UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS AGRÁRIAS DEPARTAMENTO DE FITOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM AGRONOMIA/FITOTECNIA
LINEKER DE SOUSA LOPES
CONDICIONAMENTO FISIOLÓGICO DE SEMENTES DE MAMONA COMO MEIO DE ATENUAR OS EFEITOS DO ESTRESSE SALINO NA
GERMINAÇÃO E ESTABELECIMENTO DA PLÂNTULA
LINEKER DE SOUSA LOPES
CONDICIONAMENTO FISIOLÓGICO DE SEMENTES DE MAMONA COMO MEIO DE ATENUAR OS EFEITOS DO ESTRESSE SALINO NA
GERMINAÇÃO E ESTABELECIMENTO DA PLÂNTULA
Dissertação de mestrado apresentada ao Programa de Pós-Graduação em Agronomia/Fitotecnia, da Universidade Federal do Ceará, como requisito parcial para obtenção do Título de Mestre em Agronomia. Área de concentração: Fitotecnia.
Orientador: Prof. Dr. Enéas Gomes Filho
Dados Internacionais de Catalogação na Publicação Universidade Federal do Ceará
Biblioteca de Ciências Humanas
L854c Lopes, Lineker de Sousa.
Condicionamento fisiológico de sementes de mamona como meio de atenuar os efeitos do estresse salino na germinação e estabelecimento da plântula / Lineker de Sousa Lopes. – 2013.
94 f. : il. color., enc. ; 30 cm.
Dissertação (mestrado) – Universidade Federal do Ceará, Centro de Ciências Agrárias, Departamento de Fitotecnia, Programa de Pós-Graduação em Agronimia/Fitotecnia, Fortaleza, 2013.
Área de Concentração: Fisiologia de Sementes. Orientação: Prof. Dr. Enéas Gomes Filho.
1. Ricinus communis. 2. Salinidade. 3. Condicionamento fisiológico. 4. Estresse oxidativo. . Título.
CONDICIONAMENTO FISIOLÓGICO DE SEMENTES DE MAMONA COMO MEIO DE ATENUAR OS EFEITOS DO ESTRESSE SALINO NA GERMINAÇÃO E
ESTABELECIMENTO DA PLÂNTULA
Dissertação de mestrado apresentada ao Programa de Pós-Graduação em Agronomia/Fitotecnia, da Universidade Federal do Ceará, como requisito parcial para obtenção do Título de Mestre em Agronomia. Área de concentração: Fitotecnia.
À FUNCAP, pelo apoio financeiro com a manutenção da bolsa de mestrado. Ao INCTsal, pelo apoio financeiro.
Ao professor Enéas Gomes Filho pela fundamental orientação, amizade, confiança e incentivo à minha formação.
Aos professores participantes da Banca examinadora, Prof. Dr. Marlos Alves Bezerra e Prof. Dr. Alexandre Bosco de Oliveira, pelo tempo concedido, pelo valiosas colaborações e sugestões.
Aos professores e funcionários dos departamentos de Fitotecnia e de Bioquímica e Biologia Molecular, que direta ou indiretamente auxiliaram na minha formação científica.
Ao PET-Agronomia que em muito contribuiu para minha formação acadêmica durante a graduação.
Ao meu pai, Francisco José Lopes, pela educação, incentivo, companheirismo, sabedoria, compreensão e confiança.
À minha mãe, Maria Gêrda Araújo de Sousa Lopes, pelo esforço, educação, incentivo, companheirismo, sabedoria e compreensão.
Aos meus avôs, Adelson Araújo e Estevão Lopes (in memorian). Às minhas avós, Teresa Araújo e Aurelina Lopes.
À minha irmã, Alynne Lopes, pelo apoio e amizade.
À Camila Brasil, pelo amor, companheirismo e compreensão. À Alan Martins, pela amizade.
Aos meus primos, Ygor e Iuri Alves, pela amizade e ajuda.
À Viviane Ruppenthal, Elton Marques e Franklin Aragão, pela ajuda e amizade.
“Nenhuma grande descoberta foi feita jamais sem um palpite ousado”
atenuar os efeitos do estresse salino na germinação e estabelecimento da plântula. Orientador: Enéas Gomes Filho. Fortaleza: ufc. 90f. (dissertação)
RESUMO
A mamona (Ricinus communis) é uma cultura em ascensão que precisa de técnicas de produção adequadas à suas condições de cultivo e ao seu nicho de mercado. Essa cultura tem sido estudada com a finalidade de melhor adequá-la às condições ambientais do Norte e Nordeste brasileiros, regiões estas com problemas vigentes e potenciais de salinidade. Desse modo, a caracterização dos efeitos deletérios da salinidade em sementes e plântulas de mamona e a avaliação do uso das técnicas de condicionamento fisiológico de sementes podem convergir em uma forma de se reduzir custos com replantios, uniformizar a plantação e preparar as plântulas para melhor enfrentar o estresse salino. O objetivo geral da pesquisa foi caracterizar os efeitos deletérios da salinidade em sementes de mamona e avaliar o uso das técnicas de condicionamento fisiológico de sementes como meio de minimizá-los. A salinidade afetou a germinação das sementes de mamona cv. BRS-Energia apenas a partir do Ψs de -0,4 MPa de soluções de NaCl, porém, o crescimento das plântulas foi afetado sob Ψs de -0,16 MPa. A curva de embebição de água pelas sementes sob salinidade, mostrou-se desacelerada com o tempo de exposição à salinidade. A atividade da enzima dismutase do superóxido não foi afetada da salinidade, enquanto das enzimas catalase e peroxidase do ascorbato foram responsivas à salinidade. Não foi detectada atividade da enzima peroxidase do guaiacol nas sementes de mamona. Apesar dos osmocondicionamentos com CaCl2, NaNO2 e PEG-6000, testados sob condições salinas, terem demonstrado bons resultados, o condicionante com maior destaque foi o NaNO3, sendo este portanto o mais recomendado para as sementes de mamona cv. BRS-Energia.
salt stress on germination and seedling establishment. Leader: Enéas Gomes Filho. Fortaleza: UFC. 90f. (dissertation)
ABSTRAT
Castor bean (Ricinus communis) is a rise culture that needs production techniques appropriate to its growth conditions and niche market. Several researches have been conduct to adapt its development in the North and Northeast regions of the Brazil, which are regions with increase salinity problem. Thus, the seed and seedling salinity effects characterization and the evaluation of seed priming techniques can reduce the replanting costs and improve emergency uniformity, preparing the plant to response the salt stress. The overall objective of the research was to characterize the deleterious effects of salinity on castor bean seeds and evaluate the use of the techniques of seed priming as a means of minimizing them. Salinity affected the germination of castor bean cv. Energy only from the osmotic potential (Ψs) of -0.4 MPa, but seedling growth was affect under Ψs of -0.16 MPa. The water imbibition curve by the seed under salinity was decelerate with time of exposure to salinity. The enzymatic activity of superoxide dismutase are independent of salinity, while catalase and ascorbate peroxidase was responsive to salinity. There were no detectable enzyme activity of guaiacol peroxidase in the seeds of castor bean. However, despite osmopriming with CaCl2, NaNO2 and PEG-6000, tested under saline conditions, have shown good results, most notably the priming was NaNO3, this being the most recommended as priming to castor bean seeds.
