UNIVERSIDADE FEDERAL DO CEARÁ DEPARTAMENTO DE BIOLOGIA
PÓS-GRADUAÇÃO EM ECOLOGIA E RECURSOS NATURAIS
WANESSA NEPOMUCENO FERREIRA
ECOFISIOLOGIA DA GERMINAÇÃO E DO CRESCIMENTO INICIAL DE Piptadenia stipulacea (Benth.) Ducke E Anadenanthera colubrina (Vell.) Brenan
WANESSA NEPOMUCENO FERREIRA
ECOFISIOLOGIA DA GERMINAÇÃO E DO CRESCIMENTO INICIAL DE Piptadenia stipulacea (Benth.) DuckeE Anadenanthera colubrina (Vell.) Brenan
Dissertação submetida à Coordenação do Programa de Pós-Graduação em Ecologia e Recursos Naturais da Universidade Federal do Ceará, como requisito parcial para obtenção do grau de Mestre em Ecologia e Recursos Naturais.
Orientador: Prof. Dr. Sebastião Medeiros Filho/UFC Co-orientadora: Profª. Dra. Roberta Boscaini Zandavalli/UFC
F444e Ferreira, Wanessa Nepomuceno
Ecofisiologia da germinação e do crescimento inicial de Piptadenia stipulacea (Benth.) Ducke e Anadenanthera colubrina (Vell.) Brenan / Wanessa Nepomuceno Ferreira. – 2011.
65 f. : il. color., enc.
Orientador: Prof. Dr. Sebastião Medeiros Filho
Co-orientadora: Profa. Dra. Roberta Boscaini Zandavalli Área de concentração: Ecologia e Recursos Naturais
Dissertação (Mestrado) – Universidade Federal do Ceará, Centro de Ciências, Departamento de Biologia, Fortaleza, 2011.
1. Ecofisiologia vegetal 2. Germinação I. Medeiros Filho, Sebastião (Orient.) II. Zandavalli, Roberta Boscaini (Co-orient.) III. Universidade Federal do Ceará – Programa de Pós-Graduação em Ecologia e Recursos Naturais III.Título
CDD 577
WANESSA NEPOMUCENO FERREIRA
ECOFISIOLOGIA DA GERMINAÇÃO E DO CRESCIMENTO INICIAL DE Piptadenia stipulacea (Benth.) DuckeE Anadenanthera colubrina (Vell.) Brenan
Dissertação submetida à Coordenação do Programa de Pós-Graduação em Ecologia e Recursos Naturais da Universidade Federal do Ceará, como requisito parcial para obtenção do grau de Mestre em Ecologia e Recursos Naturais.
AGRADECIMENTOS
Com um sentimento de gratidão imensurável deixo aqui os meus sinceros agradecimentos:
A Deus, por renovar minha fé a cada dia e fazer-me perseverar mesmo quando as “pedras no caminho” pareciam-me intransponíveis;
Ao meu orientador Dr. Sebastião Medeiros Filho e Co-orientadora Dra. Roberta Boscaini Zandavalli pela orientação, paciência e força que transmitiram em todos os momentos; obrigada de coração;
Ao CNPq e a CAPES pela concessão das bolsas, respectivamente, no primeiro e segundo ano do curso;
Aos componentes da banca examinadora, Dr. Rogério Parentoni e Dra. Mª Clarete, pela disponibilidade de participar e contribuir com sugestões e comentários a cerca dessa dissertação;
Ao prof. Marcos Esmeraldo por disponibilizar gentilmente o espaço físico do Núcleo de Ensino e Pesquisa em Agricultura Urbana – NEPAU para realização do experimento de irradiância e pelos valiosos ensinamentos, confiança, incentivo e amizade;
Ao amigo Carlos Barboza pelas inestimáveis colaborações nas mais diversas fases deste trabalho, sempre com amizade e profissionalismo, sem medir esforços para simplesmente ajudar;
Ao Sr. Manoel e família, moradores da Fazenda Não Me Deixes, pela atenção e acompanhamento em campo;
A todos os integrantes do NEPAU, em especial ao Magnum, Cleriston, Cristiano, Mazinho, George e Edilson, que me receberam e ajudaram de coração;
A todos do Laboratório de Análises de Sementes: Salete, Batista, Elizita, Cássia, Franklin, Luiz Carlos, Carla, Wener, Paulinha, Estela, Alexandra, Lineker, Ronaldo, Adriana, Felipe, Denise, Neurilan e, especialmente, a Diego, Selma, Wendney e a professora Clarete pela atenção, preocupação e ajuda em momentos difíceis; todos vocês vão ficar pra sempre;
À toda minha família pelo amor, força e incentivo sempre; “Mainha”, a senhora é meu porto seguro;
À minha grande amiga Camila, que morou comigo durante os dois anos, pelo apoio, atenção, respeito e carinho; você é uma verdadeira irmã;
A todos os professores e alunos do Programa de Pós-Graduação em Ecologia e Recursos Naturais/UFC. Em destaque, aos alunos Ubirajara, Belchior e Paulo pela amizade e por estarem ao meu lado em momentos de descontração e alegria, mas, sobretudo, nos momentos de aflição e preocupação. Em especial, a Ingrid, Giovana e Juliana, não só pelas oportunas discussões ecológicas, mas também pelas conversas sobre a vida e amizades tão gratificantes; torço pelo sucesso de todos vocês;
“Talvez não tenhamos conseguido fazer o melhor, mas lutamos para que o melhor fosse feito. Não somos o que deveríamos ser.
Não somos o que iremos ser... Mas graças a Deus, não somos o que éramos!”
RESUMO
Este estudo apresenta a hipótese de que P. stipulacea é uma espécie característica de estágio inicial de sucessão, enquanto A. colubrina de estágios mais avançados. Para caracterizar as duas espécies, estudaram-se alguns atributos utilizados na classificação de grupos ecológicos de florestas tropicais úmidas, respondendo as seguintes questões: a) as espécies apresentam dormência de sementes? b) quais as condições de temperatura e luz para germinação? c) as sementes apresentam tolerância à dessecação? d) quais as características de crescimento das plântulas sob diferentes graus de irradiância? Dessa forma, o trabalho apresentou as seguintes previsões: a) as sementes de P. stipulacea apresentam dormência, enquanto as de A. colubrina não apresentam; b) as sementes de P. stipulacea germinam em uma ampla faixa de temperatura, enquanto as de A. colubrina germinam em uma faixa de temperatura mais estreita; c) as sementes de P. stipulacea toleram baixos níveis de dessecação, enquanto as de A. colubrina não toleram; d) as plântulas de P. stipulacea crescem melhor em maiores níveis de irradiância, enquanto as de A. colubrina crescem melhor em menores níveis de irradiância. Os frutos foram coletados em área de caatinga situada na fazenda “Não Me Deixes”, no município de Quixadá-CE, e os experimentos foram realizados no Laboratório de Análise de Sementes e no Núcleo de Ensino e Pesquisa em Agricultura Urbana. Resultados do teste preliminar de germinação mostraram que sementes de P. stipulacea apresentam dormência, sendo melhor superada com escarificação química por 10 minutos. Já as sementes de A. colubrina não possuem dormência. O experimento de temperatura e luz revelou que as duas espécies apresentam a mesma amplitude de germinação (10ºC a 40ºC), com melhor germinação a 30ºC, porém, nas menores temperaturas P. stipulacea apresentou germinação muito baixa. Quanto à tolerância a dessecação, as sementes de P. stipulacea suportaram secagem até 5,16% de umidade sem influenciar a germinação, representando comportamento ortodoxo. Já com A. colubrina a germinação diminuiu à medida que as sementes perderam água, porém, mesmo com teor de água de 5,81%, a germinação permaneceu acima de 80%, indicando que esta espécie também apresenta sementes ortodoxas. No experimento de irradiância, plântulas de P. stipulacea apresentaram maior diâmetro a pleno sol e, em todos os tratamentos, acumularam maior biomassa para o caule. A. colubrina apresentou maior altura e diâmetro em 50% de sombra e maior número de folhas e biomassa da parte aérea em 70% de sombra, indicando que esta espécie tolera níveis intermediários de sombra. Sugere-se que características utilizadas na classificação de grupos ecológicos de florestas tropicais úmidas também possam ser utilizadas para subsidiar a identificação de qual estágio sucessional pertencem determinadas espécies da caatinga, com o objetivo de entender sua ocorrência, distribuição e desenvolvimento, para melhor conservação e uso adequado no reflorestamento de áreas degradadas.
