UNIVERSIDADE FEDERAL DO PARÁ INSTITUTO DE CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA AQUÁTICA E PESCA
Texto
Imagem
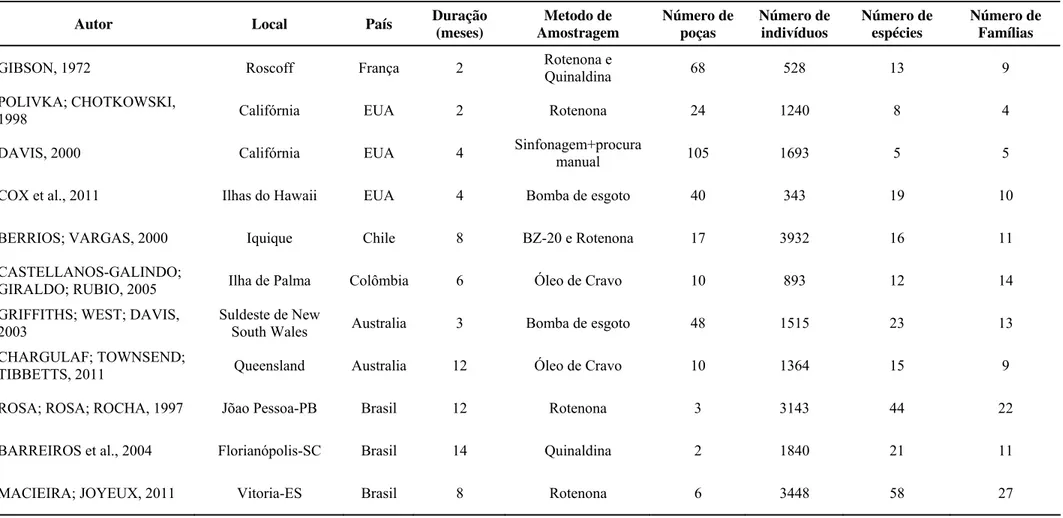






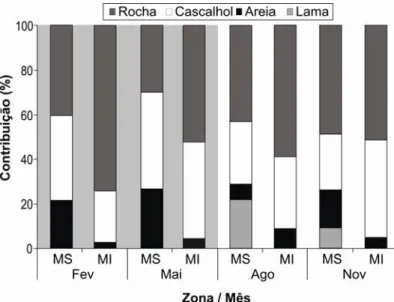


Documentos relacionados
a) Considerar apenas os cargos reportados, consoante os casos, ao regime definido pela Lei nº 2/2004, de 15 de Janeiro (republicado pela lei nº 51/2005 de 30 de Agosto e
(“Corretora”), que compreendem o balanço patrimonial em 30 de junho de 2020 e as respectivas demonstrações do resultado, do resultado abrangente, das mutações do
Petrolisthes armatus se reproduz durante o ano inteiro no estuário de Marapanim, zona costeira amazônica, sendo a presença de fêmeas ovígeras mais intensa em agosto
Detalhes mais próximos da teleoconcha, evidenciando na segunda volta costelas axiais fortes e suas micro linhas
Fonte: Trabalho de campo (2013, 2014, 2015).. O pescador que menos conhece o ambiente da pesca procura se apropriar dos pontos a partir da observação do outro. Quando verifica
Densidade e biomassa (média ± ep) de Nematopalaemon schmitti, Farfantepenaeus subtilis, Xiphopenaeus kroyeri e Acetes marinus nos três setores do Estuário
Exemplares de Hypancistrus zebra foram capturados na região do rio Xingu, na área de distribuição da espécie, entre a localidade de Gorgulho da Rita e a vila
Para a o período I as coletas totalizaram cerca de dezenove pontos nos ambientes (Lago, Parana e Ressaca) e habitats (macrófitas aquáticas, galhada, margens e praias) dentro da