1. INTRODUÇÃO ... 16
2. REVISÃO DE LITERATURA ... 19
2.1. A mamona ... 19
2.1.1. Cultivares de mamona ... 19
2.1.2. Aplicabilidade da Mamona ... 20
2.2. Salinidade ... 21
2.2.1. Salinidade na agricultura ... 21
2.2.2. Impacto do estresse salino sobre a cultura da mamona... 22
2.3. Estresse oxidativo... 23
2.3.1. Indicadores de estresse oxidativo ... 23
2.3.2. Atuação das ROS ... 24
2.3.3. Mecanismos enzimáticos de eliminação das ROS ... 27
2.4. Tecnologia de sementes ... 29
2.4.1. Qualidade de sementes ... 29
2.4.2. Dormência, germinação e vigor de sementes ... 30
2.4.3. Curva de embebição ... 31
2.4.4. Condicionamento de sementes ... 32
3. MATERIAL E MÉTODOS ... 35
3.1. Estratégia experimental ... 35
3.2. Aplicação dos tratamentos de quebra de dormência ... 37
3.3. Teste de germinação ... 38
3.4. Índice de velocidade de germinação (IVG) ... 38
3.5. Cálculo do tempo médio de germinação (TMG) ... 39
3.6. Testes de vigor ... 39
3.6.1. Teste de condutividade elétrica (TCE) ... 39
3.6.2. Teste de comprimento e massa seca de plântula ... 40
3.7. Determinação da umidade nos órgãos vegetais ... 40
3.8. Curva de embebição ... 40
3.9. Determinação dos teores dos íons Na+ e K+ ... 41
3.10. Determinação de lipídios... 41
3.11. Extrato para determinação dos teores de H2O2, MDA e atividade enzimática ... 41
3.12. Determinação dos teores de H2O2 ... 42
3.13. Determinação de malondialdeído (MDA) ... 42
3.14.2. Atividade enzimática da peroxidase do ascorbato ... 43
3.14.3. Atividade enzimática da peroxidase do guaiacol ... 43
3.14.4. Atividade enzimática da dismutase do superóxido... 44
3.15. Condicionamento fisiológico de sementes de mamona ... 44
3.16. Estatística ... 45
4. RESULTADOS E DISCUSSÃO ... 46
4.1. Quebra de dormência ... 46
4.2. Efeitos da salinidade sobre a germinação ... 48
4.3. Efeitos da salinidade sobre crescimento de plântula ... 51
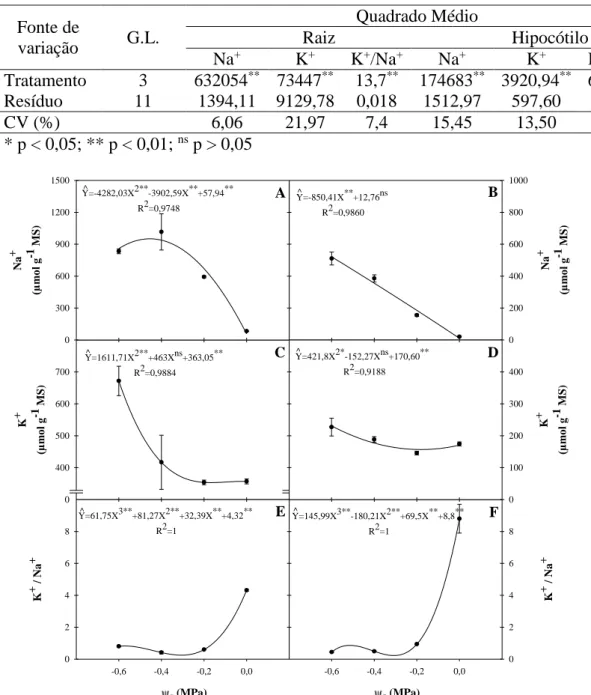
4.4. Efeitos da salinidade sobre a homeostase K+/Na+ ... 53
4.5. Efeitos da salinidade sobre a absorção de água pelas sementes... 55
4.6. Efeitos da germinação e salinidade nos teores de lipídios, H2O2 e MDA... 59
4.7. Efeitos da germinação e da salinidade na atividade das enzimas antioxidantes ... 64
4.8. Monitoramento da secagem de sementes condicionadas ... 68
4.9. Avaliação dos benefícios do condicionamento fisiológico ... 71
4.10. Efeitos da salinidade sobre germinação de sementes osmocondicionadas ... 74
4.11. Efeitos da salinidade sobre o crescimento das plântula oriundas de sementes osmocondicionadas ... 78
4.12. Efeitos da salinidade sobre a homeostase K+/Na+ em plântulas oriundas de sementes osmocondicionadas ... 80
4.13. Considerações finais... 81
5. CONCLUSÕES ... 83
com potenciais osmótico de -0,6 MPa... 74 Tabela 11 – Resumo do quadro de ANOVA para as variáveis de crescimento de plântulas oriundas de sementes osmocondicionadas de mamona sob condições salinas
(Ψs = -0,6 MPa) como: massa seca da raiz (MSR) e do hipocótilo (MSH), relação massa seca da raiz/hipocótilo (MSR/MSH), comprimento da raiz principal (CR) e do hipocótilo (CH) e relação comprimento da raiz/hipocótilo (CR/CH)... 78 Tabela 12 – Resumo do quadro de ANOVA para os teores de Na+, K+ e relação de K+/ Na+ na raiz e no hipocótilo de plântulas oriundas de sementes de mamona submetidas a
Figura 1- Padrão trifásico de capitação de água pelas sementes durante a germinação, Fonte: Bewley e Black (1978)... 32 Figura 2 – Percentual de germinação, sementes mortas e dormentes (A), índice de velocidade de germinação (IVG) (B) e tempo médio de germinação (TMG) (C), de sementes de mamona cv. BRS-Energia submetida a tratamentos pré-germinativos: (
) sementes intactas, ( ) sementes sem carúncula, ( ) sementes lixadas, ( ) sementes que sofreram choque mecânico e ( ) sementes com aplicação de ácido sulfúrico...47 Figura 3 – Percentual de germinação de sementes de mamona cv. BRS-Energia em
180 (B) horas de germinação, Fortaleza, 2013... 61 Figura 12 – Teores de malondialdeído (MDA) em sementes de mamona cv. BRS-Energia germinando em soluções salinas de NaCl com potenciais osmóticos (s) de 0 MPa (A), de -0,2 MPa (B), de -0,4 MPa (C) e de -0,6 MPa (D), em função do tempo de embebição... 62 Figura 13 – Teores de malondialdeído (MDA) em sementes de mamona cv.
BRS-Energia germinando em soluções de NaCl nos Ψs entre 0 e -0,6 MPa com 60 (A), 120 (B) e 180 (C) horas após semeadura... 63 Figura 14 – Atividade da catalase (CAT) em sementes de mamona cv. BRS-Energia durante a germinação em soluções salinas de NaCl com potenciais osmóticos (s) de 0 MPa (A), de -0,2 MPa (B), de -0,4 MPa (C) e de -0,6 MPa (D)...64 Figura 15 – Atividade da enzima catalase (CAT) em sementes de mamona cv. BRS-Energia germinando em soluções de NaCl nos Ψs entre 0 e -0,6 MPa com 24 (A), 60 (B) e 120 (C) horas de embebição... 66 Figura 16 – Atividade da peroxidase do ascorbato (APX) em sementes de mamona cv. BRS-Energia germinando em soluções salinas de NaCl com potenciais osmóticos (s) de 0 MPa (A), de -0,2 MPa (B), de -0,4 MPa (C) e de -0,6 MPa (D)... 66 Figura 17 – Atividade da peroxidase do ascorbato (APX) em sementes de mamona cv. BRS-Energia sob Ψs entre 0 e -0,6 MPa com 180 horas de embebição... 67 Figura 18 – Atividade da dismutase do superóxido (SOD) em sementes de mamona cv. BRS-Energia durante a germinação... 67 Figura 19 – Curva de secagem das sementes de mamona após seu condicionamento com H2O (A), NaCl (B), CaCl2 (C), KCl (D), NaNO2 (E), NaNO3 (F), KH2PO4 (G), NaSiO3 (H), PEG-6000 (I) e H2O2 (J)... 70 Figura 20 – Testes de vigor realizados com sementes de mamona condicionadas com H2O, NaCl, CaCl2, KCl, NaNO2, NaNO3, KH2PO4, NaSiO3, PEG-6000 e H2O2. A, condutividade elétrica (CE); B, plântulas normais (PN); C, comprimento do hipocótilo (CH); D, comprimento da raiz (CR); E, relação CR/CH; F, massa seca do hipocótilo (MSH); G, massa seca da raiz (MSR) e H, relação MSR/MSH... 72 Figura 21 – Percentual de germinação de sementes osmocondicionadas de mamona sob
1. INTRODUÇÃO
A utilização de óleos vegetais na produção de combustível renovável tem sido uma importante iniciativa nos últimos tempos para a substituição do diesel. O biodiesel produzido a partir de óleos vegetais foi introduzido na matriz energética brasileira através da Lei N° 11.097 publicada no Diário Oficial da União em 13 de janeiro de 2005. De acordo com esta lei, foi fixado em 5% o percentual mínimo obrigatório de adição do biodiesel ao óleo diesel comercializado ao consumidor final a partir de 2008.
Em setembro de 2011, o Brasil conquistou a posição de maior consumidor de
biodiesel do mundo e, atualmente, o país é o segundo maior produtor mundial, com uma
produção anual de 2,4 bilhões de metros cúbicos, estando na eminência de passar a
Alemanha, que é a maior produtora (TAGUCHI, 2012)
São muitas as espécies com potencial para fornecer óleo para a produção de biodiesel como: algodão, amendoim, babaçu, dendê, girassol, macaúba, mamona e pinhão-manso. Entretanto, 70% do biodiesel produzido no país ainda é proveniente do óleo de soja, cultura de monocultivo, centralizadora de renda e sob fortes efeitos mercadológicos, devido ao amplo uso para alimentação animal e humana (BRASIL, 2009b). No entanto, o governo brasileiro pretende tornar a mamona a principal matriz para o programa de produção de biodiesel na região norte e nordeste, onde a cultura é mais disseminada.
A estabilidade do preço e a garantia dos contratos firmados com as indústrias têm feito a cultura se expandir além de Irecê/BA (região produtora mais tradicional). O número de agricultores cultivando essa oleaginosa passou de 5 mil, em 2005, para 41,5 mil, em 2010. No estado do Ceará, onde há um programa de incentivo, já são cerca de 20 mil produtores, a maioria agricultores familiares (EMBRAPA, 2012a). Entretanto, o Brasil atualmente importa óleo de mamona e derivados, sendo que na safra 2010/2011, 5.120 toneladas de óleo de mamona chegaram aos portos brasileiros. Assim, o Brasil precisa se organizar para que a mamona seja uma cultura competitiva, aumentando a produtividade, a área plantada e a diversificação dos produtos (EMBRAPA, 2012a).