ABSTRACT
This study presents the hypothesis that P. stipulacea is a species characteristic of early stages of succession, while A. colubrina in later stages. To characterize the two species studied are some attributes used in classification of ecological groups of tropical rainforests, answering the following questions: a) species have seed dormancy? b) what conditions of temperature and light for germination? c) the seeds are desiccation tolerance? d) the characteristics of seedling growth under different levels of irradiance? Thus, the work made the following predictions: a) seeds of P. stipulacea present dormancy, while those of A. colubrina not present; b) seeds of P. stipulacea germinate over a wide temperature range, while those of A. colubrina germinate in a narrow temperature range; c) the seeds of P. stipulacea tolerate low levels of desiccation, while those of A. colubrina not tolerate; d) seedlings of P. stipulacea grow best at higher irradiance levels, while those of A. colubrina grow best at lower levels of irradiance. Fruits were collected in the caatinga area located on the farm "Não Me Deixes” city of Quixada-CE, and the experiments were performed at the Seed Analysis Laboratory and the Core Teaching and Research in Urban Agriculture. Results of preliminary testing of germination showed that seeds of P. stipulacea present dormancy and are better solved with chemical scarification for 10 minutes. The seeds of A. colubrina have no dormancy. The experiment temperature and light was revealed that the two species have the same range of germination (10°C to 40ºC), with best germination at 30°C, but at lower temperatures P. stipulacea showed germination very low. As for tolerance to desiccation, seeds of P. stipulacea bore drying up 5.16% moisture without influencing germination, representing orthodox behavior. Already A. colubrina germination decreased as the seeds lost water, but even with a water content of 5.81%, germination remained above 80%, indicating that this species also has orthodox seeds. In the irradiance experiment, seedlings of P. stipulacea showed larger diameter in full sun, and all treatments accumulated more biomass to the stem. A. colubrina showed greater height and diameter in 50% shade and a larger number of leaves and shoot biomass by 70% shade, indicating that this species tolerates intermediate levels of shade. It is suggested that features used for classification of ecological groups of tropical forests could also be used to support the identification of which stage of succession belongs to certain species of caatinga, with the aim of understanding their occurrence, distribution and development, for better conservation and use suitable for the reforestation of degraded areas.
LISTA DE FIGURAS
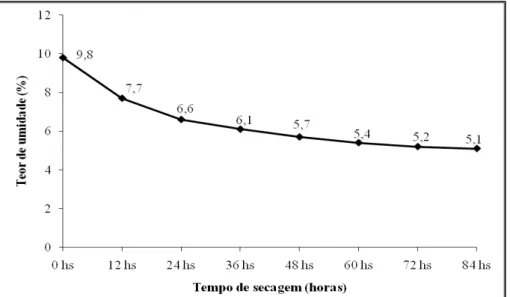
Figura 1 Curva de secagem representando o comportamento do teor de água das sementes de Piptadenia stipulacea em função do tempo de secagem...
38 Figura 2 Curva de secagem representando o comportamento do teor de água das
sementes Anadenanthera colubrina em função do tempo de secagem...
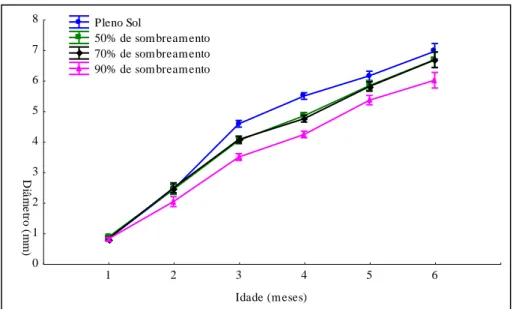
40 Figura 3 Altura das mudas de Piptadenia stipulacea até seis meses de idade,
para diferentes níveis de sombreamento...
42 Figura 4 Diâmetro do coleto das mudas de Piptadenia stipulacea até seis meses
de idade, para diferentes níveis de sombreamento...
43 Figura 5 Número de folhas das mudas de Piptadenia stipulacea até seis meses
de idade, para diferentes níveis de sombreamento...
44 Figura 6 Altura das mudas de Anadenanthera colubrina até seis meses de idade,
para diferentes níveis de sombreamento...
46 Figura 7 Diâmetro do coleto das mudas de Anadenanthera colubrina até seis
meses de idade, para diferentes níveis de sombreamento...
47 Figura 8 Número de folhas das mudas de Anadenanthera colubrina até seis
meses de idade, para diferentes níveis de sombreamento...
LISTA DE TABELAS
Tabela 1 Porcentagem de emergência (PE), índice de velocidade de emergência (IVE) e tempo médio de emergência (TME) de sementes de Piptadenia stipulacea submetidas a diferentes tratamentos de superação de dormência. Fortaleza, CE, 2011...
33 Tabela 2 Resumo da Análise de variância dos dados de porcentagem de
germinação (PG), índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG) de sementes de Piptadenia stipulacea, submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011...
35 Tabela 3 Médias da porcentagem de germinação (PG), índice de velocidade de
germinação (IVG) e tempo médio de germinação (TMG) de sementes de Piptadenia stipulacea, submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011...
35 Tabela 4 Resumo da Análise de variância dos dados de porcentagem de
germinação (PG), índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG) de sementes de Anadenanthera colubrina, submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011...
36 Tabela 5 Médias da porcentagem de germinação (PG), índice de velocidade de
germinação (IVG) e tempo médio de germinação (TMG) de sementes de Anadenanthera colubrina, submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011...
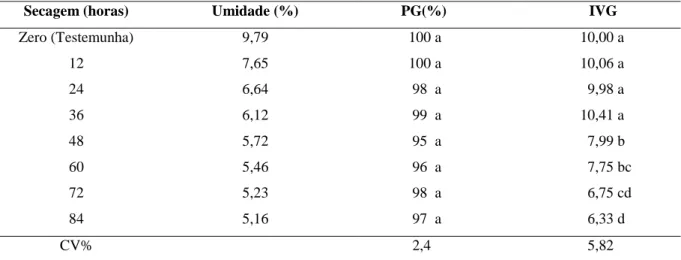
37 Tabela 6 Valores médios de umidade, porcentagem de germinação (PG) e índice
de velocidade de germinação (IVG) de sementes de Piptadenia stipulacea, submetidas a diferentes tratamentos de dessecação. Fortaleza, CE, 2011...
39 Tabela 7 Valores médios de umidade, porcentagem de germinação (PG) e índice
de velocidade de germinação (IVG) de sementes de Anadenanthera colubrina, submetidas a diferentes tratamentos de dessecação. Fortaleza, CE, 2011...
40
Tabela 8 Análise de variância para as variáveis altura, diâmetro e número de folhas das plantas de Piptadenia stipulacea submetidas a diferentes tratamentos de sombreamento. Fortaleza, CE, 2011...
41 Tabela 9 Efeito dos diferentes níveis de sombreamento sobre as variáveis de
peso da matéria seca (g) das plantas jovens de Piptadenia stipulacea. Fortaleza, CE, 2011...
45 Tabela 10 Análise de variância para as variáveis altura, diâmetro e número de
folhas das plantas de Anadenanthera colubrina submetidas a diferentes tratamentos de sombreamento. Fortaleza, CE, 2011...
46 Tabela 11 Efeito dos diferentes níveis de sombreamento sobre as variáveis de
peso (g) da matéria seca das plantas jovens de Anadenanthera colubrina. Fortaleza, CE, 2011...
SUMÁRIO
1 INTRODUÇÃO... 11
2 REVISÃO DE LITERATURA... 15
2.1 Germinação e dormência... 15
2.2 Influência da temperatura e da luz na germinação de sementes... 17
2.3 Tolerância à dessecação em sementes... 19
2.4 Crescimento inicial de plântulas em diferentes níveis de irradiância.... 21
3 MATERIAL E MÉTODOS... 23
3.1 Preparo de exsicatas e coleta dos frutos... 23
3.2 Espécies focais... 24
3.2.1 Piptadenia stipulacea (Benth.) Ducke... 24
3.2.2 Anadenanthera colubrina (Vell.) Brenan var. cebil (Griseb.) Altshul... 24
3.3 Teste preliminar de germinação e superação da dormência... 25
3.4 Influência da temperatura e da luz na germinação de sementes... 27
3.5 Tolerância à dessecação em sementes... 28
3.6 Crescimento inicial de plântulas em diferentes níveis de irradiância.... 30
4 RESULTADOS E DISCUSSÃO... 32
4.1 Teste preliminar de germinação e superação da dormência... 32
4.2 Influência da temperatura e da luz na germinação de sementes... 34
4.3 Tolerância à dessecação em sementes... 38
4.4 Crescimento inicial de plântulas em diferentes níveis de irradiância.... 41
5 CONSIDERAÇÕES FINAIS... 50
REFERÊNCIAS... 52
1 INTRODUÇÃO
A caatinga do nordeste brasileiro, com 735.000km2, é um mosaico de arbustos espinhosos e florestas sazonalmente secas que cobre a maior parte dos estados do Piauí, Ceará, Rio Grande do Norte, Paraíba, Pernambuco, Alagoas, Sergipe, Bahia e a parte nordeste de Minas Gerais, no vale do Jequitinhonha (LEAL et al., 2005). Esta vegetação está submetida a temperaturas médias anuais constantes e altas, 26°C em média, e precipitação pluviométrica anual baixa (até 750mm em 50% do território) e irregular, concentrada em três a quatro meses consecutivos (NIMER, 1989).
