UNIVERSIDADE DO CENTRO DE
ALGARVE INVESTIGACIÓNS
FACULDADE DE MARIÑAS (CIMA)
CIÊNCIAS DO MAR Consellería de Pesca,
E DO AMBIENTE Marisqueo e Acuicultura
XUNTA DE GALICIA
“CRECIMIENTO Y REPRODUCCIÓN DE LA OSTRA
RIZADA, Crassostrea gigas (Thunberg, 1793), CULTIVADA
EN INTERMAREAL Y EN BATEA EN GALICIA (NW
ESPAÑA)”
(Dissertação para a obtenção do grau de mestre em Aquaculture
and Fisheries)
ARACELI ESCUDEIRO ROSSIGNOLI
FARO
UNIVERSIDADE DO CENTRO DE
ALGARVE INVESTIGACIÓNS
FACULDADE DE MARIÑAS (CIMA)
CIÊNCIAS DO MAR Consellería de Pesca,
E DO AMBIENTE Marisqueo e Acuicultura
XUNTA DE GALICIA
“CRECIMIENTO Y REPRODUCCIÓN DE LA OSTRA
RIZADA, Crassostrea gigas (Thunberg, 1793), CULTIVADA
EN INTERMAREAL Y EN BATEA EN GALICIA (NW
ESPAÑA)”
(Dissertação para a obtenção do grau de mestre em Aquaculture and
Fisheries)
ARACELI ESCUDEIRO ROSSIGNOLI
FARO
2006
Os investigadores Drs, Dna Maria Teresa Vieira Coelho Dinis da
Universidade do Algarve (Faro, Portugal) e D. Alejandro Guerra
Diaz, do Centro de Investigacións Mariñas (Pontevedra, España).
CERTIFICAN
Que o traballo realizado por ARACELI ESCUDEIRO
ROSSIGNOLI, no Centro de Investigacións Mariñas (CIMA) da
Xunta de Galicia, titulado "Crecimiento y reproducción de la ostra
rizada, Crassostrea gigas (Thunberg, 1793), cultivada en
intermareal y en batea en Galicia (NW España)", foi realizado baixo
a súa tutoría e cumpre as condicións para ser presentado como
Dissertaçao para el Master International in Aquaculture and
Fisheries.
Para que así conste firman o presente Vilanova de Arousa, a
27 de xaneiro de 2006.
y Universidade do Algarve (Faro, Portugal).
ORIENTADORES: Dr. Alejandro Guerra Díaz y Dra. Maria Teresa Vieira Coelho
Dinis.
FECHA: 27 de enero de 2006.
TÍTULO DA DISSERTAÇÃO: “CRECIMIENTO Y REPRODUCCIÓN DE LA
OSTRA RIZADA, Crassostrea gigas (Thunberg, 1793), CULTIVADA EN INTERMAREAL Y EN BATEA EN GALICIA (NW ESPAÑA)”.
_______________________________________________
JÚRI:
Presidente: Doutor Karim Erzini, Professor Associado da Faculdade de Ciências do
Mar e do Ambiente da Universidade do Algarve.
Vogais: Doutor Alejandro Guerra Díaz, Director do Centro de Investigações
Marinhas, Direcção Peral de Inovação e Desenvolvimento Pesqueiro – Espanha;
Doutora Maria Teresa Vieira Coelho Dinis, Professora Catedrática da Faculdade de Ciencias do Mar e do Ambiente da Universidade do Algarve; Doutora Ana Manuel Pereira Almeida Costa, Profesora Auxiliar da Universidade de Évora.
AGRADECIMIENTOS
Al Dr. Alejandro Guerra Díaz, por la dirección de este proyecto de master.
A la Dra. Maria Teresa Dinis, codirectora de este proyecto, por sus útiles
consejos.
A David Iglesias, sin el cual este trabajo no hubiera sido posible.
A los ostricultores Anselmo Bugallo, César García y Jacinto Piñeiro, por su
inestimable cooperación en las labores de cultivo en batea.
A las asistencias técnicas de la cofradía de Vilaxoán, por la colaboración
en las labores de cultivo en intermareal.
A Mª Isabel Meléndez y Elena Penas, por la realización del procesamiento
de las ostras.
A Pilar Fraguela, por la aportación de numerosa bibliografía y
documentación relevante.
ÍNDICE
I. INTRODUCCIÓN GENERAL...1
I.1. Clasificación taxonómica de Crassostrea gigas...2
I.2. Descripción de Crassostrea gigas...3
I.3. Sexualidad y ciclo reproductivo de C.gigas...9
I.4. Ecología, Hábitat y Distribución...11
I.5. Antecedentes y situación actual del cultivo...14
I.6 Técnicas de cultivo...18
I.6.1 Producción de semilla...18
I.6.2 Preengorde y engorde...20
I.7.Contexto económico.
.....25
II. JUSTIFICACIÓN Y OBJETIVOS...29
III. MATERIAL Y MÉTODOS ...31
III.1. Origen de las ostras y localización de los puntos de estudio...32
III.2. Métodos y condiciones de cultivo. Plan de trabajo...34
III.3. Evaluación de la condición gonadal...34
III.4. Variables biométricas e índices de condición...35
III.5. Parámetros ambientales...37
III.6. Reclutamiento natural...37
IV. RESULTADOS...39
IV.1. Evaluación de la condición gonadal...40
IV.1.1. Sexualidad y relación entre sexos (sex ratio)...40
IV.1.2. Ciclo reproductivo...43
IV.2. Variables biométricas e índices de condición...51
IV.3. Parámetros ambientales...55
IV.4. Reclutamiento natural...
.....56
V. DISCUSIÓN...57
VI. CONCLUSIONES...68
VII. REFERENCIAS BILIOGRÁFICAS...70
VIII. ANEXOS...81
I.-INTRODUCCIÓN GENERAL
I.1. Clasificación taxonómica
Los moluscos constituyen el mayor filo de invertebrados después de los artrópodos y en él se han descrito más de 50000 especies actuales. Los animales que integran este filo son algunos de los invertebrados más llamativos y mejor conocidos ya que entre ellos destacan especies de almejas, ostras, calamares, pulpos y caracoles (Ruppert & Barnes, 1995).
La “Ostra Rizada o del Pacífico” en la cual se centra el presente estudio, pertenece, a este filo y según Hayward, et al. (1998), se clasifica taxonómicamente de la siguiente manera :
Phylum: Mollusca, (Cuvier, 1798)
Clase: Pelecypoda o Bivalvia, (Linnaeus, 1758) Subclase: Pteriomorphia, (Beurlen, 1944). Orden: Pterioida, (Newell, 1965)
Familia:Ostreidae, (Rafinesqui, 1815) Género: Crassostrea, (Sacco, 1897)
Especie: Crassostrea gigas: (Thunberg, 1793)
I.2. Descripción de Crassostrea gigas
Presenta un tamaño que puede alcanzar los 300 milímetros de longitud (incluso en casos especiales hasta los 400 mm) (Hayward, et al. 1998). Las formas de la concha son muy variadas y dependen del tipo de sustrato al que se fije.
Figura 1: Crassostrea gigas (Thunberg, 1793). Fuente: Fundación Alfonso Martín Escudero (2002)
Generalmente, la concha se caracteriza por ser más larga que ancha y presentar una ornamentación característica formada por costillas radiales gruesas que se distribuyen de manera concéntrica por las valvas acentuando el aspecto rugoso de la ostra. Las valvas son ambas convexas y bastante similares entre si aunque la inferior o izquierda es ligeramente más convexa que la superior o derecha (Figura 1). Presenta un
umbo prominente y frecuentemente enrollado en la valva izquierda, siendo esta una
característica distintiva de la especie. Su coloración externa es blanca con vetas marrones, a veces con irisaciones gris verdosas y destellos azules (Matus, 2004). La
charnela, por donde se articulan las valvas, está ondulada en el adulto sin llegar a
formar los dientes característicos (Ruiz, 1992).
Presenta un solo músculo abductor (monomiaria) (Figura 2) formado por, fibras lisas o tónicas (opaco), responsables de las contracciones lentas que mantienen las
valvas cerradas durante largos períodos de tiempo, y fibras estriadas o fásicas (semitranslúcidas), encargadas del rápido cierre de las valvas que se producen cuando el animal es molestado por cualquier motivo. La impresión que el músculo provoca, se localiza cerca del margen posterior de la valva y más cercana al margen ventral que a la charnela; tiene aspecto reniforme y aparece con una coloración púrpura intenso característica (Figura 1).
El manto, formado por dos lóbulos denominados "lóbulos del manto o paleales", es una fina lámina de tejido secretor situado bajo las valvas que se fija a la región anterodorsal del cuerpo desde donde se continúa hacia la boca y palpos labiales envolviendo completamente todo el cuerpo del animal y formando lo que se denomina “capuchón cefálico” (Grassé, et al. 1976) (Figura 2). La cavidad del manto o cavidad
paleal es una cámara interna que se forma como consecuencia de la compresión dorso –
ventral de los lóbulos del manto y cuya misión es, de alguna manera, proteger los órganos de alimentación y reproducción. También a dicha cavidad van a desembocar los conductos excretores, genitales y de la masa visceral, acentuando aún más su importancia.