As principais cultivares recomendadas para a região Nordeste são a BRS-Nordestina, a BRS-Paraguaçu e a BRS-Energia pois apresentam um produção média de 1500 Kg ha-1 nas condições da região (EMBRAPA, 2012b). A cultivar BRS-Energia é diferente das outras duas por ser um cultivar de ciclo curto, o que contribui para redução de risco de cultivo na condição semiárida. No entanto, além das condições climáticas outros fatores podem prejudicar o desenvolvimento da cultura, sendo estes bióticos (pragas, patógenos) ou abióticos (temperatura, umidade, radiação solar, salinidade).
sodificadas. Assim, estratégias de manejo que visem aumentar a probabilidade de sucesso no cultivo da mamona nessas regiões são de grande importância.
Técnicas de condicionamento fisiológico têm sido capazes de dar mais vigor às sementes e podem ser potencialmente uma forma de reduzir custos com replantios, bem como ser útil para uniformizar a plantação e preparar as plântulas para melhor enfrentar estresses abióticos e bióticos (HARRIS et al., 2001). Segundo Conrath et al. (2002), um melhor conhecimento dos mecanismos moleculares de condicionamento fisiológico será fundamental para melhorar a capacidade das plantas de perceberem estímulos de estresses bióticos e abióticos mais rapidamente, de modo a lidar com diferentes formas de estresse de forma mais eficiente e de uma forma natural.
Diante do exposto, o objetivo geral da pesquisa foi caracterizar os efeitos deletérios da salinidade na germinação das sementes e estabelecimento das plântulas de mamona e avaliar o uso das técnicas de condicionamento fisiológico de sementes como meio de minimizá-los.
Pretende-se especificamente nessa pesquisa:
Determinar a melhor metodologia de quebra de dormência para sementes de mamona cv. BRS-Energia;
Caracterizar os efeitos da salinidade sobre a germinação de sementes de mamona cv. BRS-Energia;
Determinar quais enzimas atuam no combate as espécies reativas de oxigênio durante a germinação e quais enzimas respondem ao estresse salino;
2. REVISÃO DE LITERATURA
2.1. A mamona
A mamoneira (Ricinus communis L.) pertence à classe das Dicotiledôneas, família Euphorbiaceae (WEISS, 1983), é uma planta oleaginosa de relevante importância econômica e social. É bastante tolerante à escassez de água, mas exigente em calor e luminosidade. Está disseminada em quase todo o Nordeste brasileiro, local cujas condições climáticas são propícias ao seu desenvolvimento e crescimento da mamoneira
(EMBRAPA, 2012b).
2.1.1.Cultivares de mamona
Existem várias cultivares de mamoneira disponíveis para o plantio em nosso país, variando em porte, deiscência dos frutos, tipo dos cachos e outras características. Conforme EMBRAPA (2004), a pesquisa com cultivares começou no Estado de São Paulo, em 1937, com os trabalhos desenvolvidos pelo Instituto Agronômico de Campinas (IAC). Com as seleções realizadas, o IAC lançou a cultivar IAC 38, de porte anão, frutos deiscentes, ciclo de 200 dias (em média) nas condições do Estado de São Paulo, e capacidade para produzir até mais de 2 t/ha de bagas (sistema de sequeiro), com 41% de óleo nas sementes e de frutos deiscentes.
Em 1974, a Empresa Baiana de Desenvolvimento Agrícola (EBDA) lançou no mercado várias cultivares para as condições do Nordeste, em que as principais foram: Sipeal 1, Sipeal 9, Sipeal 28 e Sipeal 12. Dessas, a de maior destaque foi a Sipeal 28, de porte médio, frutos deiscentes, teor de óleo nas sementes de 47,3% e produtividade média em condições de sequeiro de 1300 kg/ha de bagas. Em 1998 e 1999, a EMBRAPA lançou as cultivares BRS-149 Nordestina e BRS-188 Paraguaçu para as condições edafoclimáticas do semi-árido nordestino. Tratavam-se de cultivares adaptadas para a cultura familiar do Nordeste, pois apresentava frutos semideiscentes, que não liberam totalmente as sementes após maduras, facilitando a colheita manual.
Em 2007, a Embrapa algodão apresentou a variedade BRS-Energia, caracterizada por sua precocidade no cultivo, que permite ao agricultor uma colheita de cachos na metade de tempo previsto para a colheita das outras cultivares disponíveis no mercado. A BRS-Energia foi fruto de uma pesquisa iniciada em 2000, com experimentos feitos na Paraíba, Ceará, Rio Grande do Norte e Rondônia (EMBRAPA, 2012c).
2.1.2. Aplicabilidade da Mamona
O óleo extraído das sementes de mamona é solúvel em álcool e possui inúmeras aplicações na indústria, sendo usado na fabricação de plásticos, fibras sintéticas, tintas, esmaltes, entre outros (FONSECA et al., 2004). Além disso, também tem seu uso na biomedicina, na elaboração de próteses, com destaque em cirurgias ósseas de mama e de próstata (BDMG, 2000).
têm despertado interesse como uma alternativa promissora para a produção de biodiesel, um combustível biodegradável, renovável e ecologicamente correto.
O óleo de mamona é rico em ácido ricinoléico (ácido 12-hidróxi-cis-octadeca-9-enóico), o qual representa quase 90% do total de ácidos de graxos da semente. Essa composição confere ao biodiesel de mamona uma excelente estabilidade à variação de temperatura, não congelando em baixas temperaturas, e com boa estabilidade oxidativa, já que a autoxidação depende do número e da localização de ligações duplas de metilenos interrompidos (MOSER, 2009). Além disso, o óleo da mamona tem 30% a mais de lubricidade que os outros óleos, podendo reduzir a emissão de diversos gases causadores do efeito estufa, a exemplo do gás carbônico e enxofre. Conclui-se, portanto, tratar-se de um óleo especial e com mercado garantido no mundo moderno (BELTRÃO, 2003). Em contrapartida, apresenta altos valores de massa específica e viscosidade cinemática que limitam seu uso como biocombustível (SILVA et al., 2010).
2.2.Salinidade
2.2.1. Salinidade na agricultura
nessa área o semiárido nordestino (FAO, 2010). Segundo Chinnusamy, Jagerndorf e Zhu (2005), a maioria das culturas é afetada quando a condutividade elétrica do extrato de saturação do solo (CEes) é superior a 3 dS m-1. Mas, para outros autores, isso ocorre quando a condutividade elétrica do extrato de saturação é igual ou superior a 2 dS m-1 (HOLANDA et al., 2010).
Solos salinos são os que possuem pH inferior a 8,5, percentagem de sódio trocável (PST) inferior a 15% e condutividade elétrica do extrato de saturação (CEes) entre a 4 dS m-1 e 7dS m-1, e solo com CEes acima de 7 dS m-1 é considerado sálico e, letalmente tóxico, à maioria das plantas (RICHARDS, 1954; SCHNEIDER; SPERA, 2009). É importante destacar, que o grau das injurias causadas pela salinidade do solo dependem da espécie vegetal, variedade, estágio de crescimento, fatores ambientais e da natureza do sal, o que significa em si, um problema para se definir solos salinos com precisão (TAIZ; ZEIGER, 2013).
O sal provoca três ações estressantes sobre os tecidos e órgãos das plantas superiores: déficit hídrico, como resultado da alta concentração de íons no ambiente radicular; estresse iônico, o qual decorre, em grande parte, de alterações nas relações Na+/K+ e de concentração excessiva de íons que são prejudiciais ao metabolismo celular (HORIE; SCHROEDER, 2004); e desequilíbrio nutricional, causado pelos distúrbios na absorção ou distribuição dos nutrientes (YAHYA, 1998).
2.2.2. Impacto do estresse salino sobre a cultura da mamona
Como todas as outras espécies de planta, a mamona tem limites suportáveis de salinidade. Conforme Cavalcanti et al. (2004) e Cavalcanti et al. (2005),a razão de área foliar, a eficiência quântica do fotossistema II, a altura, o número de folhas e o diâmetro de caule de plantas de mamona cv. BRS-149 Nordestina, com até trinta dias de emergência, não são afetados pela salinidade da água de irrigação de até 4,7 dS m-1. Contudo, ocorrem decréscimos na fitomassa da parte aérea (6,0%), na área foliar (6,6%) e no consumo de água pelas plantas (6,3%) com o aumento unitário da condutividade elétrica da água de irrigação (CEa). Similarmente, níveis de salinidade entre 0,8 e 4,8 dS m-1 não afetaram o crescimento em altura das plantas de mamoneira cv. BRS-Energia até os primeiros 50 dias após a emergência. Entretanto, níveis de salinidade iguais ou a superiores a 4 dS m-1 após 50 dias de emergência, começaram a afetar o crescimento das plantas, reduzindo assim suas alturas (SANTOS et al., 2010).