Historicamente, a caatinga sofreu impactos do processo de uso e ocupação do solo, que a degradaram paulatinamente. Ainda hoje, as atividades antrópicas da maneira como são desenvolvidas, não são compatíveis com as condições do ambiente ou com o regime pluviométrico da região (SOUZA, 2006). Para acentuar ainda mais este problema, registra-se o número bastante reduzido de unidades de conservação e a insuficiência do conhecimento científico (TABARELLI; VICENTE, 2002).
Para subsidiar ações de conservação na caatinga é necessário estruturar o conhecimento já existente e buscar novas informações que permitam a compreensão dos padrões e processos ecológicos que operam na região. Dentre os estudos conduzidos na caatinga, cabe salientar os do componente vegetal, como: estratégias de germinação (BARBOSA, 2003), fenologia e síndromes de dispersão (GRIZ; MACHADO, 2001, BARBOSA; BARBOSA; LIMA, 2003), influência de diferentes níveis de luz no crescimento inicial de espécies vegetais (RAMOS et al., 2004, RODRIGUES et al., 2007, MELO; CUNHA, 2008) e composição florística e fitossociologia (LEMOS; RODAL, 2002, ALCOFORADO-FILHO; SAMPAIO; RODAL, 2003). Entretanto, são necessários mais estudos sobre características de história de vida das espécies, que permitam entender sua distribuição e importância em processos ecológicos.
Os estudos sobre história de vida buscam explicar como organismos evoluíram, alcançando o sucesso reprodutivo. História da vida é geralmente definida como um conjunto de estratégias evoluídas, inclusive adaptações comportamentais, anatômicas e fisiológicas, que podem influenciar diretamente na sobrevivência e sucesso reprodutivo dos organismos (RICKLEFS; WIKELSKI, 2002).
foram categorizadas em grupos ecológicos distintos. Segundo Paula et al. (2004), estas distinções são ferramentas essenciais para a compreensão da sucessão ecológica. Existem diversas classificações de grupos ecológicos (BUDOWSKI, 1965; BAZZAZ, 1979; SWAINE; WHITMORE, 1988; JARDIM; SOUZA; SILVA, 1996) que se diferenciam quanto às denominações e as características utilizadas. No entanto, todas se baseiam na divisão das espécies florestais entre aquelas de estádios iniciais e tardios da sucessão. Budowski (1965), com base em vinte e uma características das espécies que compõe o ecossistema de florestas tropicais úmidas propôs classificá-las em pioneiras, secundárias iniciais, secundárias tardias e climáceas. Enquanto, Swaine e Whitmore (1988) definiram, com base na germinação de sementes e estabelecimento de plântulas, dois grupos principais: das espécies pioneiras e não pioneiras (clímax), sendo que este último grupo é subdividido em espécies clímax exigentes de luz e clímax tolerantes à sombra. E mais recentemente, Whitmore (1990) mostrou a classificação pioneira/clímax com base em 16 características das espécies tropicais.
Rodrigues (1995) comenta que as espécies pioneiras têm a função de regenerar ambientes perturbados. Estas espécies são boas colonizadoras, devido a sua capacidade de dispersão e alta fecundidade que permitem seu estabelecimento antes da chegada das espécies clímax. No outro extremo das pioneiras, têm-se as climáceas, que são as espécies finais na substituição seqüencial da sucessão. Entre os dois extremos existe um grande número de espécies com características ou adaptações ecológicas intermediárias. Quando as características são mais parecidas com as pioneiras, estas espécies são chamadas de secundárias iniciais; quando apresentam características mais próximas das espécies clímax, são denominadas secundárias tardias.
Observa-se em trabalhos fitossociológicos realizados em áreas de caatinga que, assim como espécies pioneiras e tardias de florestas tropicais úmidas, algumas espécies da caatinga tais como, Croton sonderianus, Caesalpinia pyramidalis e Aspidosperma pyrifolium
(PEREIRA et al., 2001; ANDRADE et al., 2005) são mais abundantes em áreas perturbadas, enquanto outras espécies como Myracrodruon urundeuva e Amburana cearensis já foram relatadas como exclusivas em áreas conservadas (BARBOSA et al., 2007; PEREIRA et al., 2001). Diante disso, acredita-se que algumas características utilizadas nas classificações de grupos ecológicos de florestas tropicais úmidas, também possam oferecer subsídios para compreender porque algumas espécies da caatinga se comportam de modo diferente em áreas perturbadas e conservadas.
trabalhos fitossociológicos revelou que a primeira apresenta maiores valores de abundância em áreas que foram submetidas a grandes níveis de perturbação, em detrimento da segunda. Um estudo realizado na Serra do Monte, Boqueirão, PB, em uma área com focos de antropização, mostrou que P. stipulacea apresentou densidade de 52.5 ind.ha-1 e freqüência absoluta de 40%, valores superiores aos de A. colubrina com 32,5 ind.ha-1 e 15%, respectivamente. Os autores consideraram A. colubrina como uma espécie tardia e inferiram que essa vegetação poderia estar evoluindo em direção a uma condição mais avançada de desenvolvimento, passando por um processo de regeneração e sucessão (QUEIROZ et al., 2006). Outro estudo de regeneração natural em um remanescente de caatinga, sob diferentes níveis de perturbação, constatou uma alta densidade e freqüência de P. stipulacea,
principalmente na área mais antropizada, sendo considerada pelos autores como uma espécie tolerante e indicadora de elevados níveis de perturbação (PEREIRA et al., 2001). Em uma análise da cobertura de duas fitofisionomias de caatinga, com diferentes históricos de uso, Andrade et al. (2005) verificaram que A. colubrina foi encontradaapenas na área considerada relativamente conservada, enquanto que na área em regeneração há 30 anos não houve registros da espécie.
Sabe-se que a estrutura das populações vegetais depende de muitos fatores bióticos e abióticos. Entretanto, Pereira et al. (2001) afirmam que, considerando a extensão e a homogeneidade de algumas áreas de caatinga, particularmente no que se refere às condições de relevo, solos e clima, não é de se esperar que existam fatores restritivos à ocorrência das espécies em determinados locais. Portanto, a distribuição destas em áreas em diferentes estágios de sucessão, pode ser entendida como uma resposta das mesmas à perturbação antrópica. Considerando-se que estas respostas estão relacionadas a características ecofisiológicas, este estudo tem como hipótese que Piptadenia stipulacea (Benth.) Ducke caracteriza-se como uma espécie de estágio inicial de sucessão, enquanto Anadenanthera colubrina (Vell.) Brenan var. cebil (Griseb.) Altshul de estágios mais avançados, visto que seus parâmetros fitossociológicos possuem diferentes valores em áreas de caatinga perturbada e conservada.
a) as espécies apresentam dormência de sementes?
b) quais as condições de temperatura e luminosidade para a germinação de suas sementes? c) as sementes das espécies apresentam tolerância à dessecação?
d) quais as características de crescimento das plantas jovens, em viveiros, sob diferentes graus de irradiância?
Assim, este estudo seguiu o seguinte diagrama:
Dormência de sementes
Quase sempre presente
Quase sempre ausente
P. stipulacea A. colubrina Apresenta dormência Não apresenta dormência Temperatura de germinação Maior amplitude (característica de clareiras) Menor amplitude (característica de
sub-bosque)
Germinam em uma ampla faixa de
temperatura Germinam em uma faixa de temperatura
mais estreita
Tolerância a dessecação de
sementes
Ortodoxa (toleram dessecação a baixos
níveis de umidade) Quase sempre recalcitrante (não toleram a dessecação a
níveis de umidade muito baixos)
Toleram baixos níveis de dessecação
Não toleram baixos níveis de dessecação
estreita
Tolerância das plantas à
sombra
Muito intolerante
Tolerante (exceto no estágio adulto)
Crescem melhor em maiores níveis de
irradiância
Crescem melhor em menores níveis de
2 REVISÃO DE LITERATURA
2.1 Germinação e dormência
O estudo das exigências necessárias para que a germinação ocorra é fundamental para a compreensão de processos como sucessão e regeneração natural. A germinação pode ser definida como a retomada do crescimento do embrião de uma semente em condições favoráveis de hidratação, temperatura e oxigênio. Este processo é iniciado pela absorção de água pela semente e terminando com o alongamento do eixo embrionário, geralmente a radícula (LABOURIAU, 1983). Diversos fatores bióticos e abióticos, como temperatura, luz, água, salinidade, dispersão e predação atuam em conjunto, regulando o processo de germinação. Além destes fatores, características da própria semente, como tamanho, viabilidade, conteúdo de água, tolerância à dessecação, dormência, espessura da testa, entre outros, também influenciam o processo de germinação (BASKIN; BASKIN, 1998; FENNER; THOMPSON, 2005).