La ostra del Pacífico también se caracteriza por presentar una cámara o pasaje
promial, que no es más que un espacio que se forma sobre la región pericárdica, entre
el músculo aductor y la charnela, y que establece la separación entre la masa visceral y el lóbulo derecho del manto. La presencia de esta cámara ha sido observada por diversos autores a lo largo del tiempo (Nelson, 1938; Thomson, 1954; Walne, 1980) y constituye según Galtsoff (1964) un carácter taxonómico primordial a la hora de diferenciar entre los dos géneros más importantes de la familia Ostreidae; el género
Ostrea, sin pasaje promial y el género Crassostrea, con cámara promial. Son muchas
las funciones que se le han atribuido a esta cámara, entre ellas, destaca el que sea un área de descarga adicional para el agua filtrada por las branquias (Galtsoff, 1964; Quayle & Newkirk, 1989; Mitchell, et al. 2000) ayudando a la expulsión de pseudoheces. Es precisamente esta capacidad de producir pseudoheces la que permite a las especies del género Crassostrea crecer y sobrevivir en aguas que presentan alta turbidez de una manera mucho mas exitosa con respecto a otras especies.
Los palpos labiales son 4 láminas (2 pares) tisulares aplanadas que se encuentran situadas en la parte anterior del cuerpo a ambos lados de la boca y fusionados por su parte proximal al manto y a la masa visceral (Figura 2).
Figura 2: Órganos internos de Crassostrea gigas. Fuente: Mitchell et al. (2000).
A,Mu-Músculo abductor; F-Fusión de los lóbulos del manto con las branquias; G-Branquias; H-corazón; Hi-Ligamento; L.M.-Manto izquierdo;L.P-Palpos labiales; M-Boca; P-Cámara promial; R-Recto; R.M- Manto derecho; S-Concha; T-Tentáculos.
Las branquias son de tipo lamelibranquias (Figura 2). Están formada por dos pares de lamelas (4 filamentos branquiales), cada uno de los cuáles forma una hemibranquia que se pliega en forma de U, y que a su vez se unen mediante puentes interlamelares formando una estructura, que en corte transversal, tiene aspecto de W. Estas tienen numerosas funciones: respiración, alimentación (1ª selección del alimento) y como señalaba Galstoff (1964), reproducción, ya que son los encargados de dispersar los gametos durante el desove y, en las especie larvíparas (Ostrea edulis), proporcionan el medio idóneo para la fertilización y posterior incubación de los ovocitos.
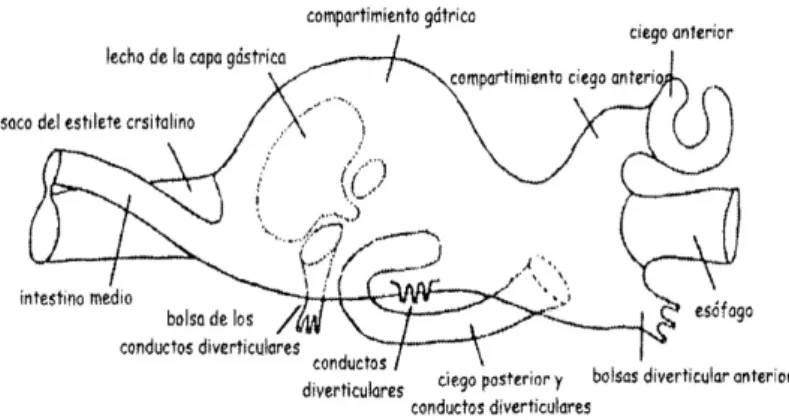
El tubo digestivo (Figura 3) está formado por un esófago corto y ciliado, un amplio estómago con numerosos surcos, divertículos y crestas , un intestino con surcos y un ano, situado sobre el flanco del músculo abductor. Rodeando al estómago se encuentra la glándula digestiva o hepatopáncreas que es donde se asimilan y digieren intracelularmente los alimentos (Galstoff, 1964). En el interior de uno de los surcos intestinales aparece una varilla compacta semitransparente formada por una matriz proteica y denominada estilo cristalino que se mueve libremente en el interior de un saco ciliado, el saco del estilo. Se establece un flujo bidireccional continuo de entrada de material asimilable y salida de desechos (Galtsoff ,1964; Walne, 1980).
Las especies del género Crassostrea, al igual que el resto de los bivalvos característicos del intermareal, solo se alimentan durante la pleamar y en consecuencia todos sus procesos digestivos están obligados también a seguir el ritmo de las mareas.
Crassostrea gigas como filtrador que es, ingiere, básicamente, fitoplancton
microscópico, detritos y materia particulada que encuentra en el medio marino.
Figura 3 : Esquema de las estructuras del aparato digestivo de C.gigas. Fuente: Evseev, et al. (1996).
El sistema circulatorio está tabicado, es abierto y está formado por un corazón de color marfil o blanco – amarillento, encerrado en una cavidad pericárdica con un ventrículo que se pliega rodeando al intestino a nivel del recto y dos aurículas laterales marrón claro (Figura 2).
El aparato excretor está formado por dos nefridios con una estructura en forma de U, situados entre la cavidad pericárdica y las branquias (Grassé, et al. 1976).
El sistema nervioso de las ostras presenta simetría bilateral y está formado por dos pares de ganglios: cerebral y visceral, y dos pares de largos cordones nerviosos que atraviesan todo el cuerpo (Ruppert & Barnes, 1995) Los únicos órganos sensoriales importantes que presenta Crassostrea son unos tentáculos paleales, localizados en el borde del manto que presentan células táctiles y quimiorreceptoras.
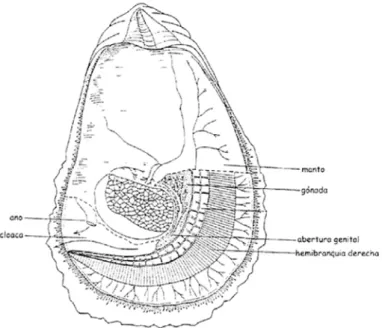
El sistema reproductor presenta una gónada situada entre la epidermis y los divertículos digestivos que, antes de que comience el desarrollo de los productos sexuales (en reposo), consiste en una serie de túmulos entremezclado con el tejido
conectivo (Figura 4). La pared externa de cada túbulo, es decir, la mitad más próxima a la superficie del cuerpo, está revestida de epitelio ciliado, mientras que la pared opuesta, más cercana a los divertículos, lo está por una capa doble o triple de ovogonias y espermatogonias distinguibes en esa fase indiferenciada. (Cole, 1942; Román, 1992).
Figura 4 : Anatomía interna de un ejemplar maduro de Crassostrea Virginica. Fuente: Galtsoff (1964)
Cuando la gónada está madura constituye un órgano de gran tamaño y aparece en forma de racimos de túmulos (folículos) que emergen a lo largo de la superficie dorsal del cuerpo para formar una estructura continua que acompaña a la masa visceral y se extiende centralmente hasta el ápice del proceso pilórico (Galtsoff, 1964). En ostras sexualmente maduras la superficie que cubre la gónada llega a ser tan fina que se observa una maraña de finos canales genitales. Los canales genitales van aumentando su diámetro gradualmente hasta que convergen en un amplio gonoducto a través del cual se descargan las células sexuales.
I.3. Sexualidad y ciclo reproductivo de C. gigas
En 1943, Coe estableció una clasificación de los distintos tipos de reproducción sexual que pueden presentarse en los bivalvos; y en ella incluye a Crassotrea gigas como un claro ejemplo de hermafroditismo asincrónico alternante. Analizando cada uno de estos términos por separado nos encontramos con que; es hermafrodita porque produce ambos tipos de gametos masculinos y femeninos a lo largo de su ciclo reproductivo; es asincrónica porque los gametos se desarrollan en una misma gónada pero en momentos diferentes a lo largo de la etapa reproductiva y es alternante porque los numerosos cambios de sexo que tienen lugar a lo largo de toda la vida del animal se desarrollan al azar de tal forma que es muy difícil saber que sexo tendrá el individuo en ciclos consecutivos. Además, a esta denominación de hermafroditismo asincrónico alternante también se le puede añadir otro término y es el de protándrico (Román, 1992; Galtsoff; 1964; Coe, 1943) ya que el primer desarrollo sexual en individuos jóvenes es siempre como macho.
El ciclo reproductivo es definido por Seed (1976) como el ciclo completo de sucesos que ocurren a partir de la activación de la gónada desde el principio hasta el fin de la gametogénesis, y que finalizan con el desove. Puede ser dividido en un período reproductivo , que comienza con la iniciación de la gametogénesis y culmina con la emisión de los gametos, y un período vegetativo o de descanso, durante el cual los gametos residuales son eliminados. El período reproductivo incluye la acumulación de nutrientes para ser utilizados durante la gametogénesis, la proliferación de células goniales y su diferenciación (gametogénesis), la acumulación de gametos maduros y su liberación. (Román, 1992).