2.3. Estresse oxidativo
2.3.1. Indicadores de estresse oxidativo
O estresse oxidativo é definido como um aumento no nível das espécies reativas de oxigênio (ROS, do inglês, Reactive Oxygen Species), que ocorre em função do desequilíbrio na relação entre compostos antioxidantes e compostos oxidantes (ZHU, 2002). As ROS mais comumente encontradas são o oxigênio singleto (1O2), o peróxido de hidrogênio (H2O2) e os radicais superóxido (•O2-) e hidroxil (HO•). Elas são consideradas subprodutos inevitáveis do metabolismo celular aeróbico e dos processos fotoxidativos (NOCTOR; FOYER, 1998) e sua produção ocorre principalmente nas mitocôndrias (MØLLER, 2001), nos cloroplastos (FOYER; NOCTOR, 2000) e nos peroxissomos (DEL RIO et al., 2002).
reduzindo o superóxido a H2O2. O superóxido é produzido nas mitocôndrias quando a taxa de entrada de elétrons na cadeia respiratória e sua taxa de transferência não são coordenadas, sendo os elétrons transferidos ao oxigênio pela ubiquinona parcialmente reduzida nos complexos I e III. O superóxido também pode ser produzido no apoplasto pela ação da enzima NADPH oxidase localizada na plasmalema (NELSON; COX, 2011).
2.3.2. Atuação das ROS
O peróxido de hidrogênio e outras ROS eram vistas unicamente como metabólitos tóxicos para a célula. Isto porque, quando em altas concentrações, essas moléculas são tóxicas, capazes de ocasionar danos aos diversos componentes das células, como DNA, lipídios e proteínas (APEL; HIRT, 2004). Sob condições ótimas de crescimento, sua produção é mantida em baixos níveis. Entretanto, sob condições adversas, como nos estresses hídrico e salino, sua taxa de produção pode ser drasticamente aumentada (VAIDYANATHAN et al., 2003). Porém, estudos têm demonstrado que as ROS, especialmente o H2O2, atuam como moléculas sinalizadoras e são produzidas e controladas pelo metabolismo, sendo benéficas a baixas concentrações e prejudiciais quando em excesso (UCHIDA et al., 2002; GECHEV; HILLE, 2005; QUAN et al., 2008).
2002). Em função disso, as plantas desenvolveram mecanismos para manter a relação produção/eliminação de ROS constante no interior das células (APEL; HIRT, 2004), utilizando para isso sistemas de defesa enzimáticos e não enzimáticos (ASADA, 1999).
O peróxido de hidrogênio (H2O2) é um componente vital para o desenvolvimento, metabolismo e homeostase de diferentes organismos (BIENERT et al., 2006). Nas plantas, o H2O2 é uma das ROS mais estáveis, estando envolvida nos processos de aclimatação, defesa e desenvolvimento (SLESAK et al., 2007). O H2O2 pode atuar como um sinalizador por ter meia-vida longa quando comparada às de outras ROS, e não ser, diferentemente, por exemplo do superóxido, um radical livre e não apresentar carga (HALLIWELL, 2006). Dessa forma, o mesmo pode atuar como uma molécula sinalizadora capaz de percorrer longas distâncias até o seu alvo (VRANOVÁ et al., 2002). Além disso, canais transportadores de água na membrana plasmática, conhecidos como aquaporinas, podem facilitar o movimento de H2O2 transmembranar (HENZLER; STEUDEL, 2000).
De acordo com Bienert et al. (2006), o H2O2 atua nas células como um mensageiro secundário, aumentando o fluxo de íons Ca2+ e modificando o padrão de proteínas e de expressão gênica. Para Agarwal et al. (2005), o H2O2 produzido em resposta à aplicação exógena de Ca2+ ou de moléculas sinalizadoras (ABA e ácido salicílico) pode induzir a síntese ou ativação de fatores de transcrição que estão associados à indução de várias enzimas antioxidativas.
como sinalizador ou se a oxidação de outras moléculas pelo mesmo é necessária para gerar o sinal intracelular (DESIKAN et al., 2003).
O H2O2 atua como regulador de vários processos fisiológicos, como o fortalecimento da parede celular, a senescência, a produção de fitoalexinas, a fotossíntese, a abertura estomática e o ciclo celular. Contudo, os efeitos biológicos do H2O2 mostram-se dependentes não apenas de sua concentração, mas também do seu sítio de produção, do estádio de desenvolvimento da planta e da prévia exposição da planta a outros tipos de estresse (PETROV; VAN BREUSEGEM, 2012).
O radical hidroxila causa danos ao DNA, RNA, às proteínas, lipídios e membranas celulares do núcleo e mitocondrial. No DNA ele ataca tanto as bases nitrogenadas quanto a desoxirribose. O ataque ao açúcar pode ser realizado por abstração de um dos átomos de hidrogênio e quase sempre leva à rupturada cadeia de DNA (BARREIROS et al., 2006).
A forma mais deletéria do oxigênio ao organismo é o oxigênio singleto (1O2), pois é a causa ou o intermediário da toxicidade fotoinduzida do O2 em organismos vivos. Os compostos naturais mais reativos frente ao 1O2 são oscarotenóides, devido as múltiplas insaturações conjugadas. Assim, o 1O2 reage mais lentamente com os ácidos graxos que com o β-caroteno, e quanto maior o número de insaturações presentes nos ácidos graxos, mais rapidamente eles irão reagir. Essa reação se dá por incorporação do oxigênio à cadeia com consequente migração da ligação dupla, formando ácidos hidroperóxidos (BARREIROS et al., 2006).
redução da ubiquinona para ubiquinol, no interior da mitocôndria. Por fim, o radical ânion superóxido funciona como sinalizador molecular através da sua capacidade de oxidar grupos –SH em ligações dissulfeto, podendo ativar e desativar enzimas que contenham metionina (BARREIROS et al., 2006).
2.3.3. Mecanismos enzimáticos de eliminação das ROS
As plantas possuem mecanismos específicos para a eliminação das ROS, os quais incluem a ativação de enzimas, como a catalase (CAT), as peroxidases e a dismutase do superóxido (SOD), bem como aquelas que participam do ciclo ascorbato-glutationa [peroxidase do ascorbato (APX), redutase da glutationa (GR), redutase do monodesidroascorbato (MDHAR) e redutase do desidroascorbato (DHAR)] (FOYER; NOCTOR, 2003).
A SOD é uma metaloproteína responsável pela dismutação do radical superóxido em peróxido de hidrogênio e oxigênio molecular, sendo considerada a primeira linha de defesa das plantas contra o estresse oxidativo (ALSCHER; ERTURK; HEATH, 2002). Nas plantas superiores, a SOD apresenta-se na forma de três isoenzimas: Cu/Zn-SOD, Mn-SOD e Fe-SOD que são classificadas de acordo com o íon metálico no grupo prostético (ASADA, 1999; ARORA; SAIRAM; SRIVASTAVA, 2002). Além disso,
Scandalios (2002) e Ashraf (2009) afirmam que existe ainda um quarto tipo de SOD, cujo íon metálico é o níquel (Ni).
correlacionaram com a subsequente tolerância dessa espécie à salinidade (PARIDA; DAS; MOHANTY, 2004).
Meloni et al. (2003), trabalhando com duas cultivares de algodão com tolerância diferencial à salinidade, mostraram que a cultivar mais tolerante apresentou maior indução do sistema enzimático antioxidativo envolvendo a SOD, a GR e a G-POD (peroxidase do guaiacol, também conhecida como GPX), que permitiu menor dano oxidativo, como comprovado pela menor taxa de peroxidação de lipídios de membrana, em comparação à cultivar sensível.
O H2O2 gerado a partir da atividade da SOD pode ser tóxico e precisa ser removido da célula em reações subsequentes. Nas plantas, as enzimas APX, CAT e GPX são os principais reguladores dos níveis intracelulares de H2O2 (FOYER; NOCTOR, 2003). Azevedo Neto et al. (2005), trabalhando com plantas de milho sob condições de estresse salino, mostraram que as atividades dessas enzimas aumentaram substancialmente no genótipo tolerante, em relação ao genótipo sensível, indicando que sob estresse, o mecanismo removedor de H2O2 é mais eficaz no genótipo tolerante. Além disso, esses autores afirmaram que a CAT foi a enzima removedora de H2O2 mais importante nas folhas, enquanto a GPX foi a mais importante nas raízes.
Da mesma forma que a CAT, a APX desempenha um papel vital na defesa das plantas contra o estresse oxidativo. A APX utiliza especificamente o ascorbato como doador de elétrons na reação de eliminação do H2O2, protegendo as células contra os efeitos nocivos dessa substância (ASADA, 1992; SHIGEOKA et al., 2002, FOYER; NOCTOR, 2005). A APX está localizada em diversos compartimentos celulares, como citosol, cloroplastos, glioxissomos, mitocôndrias e peroxissomos (SHIGEOKA et al., 2002). Diferentes isoformas de APX são encontradas nas plantas, podendo ser solúveis, como as encontradas no citosol e estroma, ou associadas às membranas, como aquelas presentes nos microcorpos, tilacóides e mitocôndrias (ASADA, 1992).
2.4. Tecnologia de sementes
2.4.1. Qualidade de sementes
Sementes de qualidade são aquelas com capacidade de apresentarem bom desempenho no campo, com altos níveis de germinação, sob as mais variadas condições, originando plantas vigorosas e potencialmente produtivas no menor tempo possível (CARVALHO; NAKAGAWA, 2000).
2.4.2. Dormência, germinação e vigor de sementes
A dormência pode ser definida como uma falha de uma semente intacta e viável em germinar sob condições aparentemente favoráveis à germinação (DE CASTRO; HILHORST, 2000). Entretanto, o impedimento estabelecido pela dormência também pode encarado como uma estratégia benéfica, pela distribuição da germinação ao longo do tempo, aumentando a probabilidade de sobrevivência da espécie (FOWLER; BLANCHETTI, 2000).