A ausência de germinação de uma semente viável em condições usualmente consideradas como favoráveis à germinação, ou seja, em condições ideais de hidratação, temperatura e oxigênio, é definida como dormência. A dormência é causada por um ou mais bloqueios situados na própria semente que precisam ser superados para que a germinação ocorra (BASKIN; BASKIN, 1998). Pode-se dizer que a principal função da dormência é impedir a germinação em condições desfavoráveis ao estabelecimento das plântulas, sendo uma característica importante na manutenção das espécies (FENNER; THOMPSON, 2005). A dormência distribui a germinação no tempo, fazendo com que a emergência de plântulas ocorra em intervalos irregulares, diminuindo a competição e aumentando a probabilidade de sobrevivência das plântulas (BEWLEY; BLACK, 1994).
Fowler e Bianchetti (2000) afirmaram que os dois tipos de dormência são: endógena ou embrionária e exógena ou tegumentar.
hormonais no embrião, ou seja, tanto a redução da concentração dos inibidores como a síntese de fitohormônios promotores da germinação. Assim, métodos que atuem impedindo a ação dos primeiros ou que aumentem a concentração dos promotores são os mais recomendados (DIAS, 2005).
A dormência exógena é causada pela impermeabilidade do tegumento à água e gases. Essa impermeabilidade é bastante comum e está associada a espécies de diversas famílias botânicas, sendo constatada com maior freqüência em sementes de espécies de Leguminosae (CARVALHO; NAKAGAWA, 2000). A maioria das espécies de ocorrência no semiárido nordestino também apresenta a dormência tegumentar, considerada um mecanismo de ajuste ecofisiológico inibidor da embebição rápida durante a ocorrência de chuvas irregulares na estação seca (ARAÚJO; FERRAZ, 2003; ARAÚJO et al., 2006; LIMA et al.
2007). Cerca de 67 a 70% das espécies das florestas tropicais decíduas, dos cerrados e das caatingas apresentam dormência física, sendo tais percentuais elevados quando comparados às florestas tropicais úmidas e semi-deciduais (18-40%) (CARDOSO, 2004; LEAL et al., 2008). Segundo Vázquez-Yanes e Orozco-Segovia (1993), nas florestas topicais úmidas a dormência é uma característica comum entre espécies tipicamente pioneiras.
A dormência exógena pode ser superada por processos de escarificação, a qual consiste em qualquer tratamento que resulte na ruptura ou enfraquecimento do tegumento, permitindo a entrada de água e gases e, assim, dando inicio ao processo germinativo (MAYER; POLJAKOFF-MAYBER, 1989). Em ambiente natural, a escarificação pode ocorrer pelo aquecimento úmido ou seco do solo por temperaturas alternadas, permitindo, assim, a entrada de água para o interior da semente. Esse processo pode ocorrer também, pela ação de ácidos durante a ingestão das sementes por animais dispersores, além da ação dos microorganismos do solo (VAZQUEZ-YANES; OROZCO-SEGOVIA, 1993).
2.2 Influência da temperatura e da luz na germinação de sementes
A temperatura é considerada de grande importância na germinação, pois exerce forte influência na sua velocidade e uniformidade, e está relacionada com os processos bioquímicos (CARVALHO; NAKAGAWA, 2000). A temperatura pode regular a germinação por três maneiras: determinando a capacidade e taxa de germinação; removendo a dormência primária ou secundária; e induzindo dormência secundária (BEWLEY; BLACK 1994). Brändel e Schütz (2005) também afirmaram que a temperatura influencia a absorção de água pela semente e as reações bioquímicas que regulam o metabolismo envolvido neste processo, principalmente a entrada de oxigênio até o embrião, afetando a porcentagem, velocidade de germinação, viabilidade, e quebra ou indução de dormência.
Cada espécie possui uma faixa de temperatura na qual a germinação ocorre, ou seja, acima da temperatura máxima e abaixo da temperatura mínima não há germinação. Desta forma, existem três temperaturas cardinais para a germinação. A temperatura máxima, mínima e ótima. A faixa ótima de temperatura é aquela na qual há uma maior porcentagem de germinação em um menor intervalo de tempo (BEWLEY; BLACK, 1994).
Temperaturas elevadas e flutuações na temperatura são fatores importantes na germinação de sementes com tegumento duro, uma vez que provocam rachaduras no tegumento, permitindo a entrada de água e gases (BASKIN, 2003). Porém, temperaturas elevadas podem ser letais, já que elas desnaturam estruturas celulares e causam um rápido envelhecimento da semente, enquanto que baixas temperaturas reduzem o metabolismo, gerando um atraso na germinação e uma redução da germinação (PROBERT, 2000; ZAIDAN; BARBEDO, 2004). Além disso, altas temperaturas aumentam a evaporação da água no solo, diminuindo a disponibilidade de água para o processo germinativo (BRESHEARS et al., 1998).
De forma geral, espécies tropicais germinam em temperaturas mais elevadas do que espécies de ambientes temperados (FENNER; THOMPSON, 2005). Para algumas espécies encontradas na caatinga, tais como, Cassia excelsa, Mimosa caesalpiniaefolia e
Amburana cearensis a temperatura ótima foi de 30ºC (JELLER; PEREZ, 1999; ALVES et al., 2002; GUEDES et al., 2010).
havendo também, as que são indiferentes, ou seja, que não apresentam sensibilidade à luz (OROZCO-SEGOVIA; VÁZQUEZ-YANES, 1989). A germinação das sementes, em relação à luz, é uma resposta ecofisiológica da espécie e tem correspondência com o seu posicionamento no estádio sucessional da floresta (JESUS; PINA-RODRIGUES, 1991). As sementes de espécies pioneiras fotoblásticas respondem com germinação plena apenas quando são submetidas à luz vermelha, enquanto as dos demais grupos ecológicos, como as secundárias e as clímax, têm a capacidade de germinar à sombra do dossel, sem luz solar direta (KAGEYAMA; VIANA, 1991). Em algumas espécies, o requerimento de luz para germinação das sementes é influenciado pela temperatura (SMITH, 1975).
Maia et al. (2008) verificaram que Hyptis suaveolens, espécie distribuída em regiões tropicais e subtropicais, e comum em locais que foram submetidos à ação antrópica, apresenta maior germinação de sementes na presença de luz e em temperaturas altas (25 e 30ºC). Araújo Neto, Aguiar e Ferreira (2003) estudando a germinação de uma espécie arbórea, Acácia polyphylla, característica dos estádios iniciais da sucessão em Floresta Estacional Semidecidual (PAULA et al., 2004), verificaram que a porcentagem de germinação não foi afetada pelos diferentes fotoperíodos de luz branca testados. Entretanto, o fotoperíodo igual ou superior a quatro horas favoreceu o crescimento inicial das plântulas. A germinação não foi afetada pela luz com baixa relação vermelho/vermelho-extremo, mas na ausência de luz as sementes germinaram em menores porcentagens e velocidade.
Sementes de Miconia theazeans, espécie encontrada na Floresta Atlântica, apresentaram forte dependência da presença de luz branca para a indução da germinação com a necessidade de 4-6 horas de luz diária para a máxima indução do processo. Entretanto, com a alternância de temperaturas de 30 e 20°C, o fotoperíodo de 2 horas é suficiente para a indução da germinação sendo o máximo de indução obtida a partir de 4 horas diárias. Estes parâmetros indicam que Miconia theazeans é uma espécie importante que coloniza clareiras e áreas perturbadas em uma floresta natural (GODOI; TAKAKI, 2007).
2.3 Tolerância à dessecação em sementes
Assim como a temperatura e a luminosidade, outros fatores físicos como a disponibilidade hídrica do solo promovem o aparecimento de características germinativas diversificadas (VÁZQUEZ-YANES; OROZCO-SEGOVIA 1996).