La gametogénesis de varias especies de ostreidos ha sido suficientemente descrita (Cole, 1942; Galtsoff, 1964) y sigue las pautas generales de los demás pelecípodos. Las células germinales denominadas gonocitos, dan lugar, por medio de divisiones mitóticas, a las células goniales, espermatogonias y ovogonias, que producirán , a su vez, los ovocitos y espermatocitos primarios. Estos sufren una serie de cambios nucleares característicos durante la profase de la primera división meiótica , que implican duplicación, emparejamiento, condensación y separación parcial de los cromosomas homólogos. En la espermatogénesis, los espermatocitos primarios tras sufrir la primera división meiótica dan lugar a los espermatocitos secundarios. La segunda división meiótica tiene lugar poco después y produce espermátidas haploides (4 por cada espermatocito primario) que a su vez, mediante una serie de reorganizaciones celulares, se transforman en espermatozoides maduros.
En los ovocitos primarios, cuando estos alcanza el estado diploteno, se detienen los cambios nucleares y los cromosomas, emparejados y parcialmente separados, se muestran dispersos dentro de los núcleos engrosados (vesícula germinal). El desarrollo subsiguiente continúa a través de dos fases distintas: síntesis intensa de RNA y vitelogénesis (crecimiento por acumulación de vitelo y otras sustancias nutritivas como glucógeno). Una vez que el ovocito alcanza la madurez, se separa de la pared del folículo y migra hacia el lumen donde permanece a nivel de la metafase de la primera división meiótica, hasta el desove (Eckelbarger & Davis, 1996). Cuando este tiene lugar se expulsan ovocitos primarios totalmente desarrollados, diploides, y el resto de las etapas de la meiosis se completan en el medio externo. Son, por tanto, especies ovíparas con fecundación externa.
Tras la fertilización, los individuos tienen un tamaño aproximado de 70 micras de tamaño y comienzan rápidamente el desarrollo larvario. Inicialmente, a las pocas horas, se forma una larva trocófora que nada libremente y que se alimenta, preferentemente, de los restos de vitelo. A las 48 horas, comienza la fase de larva velíger, que es planctónica y en la que se desarrollan dos valvas que encierran completamente el cuerpo y comienza ha aparecer un sistema de órganos (estómago, esófago, intestino) que favorecen la alimentación exógena. Hacia el final del estadio larvario de natación libre, con un tamaño aproximado es de 300 micras, un pie activo y una glándula funcional secretora de cemento, comienzan a desarrollarse. Una mancha ocular aparece a cada lado del cuerpo. Una vez que los individuos han alcanzado este grado de desarrollo, sufren metamorfosis y se fijan al sustrato excretando una pequeña gota de cemento en la valva inferior. En este momento, el pie ya no es necesario y su desaparición marca la transición de la vida libre al modo de vida sedentario (Galtsoff, 1964).
La puesta es dependiente de la temperatura, salinidad y disponibilidad de alimento y ocurre, generalmente en verano.
I.4. Ecología, Hábitat, y Distribución
Las especies adultas de Crassotrea gigas se caracterizan por poseer una extraordinaria capacidad para adaptarse a grandes variaciones en los parámetros ambientales del agua, especialmente, las de aquellos que afectan a su distribución como son la temperatura y la salinidad; siendo, en general, eurihalinas y estuarinas.
Han sido numerosos los estudios que han demostrado que pequeñas variaciones en la temperatura y disponibilidad de alimento, tanto estacional como anualmente, influyen
notablemente sobre el crecimiento y el esfuerzo reproductivo de las ostras (Brown & Hartwick, 1988; Hofmann, et al. 1992) .
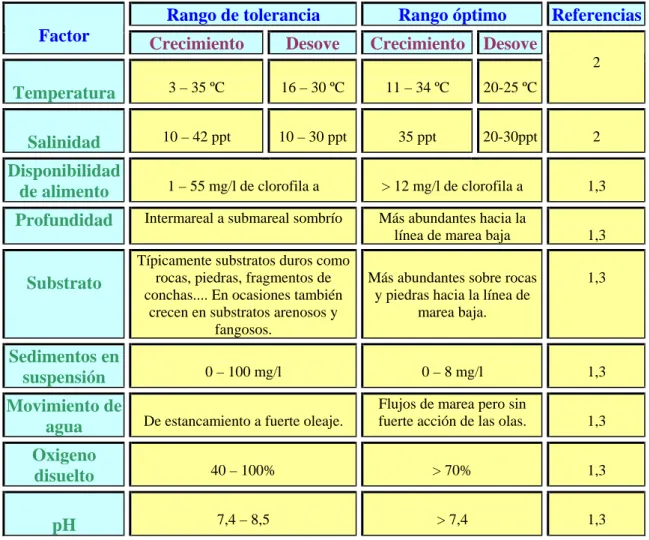
Las tolerancias a las condiciones medioambientales de un adulto de C. gigas se resumen en la Tabla 1. Es conveniente resaltar que según las diferentes regiones del mundo, las condiciones del medio necesarias para la iniciación de la gametogénesis y la puesta son diferentes; por ello en la Tabla 1 se han recogido los intervalos más amplios que abarcan, aproximadamente, la totalidad de todos los rangos de tolerancia, independientemente de la situación geográfica. La fertilización ocurre entre 15 y 30 ºC (Mitchell, 2000) , y el rango de temperaturas en el que el desarrollo larvario tiene lugar está delimitado entre 19-27 ºC con su óptimo a 21-22 ºC (Imai, 1980) . La salinidad ha sido identificado como el principal factor en el control del desarrollo larvario y aunque hay mucha variación sobre los rangos de tolerancia respecto a este factor, Mann, et al. (1991) lo establecieron entre 19 y 35 ppt. Además Héral & Deslous – Paoli (1991) establecieron que la temperatura necesaria para el desarrollo de la larva aumenta a medida que lo hace la salinidad. Las condiciones adecuadas para el desarrollo larvario deben mantenerse entre 15 y 30 días, que es el tiempo medio transcurrido desde el momento de la fertilización hasta que la larva se fija (Quayle & Newkirk, 1989; Mitchell, 2000).
Tabla 1.Rangos de tolerancia de factores medioambientales de Crassostrea gigas.
Referencias: 1. Coleman, 1996; 2. Mann, et al. (1991); 3.Mitchell, et al. (2000).
En la actualidad la especie Crassostrea gigas se encuentra distribuida por el océano Pacífico, en Japón, Korea y China, a lo largo de la costa Pacífica de Norte América desde Alaska hasta California (Shafee & Sabatie, 1986; Steele & Mulcahy, 1999), en Chile y Perú (Escapa, et al. 2004), en Costa Rica y Nueva Zelanda (Mann, et al. 1991) y en Australia (Mitchell, et al. 2000). También está presente en Brasil y Argentina (Escapa, et al. 2004); en Rusia en el mar de Okhotsk (Steele & Mulcahy, 1999); y en Europa, donde aparece en el Mar del Norte y se extiende por el
Rango de tolerancia Rango óptimo Referencias
Factor Crecimiento Desove Crecimiento Desove
Temperatura 3 – 35 ºC 16 – 30 ºC 11 – 34 ºC 20-25 ºC
2
Salinidad 10 – 42 ppt 10 – 30 ppt 35 ppt 20-30ppt 2
Disponibilidad
de alimento 1 – 55 mg/l de clorofila a > 12 mg/l de clorofila a 1,3
Profundidad Intermareal a submareal sombrío Más abundantes hacia la
línea de marea baja 1,3
Substrato
Típicamente substratos duros como rocas, piedras, fragmentos de conchas.... En ocasiones también
crecen en substratos arenosos y fangosos.
Más abundantes sobre rocas y piedras hacia la línea de
marea baja.
1,3
Sedimentos en
suspensión 0 – 100 mg/l 0 – 8 mg/l 1,3
Movimiento de
agua De estancamiento a fuerte oleaje.
Flujos de marea pero sin
fuerte acción de las olas. 1,3
Oxigeno
disuelto 40 – 100% > 70% 1,3
pH 7,4 – 8,5 > 7,4 1,3
Mediterráneo hasta Marruecos teniendo representación en Irlanda, Inglaterra, Italia, Francia, Israel, Portugal y España (Mann, et al. 1991; Ruiz, et al. 1992).
I.5. Antecedentes y situación actual del cultivo
El cultivo de moluscos bivalvos es de los más antiguos entre los animales marinos, además del más intenso y el de mayor expansión a través del mundo (Pardellas & Polanco, 1987). Dentro de las ostras, las más cultivadas para el consumo humano pertenecen a tres géneros: Crassostrea (Sacco, 1897); Saccostrea (Dollfus & Dautzenberg, 1920) y Ostrea (Linné, 1758).