As sementes de mamona apresentam germinação lenta e desuniforme (OLIVEIRA et al., 2004), ficando por mais tempo expostas aos patógenos do solo e às intempéries, o que resulta em estande final irregular. A irregularidade na emergência das plântulas de mamona em campo, e também em laboratório, tem sido atribuída à dificuldade de absorção de água pelas sementes, devido à espessura e rigidez do tegumento ou a uma possível dormência pós-colheita (LAGO et al., 1979). Lago et al., (1979) ao avaliarem a qualidade fisiológica de sementes de mamona, cultivares Guarani e IAC-38, constataram variação de 9 a 66% de sementes não germinadas.
É importante ressaltar que outros fatores influenciam a emergência das plântulas de mamona, tais como: temperatura, umidade, características físicas do solo, profundidade de plantio e disponibilidade de oxigênio, salinidade, presença de insetos e patógenos (VILELA et al., 1991). A salinidade também pode contribuir para uma queda no percentual de emergência ou na velocidade de emergência, pelo efeito de redução do potencial osmótico do solo e também pelo efeito tóxico de seus íons.
emergência das plantas em campo apresentam significativos reflexos sobre a produção. A uniformidade de desenvolvimento das plantas pode ser afetada por fatores como compactação do solo, contato com herbicidas, profundidade e densidade de semeadura, distribuição desuniforme de fertilizantes e pela desuniformidade de emergência de plântulas, entre outros fatores.
2.4.3. Curva de embebição
O processo de embebição das sementes é físico, relacionado basicamente às
propriedades coloidais dos seus constituintes e às diferenças de potencial hídrico (Ѱw)
entre a semente e o meio externo. O potencial hídrico é composto pelo potencial de
pressão (Ѱp), potencial osmótico (Ѱs), potencial gravitacional (Ѱg) e o potencial
matricial (Ѱm) (BEWLEY; BLACK, 1978).
O potencial de pressão (Ѱp) numa célula ocorre porque a entrada de água aumenta
o conteúdo celular, manifestando uma pressão contraria ao fluxo de água devido a resistência a expansão da parede celular (REICHARDT, 1985). O potencial osmótico
(Ѱs) representa à capacidade dos solutos, quando dissolvidos, de reduzir a energia livre do sistema, enquanto o potencial matricial (Ѱm) representa a capacidade de sólidos ou substâncias insolúveis, de reduzirem o Ѱw. O componente matricial é particularmente importante em estágios iniciais de absorção de água pelas sementes secas, enquanto ainda há pouca atuação do componente osmótico por não haver plena embebição da semente (PIMENTA, 2004).
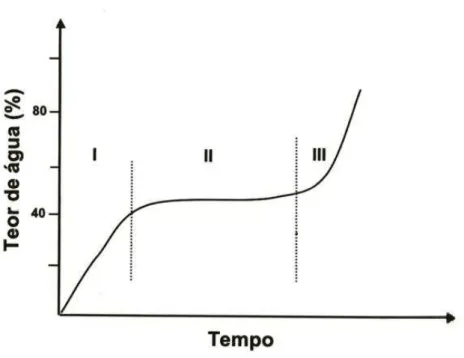
velocidade de hidratação, sendo sua ocorrência dependente da espécie estudada. Na terceira fase, torna-se visível a retomada de crescimento do embrião pela protrusão da raiz principal (Figura 1).
Figura 1- Padrão trifásico de capitação de água pelas sementes durante a germinação, Fonte: Bewley e Black (1978).
2.4.4. Condicionamento de sementes
O potencial fisiológico pode ser recuperado com a absorção de água pela semente durante as fases I e II, sem permitir a protrusão radicular ou início da fase III, do modelo trifásico de Bewley e Black (1978). A hidratação ativa a digestão das reservas e sua translocação e assimilação, levando as sementes do lote a um estado metabólico relativamente uniforme (BEWLEY; BLACK, 1982).
A expressão seed enhancement é utilizada no meio industrial para nomear um conjunto de técnicas ou processos destinados a realçar a qualidade ou beneficiar o desempenho de lotes de sementes e/ou plântulas produzidas (TAYLOR, 1998). Não há, ainda, uma proposta concreta para a tradução da expressão seed enhancement Marcos Filho (2005), sugere o uso da expressão condicionamento de sementes para nominar este conjunto de técnicas. O condicionamento fisiológico (priming) é apenas uma técnica do condicionamento de sementes, que visa elevar o potencial fisiológico da semente através de um pré-umedecimento.
A eficiência da técnica foi comprovada em várias espécies como alface (EIRA; MARCOS FILHO, 1990), milho-doce (PARERA; CANTLIFFE, 1994), brócolis (JETT
et al., 1996), berinjela (TRIGO; TRIGO, 1999), algodão (QUEIROGA et al., 2011), girassol (BARROS; ROSSETTO, 2009), cenoura (CARNEIRO et al., 1999), pimentão (ROVERI JOSÉ et al., 2000) e cebola (CASEIRO et al., 2004).
3. MATERIAL E MÉTODOS
3.1.Estratégia experimental
A pesquisa foi realizada no Laboratório de Fisiologia Vegetal, localizado no Departamento de Bioquímica e Biologia Molecular da Universidade Federal do Ceará. As sementes de mamona utilizadas foram da variedade BRS-Energia safra 2011 fornecidas pela EMBRAPA Agroenergia.
O estudo das sementes de mamona foi divido em cinco etapas: (1) Quebra de dormência; (2) Caracterização do estresse salino sobre a semente e plântula de mamona; (3) Caracterização do perfil de lipídios, de H2O2 e de peroxidação de lipídios de membrana durante a germinação e sob condições salinas; (4) Ação das enzimas antioxidativas; e a (5) Ação do condicionamento fisiológico de sementes sobre os efeitos do estresse salino.
Em todas as etapas, as sementes foram postas para germinar em câmaras de germinação tipo BOD, com temperatura a 25±1 °C e fotoperíodo de 12 h de luz e 12 h de escuro, em delineamento experimental inteiramente casualizado (DIC).
Na primeira etapa, quebra de dormência, as sementes de mamona foram pré-tratadas, e logo após submetidas a um teste de germinação e cálculo do índice de velocidade de germinação (IVG) e do tempo médio de germinação (TMG) com o objetivo de avaliar a eficiência do pré-tratamento.
tratamentos salinos foram utilizadas soluções de NaCl com Ψs de 0; 0,2; 0,4; 0;6 e -0,8 MPa, os quais foram calculados segundo a equação de potencial osmótico de Van't Hoff (Tabela 1).
Equação de Van't Hoff:
Ψs = -RTCsI I = 1 + α(q-1) Onde:
Ψs = pressão osmótica da solução (0,987 ≈ 1 atm = 0,1 Mpa);
R = constante universal dos gases perfeitos (0,08205 L atm mol-1 K-1); T = temperatura em Kelvin (TºK = TºC + 273);
Cs = concentração do soluto (M ou mol L-1); I = fator de correção de Van't Hoff;
α = grau de ionização;
q = número total de íons liberados na ionização de um composto (q=2 para o NaCl).
Tabela 1–Tabela de conversão do Ψs com a concentração de NaCl e a condutividade elétrica (CE)
Ψs Cloreto de sódio (NaCl) CE*
MPa g/L mM dS m-1
-0,2 2,36 40,36 4,06
-0,4 4,72 80,72 8,12
-0,6 7,08 121,09 12,19
-0,8 9,44 161,45 16,25
*Condutividade elétrica calculada a partir de um curva padrão com NaCl.
lipídios de membrana e sua relação com a produção de H2O2, sendo esta consequente ou
não do processo de β-oxidação de ácidos graxos.
Na quarta etapa, ação das enzimas antioxidativas, foram feitas determinações da atividade das enzimas catalase (CAT), peroxidase do ascorbato (APX), peroxidase do guaiacol (GPX ou G-POD) e dismutase do superóxido (SOD), durante a germinação e
sob os Ψs de 0 a -0,6 MPa. O objetivo dessa etapa foi determinar que enzimas estão envolvidas no combate às espécies reativas de oxigênio nas sementes de mamona e quais delas respondem ao estresse salino.
Na quinta etapa, ação do condicionamento fisiológico de sementes sobre os efeitos do estresse salino, as sementes de mamona foram condicionadas fisiologicamente por 24 h com soluções de NaCl, CaCl2, KCl, NaNO2, NaNO3, KH2PO4, NaSiO3 e PEG-6000 a Ψs de -0,2 MPa, com H2O2 a 10 mM e com H2O. Após a aplicação dos condicionantes, as sementes foram secas em BOD com temperatura de 25±1 °C e 50% de umidade relativa até retornarem a umidade inicial.
Com essas sementes, foram realizados testes de vigor (teste de comprimento de plântula e de condutividade elétrica (TCE)) e foram determinados o percentual de plântulas normais e massa seca do hipocótilo e raiz, na ausência de salinidade, para se selecionar os condicionantes que produzissem efeitos benéficos sobre as sementes de mamona. As sementes com os condicionantes selecionados foram submetido ao Ψs de -0,6 MPa da solução de NaCl para realização de um teste de germinação e de vigor, sendo também realizado o cálculo do índice de velocidade de germinação (IVG) e o tempo médio de germinação (TMG), além de se determinar os teores de Na+ e K+ com o objetivo de se avaliar a atuação dos condicionantes selecionados em condições salinas e selecionar o melhor dentre eles.