A água é um fator crucial que regula o processo de germinação, pois este se inicia somente com a embebição. Uma baixa disponibilidade hídrica, principalmente no começo da embebição, inviabiliza a seqüência dos eventos germinativos da semente durante a absorção de água, conseqüentemente podendo diminuir a porcentagem e a velocidade de germinação ou até mesmo impedir que este processo ocorra (CASTRO; HILHORST, 2004). Além disso, uma baixa disponibilidade hídrica pode causar a dessecação de sementes, provocando a morte das mesmas. Entretanto, é preciso considerar que o grau de tolerância à dessecação e quantidade de água exigida para germinação variam de acordo com a espécie, influenciando a distribuição da mesma. É observado que sementes de espécies de ambientes áridos, geralmente, são tolerantes à dessecação e conseguem germinar em uma menor disponibilidade hídrica, se comparadas às sementes de espécies provenientes de ambientes úmidos, que geralmente são sensíveis à dessecação (EVANS; ETHERINGTON, 1990; FACELLI; LADD, 1996; PAMMENTER; BERJAK, 2000; TWEDDLE et al., 2003).
Segundo Roberts (1973), as sementes podem ser classificadas fisiologicamente em dois grupos distintos: no primeiro estão as ortodoxas, que se mantém viáveis após dessecação até um grau de umidade em torno de 5% e podem ser armazenadas sob baixas temperaturas por um longo período; no segundo grupo têm-se as recalcitrantes, ou sementes sensíveis à dessecação, que não sobrevivem com baixos níveis de umidade, o que impede o seu armazenamento por longo prazo.
Segundo Ellis, Hong e Roberts (1990), as sementes podem ser classificadas em um terceiro grupo, no qual apresentam um comportamento de armazenamento intermediário ao ortodoxo e ao recalcitrante. De acordo com Hong e Ellis (1996), as sementes que apresentam comportamento intermediário toleram a desidratação até 7,0% a 10% de umidade e não toleram baixas temperaturas durante período de tempo prolongado. Assim, a classificação das sementes quanto à capacidade de armazenamento depende de estudos de tolerância à dessecação e do armazenamento sob temperaturas baixas.
sementes adquirem tolerância à dessecação, que é mantida após a dispersão (BEWLEY; BLACK, 1994). A secagem até o grau de umidade de 5,0% a 15,0% pode levar as sementes a um estado de quiescência, o que causa redução do metabolismo até níveis mínimos, permitindo que elas permaneçam vivas, mesmo sob condições adversas. Após a hidratação, estas sementes podem retomar o metabolismo direcionado para o processo de germinação (BEWLEY; BLACK, 1994). Por outro lado, as sementes recalcitrantes não sofrem a secagem no final da maturação e são dispersas com elevado grau de umidade permanecendo metabolicamente ativas, sensíveis à secagem e podendo germinar logo após a dispersão (PAMMENTER; BERJAK, 2000).
Existe uma associação entre o comportamento das sementes durante o armazenamento e os grupos ecológicos a que as espécies pertencem. Swaine e Whitmore (1988) separaram as espécies em dois grupos ecológicos principais: espécies pioneiras e espécies clímax. As sementes de espécies pioneiras necessitam de alta intensidade de luz para a germinação, geralmente apresentam dormência, principalmente causada por tegumento impermeável e possuem alta longevidade. Estas espécies regeneram-se por meio do banco de sementes no solo e podem ser armazenadas durante longo período (KAGEYAMA; VIANA, 1991), o que corresponde ao comportamento ortodoxo (ROBERTS, 1973). As sementes de espécies clímax, que não necessitam de luz direta para germinação e posterior crescimento da plântula, geralmente não apresentam dormência, possuem reduzida longevidade e regeneram-se, principalmente, por meio do banco de plântulas. Dentro deste grupo podem ser encontradas as sementes classificadas como recalcitrantes (KAGEYAMA; VIANA, 1991; PAMMENTER; BERJAK, 2000).
Carvalho, Silva e Davide (2006), classificaram 22 sementes de espécies florestais quanto ao comportamento no armazenamento e encontraram 17 espécies ortodoxas e cinco espécies recalcitrantes. O comportamento ortodoxo distribuiu-se em cinco espécies pioneiras, nove espécies clímax exigentes de luz para o crescimento da plântula e três espécies clímax tolerantes à sombra. As cinco sementes classificadas como recalcitrantes distribuíram-se em duas espécies clímax tolerantes à sombra e três espécies clímax exigentes de luz.
2.4 Crescimento inicial de plântulas em diferentes níveis de irradiância.
Assim como as características de germinação, o estudo do estabelecimento das plântulas é importante, pois o estádio de plântula é uma fase decisiva para a sobrevivência de um indivíduo e para a distribuição espacial de uma população (LARCHER, 2004). Conforme o mesmo autor, durante essa fase a plântula exige um pleno suprimento de nutrientes necessários para garantir o aumento de energia e metabólitos utilizados na biossíntese, bem como um estado de hidratação suficiente para manter a turgescência durante o rápido crescimento em extensão e a diferenciação da parede celular.
Um dos fatores ambientais essenciais para o estabelecimento de algumas espécies vegetais é a disponibilidade de luz. Esta desempenha papel fundamental na organização das comunidades, sendo apontada como um dos fatores mais importantes para as estratégias de crescimento e regeneração de plantas. A adaptação destas à luz durante a fase inicial do desenvolvimento condiciona mudanças em sua estrutura e metabolismo que determinarão o sucesso do estabelecimento e crescimento de espécies vegetais (TURNBULL, 1991; AGYEMAN; SWAINE; THOMPSON, 1999).
A capacidade de algumas espécies em manter populações de plântulas e plantas jovens no sub-bosque de florestas está associada à sua sobrevivência sob baixas irradiâncias, ou seja, tolerar o sombreamento (WALTERS; REICH 2000). Segundo Valladares et al. (2000), florestas tropicais possuem um mosaico de diferentes ambientes de luz, variando de sub-bosques sombreados a clareiras de vários tamanhos causadas por queda de árvores, levando a uma grande heterogeneidade de adaptações a esses diferentes gradientes de luz.
Segundo Barbosa (2003), na fase de germinação algumas espécies da caatinga se comportam como tolerantes à sombra tendo em vista as condições ambientais desse tipo de vegetação na época chuvosa (dois a quatro meses), que apresenta cobertura das copas, maior quantidade de estrato herbáceo e serrapilheira. E, posteriormente, com a chegada do período de estiagem (sete a oito meses), há queda das folhas na maioria das espécies, proporcionando grande exposição à luz. Portanto sobreviverão as plântulas com adaptações para resistir ao estresse hídrico.
luminosidade, principalmente quanto ao crescimento vegetativo da parte aérea e à sobrevivência das mudas.
Finegan (1984) afirmou que as espécies pioneiras apresentam nos primeiros anos de
desenvolvimento, maior crescimento vegetativo do que as espécies não pioneiras. Em um estudo
com 10 espécies pioneiras e 10 não-pioneiras de Mata Atlântica, Ferez (2010), observou que as
espécies pioneiras apresentaram uma menor alocação de biomassa nas raízes, do que as espécies
não-pioneiras, respectivamente 26 e 32%. Indicando que provavelmente as espécies pioneiras têm
entre suas estratégias de sobrevivência uma maior alocação para a parte aérea, garantindo a sua
captura de luz frente às demais espécies competidoras. Já as não-pioneiras por naturalmente
ocorrerem em ambientes mais sombreados tendem a investir mais no sistema radicular enquanto
aguardam a oportunidade do aparecimento de uma clareira.
Ressalta-se que conhecer as características do crescimento inicial de plântulas sob diferentes níveis de irradiância é essencial para o entendimento do processo sucessional e para a avaliação do potencial de espécies nativas em eventuais programas de revegetação.
3 MATERIAL E MÉTODOS
3.1 Preparo de exsicatas e coleta de frutos
Amostras de ramos com flores e frutos de Piptadenia stipulacea (Benth.) Ducke e
Anadenanthera colubrina (Vell.) Brenan var. cebil (Griseb.) Altshul foram coletadas entre abril e maio/2010 de plantas matrizes localizadas em área de caatinga, situada na fazenda “Não Me Deixes”, Reserva Particular do Patrimônio Natural – RPPN (4°49’34’’S, 38°58’9’’W e 210 m de altitude), em Quixadá, Ceará. O município apresenta clima caracterizado como tropical quente semiárido, temperatura média de 26-28ºC e pluviometria média anual de 838 mm, com chuvas concentradas de fevereiro a abril (IPECE, 2010).
Após coleta, o material botânico foi herborizado e P. stipulacea e A. colubrina
foram depositadas como exsicatas, respectivamente, sob números 47.038 e 46.898 no Herbário Prisco Bezerra – EAC, pertencente ao Departamento de Biologia da Universidade Federal do Ceará.
Quanto às coletas de frutos para a realização dos experimentos, foram realizadas viagens freqüentes à RPPN de Quixadá (dezembro/2009 a setembro/2010), quando coletou-se, manualmente, frutos maduros de pelo menos sete indivíduos adultos de cada espécie, visando obter uma variabilidade genética na amostra.