La ostra plana (Ostrea edulis), que se consume desde hace siglos, siempre ha tenido una gran aceptación en antiguas culturas lo que ha contribuido a la proliferación de normas reguladoras para su extracción y cultivo (Guerra, 1995). En el principio de nuestra era ya se enviaban ostras a Roma y durante la Edad Media y el Renacimiento comienzan en Francia a explotarse los criaderos naturales. A partir del siglo XVIII, los bancos naturales de las costas Atlánticas comienzan a sobreexplotarse y aparecen las primeras disposiciones que prohíben en toda la costa francesa la recolección de ostra durante el período de reproducción. (Héral, 1991).
Un siglo después, continua aumentando el esfuerzo de pesca debido al incremento de la demanda de semilla natural, marcado por el desarrollo del comercio. Estos factores añadidos a una época de condiciones climatológicas bastante desfavorables y un aumento notable de depredadores y competidores conllevaron a diezmar casi al completo el cultivo de Ostrea edulis en Francia. Fue por esto que a partir de mediados del siglo XIX comienzan a importarse ejemplares de Crassostrea
angulata (ostra portuguesa) en la Bahía de Arcachon pero debido a un accidente de
navegación del barco “Le Morlaisien”, esta especie coloniza gran parte de la costa
sudoeste francesa compitiendo con Ostrea edulis y desplazándola en gran medida de su hábitat (Matus, 2004).
Posteriormente, en 1920, comenzaron a sucederse una serie de fenómenos de mortandad masiva debido, probablemente, a enfermedades y a regimenes de temperaturas anormales que conllevaron a una paralización casi total de la ostricultura francesa al impedirse la reproducción y reducirse al máximo la disponibilidad de semilla (Héral, 1991). Por ello a partir de ahí, el desarrollo intensivo de la ostricultura se basó prácticamente en el cultivo de C.angulata reduciéndose al mínimo la producción de O.
edulis. Así, en 1960, la producción de C.angulata en la costa Atlántica era de 85.000
toneladas frente a 28.000 toneladas de O. edulis.
Pero los fantasmas de las enfermedades volvieron a resurgir hacia 1960 cuando la ostra portuguesa comienza a padecer una serie de infecciones tales como, necrosis viral de las branquias (GNV) e infecciones hemocíticas (HIV) (Mann, et al. 1991; Marteil, 1979). Esto conlleva de nuevo, en torno a 1970, a la reducción drástica de las poblaciones naturales de C. angulata en las costas francesas.
Ante este problema, los ostricultores franceses comienzan a plantearse la introducción de un especie foránea para sustituir a la ostra portuguesa. De esta forma en 1967, aparecen las primeras importaciones en Francia de la ostra japonesa, Crassostrea
gigas, originaria del Pacífico. Esta especie demostró desde un primer momento unas
elevadísimas tasas de crecimiento, de reproducción y de resistencia a enfermedades con respecto a otras ostras. No obstante, cuando en 1977 se produjeron mortalidades limitadas en la laguna de Arcachon , se pudo demostrar la existencia en C.gigas de lesiones virales idénticas a las encontradas en C.angulata en 1970, lo que llevó a reconsiderar la resistencia de la ostra japonesa a ciertos virus (Héral, 1991). La
infección de C.gigas por el copépodo Mytilicola orientalis es un hecho reciente. Este pequeño crustáceo que vive en el tracto digestivo de la ostra fue observado en la Bahía de Arcachon por His (1977) y en la de Marennes- Olerón por Deslous – Paoli (1981); Pero parece que su presencia no es significativamente importante ya que no fue encontrada relación alguna entre infestación y mortalidad (Figueras & Villalba, 1988) y únicamente podría afectar al índice de condición de la ostra japonesa en primavera y en invierno (Héral, 1991).
La introducción de C. gigas aportó una serie de problemas biológicos, ya que con ellas se introdujeron, también, como fauna asociada otras especies japonesas entre las que se encuentran el anélido Hydroides ezoensis, el cnidario Aiptasia pulchella, el molusco Anomia chinensis o los cirrípedos Balanus amphitrite y Balanus albicostatus. (Pardellas & Polanco,1987)
En 1984, la producción de ostras francesas (100.000 toneladas) estuvo constituida por un 98% de la ostra japonesa C. gigas, cultivada sobre una superficie de 20.000 hectáreas (Héral, 1991); debido principalmente a los ataques sucesivos que entre 1974 y 1979 sufrieron los individuos de O.edulis por parte de sendos parásitos, Martelia
refringens, en 1974 y Bonamia ostreae, en 1979 y que se propagaron por casi la
totalidad de los centros de producción, comprometiendo su cultivo.
En Galicia la existencia desde antiguo de esta especie se constata por los abundantes restos de valvas encontradas en los “concheiros” de los castros celtas, también en las cartas marinas se citan numerosos fondos rocosos con el nombre “ostreira” que indicaban la presencia de bancos naturales (Guerra, 1995). Más recientemente la abundancia se refleja en las exportaciones que se efectuaron en el siglo XIX para repoblar las áreas ostrícolas de Arcachon (Francia) (Navaz, 1942).
A pesar de que la ostra plana fue una de las especies comerciales cultivadas de mayor importancia en el litoral gallego, no se libró del progresivo agotamiento que sus bancos naturales comenzaron a sufrir en Galicia en el siglo XVIII, paralelamente a sus análogos franceses; aunque no fue realmente hasta los años 50 del siglo pasado, cuando se produce prácticamente la desaparición total de estos bancos. Fue por esto que a partir de 1960 comienzan a engordarse en las costas gallegas ostras procedentes de Francia (Matus, 2004) y a ponerse en marcha diferentes métodos de obtención de semilla.
Los primeros estudios de captación natural de semilla a nivel experimental datan de los años 30 y 50. Inicialmente estas experiencias tuvieron éxito, especialmente en las Rías Altas hasta 1980 (Saavedra, 1982), pero fue a partir de este año, cuando comienza a detectarse una disminución progresiva de la captación de la semilla y unas elevadas mortandades. Dichas mortandades, coincidentes en el tiempo con las manifestadas en los parques ostrícolas franceses, son achacadas a fenómenos de infección a través de las importaciones de ejemplares desde Francia, (Montes, et al. 1990). Debido a esto la captación natural de O.edulis comienza a ser insuficiente como para mantener una producción estable y rentable, por lo que su cultivo se basa únicamente en el engorde de semilla y ejemplares adultos importados de terceros países como Francia o Italia. Por primera vez, Galicia, que tradicionalmente había sido exportadora de ostra utilizada en la repoblación de los bancos franceses, pasaba a convertirse en importador en los años 60 del siglo XX. Es en este momento cuando se empiezan a plantear alternativas parciales al cultivo de O.edulis y de esta forma se introduce la especie de ostra alóctona, originaría del Pacífico, Crassostrea gigas, conocida en Galicia como “ostra rizada”. La autorización de su cultivo en aguas gallegas, estuvo precedida de fuertes polémicas, debido sobre todo a la ausencia de conocimiento y datos concretos sobre su desarrollo
(Molares, et al. 1986), si bien en ensayos previos se había puesto de manifiesto la favorable respuesta al cultivo de esta especie (Guerra, et al. 1987).
I.6.Técnicas de cultivo
I.6.1. Producción de semilla:
La obtención de semilla para los cultivos puede hacerse de tres formas: pesca reglamentaria de juveniles en los criaderos autorizados, captación natural de juveniles producidos por los reproductores tanto de los bancos naturales como de los cultivados y compra de semilla a un criadero-semillero (Héral, 1991).
En Francia el método más utilizado es la captación natural mediante colectores (Marteil, 1979). La obtención mediante criaderos es secundaria y únicamente se recurre a ella cuando existe déficit en los colectores. Los métodos de captación de semilla de ostra prácticamente no han cambiado desde su inicio en las costas francesas en 1875, lo único que ha ido variando son los tipos de colectores y los materiales empleados para su construcción.
La especie Crassostrea gigas es poco exigente a la hora de elegir el tipo de sustrato sobre el que fijarse, el único requisito necesario es que la superficie seleccionada debe estar exenta de suciedad, algas y lodo (Goulletquer & Héral, 1997).
Los colectores mas antiguos que se conocen son los denominados de “teja árabe” hechos a base de tejas de arcilla comprimida y cocida, semicilíndricas y más o menos porosas, que se colocan perpendiculares 2 a 2 y se unen mediante un hilo de hierro que se retuerce y se engancha en los extremos (Figura 5.B). Las tejas se encalan no solo para favorecer la separación posterior de la ostra nueva del colector, sino también para proporcionar una mayor limpieza y crear un efecto más atractivo para las larvas.
Otro tipo de colectores son los fabricados a partir de fibrocemento. Estos, también están encalados y en comparación con los de teja árabe, presentan la ventaja de que el material es menos frágil que la arcilla. Las tejas de fibrocemento se instalan preferentemente sobre mesas o soportes metálicos.