Os pré-tratamentos para quebra de dormência foram com (1) remoção da carúncula: a carúncula de cada semente foi removida com o auxílio de um estilete, deixando-se a micrópila; (2) escarificação do tegumento com lixa: utilizando-se uma
lixa d’água n°1 efetuou-se a escarificação nos quatro lados do tegumento (BRASIL, 2009a); (3) escarificação do tegumento por choque mecânico: utilizando-se um recipiente hermeticamente fechado, sementes e pedrisco foram agitados a fim de provocar choque entre eles para causar escarificação do tegumento; e (4) escarificação química: utilizou-se o H2SO4 concentrado por um tempo de exposição de 5 min (suficiente para amolecer o tegumento), e depois as sementes foram lavadas com água corrente pelo mesmo tempo a que ficaram expostas ao ácido (BRASIL, 2009a).
3.3.Teste de germinação
Foram preparados 8 repetições de 50 sementes postas para germina em papel tipo germitest umedecido com água destilada ou solução de NaCl em quantidade equivalente a 2,5 vezes ao seu peso. Os rolos formados foram acondicionados em câmaras de germinação tipo BOD sob condições controladas, com temperatura a 25±1 °C e fotoperíodo de 12 h de luz e 12 h de escuro simulando as condições fotoperiódicas encontradas próximo à linha do Equador durante todo ano. A primeira contagem foi realizada no sétimo dia de embebição e novas contagens diárias até o décimo quarto dia (BRASIL, 2009a), com exceção da primeira contagem realizada na quinta etapa que foi realizada no quarto dia prevendo-se que os condicionantes pudessem acelerar o início da germinação. Foram consideradas mortas às sementes que, ao longo do teste, apresentavam-se necrosadas. Por outro lado, as sementes foram consideradas dormentes quando, ao final do teste, não foram capazes de germinar. As plântulas consideradas normais não apresentavam falta de partes ou deformações na formação de seus órgãos.
O IVG foi calculado pelo somatório do número de sementes germinadas a cada dia, dividido pelo número de dias decorridos entre a semeadura e a germinação, de acordo com a fórmula definida por Maguire (1962).
IVG = ∑n NiGi
i=1 , onde
Gi = número de sementes germinadas a cada dia de contagem; Ni = número de dias decorridos da semeadura;
n = n-ézimo tempo de contagem.
3.5. Cálculo do tempo médio de germinação (TMG)
O TMG foi calculado pelo somatório do produto do número de sementes germinadas com o número de dias decorridos entre a semeadura e a germinação dividido pelo somatório do número das sementes germinadas, a cada dia, através da fórmula proposta por Labouriau (1983), sendo os resultados expressos em dias.
TMG = ∑n Gi ∗ Ni
i=1 ⁄∑ Gn, onde
Gi = número de sementes germinadas a cada dia de contagem;
Ni = tempo decorrido entre o início da germinação e a n-ésima contagem; n = n-ézimo tempo de contagem.
3.6.Testes de vigor
3.6.1. Teste de condutividade elétrica (TCE)
3.6.2. Teste de comprimento e massa seca de plântula
Foram semeadas 10 sementes, em rolo de papel, dispostas alternadamente, com a carúncula voltada para baixo, seguindo o mesmo procedimento do teste de germinação, mas com avaliação, para mamona, apenas no décimo quarto dia. O princípio deste teste considera que sementes mais vigorosas apresentam plântulas também mais vigorosas (VIEIRA; KRZYZANOWSKI, 1999). Assim, o vigor das plântulas é avaliado por meio do comprimento e massa seca do hipocótilo e raiz.
As medidas de comprimento foram realizadas com auxílio de paquímetro digital. Para determinar a massa seca, as plântulas normais foram colocadas em sacos de papel devidamente identificadas e levadas para secar em estufa com circulação forçada de ar, regulada à temperatura de 105±2°C por 24 h.
3.7.Determinação da umidade nos órgãos vegetais
Os órgãos vegetais (hipocótilo e raiz) foram postos para secar em estufa à temperatura de 105±2°C por 24 h. Essa determinação foi usada para cálculos de concentração e atividade enzimática com base em massa seca (MS).
3.8.Curva de embebição
As sementes foram postas para embeber em caixas tipo Gerbox (11 cm x 11 cm x 3 cm). Cada tratamento de solução de NaCl (0; -0,2; -0,4 e -0,6 MPa) continha dez repetições com trinta sementes. As sementes, nas caixas Gerbox, foram postas em câmaras de germinação tipo BOD, sob condições controladas, com temperatura de 25±1 °C e fotoperíodo de 12 h de luz e 12 h de escuro.
3.9.Determinação dos teores dos íons Na+ e K+
Para a determinação dos teores dos íons inorgânicos, 10 mg do pó seco de hipocótilo e de raízes foram homogeneizados com 1,0 mL de água desionizada, em tubos de Eppendorfs, os quais foram mantidos à temperatura ambiente de 25±3 °C, durante 1 h, com agitações a cada 10 min. Após esse tempo, as amostras foram centrifugadas a 3.000 x g por 15 min, à temperatura de 25°C. O sobrenadante foi coletado e armazenado a -25°C até sua utilização, sendo o precipitado descartado. Os teores de Na+ e K+ foram determinados segundo Malavolta, Vitti e Oliveira (1989), com o auxílio de um fotômetro de chama [Micronal, modelo B462 (São Paulo/SP, Brasil)]. Os teores iônicos foram expressos em µmol g-1 MS.
3.10. Determinação de lipídios
Os lipídios foram extraídos quimicamente com hexano a frio. As amostras de 0,2 g de massa seca foram misturados com 10 mL de hexano por 24 h e depois por mais 20 min para remoção dos lipídios ainda junto ao macerado. A cada tempo de exposição, as amostras foram centrifugados e os sobrenadantes colocados em um tubo de ensaio de massa conhecida. Para determinação do teor de lipídios, deixou-se o hexano evaporar à temperatura ambiente de 25°C, sendo os tubos novamente pesados até que o peso se mantivesse constante. Por diferença com o peso conhecido do tubo de ensaio, estimou-se a quantidade de lipídios extraído. O teor de lipídios foi expresso em mg g-1 de MS.
3.11. Extrato para determinação dos teores de H2O2, MDA e atividade
enzimática
6oC para se obter um sobrenadante límpido (extrato). Todas as etapas do processo de extração foram realizadas a uma temperatura de aproximadamente 6°C. O extrato foi armazenado em ultrafreezer, à temperatura de -80°C, até sua utilização.
3.12. Determinação dos teores de H2O2
Os teores de H2O2 foram determinados conforme Sergiev, Alexieva e Karanov (1997). O meio de reação foi constituído de 0,5 mL de extrato; 0,5 mL de tampão fosfato de potássio a 10 mM, pH 7; e 1 mL de KI a 1 M. A mistura foi agitada e mantida em repouso ao abrigo da luz, à temperatura de 25°C por 60 min. Em seguida, procedeu-se a quantificação dos teores de H2O2 através das medidas de absorbância a 390 nm, utilizando-se uma curva padrão feita com concentrações conhecidas de H2O2. Os resultados foram expressos em µmol g-1 MS. A MS foi estimada a partir da relação de massa fresca e massa seca das sementes.
3.13.Determinação de malondialdeído (MDA)
3.14.Determinação de enzimas antioxidantes
3.14.1.Atividade enzimática da catalase
A atividade total da CAT (CE 1.11.1.6) foi medida de acordo com Beers e Sizer (1952), com pequenas modificações. Para o meio de reação, a 1290 L de tampão fosfato de potássio a 0,1 M, contendo EDTA a 0,1 mM (pH 7,0) a 30°C, foram adicionados 60 L de H2O2 a 0,5 M e 150 L do extrato enzimático. Após 1 min de reação a atividade enzimática da CAT foi determinada por meio do desaparecimento do H2O2, sendo mensurada a partir de leituras de absorbância em 240 nm e usando o coeficiente de extinção molar do H2O2 (36 M-1 cm-1). Os resultados foram expressos em mmol H2O2 g-1 MS min-1.
3.14.2.Atividade enzimática da peroxidase do ascorbato
A atividade total da APX (EC 1.11.1.1) foi determinada de acordo Nakano e Asada (1981). Para o meio de reação, a 1100 L de tampão fosfato de potássio a 50 mM, contendo EDTA a 50 µM (pH 6,0) a 30°C, foram adicionados 300 L do extrato enzimático, 50 L de H2O2 a 30 mM e 50 L de ácido ascórbico a 15 mM. Após 2 min de reação, a oxidação do ácido ascórbico foi determinada medindo-se a absorbância a 290 nm e utilizando seu coeficiente de extinção molar (2,8 mM-1 cm-1). Os resultados foram expressos em mmol H2O2 g-1 MS min-1, considerando a relação de 2 moles de ascorbato para 1 mol de H2O2.