3.2 Espécies focais
3.2.1 Piptadenia stipulacea (Benth.) Ducke
É uma espécie da família Mimosaceae, conhecida popularmente como jurema branca, que ocorre na caatinga, do Piauí até a Bahia (BRAGA, 1976). Esta árvore é pequena, com cerca de 2-4m de altura, casca castanho-claro, fortemente armada por acúleos vigorosos. Possui folhas alternas, compostas, com 10-16 pares de pinas opostas, cada pina com 2 a 5,5cm de comprimento e com 25-40 pares de folíolos verde-claros, foscos, oblongos, com 3-8mm de comprimento. As flores em espigas possuem de 4-8cm de comprimento, de cor alva, na extremidade dos ramos onde se encontram até três espigas por axila de folha. Seu fruto é uma vagem de cor castanho-pálido, com 8-12cm de comprimento, com superfície ondulada nas áreas onde ficam as sementes. Contém 2-12 sementes ovais pequenas, por vagem, de cor marrom. Segundo Barbosa, Barbosa e Lima (2003) esta espécie apresenta fruto de consistência seca, a semente é a unidade de dispersão, sendo que esta caracteriza-se como autocórica. A floração ocorre no período chuvoso e a frutificação no período seco para o chuvoso.
Pereira et al. (2001) afirmaram que o comportamento da P. stipulacea reflete, nitidamente, os efeitos das perturbações a que uma vegetação foi submetida, mostrado-se bastante tolerante a elevados níveis de perturbação.
3.2.1 Anadenanthera colubrina (Vell.) Brenan var. cebil (Griseb.) Altshul
A. colubrina caracteriza-se como uma espécie decídua. Segundo Maia (2004),nos primeiros dias do período seco, inicia-se a queda das folhas que só serão produzidas novamente no final da estação seca/início da chuvosa. A floração acontece no período seco e, ao final dessa estação, ocorre a frutificação, seguida da maturação e dispersão.
Esta espécie possui grande demanda no mercado, sendo utilizada como ornamental, fornecedora de tanino e mel, forrageira, energética, resinífera, madeireira e, inclusive, medicinal. Esse uso intenso, até por indústrias, coloca em risco a existência dessa e de outras espécies, aliado à ausência de cultivos ou de métodos de propagação, ao desconhecimento da real distribuição das populações nativas e à ausência de estudos que avaliem o impacto do extrativismo na estrutura e biologia dessas populações (ALBUQUERQUE; ANDRADE, 2002).
Alguns autores (SILVA et al., 2003; PAULA et al., 2004) desenvolveram pesquisas em floresta estacional semidecidual, envolvendo classificação de espécies em grupos ecológico, e citaram Anadenanthera colubrina como uma espécie secundária inicial, de acordo com Gandolfi, Leitão Filho e Bezerra (1995).
3.3 Teste preliminar de germinação e superação da dormência
Para verificar se as espécies apresentavam dormência, foi realizado um teste preliminar de germinação em dezembro/2009. No Laboratório de Análises de Sementes da UFC, as sementes selecionadas de P. stipulacea e A. colubrina foram colocadas em placas de Petri de 15 cm de diâmetro, previamente esterilizadas e forradas com dupla camada de papel filtro umedecido com água destilada, com volume correspondente a duas vezes o peso do papel. As placas foram colocadas em câmara tipo BOD, com temperatura constante de 30ºC e fotoperíodo de 12hs, monitoradas diariamente, durante 30 dias, e umedecidas com água destilada, sempre que necessário. Ao final do experimento, calculou-se a porcentagem de germinação para cada espécie.
dessecação e irradiância, bem como subsidiar futuros trabalhos de germinação e propagação de mudas desta espécie.
Dessa forma, o ensaio constou de seis tratamentos: escarificação mecânica com lixa; imersão em ácido sulfúrico por 5, 10 e 15 minutos (seguidas da lavagem em água corrente até a total retirada do ácido); imersão em água quente (a 80º C até o esfriamento da água a temperatura ambiente) e sementes sem nenhum tratamento (testemunha).
Depois de submetidas aos tratamentos, as sementes foram levadas para o Núcleo de Ensino e Pesquisa em Agricultura Urbana – NEPAU da UFC e semeadas em bandejas plásticas de 200 células, contendo mistura de areia e húmus na proporção volumétrica de 3:1. O experimento foi conduzido em casa de vegetação, onde foram feitas irrigações diárias, por meio de micro-aspersão e nebulização, para manter a umidade adequada à germinação das sementes, sempre na mesma quantidade para todas as bandejas.
Foram realizadas contagens diárias das plântulas emergidas que apresentavam cotilédones expandidos. As contagens foram cessadas quando não ocorreu mais emergência em nenhum dos tratamentos, ou seja, aos 16 dias após a semeadura. Em seguida avaliou-se as seguintes variáveis:
Porcentagem de emergência - calculada de acordo com a fórmula (1).
PE = (N/A) x 100 (1)
Em que:
PE - Porcentagem de emergência,
N - Número total de plântulas emergidas,
A - Número total de sementes semeadas.
Índice de velocidade de emergência - é fundamentado na emergência diária, e foi obtida através da fórmula (2) proposta por Maguire (1962).
IVE = (E1 + E2 + ... + En) / ( N1 + N2 +...+ Nn) (2) Em que:
IVE - Índice de velocidade de emergência,
E1, E2 ... En - Número de plântulas normais emergidas na primeira, na segunda até a última
contagem.
Tempo médio de emergência - é expresso como o tempo médio necessário para atingir o número de emergência máxima, e foi obtida através da fórmula (3) proposta por Laboriau (1983).
TME = Σniti/ Σni (3) Em que:
TME -tempo médio de emergência,
ni-número de plântulas emergidas no intervalo entre cada contagem,
ti -tempo médio decorrido entre o início da emergência e a i-ésima contagem.
Adotou-se um delineamento experimental inteiramente casualizado com seis tratamentos de 100 sementes cada, distribuídas em quatro repetições de 25 sementes. Os dados foram submetidos à análise de variância e as médias comparadas pelo teste de Tukey a 5% de probabilidade (BANZATTO; KRONKA, 2006), utilizando-se o programa SISVAR (FERREIRA, 2008).
3.4 Influência da temperatura e luz na germinação de sementes
Para este experimento, as vagens das espécies focais foram coletadas em agosto/2010. Após serem beneficiadas, apenas as sementes de P. stipulacea foram submetidas ao tratamento de superação de dormência (ácido sulfúrico por 10 minutos) e, em seguida, as sementes de ambas as espécies foram colocadas em placas de Petri de 15 cm de diâmetro, previamente esterilizadas e forradas com dupla camada de papel filtro umedecido com água destilada, com volume correspondente a duas vezes o peso do papel. As placas foram monitoradas diariamente e umedecidas com água destilada, sempre que necessário.
As avaliações foram realizadas a cada 24 horas por um período de 30 dias, adotando-se como caráter germinativo a protrusão da radícula a 2 mm de comprimento. Ao final de cada ensaio, calculou-se a porcentagem de germinação – PG, o índice de velocidade de germinação – IVG (MAGUIRE, 1962) e o tempo médio de germinação – TMG (LABORIAU,1983).
Adotou-se um delineamento experimental inteiramente casualizado, em esquema fatorial 2x10 (duas condições de luz e dez temperaturas), com quatro repetições de 25 sementes, totalizando 100 sementes por tratamento para cada espécie. Os dados foram submetidos à análise de variância e as médias comparadas pelo teste de Tukey a 5% de probabilidade (BANZATTO; KRONKA, 2006), utilizando-se o programa SISVAR (FERREIRA, 2008).
3.5 Tolerância à dessecação em sementes
Para este experimento os frutos das duas espécies foram coletados em setembro/2010, antes da deiscência, quando as vagens de P. stipulacea adquiriram cor castanho-pálido e as de A. colubrina cor marrom-escura.
Após serem beneficiadas, amostras de sementes frescas (zero horas de secagem), de ambas as espécies, foram utilizadas como testemunhas para avaliar o teor de água inicial e teste de germinação (descritos a seguir). Em seguida, todas as sementes das duas espécies foram submetidas à secagem em câmara com ventilação forçada e ar aquecido a 35ºC. Para se obter tratamentos com diferentes teores de umidade, amostras de sementes foram retiradas da câmara após 12, 24, 36, 48, 60, 72 e 84 horas de secagem. Para facilitar a liberação da água no ambiente da câmara, as sementes foram colocadas em telas metálicas horizontais, fixadas na posição mediana de caixas tipo gerbox. Para que as sementes alcancem baixos conteúdos de água, foi coloca sílica gel sob as telas, que era trocada diariamente, antes de ocorrer mudança na coloração do seu indicador de umidade.