A partir de 1960 comienzan a tener gran importancia los colectores de plástico, por ser un material resistente, no susceptible a la putrefacción y de fácil manejo. Las formas de este tipo de colectores son muy variadas; redes de malla, tubos (Figura 5.C), placas, barras o discos. Estos últimos constituyen los denominados “ sombreritos chinos” (Figura 5.A); Son unos conos encalados , de 30 centímetros de diámetro y 10 centímetros de altura que se colocan en batería superpuestos unos encima de otros.
También se usan como colectores, conchas de ciertos moluscos como las propias ostras, vieiras o mejillones, especialmente en parques de aguas profundas. Una vez limpias las conchas se utilizan para preparar unos sacos de plástico enrejados, semirrígidos y de malla amplia que se colocan sobre unas mesas o soportes. Por último, también se pueden usar como colectores, estructuras de cartón a los que se somete a una especie de encalado, mezclando cal con cemento y arena. De esta forma se vuelven impermeables y más resistentes.
La monitorización y el control de la abundancia larvaria, así como de las condiciones ambientales, son factores clave en el despliegue de los colectores para maximizar el reclutamiento. El éxito de la captación en medio natural radica, entre otros factores, en la abundancia de progenitores, de ahí que sea muy común encontrarse en el caso de la ostra, con problemas serios a la hora de plantearse un proyecto serio y rentable de obtención de semilla por este sistema.
A B
C
Figura 5: Tipos de colectores más empleados en Francia para la captación de semilla de ostra rizada. Fuente: CIMA. A. Sombreritos chinos. B. Tejas árabes. C. Tubos de PVC.
I.6.2. Preengorde y Engorde:
En las etapas de preengorde y engorde, se utiliza diferentes técnicas dependiendo de las características del área de cultivo. La duración de cada etapa se encuentra sujeta a la densidad del cultivo y a la capacidad de carga del ecosistema.
Transcurrido de 6 a 18 meses tras la fijación , se procede al despegue de la ostra joven de los colectores (Marteil, 1979). Este puede hacerse o bien a mano o bien de forma automatizada; esta última tiene la ventaja de reducir la mano de obra pero es de la forma en la que se estropean más ejemplares, llegando en ocasiones a causar un 25% de mortalidades.
En el caso de los colectores encalados, la ostra se despega de la teja o similar, se lava y se clasifica por tamaños, quedando así lista para ser sometida al proceso de engorde.
Las ostras de los colectores hechos a base de conchas de otros moluscos, no necesitan ser despegadas. Simplemente cuando han transcurrido de 6 a 12 meses tras la fijación de la larva, se esparce el contenido de los sacos o barras directamente por el parque de cultivo (Grizel, et al. 1979). La semilla fijada seguirá creciendo a medida que la concha sobre la que se asienta se vaya rompiendo, sin perturbar el desarrollo normal de la ostra.
Cuando se utilizan técnicas de captación que exigen el despegue de la ostra joven de los colectores,la fase de engorde, que comienza en el segundo año de cultivo, puede realizarse sobre el fondo, en cultivos sobreelevados o en suspensión. Generalmente se cultivan hasta los 3 años de edad.
El cultivo en parque se desarrolla en la zona intermareal y consta de una serie de parcelas que en el caso de la ostricultura se suelen delimitar mediante verjas de plástico. Dichas parcelas se preparan mediante, limpiezas de fondo, eliminación de depredadores, y en casos necesarios aportes de sedimentos apropiados. Las ostras se disponen sobre los fondos así adaptados, metidas en unas bolsas de malla de material plástico, que se colocan directamente sobre el sustrato. La instalación de la semilla ha de hacerse de forma que quede bien repartida, procurando que la densidad de siembra sea acorde con la capacidad alimentaria del parque en cuestión. En el caso de C.gigas, la densidad media de su cultivo durante las etapas de preengorde y engorde es de 5 y 7 Kg/m2(Goulletquer & Héral, 1997).
En el período de tiempo que transcurre desde la época de siembra hasta la salida de la ostra para su comercialización, es fundamental evitar la proliferación de los depredadores y remover regularmente a los individuos con rastrillos u horcas para evitar que queden cubiertas por lodo.
El cultivo sobreelevado se lleva a cabo en las zonas de parques que quedan en
seco durante las etapas de mareas (Figura 6.A, 6.B). Para ello se pueden emplear cajas o casilleros ostrícolas, aunque los más utilizado son unos sacos de redes denominados “pochones”, cuya abertura se va aumentando a medida que avanza el cultivo, y que se sostienen por unas mesas o parrillas situadas sobre el sustrato a algunos centímetros de altura. Los sacos, de un tamaño estándar de 1 metro por 0,5 metros, se sitúan yuxtapuestos en filas sobre las mesas de 50 centímetros de altura y 4 metros de longitud (Héral, 1991; Marteil, 1979). Periódicamente se les da la vuelta a los sacos para impedir que proliferen las algas y se desdoblan cuando la carga se hace demasiado grande. Este tipo de cultivo sobreelevado en “pochones” presenta una serie de ventajas con respecto a otros; ya que con él se logran mayores tasas de crecimiento, mejor calidad, más facilidad de explotación y menores mortandades durante las épocas de temporales.
Aunque es muy importante mantener una densidad óptima de ejemplares y una correcta limpieza de las instalaciones ya que sino esto puede acarrear grandes inconvenientes como son la disminución de las tasas de crecimiento, aumento de las deposiciones de lodo o imposibilidad de utilización de los cultivos sobreelevados durante ciertos períodos del año.
Finalmente el cultivo en suspensión es un tipo de cultivo tridimensional, en el que se utiliza un volumen y no una superficie, con lo que se ve multiplicada su rentabilidad (Figura 7.A, 7.B). Esta modalidad permite un crecimiento que, en
ocasiones, es de 2 a 3 veces superior al obtenido mediante otros sistemas (Pardellas & Polanco, 1987) y posiblemente esto se deba a que, por este sistema, las ostras no solo están en continúa inmersión con lo que se alimentan constantemente sino que también se evitan las perturbaciones climáticas, que en otros cultivos se generan como consecuencia de estar sometidas a períodos de sequía. La suspensión puede hacerse sobre estructuras pesadas (fijas o flotantes) o sobre estructuras ligeras (barrotes flotantes o boyas). En Galicia lo más utilizado son unos artefactos flotantes denominados “bateas”. Estas están formadas fundamentalmente por una serie de flotadores de madera, hierro o poliuretano que soportan un emparrillado (de madera o poliuretano) de forma más o menos rectangular, del cuál cuelgan unas cuerdas sobre las que se sitúan las ostras directamente o los sistemas, cestas o sacos, que las contengan. Las bateas se fondean con una o dos cadenas que actúan de ancla, evitando que la estructura se desplace con las mareas o con los temporales. Cuando las ostras se colocan directamente sobre las cuerdas de la batea, se fijan con una mezcla de cemento en grupos de tres, de tal forma que la valva inferior o izquierda es la que queda adherida a la cuerda. En otras ocasiones, las ostras son introducidas en cestas planas cilíndricas de material plástico o en sacos de red, con un canal en el medio por el que se introduce la cuerda; Así se van formando pilares y se cuelgan sobre el emparrillado de la batea.
La ventaja de este sistema de cultivo es que se pone a disposición de la ostra toda la capa de agua y el inconveniente es que al estar en constante inmersión se favorece el incremento de competidores tróficos (ascidias) y algas (sargazos) sobre las estructuras de cultivo.
Figura 6 : Cultivo de ostra rizada en intermareal. Fuente: CIMA.
A. Detalle de las ostras dispuestas en el interior de los sacos ostrícolas (pochones). B. Mesas de cultivo en sobreelevado ubicadas en la playa de Corón (Vilaxoán).
A B
A B
Figura 7 : Cultivo de ostra suspendido en batea. Fuente: CIMA.
A. Cestos ostrícolas dispuestos en cuerdas. B. Ubicación de los cestos suspendidos en batea.
Una de las principales consecuencias del cultivo en batea es que la ostra al permanecer tanto tiempo en inmersión, ve disminuida su capacidad para cerrar efectivamente las valvas y presenta por ello una importante pérdida de agua en el proceso de transporte hacia el lugar de comercialización. Incluso en ocasiones, el productor que las compra se ve obligado a volver a sumergir las ostras de manera previa a su comercialización con los gastos asociados que esto conlleva. Por este motivo, se comenzaron a realizar en Galicia experiencias de afinado o de resistencia a la desecación consistentes en llevara cabo una serie de protocolos de sometimiento de las
ostras a períodos de emersión en la propia batea. Estas experiencias, reciben su nombre en referencia al proceso de “affinage” que practican los ostricultores franceses en estanques (Figura. 8) con el fin de mejorar las propiedades organolépticas de sus ostras incrementando así, tanto su calidad final como su valor en el mercado (Marteil, 1979)
Figura 8: Estanque de afinado en la Île d`Oleron (Francia). Fuente: CIMA.