3.14.3.Atividade enzimática da peroxidase do guaiacol
A atividade total da GPX (EC 1.11.1.7) foi determinada tal como descrito por
do guaiacol foi determinada medindo-se a absorbância a 470 nm e utilizando seu coeficiente de extinção molar (26,6 mM-1 cm-1).
3.14.4.Atividade enzimática da dismutase do superóxido
A atividade total da SOD (EC 1.15.1.1) foi determinada medindo a sua capacidade para inibir a redução fotoquímica do azul de nitrotetrazólio (NBT), como descrito por Giannopolitis e Ries (1977). Para o meio de reação, a 1000 L de tampão fosfato de potássio a 0,05 M (pH 7,8), contendo EDTA a 0,1 mM e metionina a 19,5 mM, foram adicionados 50 L do extrato enzimático, 150 L de NBT a 750 µM e 300
L de riboflavina a 10 µM. A reação foi conduzida a 25°C numa câmara iluminada com duas lâmpadas fluorescentes de 20 W, por 15 min. Uma unidade de atividade da SOD (U) foi definida como a quantidade de enzima necessária para causar uma inibição de 50% da taxa de fotorredução do NBT, sendo as leituras espectrofotométricas realizadas a 560 nm . Os resultados foram expressos em U g-1 MS.
3.15.Condicionamento fisiológico de sementes de mamona
O condicionamento foi realizado com água desionizada, soluções salinas de NaCl, CaCl2, KCl, NaNO2, NaNO3, NaSiO3, KH2PO4 no Ψs de -0,2 MPa, calculado pela equação de Van't Hoff, e conferido com o auxílio de um osmômetro (Wescor Vapor Pressure 5500), solução de PEG-6000 a Ψs igual -0,2 MPa, conforme Villela et al.
(1991) e com peróxido de hidrogênio na concentração de 10 mM (GORDIM et al., 2012).
3.16.Estatística
4. RESULTADOS E DISCUSSÃO
4.1.Quebra de dormência
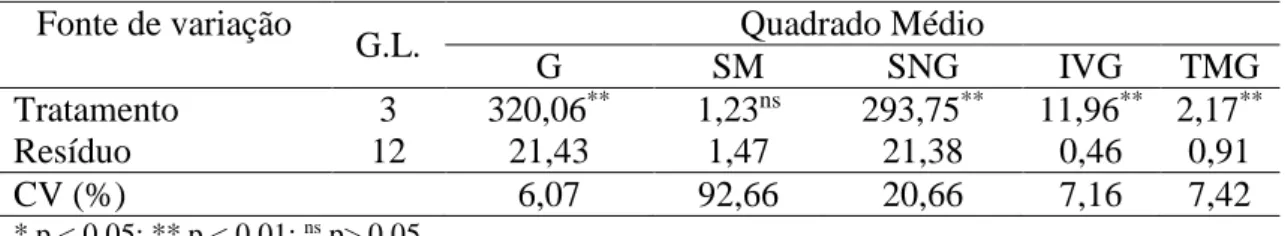
Os tratamentos pré-germinativos para quebra da dormência de sementes de mamona diferiram quanto ao percentual de germinação, de sementes não germinadas, ao índice de velocidade de germinação (IVG) e ao tempo médio de germinação (TMG) (Tabela 2).
Tabela 2 – Resumo do quadro de ANOVA para as variáveis percentual de germinação (G), de sementes mortas (SM), de sementes não germinadas (SNG); índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG) de sementes de mamona cv. BRS-Energia submetidas a tratamento para quebra de dormência
Fonte de variação
G.L. Quadrado Médio
G SM SNG IVG TMG
Tratamento 3 320,06** 1,23ns 293,75** 11,96** 2,17**
Resíduo 12 21,43 1,47 21,38 0,46 0,91
CV (%) 6,07 92,66 20,66 7,16 7,42
* p < 0,05; ** p < 0,01; ns p> 0,05
Conforme os resultados obtidos não há dormência nas sementes de mamona cv. BRS-Energia, as sementes de mamona cv. BRS-Energia apresentaram, em média, 78% de germinação e 21% de sementes não germinadas (Figura 2A).
Figura 2 – Percentual de germinação, sementes mortas e dormentes (A), índice de velocidade de germinação (IVG) (B) e tempo médio de germinação (TMG) (C), de sementes de mamona cv. BRS-Energia submetida a tratamentos pré-germinativos: ( ) sementes intactas, ( ) sementes sem carúncula, ( ) sementes lixadas, ( ) sementes que sofreram choque mecânico e ( ) sementes com aplicação de ácido sulfúrico.
As sementes que sofreram pré-tratamento com choque mecânico foram as que apresentaram maior percentual de sementes não germinadas e menor IVG (Figura 2A, 2B). Estes resultados podem ser resultado de microdanos que inviabilizaram o desenvolvimento do embrião.
As sementes sem carúncula apresentaram melhor qualidade fisiológica. Apesar do percentual de germinação dessas sementes não diferir das sementes intactas, as sementes sem carúncula germinaram mais rapidamente, sendo o IVG 21% maior e o TMG 14% menor que os das sementes intactas (Figura 2).
Segundo Sousa et al. (2009), a retirada da carúncula e a escarificação com ácido sulfúrico em sementes de mamona não aumentou a percentagem de germinação das
Pré-tratamentos
%
0 20 40 60 80 100
a a a
b
b b b b
b
b b
a
Pré-tratamentos
IV
G
0 3 6 9 12 15
Pré-tratamentos
T
M
G (dia)
0 3 6 9 12 15
b a
ab
c
ab
c b
a
A
B C
c c
Germinação Sementes mortas Sementes dormentes
variedades Al-Guarany, Mirante e Cafeista. Já em sementes de mamona cv. Al-Guarany os pré-tratamentos com lixa e com a remoção da carúncula foram capazes de reduzir o percentual de dormência (MENDES et al., 2009). Entretanto como mostra a Figura 2A,
os pré-tratamentos germinativos não reduziram, estatisticamente, o percentual de sementes de mamona cv. BRS-Energia não germinadas. Devido a tais resultados os demais ensaios foram conduzidos com sementes intactas.
4.2.Efeitos da salinidade sobre a germinação
Conforme apresentado na tabela 3, a salinidade afetou a qualidade fisiológica das sementes de mamona. Além da dormência, a germinação é afetada pelo ambiente de germinação. A salinidade é uma fonte de estresse ambiental sobre as sementes de diversas espécies, inclusive a mamona (SEVERINO et al., 2012). As sementes de mamona cv. BRS-Energia tiveram uma redução no percentual de germinação em função
da redução do Ψs, através da adição de NaCl na solução de germinação (Figura 3). Essa redução da germinação era esperada pois o estresse salino atua prejudicando a absorção de água e favorecendo a entrada excessiva de íons nas células (BRACCINI et al., 1996).
Tabela 3– Resumo do quadro de ANOVA para as variáveis percentual de germinação (G), sementes mortas (SM), sementes não germinadas (SNG), sementes germinadas na primeira contagem (PC) e plântulas normais (PN); índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG) de sementes de mamona submetidas a solução salina com potenciais osmótico entre 0 e -0,8 MPa
Fonte de variação
G.L. Quadrado Médio
G SM SNG PC PN IVG TMG
Tratamento 4 4012,7** 3* 3805,7** 5337,5** 4343,3** 90,77** 11,31**
Resíduo 15 49,73 0,73 49,93 32,93 62,53 0,90 1,36
CV (%) 13,16 171,27 15,40 14,00 15,75 13,42 13,53
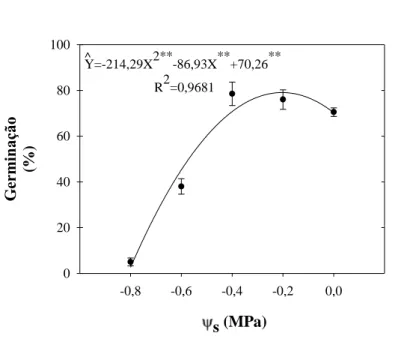
Figura 3 – Percentual de germinação de sementes de mamona cv. BRS-Energia em soluções salinas de
NaCl com potenciais osmóticos (Ψs) decrescentes.
A mamona cv. BRS-Energia não sofreu nenhum efeito deletério sobre a germinação até o Ψs de –0,4 MPa correspondente à concentração de 80 mM de NaCl. Isso foi similar ao que ocorreu com Apuleia leiocarpa (HENICKA et al., 2006). Em potencias não provocados por salinidade outras espécies apresentam menor tolerância ao efeito osmótico como as leguminosas da savana africana Combretum apiculatum, Colophospermum mopane, Acacia karroo e Acacia tortilis, que apresentam uma
redução da germinação a partir de Ψs de -0,3 MPa (CHOINSKY; TUOHY, 1993). Por outro lado, outras plantas apresentam de maior tolerância ao efeito osmótico como Prosopis juliflora, uma leguminosa da caatinga, que chega a tolerar Ψs de -1,6 MPa sem quaisquer prejuízos à germinação (PEREZ; NASSIF, 1995).
