-731. Thompson,T.E., M. B.Sankaram, and R.L.Biltonen. 1992.
Biologi-cal membrane domains. Functional significance. CommentsMol. Cell.Biophys.8:1-15.
Tocanne, J. F., L.Dupou-Cezanne, A. Lopez, and J. F.Tournier.1989. Lipid lateral diffusion and membrane organization. FEBS (Fed.
Eur.Biochem.Soc.)Lett.257:10-16.
Vaz, W. L. C., E. C. C.Melo, and T. E.Thompson. 1989. Translational diffusion andfluiddomainconnectivityinacomponent, two-phasephospholipidbilayer.Biophys.J. 56:869-876.
Vaz, W. L.C., E. C. C. Melo, and T. E.Thompson. 1990. Fluid phase
connectivityinan isomorphous,two-component, two-phase phos-phatidylcholinebilayer. Biophys.J. 58:273-275.
Wu,S. H., and H. M. McConnell. 1975. Phaseseparations in phospho-lipid membranes.Biochemistry. 14:847-854.
![FIGURE 4 Net reaction yield for a conventional bimolecular reaction asafunction of[RI/ [U], for[R2]/[l/J = 20, andNd&m = 1,5, 10,20, 50, and 100.](https://thumb-eu.123doks.com/thumbv2/123dok_br/18602834.909396/4.918.480.843.441.607/figure-reaction-yield-conventional-bimolecular-reaction-asafunction-andnd.webp)
![FIGURE 5 Relative yield of a dimerization reaction (k = 2) for [R]/](https://thumb-eu.123doks.com/thumbv2/123dok_br/18602834.909396/5.918.87.437.68.301/figure-relative-yield-dimerization-reaction-k-r.webp)
![FIGURE 7 Relative yields for the final product of two consecutive cata- cata-lyzed reactions as a function of Ndom with [El ]/[ l(] = 10](https://thumb-eu.123doks.com/thumbv2/123dok_br/18602834.909396/6.918.98.449.84.316/figure-relative-yields-final-product-consecutive-reactions-function.webp)