I.7. Contexto económico
Actualmente Crassostrea gigas es la especie de ostra más cultivada en el mundo. Según datos de la FAO, la producción mundial de ostras en el 2003 fue de 4.496.659 toneladas, lo que equivale a unos 3.794.374.000 $US en divisas. De esta producción, 4.376.802 toneladas (3.687.951.000 $US) fueron de C. gigas. Por continentes, Asia es con diferencia el mayor productor de esta especie con 4.191.925 toneladas en 2003 (3.299.645.000 $US en divisas), seguido por Europa a gran distancia con 122.540 toneladas (304.549.000 $US). Desglosando los datos por países (Tabla 2), la China es el principal productor mundial de Crassostrea gigas, seguido por Japón, República de Corea, Francia, que es el principal productor de la Ostra del Pacífico en Europa, y Estados Unidos.
Tabla 2. Principales países productores de Crassostrea gigas en el año 2003. (Fuente FAO, 2003)
País Producción (ton) Divisas ($US)
China 3.668.237 2.751.178.000
Japón 260.644 390.966.000
República de Corea 238.326 82.480.000
Francia 115.000 284.893.000
E.E.U.U. 51.012 43.675.000
La producción total de moluscos en Francia ascendió en 2002 a 187.599 toneladas, facturando en divisas unos 30.053.219 €. De esta cifra, la mayor parte correspondía a Crassostrea gigas (Tabla 3).
La producción de C.angulata ha experimentado una cierta mejoría en los últimos años, aumentando de 4.166 toneladas en 1984 a 14.000 toneladas en 1995, año en el que constituyo el 6,51% de la producción total de moluscos en Francia (FAO, 1997). Esta industria tiene lugar principalmente en la costa atlántica francesa, en la zona litoral intermareal que va desde Arcachon a Normandía, en cultivo sobreelevado en sacos ostrícolas (pochones). La producción se orienta básicamente al mercado y consumo nacional y genera un gran número de puestos de trabajo debido a su carácter artesanal. Ocupa 20.000 hectáreas enconcesión por parte del Estado, de las que 14.000 se sitúan en la zona intermareal y 6.000 en mar abierto (Grizel, 1996; Héral, 1991). Por otro lado, la producción de ostras triploides en Francia es una actividad a tener en cuenta ya que está sufriendo una gran expansión. Esto, en gran parte se debe a que tiene una calidad constante de carne, una mayor tasa de crecimiento, alcanza la talla comercial unos meses antes que los ejemplares diploides y , en general, el manejo del cultivo es menos laborioso, con lo que se aseguran la disponibilidad de un producto de calidad todo el año. El problema está en que las tasas de mortalidad parecen ser superiores, especialmente en los períodos de escasez de fitoplancton. Son individuos
estériles ya que no completan la gametogénesis por lo que presentan índices de condición muy elevados durante todo el año y, como no tienen gónadas lechosas en los meses estivales, se hacen más apetecibles para el consumidor en esta época, momento en el que la ostra diploide lo es menos ya que se encuentra en período de desove. Su desarrollo comenzó en Estados Unidos y llegó a Francia hacia 1980 lo que supuso una gran revolución para el sector ostrícola. Actualmente existen algunos países donde hasta más del 50% de la producción de semillas en criadero está constituida por triploides (Guo, et al. 1996) .
En España, actualmente se cultivan 3 especies de ostra: O.edulis; C.gigas y
C.angulata . Las producciones totales de las 2 primeras para el año 2002 se reflejan en
la tabla 3.
Tabla 3. Producción y divisas de Ostrea edulis vs Crassostrea gigas en España y Francia en el año 2002. (Fuente FAO, 2002).
España Francia
Ostras
Producción (Ton) Divisas (€) Producción (Ton) Divisas (€)
Ostrea edulis 4.565 10.971.674 6583 17.866.953
Crassostrea gigas 591 1.420.600 113.877 20.287.897
Crassostrea gigas en España, se cultiva en la costa cantábrica, Delta del Ebro,
región suratlántica y Galicia. Esta última es la principal comunidad española en producción ostrícola con un 85 – 90% del total y abastece totalmente el mercado nacional, repartiéndose las producciones entre las grandes superficies, los diferentes mercados y la restauración (Ministerio de Agricultura, Pesca y Alimentación, 1999). En 2002 las tasas de producción gallegas fueron de 369 y 3.783 toneladas para C.gigas y
O.edulis respectivamente (Fuente, Voz de Galicia 23-05-2004)
Actualmente existen en Galicia alrededor de 150 bateas dedicadas al cultivo experimental de la ostra, de las cuales salen al mercado cerca de 4.500 toneladas anuales, con una facturación aproximada de 11.000.000 €.
.
II. JUSTIFICACIÓN Y OBJETIVOS
En respuesta a las demandas del sector ostrícola, la Dirección Xeral de Recursos Mariños inicia en 2004 un proyecto coordinado destinado a valorar desde un punto de vista integral –técnico, biológico, ecológico y económico- la viabilidad del cultivo de la ostra rizada (Crassostrea gigas) en Galicia como posible complemento al cultivo de la ostra plana.
El presente trabajo que se integra dentro del citado proyecto, tiene como finalidad realizar una valoración del crecimiento y potencial reproductor de Crassostrea
gigas en las Rías Bajas gallegas. Para ello se establecieron los siguientes objetivos:
1. Definir el ciclo de desarrollo gonadal propio de ostras adultas cultivadas en aguas gallegas mediante dos sistemas de cultivo diferentes: suspendido en batea y sobreelevado en intermareal.
2. Determinar el patrón de crecimiento y el rendimiento en carne de las ostras mediante el registro de una serie de variables biométricas y el cálculo de índices de condición fisiológicos (I.C.)
3. Valorar si se produce reclutamiento de semilla de C. gigas a lo largo del período de estudio.
III. MATERIAL Y MÉTODOS
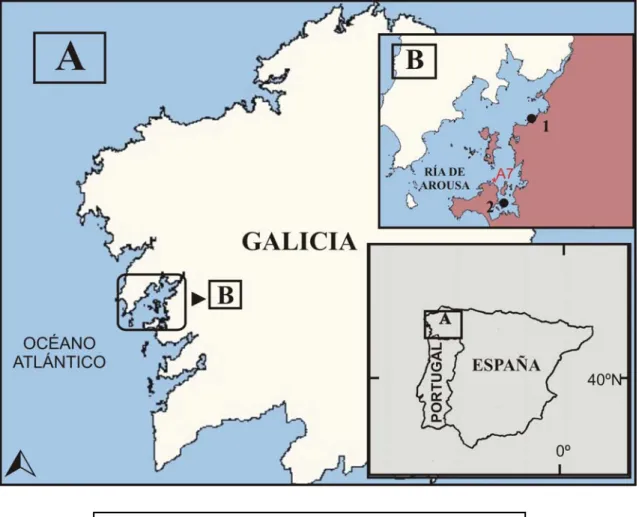
III.1. Origen de las ostras y localización de los puntos de cultivo
En febrero de 2005 se recibe en el Centro de Investigaciones Marinas (CIMA) de Corón - Vilanova de Arousa, un lote de 4000 unidades de ostra de 18 meses de edad con una talla media (± EE) de 46,8 (± 1,5) mm y un peso total medio (± EE) de 8,95 (± 0,63) g, procedentes de captación natural de la Bahía de Arcachon (Francia). Este lote se somete a un control inicial nada más llegar, basado en análisis biométrico (talla y biomasa) e histopatológico (para detectar si existía alguna alteración patológica de importancia) de una muestra de 30 individuos. Una vez tramitados los correspondientes permisos de inmersión se introducen los lotes en los dos puntos geográficos elegidos para la realización del estudio; Polígono de Grove F y Polígono de Vilaxoán. Ambos polígonos están localizados en la Ría de Arousa. Dicha ría se sitúa en la costa nor-occidental de la Península Ibérica y pertenece junto con las Rías de Vigo, Pontevedra, Arosa y Muros-Noia a las denominadas Rías Bajas gallegas (Figura 9) (Guerra, 1995). Es un estuario de 25 kilómetros de longitud, con una superficie global de 230 Km2 y un volumen aproximado de 4,3 × 103 m3
(Otto, 1975). Se caracteriza por poseer importantes corrientes de marea, cuyo flujo tiende a penetrar por el lado sur. Además sus entradas están parcialmente protegidas por islas e islotes. Las profundidades que presenta, también son importantes, si bien van disminuyendo hacia el interior, apreciándose una tendencia a la sedimentación fangosa o arenosa. Otra característica interesante de la Ría de Arousa es que tiene un profundo canal central ( de mas o menos 70 metros) que facilita la circulación, mezcla y renovación de las aguas. Además no
existen alturas próximas al litoral lo que contribuye igualmente a la renovación de las aguas por el viento, favoreciendo los afloramientos. Está abierta al océano mediante un canal, entre las llamadas Isla de Sálvora y la Península del Grove (Pardellas & Polanco, 1987). Soporta una gran producción, no sólo de mejillón, sino también de almeja, berberecho, navaja y ostra, que son objeto de explotación en los numerosos parques que existen en esta ría.