As sementes germinadas em soluções de NaCl com Ψs entre 0 e -0,4 MPa não apresentaram mortalidade ao final do teste de germinação (Figura 4A). O máximo percentual de germinação e o mínimo percentual de sementes não germinadas foram obtidos com uso de solução de NaCl a Ψs de -0,2 MPa (Figuras 3, 4B). Além disso,
quando as sementes foram postas para germinar sob Ψs até -0,4 MPa o percentual de
s (MPa)
-0,8 -0,6 -0,4 -0,2 0,0
G
er
m
in
aç
ão
(%
)
0 20 40 60 80 100
Y=-214,29X2**-86,93X**+70,26**
germinação na 1° contagem, o percentual de plântulas normais e o IVG foram iguais ou superiores, enquanto o TMG foi igual ou inferior ao de sementes postas para germinar a
Ψs de 0 MPa (Figura 4C, 4D, 4E, 4F). Entretanto, as sementes de mamona cv. BRS-Energia se mostraram melhor qualidade fisiologica quando postas para germinar em solução de NaCl a Ψs entre -0,1 e -0,2 MPa (Figura 4C, 4E).
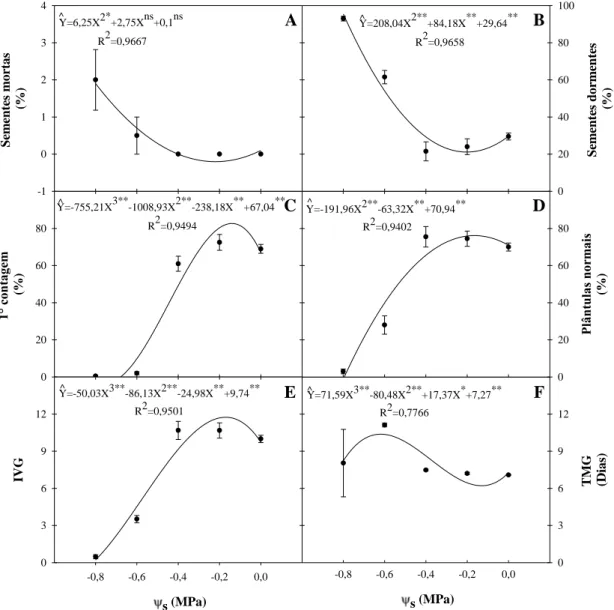
Figura 4 – Percentual de sementes mortas (A), de sementes não germinadas (B), de germinação na primeira contagem (C) e de plântulas normais (D), índice de velocidade de germinação (IVG, E) e tempo médio de germinação (TMG, F) de sementes de mamona cv. BRS-Energia em soluções salinas de NaCl com potenciais osmóticos (s) decrescentes.
Gordin et al., (2012) concluiu que a exposição ao NaCl reduz o poder germinativo das sementes de niger desde uma mínima redução no Ψs de -0,1 MPa. No caso das
s (MPa)
-0,8 -0,6 -0,4 -0,2 0,0
T M G (D ias) 0 3 6 9 12 B Se m ente s dor m ente s (% ) 0 20 40 60 80 100 A Se m ente s m or tas (% ) -1 0 1 2 3 4 C 1° c on tag em (% ) 0 20 40 60 80 D Pl ân tu las n or m ai s (% ) 0 20 40 60 80 s (MPa)
-0,8 -0,6 -0,4 -0,2 0,0
IV G 0 3 6 9 12 E F ^
Y=6,25X2*+2,75Xns+0,1ns R2=0,9667
^
Y=208,04X2**+84,18X**+29,64** R2=0,9658
^
Y=-191,96X2**-63,32X**+70,94** R2=0,9402
^
Y=-755,21X3**-1008,93X2**-238,18X**+67,04** R2=0,9494
^
Y=-50,03X3**-86,13X2**-24,98X**+9,74** R2=0,9501
^
sementes de mamona cv. BRS-Energia o poder germinativo só passou a ser afetados a partir de -0,4 MPa.
Deve-se destacar que o tratamento de salinidade de -0,8 MPa foi excluído dos experimentos conseguintes em vista da baixa germinação (±10%) e número plântulas normais (±1%) nessas condições (Figuras 3, 4D).
4.3.Efeitos da salinidade sobre crescimento de plântula
A salinidade alterou o crescimento e o acúmulo de matéria seca das partes estudadas das plântulas de mamona (Tabela 4). A salinidade trouxe contribuição positiva até o Ψs de -0,2 MPa, mas abaixo desse valor os níveis de salinidade passaram a reduzir o acúmulo de matéria seca ou crescimento das plântulas de mamona (Figura 5). Essa tendência de redução do crescimento com o aumento da CE também foi observada em plântulas de pinhão-manso submetidas à solução de NaCl ( ANDRÉO-SOUZA et al., 2010).
Tabela 4 – Resumo do quadro de ANOVA para as variáveis massas secas da raiz (MSR) e do hipocótilo (MSH), relação MSR/MSH, comprimentos da raiz (CR) e do hipocótilo (CH), e relação CR/CH de plântulas de mamona submetidas à soluções salinas com potenciais osmótico entre 0 e -0,6 MPa
Fonte de
variação G.L.
Quadrado Médio
MSR MSH MSR/MSH CR CH CR/CH
Tratamento 3 65,30** 392,99** 1,11** 625,21** 949,29** 417,02**
Resíduo 11 1,78 3,15 0,11 8,31 5,09 2,26
CV (%) 21,63 12,16 51,26 42,80 43,59 72,4
* p < 0,05; ** p < 0,01; ns p > 0,05
O crescimento da raiz teve como ponto crítico (Ψs onde os efeitos da salinidade passam a ser deletérios, levando em conta a condição controle, Ψs = 0 MPa) o Ψs de -0,31 MPa tanto para o acúmulo de massa seca quanto para o crescimento longitudinal da raiz principal (CR) (Figura 5A, 5D). Enquanto isso, o hipocótilo teve como ponto crítico para o acúmulo de massa seca (MSH) e para o crescimento longitudinal (CH) os
Ψs de -0,23 e -0,16 MPa, respectivamente (Figura 5B, 5E). O órgão da plântula de mamona cv. BRS-Energia mais sensível às reduções de Ψs com NaCl foi o hipocótilo.
Entre o Ψs -0,4 e -0,6 MPa a redução no CH foi 1,2 vezes maior que a redução no CR e a redução da MSH foi 1,4 vezes maior que a da MSR (Figura 5C, 5F).
Figura 5 – Massa seca da raiz (MSR, A) e do hipocótilo (MSH, B), relação massa seca da raiz/hipocótilo (MSR/MSH, C), comprimento da raiz principal (CR, D) e do hipocótilo (CH, E) e relação comprimento
M SH (m g) 0 6 12 18 24 30 M SR (m g) 0 3 6 9 12 15 M SR / M SH 0,0 0,6 1,2 1,8 2,4
CR (cm
) 0 3 6 9 12 s (MPa)
-0,6 -0,4 -0,2 0,0 CH (cm
) 0 2 4 6 8 s (MPa)
-0,6 -0,4 -0,2 0,0
C R / CH 0 2 4 6 8 B A C D E F ^
Y=-246,29X3**-265,43X2**-58,96X**+7,58** R2=1
^
Y=-402,97X3**-432,13X2**-79,21X*+21,44** R2=1
^
Y=6,46X2*+2,02Xns+0,42** R2=0,9224
^
Y=-187,8X3**-191,66X2**-41,82X**+7,05** R2=1
^
Y=-85,75X3**-91,23X2**-12,63X**+7,32** R2=1
^
da raiz/hipocótilo (CR/CH, F) de plântulas de mamona cv. BRS-Energia com 7 dias, oriundas de sementes germinadas em soluções salinas de NaCl com potenciais osmóticos (s) decrescentes.
A relação MSR/MSH em condições controle (Ψs = 0 MPa) foi igual a 0,42 (Figura 5C). Com a redução do Ψs até -0,16 MPa houve uma redução nessa relação de até 37,5% resultante de aumentos na MSH em maior proporção que os aumentos na MSR. Entre os Ψs de -0,16 e -0,23 MPa, o maior acúmulo relativo de massa seca passou a ser na raiz e após o Ψs de -0,23 MPa a MSH passou a ser inferior à condição controle assim como a MSR a partir de -0,31 MPa. Portanto, admite-se como aceitável apenas as variações de 0,26 a 0,42 na relação MSR / MSH entre os Ψs de 0 e -0,23 MPa por não haver numa redução no acúmulo de matéria seca das plântulas (Figura 5C). A variação de 0,99 a 1,58 na relação CR / CH é admissível entre o intervalo de 0 e -0,16 MPa, pois tanto o CR quanto o CH foram iguais ou superiores aos da condição controle (Figura 5E).
Resultados similares foram obtidos por Azevedo Neto e Tabosa (2000) e Carvalho
et al. (2012), que verificaram maior sensibilidade à salinidade no crescimento da parte aérea das plântulas de milho e soja, respectivamente, enquanto as raízes mantiveram seu desenvolvimento. A preservação das raízes pode ser vantajosa, já que as plantas com maior expansão do sistema radicular possuem tendência de serem mais resistentes aos efeitos do estresse hídrico, ao proporcionar condições para o desenvolvimento da planta e posterior recuperação da parte aérea, uma vez que a regulação da expansão foliar durante o período de exposição ao estresse osmótico ocorre em função da quantidade de água presente nas raízes (TOORCHI et al., 2009).
4.4.Efeitos da salinidade sobre a homeostase K+/Na+