Ao retirar a amostra de cada período de secagem, verificou-se o teor de água e realizou-se o teste de germinação, descritos a seguir:
fórmula (4). A pesagem foi expressa em gramas, com três casas decimais e o resultado final foi obtido através da média aritmética das porcentagens de cada uma das duas repetições retiradas da amostra.
% de Umidade (U) = 100 (P–p)/P–t (4)
Em que:
P = peso inicial, peso do recipiente e sua tampa mais o peso da semente úmida; p = peso final, peso do recipiente e sua tampa mais o peso da semente seca; t = tara, peso do recipiente com sua tampa.
Teste de germinação – foi realizado em câmaras tipo BOD, com circulação interna de água, reguladas para a temperatura constante de 30 ± 1ºC e fotoperíodo de 12 horas, pois testes preliminares mostraram que esta temperatura proporcionou 100% de germinação para ambas as espécies. Diante do conhecimento prévio de que sementes de P. stipulacea
apresentam dormência física, estas foram submetidas à quebra de dormência, utilizando-se ácido sulfúrico por 10 minutos, sabendo-se que esta técnica caracteriza-se como a melhor, conforme testes realizados em laboratório. As sementes foram colocadas em placas de Petri de 15 cm de diâmetro, previamente esterilizadas e forradas com dupla camada de papel filtro, umedecido com 5mL de água destilada, volume correspondente a duas vezes o peso do papel. As placas foram monitoradas diariamente e umedecidas sempre que necessário. O critério de avaliação utilizado foi o da protrusão da radícula (2 mm de comprimento). Ao final de cada ensaio, calculou-se a porcentagem de germinação e o índice de velocidade de germinação.
O delineamento experimental foi inteiramente casualizado, com quatro repetições de 25 sementes em cada tratamento, constando de oito tratamentos de secagem para cada espécie. Os dados foram submetidos a análises de variância e as médias comparadas pelo teste de Tukey a 5% de probabilidade (BANZATTO; KRONKA, 2006), utilizando-se o programa SISVAR (FERREIRA, 2008).
3.6 Crescimento inicial de plântulas em diferentes níveis de irradiância.
Para este experimento, as vagens das espécies focais foram coletadas no início de maio/2010. Após serem beneficiadas no Laboratório de Análises de Sementes da UFC, apenas as sementes de P. stipulacea foram submetidas ao melhor tratamento de superação de dormência (ácido sulfúrico por 10 minutos), conforme experimento de superação de dormência realizado anteriormente. Em seguida, as sementes foram conduzidas para o Núcleo de Ensino e Pesquisa em Agricultura Urbana (NEPAU) do Centro de Ciências Agrárias da Universidade Federal do Ceará, situada nas coordenadas geográficas 03º 44’ 17” S e 38º 34’ 22” W, com altitude de 12 m.
No NEPAU foi realizada a semeadura indireta das sementes de P. stipulacea e A. colubrina, em bandejas plásticas contendo mistura de solo (extraído da área de caatinga em Quixadá) e húmus na proporção volumétrica de 3:1. Esta etapa do experimento foi conduzida em casa de vegetação, onde foram feitas irrigações diárias, por meio de micro-aspersão e nebulização, sempre na mesma quantidade para todas as bandejas, para manter a umidade adequada para a emergência das plântulas.
Após atingirem cinco centímetros de altura (15 dias após semeadura), as plântulas foram transplantadas para sacos de polietileno preto-opaco de 15 x 25cm com perfurações laterais e substrato contendo solo de Quixadá e húmus na proporção volumétrica de 2:1. No dia 05/junho (30 dias após a semeadura) realizou-se a primeira coleta de dados, que se caracterizou como mensurações da altura, diâmetro do coleto e número de folhas de cada plântula, em cada espécie. A altura da plântula, medida com régua milimetrada, consistiu na distância entre o nível do solo e a gema apical. O diâmetro do coleto foi medido com o uso de paquímetro com precisão de 0,01 cm. Após esta primeira coleta, as plântulas foram submetidas aos tratamentos com diferentes graus de irradiância.
Os sacos com as plântulas foram colocados sobre uma base de 1,0 m2 feita com tijolos de cerâmica, elevada a 15cm do nível do solo e sobre cada base foi colocada uma armação de 2,0m de altura, confeccionada com ferro e madeira, e coberta com tela sombrite, permitindo a passagem de 50%, 30% e 10% da luz solar incidente, conforme especificação do fabricante das telas.
Os quatro tratamentos realizados consistiram em:
Tratamento 3 – 70% de sombreamento (30% em relação ao pleno sol). Tratamento 4 – 90% de sombreamento (10% em relação ao pleno sol).
Após um mês em que as plântulas foram submetidas às condições de tratamento citadas, procederam-se mais cinco avaliações mensais, na mesma unidade experimental, para mensuração das variáveis citadas anteriormente.
Os tratamentos foram dispostos em área experimental seguindo um delineamento em blocos ao acaso com quatro repetições, sendo cada uma delas constituída por 11 mudas. As variáveis respostas foram mensuradas no programa Statistica 7.0 (STATSOFT, 2004), utilizando uma análise de variância para medidas repetidas.
4 RESULTADOS E DISCUSSÃO
4.1 Teste preliminar de germinação e superação da dormência
O teste de germinação com P. stipulacea, mostrou que apenas 10% das suas sementes germinaram, em até 30 dias, e as demais permaneceram duras, devido ao tegumento pouco permeável a água. Então, como previsto, constatou-se que as sementes desta espécie apresentam dormência física. Segundo Vasquez-Yanes e Orozco-Segovia (1989), a dormência é uma característica típica das espécies pioneiras, que também apresentam longa longevidade natural e são capazes de formar banco de sementes.
Quanto à A. colubrina, o teste preliminar revelou que a espécie não apresenta mecanismo de dormência, visto que sua porcentagem de germinação foi de 95% em 48hs. Este dado já havia sido ressaltado na revisão feita por Barbosa (2003), a qual mostra que A. colubrina apresentou 80 a 100% de germinação após 48 horas de germinação, justificando a não aplicação de tratamentos para ativar a germinação. A autora também ressalta que talvez isso se deva ao tegumento delicado e a presença na testa da linha fissural (pleurograma), que facilita a penetração da água.
Algumas espécies que apresentam pouco retardo entre a dispersão e a germinação não apresentam dormência nas sementes e não desenvolvem um banco de sementes persistente. Silva (1998), ao observar a fenologia de A. colubrina verificou que sua frutificação se dá no início das chuvas (janeiro a fevereiro), com a dispersão das sementes em plena estação seca (novembro a dezembro), germinando facilmente e de imediato a estação chuvosa seguinte. A autora concluiu que a espécie não forma banco de sementes no solo, uma vez que não foram encontradas sementes das safras anteriores, somente da atual, reconhecidas pelo aspecto reluzente do tegumento.
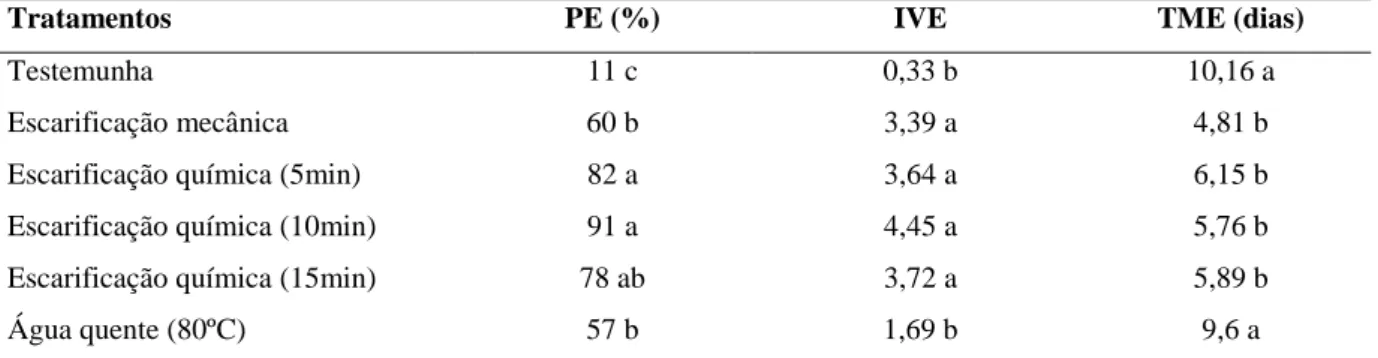
No experimento de superação de dormência das sementes de P. stipulacea, foi constatado que os tratamentos com ácido sulfúrico foram os mais eficientes em promover a germinação das sementes, apresentando resultados superiores aos demais tratamentos (Tabela 1).