Figura 9 : Ubicación de los lotes de Crassostrea gigas estudiados. Fuente : CIMA.
1. Vilaxoán (intermareal). 2. Grove F (batea). A7. Estación oceanográfica.
III.2. Métodos y condiciones de cultivo. Plan de trabajo
Entre febrero y noviembre de 2005, se recolectaron - con carácter mensual- entre 15 y 30 ejemplares en cada uno de los puntos de cultivo objeto de estudio, siendo trasladados al laboratorio en neveras isotermas. Los individuos se procesaron para los análisis correspondientes, en un período de tiempo lo más breve posible.
.
.
.
Los sistemas de cultivo empleados fueron el cultivo sobreelevado en intermareal, para el polígono de Vilaxoán y el cultivo suspendido en batea para el polígono Grove F. La distribución de los lotes se detalla en la tabla 1 de Anexos, las condiciones de cultivo y el plan de trabajo seguido en las tablas 2 y 3 de Anexos respectivamente.
III.3. Evaluación de la condición gonadal
A cada individuo muestreado se le extrajo una pieza de tejido (de aproximadamente 5 milímetros de espesor) correspondiente a una sección transversal realizada a la altura de la unión entre los palpos labiales y las branquias. Esta muestra de tejido se procesó mediante técnicas histológicas clásicas siguiendo el protocolo descrito en la tabla 4 de Anexos. Las secciones histológicas resultantes fueron examinadas al microscopio óptico con el fin de determinar la maduración gonadal.
Para cuantificar la maduración gonadal se estableció una escala propia que comprendía 6 estados; ésta se desarrolló a partir de los esquemas propuestos por Mann (1979) y por Steele & Mulcahy (1999), y se expone con detalle en el apartado IV.1.2.
En base a dicha escala se calculó un índice de maduración gonadal (I.M.G.) (Seed & Brown, 1975) mensual (para el conjunto de la población y, también, para machos y hembras por separado) como una media ponderada conforme a la siguiente categorización de los estados de maduración gonadal (Shafee & Daoudi, 1991):
Estado 0 (Gónada inactiva o reposo) :0 Estado 1 (Actividad temprana) :1
Estado 2 (Gónada en desarrollo) :2
Estado 3 (Madurez) :3
Estado 4 (Desove) :2 Estado 5 (Reabsorción) :1
Para cada muestra, el número de ostras en cada estado fue multiplicado por el “peso”atribuido a cada estado. El producto de esta multiplicación fue sumado y el resultado dividido por el número total de ostras en la muestra. El desarrollo gametogénico fue indicado por un incremento en el índice mientras que una disminución del mismo señalaba que la puesta había tenido lugar (Shaffe & Daoudi, 1991).
Asi mismo, con el fin de determinar si la relación entre sexos (sex ratios) se ajustaba a una distribución 1:1, se realizaron, mediante EXCEL, tests Chi-cuadrado de bondad de ajuste sobre las muestras mensuales, y sobre el total de individuos analizados a lo largo del período estudiado. La hipótesis nula (Ho) contrastada fue que la relación entre machos y hembras se ajustaba a 1.
III.4. Variables biométricas e índices de condición
Los ejemplares recolectados (15 -30/mes) se midieron con un calibre (0,01 mm de precisión) y se pesaron con una balanza de precisión (sd=0,01g).
Inicialmente, además de la longitud, se registró el peso total de cada uno de los individuos muestreados. A continuación se abrió cada una de las ostras y se extrajo la
carne, cuyo peso húmedo fue registrado tras un breve escurrido sobre papel secante. Estos tejidos húmedos fueron sometidos a deshidratación en estufa a 60ºC durante 72ºC, tras lo cual su peso seco fue registrado. Con las valvas se procedió de la misma forma, registrando tanto su peso húmedo como su peso seco tras el período de desecación en estufa. Todos los datos fueron archivados en EXCEL y el valor promedio, la desviación típica y el error estándar calculados para cada lote y período muestreado.
Una vez obtenidas estas variables biométricas, se procedió a realizar los cálculos de los índices de condición fisiológicos (I.C.) según la siguiente expresión (Walne, 1976; Mann & Glomb, 1978):
IC = Peso seco vianda (g) x 100/ Peso seco valvas (g)
De la misma forma que con las variables biométricas, todos los índices calculados fueron registrados en hojas de cálculo de EXCEL y se obtuvieron la media aritmética, desviación típica y error estándar para cada uno de los lotes cultivados y en cada uno de los muestreos realizados. Con el fin de valorar si existían diferencias significativas en el último mes muestreado (noviembre 2005), entre ambos lotes de cultivo en lo que respecta a las diferentes variables biométricas e índices de condición, se realizaron tests no paramétricos de Mann-Whitney, puesto que alguna de las variables no se ajustaba a una distribución normal. Asimismo se calcularon coeficientes de correlación por rangos de Spearman entre los valores de índice de maduración gonadal (IMG) y los índices de condición fisiológicos obtenidos con el fin de evaluar si existía algún tipo de relación entre la evolución del ciclo gonadal y los mismos. Para ello se empleó la aplicación informática MINITAB 14.0.
III.5. Parámetros ambientales
No fue posible disponer de los registros de condiciones oceanográficas que el
INTECMAR1 realiza en las rías gallegas, porque los mismos se editan al concluir la anualidad en curso. En su defecto, y con motivo de referenciar al menos el rango de temperatura y salinidad imperante en la Ría de Arousa –en la que se ubican ambos puntos de estudio- se emplearon los registros de la estación A7 correspondientes al año 2004 (Figura 9).
1
INTECMAR: El Instituto Tecnológico para o Control do Medio Mariño de Galicia registra,
semanalmente, parámetros ambientales a lo largo de una red de estaciones oceanográficas incluidas dentro de un programa de control, promovido por la Consellería de Pesca y Asuntos marítimos, cuya finalidad es la de tener en todo momento un conocimiento completo de las características del medio.
III.6. Reclutamiento natural
Con el fin de disponer de datos reales acerca de la viabilidad de las puestas de
Crassostrea gigas en las rías gallegas, se emprendió una campaña de disposición de
colectores en los puntos geográficos donde se estaba llevando a cabo el estudio de seguimiento del ciclo gonadal (Vilaxoán y Grove F). Para ello con periodicidad mensual entre los meses de junio y septiembre –período de puesta de C. gigas en las rías gallegas (Ruiz, et al. 1992; Rodríguez & Carrasco, 1993)- se dispusieron colectores comerciales utilizados por la industria ostrícola francesa para la captación de ostra. En el caso del cultivo sobreelevado del intermareal de Vilaxoán se emplearon dos tipos diferentes de colectores: “sombreritos chinos” y torres colectoras de semitubos plásticos, ubicados sobre parrillas metálicas sobreelevadas (Figura.10). En la batea de Grove F se colgaron únicamente torres colectoras de semitubos plásticos (Figura 11). En enero de 2006 está prevista la retirada de estos colectores y su traslado al laboratorio para proceder a la cuantificación de la cantidad de semilla de ostra fijada. Previamente en noviembre de
2005 se realizó una valoración cualitativa in situ de la presencia de fijaciones en los mismos.
Figura 10: Colectores comerciales dispuestos en intermareal (Vilaxoán) en las experiencias de captación natural desarrolladas en el proyecto: sombreritos chinos y torres colectoras de semitubos plásticos encalados. Fuente: CIMA.
Figura 11: Torres colectoras de semitubos plásticos encalados situados en una batea ( polígono Grove F). Fuente: CIMA.
IV. RESULTADOS
IV.1. Evaluación de la condición gonadal
IV.1.1. Sexualidad y relación entre sexos (sex ratio):
En las Tablas 4 y 5 se muestran el número de machos, hembras e individuos indeterminados, global y en los diferentes períodos de muestreo, observados en cada una de las zonas estudiadas. En el cultivo sobreelevado (Vilaxoán), la relación entre sexos presentó diferencias significativas a nivel global (así como en los meses de mayo y junio) con respecto a la distribución 1:1 (Tabla 4), de manera que el número de hembras fue significativamente superior al de machos. Las ostras cultivadas en suspensión (Grove F) presentaron una distribución de sexos que se ajustó a una relación 1:1 a nivel global, y a lo largo de todo el período de estudio (Tabla 5). En el muestreo correspondiente al mes de septiembre en batea, por motivos técnicos no fue posible el procesamiento histológico de los ejemplares y por ello se carece de los datos de los índices de condición gonadal de dicho mes. Los individuos de sexo indeterminado fueron predominantes en el período invernal (febrero) y en los meses de otoño. Durante la primavera y el verano no se detectó ninguna ostra de sexo indeterminado en intermareal, pero sí en batea, donde únicamente en junio no se detectó ningún individuo indeterminado (Tablas 4 y 5).
Tabla 4: Distribución de las ostras Crassostrea gigas cultivadas mediante sobreelevado en intermareal (Vilaxoán) en función de su caracterización sexual.
Hembras Machos Indeterminados Test χ2 (1) Fecha n % n % n % Ratio H/M observado Valor χ2 (G.L.2) P (3) (significación estadística) 8/02/2005 0 0 0 0 30 100 - - - 9/03/2005 6 31,6 11 57,9 2 10,5 0,5 1,47 (1) (n.s.) 0,23 11/04/2005 16 57,1 12 42,9 0 0 1,3 0,57 (1) (n.s.) 0,45 5/05/2005 16 84,2 3 15,8 0 0 5,3 8.89 (1) 0,00 (**) 8/06/2005 21 77,8 6 22,2 0 0 3,5 8,33 (1) 0,00 (**) 8/07/2005 10 71,4 4 28,6 0 0 2,5 2,57 (1) (n.s.) 0,11 23/08/2005 5 33,3 2 13,3 8 53,3 2,5 1,29 (1) (n.s.) 0,26 6/09/2005 5 33,3 2 13,3 8 53,3 2,5 1,29 (1) (n.s.) 0,26 17/10/2005 2 13,3 2 13,3 11 73,3 1 0,00 (1) (n.s.) 1,00 TOTAL 81 53,3 42 27,6 29 19,1 1,9 12,37 (1) 0,00 (**)
1 Ho: machos y hembras siguen una distribución 1:1 2 G.L.:Grados de libertad del test
3 Probabilidad de aceptar la Ho. Criterio de significación estadística al 95%: n.s., diferencias no
significativas: p>0.05; *, 0.05>p>0.01; **, 0.01>p>0.00
Tabla 5: Distribución de las ostras Crassostrea gigas cultivadas mediante suspensión en batea (Grove F) en función de su caracterización sexual.
1 Ho: machos y hembras siguen una distribución 1:1 2 G.L.:Grados de libertad del test
3 Probabilidad de aceptar la Ho. Criterio de significación estadística al 95%: n.s., diferencias no
significativas: p>0.05; *, 0.05>p>0.01; **, 0.01>p>0.00 *Valores próximos a la significación estadística
Hembras Machos Indeterminados Test χ2 (1) Fecha n % n % n % Ratio H/M observado Valor χ2 (G.L.2) P (3) (significación estadística) 8/02/2005 0 0 0 0 30 100 - - - 30/03/2005 4 21,1 12 63,2 3 15,8 0,3 4,00 (1) 0,05* (n.s.) 12/04/2005 10 55,6 5 27,8 3 16,7 2,0 1,67 (1) (n.s.) 0,20 11/05/2005 9 45,0 10 50,0 1 5,0 0,9 0,05 (1) (n.s.) 0,82 9/06/2005 19 65,5 10 34,5 0 0 1,9 2,79 (1) 0,09* (n.s.) 8/07/2005 10 52,6 8 42,1 1 5,3 1,25 0,22 (1) (n.s.) 0,64 24/08/2005 3 20,0 8 53,3 4 26,7 0,4 2,27 (1) (n.s.) 0,13 26/10/2005 1 6,7 3 20,0 11 73,3 0,3 1,00 (1) (n.s.) 0,32 TOTAL 56 41,5 56 41,5 23 17,0 1 0,00 (1) (n.s.) 1 42
Después del análisis de secciones histológicas de 291 ostras, únicamente se detectaron 4 individuos (3 en batea y 1 en intermareal) que presentaban gametos masculinos y femeninos simultáneamente (Figura 12) si bien, debido a que la viabilidad de los mismos no pudo ser determinada, se clasificaron como “potencialmente hermafroditas”.
GF GM
Figura 12: Estado de Hermafrodita potencial. Foto realizada por Araceli Escudeiro.
Corte histológico de gónada de C.gigas. Se observan simultáneamente en la gónada gametos femeninos (GF) y masculinos (GM). Barra: 100 µm.
IV.1.2. Ciclo reproductivo:
Basándose en los criterios propuestos por Mann (1979) y por Steele & Mulcahy (1999) y en observaciones propias se elaboró una escala de madurez para el seguimiento del ciclo reproductivo. Dicha escala comprende 6 estados que se exponen a continuación:
Estado 0: Gónada inactiva o reposo:
Prácticamente la totalidad de la gónada estaba ocupada por tejido conjuntivo, no distinguiéndose, o muy difícilmente la existencia de folículos periféricos a la glándula digestiva. No resultaba posible diferenciar el sexo.
Estado 1: Actividad temprana.
Se comenzaban a detectar los primeros signos de gametogénesis. Las paredes de los folículos parecían más prominentes y estaban tapizadas por las células germinales (Figuras 13 y 14 ).
F
Ov.f
oG
Figura 14: Estado 1 de Hembra. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan ovogonias (oG) surgiendo de las paredes de los folículos y algún ovocito (Ov.f) en el lumen. Barra: 50 µm.
V SsG sPI
Figura 13 : Estado 1 de macho. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan folículos gonadales abundantes (F) con espermatogonias (sG) y espermatocitos (sPI) en la pared; pero no espermatozoos. El tejido de reserva es consumido y regenerado alrededor de los vasos hemolinfáticos (V). Barra: 50 µm.
Estado 2. Gónada en desarrollo.
Los folículos ocupaban una gran parte de la masa visceral mientras que el tejido conjuntivo estaba en regresión (Figuras 15 y 16).
Figura 15 : Estado 2 de Macho. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan folículos con espermatocitos (sPI) y espermátidas (sPd) como células predominantes. Comienzan a aparecer, de manera desordenada, espermatozoides en el folículo (sPz) con las colas orientada hacia el lumen del mismo . El tejido conjuntivo (Tc) está en regresión. Barra: 100 µm.
Tc
sPI
sPdsPz Ovf.
Figura 16: Estado 2 de Hembra. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan ovocitos fijados (Ov.f) a las paredes de los folículos en proceso de aumento de tamaño. Comienza a aparecer algún ovocito libre. Los núcleos se tiñen más claro que el citoplasma Barra: 100 µm
Estado 3: Madurez.
La gónada ocupaba la mayor parte del volumen de la masa visceral. Casi no se distinguía tejido conjuntivo separando los folículos que presentaban un aspecto muy distendido (Figuras 17 y 18).
Ov.l
Figura 18: Estado 3 de Hembra. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan folículos repletos de ovocitos libres (Ov. l) con forma poligonal y distinguiéndose de manera clara sus núcleos y nucleolos. Barra: 100 µm.
sPz
Figura 17: Estado 3 de Macho. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan los folículos llenos de espermatozoides (sPz) formando la disposición en remolino típica que llena completamente el folículo. Barra: 100 µm
Estado 4: Desove.
Ha tenido lugar una liberación parcial de los gametos y como consecuencia la presión en el interior de los folículos disminuyó. En el caso de las hembras se observaban espacios libres en el interior del folículo. Los machos, por su parte presentaban un desorden generalizado de las células sexuales. (Figuras 19 y 20).
Figura 19: Estado 4 de Macho. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observa como los espermatozoides (sPz) han perdido su disposición y aparecen desordenados en el interior de los folículos que empiezan a vaciarse. Barra: 100 µm.
sPz
A.
Figura 20: Estado 4 de Hembra. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan ovocitos en estadios de madurez junto con folículos que presentan huecos en el lumen debido a los ovocitos emitidos. Algún ovocito en atresia (A.). Barra: 100 µm.
Estado 5: Reabsorción.
Los folículos degeneran y comienza un proceso de citólisis de los gametos que no han sido liberados. La gónada se encontraba en recesión siendo sustituida por tejido conjuntivo(Figuras 21 y 22).
Figura 21: Estado 5 de Macho. Foto realizada por Araceli Escudeiro . Corte histológico de gónada de Crassostrea gigas. Barra: 100 µm.
Hm
Figura 22: Estado 5 de Hembra. Foto realizada por Araceli Escudeiro. Corte histológico de gónada de Crassostrea gigas. Se observan hemocitos (Hm) atacando los folículos, que comienzan a degenerar. Proliferación de tejido conjuntivo. Barra: 100 µm.
Cuando en una sección histológica de un individuo coexistían características de diferentes estadios de desarrollo gonadal se recurrió al criterio de clasificarlo como perteneciente al estadio en el que se encontraban la mayoría de los folículos gonadales. Asimismo aquellas preparaciones histológicas que estaban muy rotas y deterioradas, fueron descartadas del estudio.
Tal y como se aprecia en la Figura 23, a través de la evolución del índice de maduración gonadal, el ciclo gametogénico de machos y hembras presentó un patrón temporal similar. Por ello ambos sexos fueron considerados de manera conjunta en la