Tabela 1 – Porcentagem de emergência (PE), índice de velocidade de emergência (IVE) e tempo médio de
emergência (TME) de sementes de Piptadenia stipulacea, submetidas a diferentes tratamentos de superação de dormência. Fortaleza, CE, 2011.
Tratamentos PE (%) IVE TME (dias)
Testemunha 11 c 0,33 b 10,16 a
Escarificação mecânica 60 b 3,39 a 4,81 b
Escarificação química (5min) 82 a 3,64 a 6,15 b
Escarificação química (10min) 91 a 4,45 a 5,76 b
Escarificação química (15min) 78 ab 3,72 a 5,89 b
Água quente (80ºC) 57 b 1,69 b 9,6 a
Médias seguidas da mesma letra, nas colunas, não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Observou-se que a porcentagem de emergência aumentou de 11% para 82% e 91% quando se utilizaram os tratamentos de escarificação química a 5 e 10 minutos, respectivamente. Em uma espécie do mesmo gênero, Piptadenia moniliformis, Benedito et al.
(2008) verificaram que os tratamentos com ácido sulfúrico por 10 e 15 minutos proporcionaram maior porcentagem de germinação, com 81% e 82%, respectivamente. Entretanto, no presente estudo, com P. stipulacea, foi constatado que o aumento no tempo de imersão em ácido sulfúrico, para 15 minutos, reduz a porcentagem de emergência (Tabela 1). Este resultado assemelha-se ao encontrado por Sampaio et al. (2001), no qual sementes de
Bowdichia virgilioides têm a emergência prejudicada com tempo de imersão acima de 11 minutos. Esse fato pode estar relacionado aos efeitos danosos do ácido sulfúrico no embrião, uma vez que Rolston (1978) relatou que a escarificação química propicia a degradação do tegumento e o aumento do período de imersão pode causar ruptura das células essenciais, o que favorece as injúrias mecânicas e a invasão de fungos, prejudicando, assim, a emergência.
OLIVEIRA, 1992), Mimosa bimucronata (RIBAS; FOSSATI; NOGUEIRA,1996) e Bauhinia monandra (ALVES et al., 2000).
A escarificação mecânica foi relativamente satisfatória, quanto ao IVE, pois não diferiu significativamente das escarificações químicas. A eficiência desse método também foi registrada nas sementes das leguminosas: Senna macranthera (SANTARÉM; ÁQUILA, 1995), Acacia mearnsii (ROVERSI et al., 2002) e Bauhinia divaricata (ALVES et al., 2004).
A imersão em água quente (80ºC) apresentou baixo índice de velocidade de emergência, sendo semelhante ao da testemunha. Este tratamento também não foi eficiente na superação da dormência de sementes de Mimosa caesalpiniaefolia (BRUNO; ALVES; OLIVEIRA, 2001). Entretanto, Smiderle, Mourão Júnior e Souza (2005) constataram a eficiência deste método na superação da dormência de Acacia margium.
Os tratamentos de escarificação química e física foram favoráveis para um menor tempo médio de emergência. Já as sementes intactas (testemunha) e tratadas com imersão em água quente (80ºC) apresentaram maior tempo, com aproximadamente 10 e 9 dias, respectivamente. Contudo, esta emergência distribuída ao longo do tempo de incubação é uma característica importante para a sobrevivência da espécie em condições naturais.
Ressalta-se que este experimento foi eficiente na determinação do melhor tratamento de superação de dormência das sementes de P. stipulacea, subsidiando os experimentos de temperatura e luz, dessecação e irradiância.
4.2 Influência da temperatura e da luz na germinação de sementes
Tabela 2 – Resumo da Análise de variância dos dados de porcentagem de germinação (PG), índice de velocidade
de germinação (IVG) e tempo médio de germinação (TMG) de sementes de Piptadenia stipulacea, submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011.
** - Significativo a 1% de probabilidade; ns - Não-significativo.
P. stipulacea apresentou germinação na faixa das temperaturas constantes de 10ºC a 40ºC. O processo germinativo foi inibido a 45ºC, indicando que sua temperatura máxima de germinação está entre 35ºC e 40ºC (Tabela 3). Os resultados obtidos concordam com a afirmação de Okusanya (1978), segundo a qual as sementes de espécies tropicais são tolerantes à alta temperatura, apresentando limite máximo igual ou superior a 35ºC, porém sensíveis à baixa temperatura, com limite mínimo superior a 5ºC.
Tabela 3 – Médias da porcentagem de germinação (PG), índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG) de sementes de Piptadenia stipulacea, submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011.
Médias seguidas da mesma letra, maiúsculas nas colunas e minúsculas nas linhas, não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Os melhores valores de porcentagem de germinação, índice de velocidade de germinação e tempo médio de germinação ocorreram na temperatura constante de 30ºC. A
Fontes de variação GL
Valor de F
PG (%) IVG TMG (dias)
Luz (L) 1 0,80 ns 0,009 ns 0,13 ns
Temperatura (T) 9 1327,84 ** 2152,43 ** 571,19 **
Interação L x T 9 1,57 ns 0,75 ns 2,88 **
Resíduo 60
CV ( % ) 6,33 6,56 11,93
PG (%) IVG TMG (dias)
Temperatura Luz Média Luz Média Luz Média
Sem Com Sem Com Sem Com
10ºC 5 9 7 e 0,08 0,13 0,10 f 16,38 Ba 18,65 Aa 17,51
15ºC 33 27 30 d 0,55 0,50 0,52 e 16,61 Aa 15,20 Bb 15,91
20ºC 99 98 98,5 ab 6,45 6,62 6,54 b 4,00 Acd 3,83 Ade 3,92
25ºC 98 98 98 abc 10,18 10,35 10,27 a 2,57 Ad 2,52 Ae 2,54
30ºC 100 100 100 a 10,38 10,08 10,22 a 2,51 Ad 2,58 Ae 2,55
35ºC 97 91 94 bc 4,70 4,46 4,58 d 5,30 Ac 5,28 Ad 5,29
40ºC 31 32 31,5 d 0,77 0,86 0,81 e 10,46 Ab 10,38 Ac 10,42
45ºC 0 0 0 f 0,00 0,00 0,00 f 0,00 Ae 0,00 Af 0,00
15-45ºC 0 0 0 f 0,00 0,00 0,00 f 0,00 Ae 0,00 Af 0,00
20-40ºC 92 93 92,5 c 5,76 5,82 5,79 c 4,11 Acd 4,11 Ade 4,11
temperatura alternada de 20-40ºC também apresentou alta porcentagem de germinação (95%), porém, a 15-45ºC o processo germinativo foi inibido (Tabela 3). Em alguns casos a alternância de temperatura pode aumentar consideravelmente a germinação, como foi verificado em sementes Bixa orellana (GOMES; BRUNO, 1992) e Trema micrantha
(CASTELLANI; AGUIAR, 1998). Todavia, também pode inibir, parcialmente, o desenvolvimento do processo de germinação, a exemplo dos resultados encontrados em sementes de Cedrela odorata (ANDRADE; PEREIRA, 1994), e Capsicum annuum (POSSE
et al., 2001).
Nas temperaturas baixas de 10ºC e 15ºC, P. stipulacea apresentou valores muito baixos de porcentagem de germinação (Tabela 3), o que indica que esta espécie pode não apresentar germinação satisfatória em sub-bosque e necessita de temperaturas mais altas, como as que ocorrem em clareiras. Segundo Vázquez-Yanes e Orozco-Segovia (1982), quando uma clareira é aberta, além da mudança na intensidade de luz, ocorre também um aumento na flutuação de temperatura diária na superfície do solo.
Maia et al. (2008), verificaram que Hyptis suaveolens não apresentou germinação abaixo de 20ºC e justificaram ao fato da espécie ser comum em locais que foram submetidos a ação antrópica, o que sugere que suas sementes estejam expostas a uma maior amplitude de temperatura.
No experimento realizado com A. colubrina verificou-se que somente a temperatura exerceu efeito significativo na porcentagem de germinação, índice de velocidade de germinação e tempo médio de germinação. As sementes mostraram-se indiferentes ao fator luz e à interação luz e temperatura, não havendo efeito significativo em nenhuma das variáveis analisadas (Tabela 4).
Tabela 4 – Resumo da Análise de variância dos dados de porcentagem de germinação (PG), índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG) de sementes de Anadenanthera colubrina,
submetidas a diferentes condições de luz e temperatura. Fortaleza, CE, 2011.
Fontes de variação GL
Valor de F
PG (%) IVG TMG (dias)
Luz (L) 1 0,9 ns 2,59 ns 1,42 ns
Temperatura (T) 9 1670,89 ** 414,89 ** 56,86 **
Interação L x T 9 1,46 ns 1,18 ns 1,05 ns
Resíduo 60
CV ( % ) 3,82 9,98 36,79