O R I G I N A L A R T I C L E
Characterization of microbial population of ‘Alheira’
(a traditional Portuguese fermented sausage) by PCR-DGGE
and traditional cultural microbiological methods
H. Albano1, I. Henriques2, A. Correia2, T. Hogg1and P. Teixeira1 1 Escola Superior de Biotecnologia, Universidade Cato´lica Portuguesa, Porto, Portugal 2 CESAM and Departamento de Biologia, Universidade de Aveiro, 3810-193 Aveiro, Portugal
Introduction
The microbiology of fermented sausages is complex. The type of microflora that develops is often closely related to the ripening technique utilized. The manufacture of fer-mented sausages has a long history in Portugal where there is a wide variety of typical preparations.
‘Alheira’ is a traditional fermented meat sausage typical of the North of Portugal (Tra´s-os-Montes). ‘Alheira de Mirandela’ is a much appreciated product and represents an important resource for this region where more than 500 tons are produced annually. ‘Alheira de Mirandela’ is in the process of name registration as ‘Traditional Guar-anteed Speciality’. The specific characteristics of the final
product mainly arise from the raw materials employed, the agro-ecosystem of the area of production and the tra-ditional technology of manufacture. ‘Alheira de Mirandela’ is produced from pork and poultry meat, lard and pork fat, wheat bread and olive oil. Ingredients are mixed with salt, garlic and spices until they form a paste which is then stuffed into natural or artificial casings (horseshoe-shaped sausages approximately 15 ± 60 mm long) and submitted to a smoking process, at low but uncontrolled temperature (generally <40!C) and uncontrolled humi-dity, for no longer than 8 days. The shelf life of ‘Alheiras’ is about 1 month if stored at 4!C in air or longer if the sausages are packed under vacuum or modified atmo-sphere. ‘Alheiras’ are cooked before consumption either
Keywords
fermented sausages, fingerprint, lactic acid bacteria, PCR-denaturing gradient gel electrophoresis.
Correspondence
Paula Teixeira, Rua Dr. Anto´nio Bernardino de Almeida, 4200-072 Porto, Portugal. E-mail: [email protected]
2008⁄ 0938: received 2 June 2008, revised 10 July 2008 and accepted 22 July 2008 doi:10.1111/j.1365-2672.2008.03947.x
Abstract
Aims: This study evaluates the microbial ecology of ‘Alheira’ by traditional microbiological analysis and a PCR-denaturing gradient gel electrophoresis (DGGE) protocol.
Methods and Results: Total microbial DNA from ‘Alheiras’ was extracted directly from the products and subjected to PCR using Eubacterial primers for 16S rDNA. The amplicons were separated by DGGE. The results demonstrated that different products of the same batch display identical profiles, whereas products from different batches of the same producer could display different DGGE profiles. ‘Alheiras’ from different producers were distinguishable based on the respective DGGE profiles. The obtained sequences from prevalent phylotypes affiliated with order Lactobacillales and order Bacillales and class Gammaproteobacteria. The same samples were subjected to traditional microbio-logical analysis. In both methods, lactic acid bacteria were dominant and were present together with other organisms, mainly members of the family Micro-coccaceae.
Conclusions: The approach explored in this study allowed the description of the microbial community present in ‘Alheira’ in particular the diversity of lac-tic acid bacteria.
Significance and Impact of the Study: This can be useful for the microbiologi-cal characterization of traditional products in order to develop new methods of quality control capable of supporting a standardization of the processes, while preserving their typical traits.
by frying, grilling or boiling, according to regional tradi-tions or consumer preferences. A wide variety of micro-organisms has already been isolated from ‘Alheiras’ by traditional methods. These are mainly lactic acid bacteria (LAB), Micrococcaceae and some pathogens, such as Liste-ria spp., Salmonella and Staphylococcus aureus (Ferreira et al. 2006; Esteves et al. 2008).
Genetic fingerprinting techniques that use DNA, gener-ated by polymerase chain reaction (PCR) procedures, applied DNA extracted directly from environmental samples, can be used to provide profiles representing the phylogenetic diversity of the microbial community in the environment being studied. This approach eliminates the necessity for traditional culture-dependent methods like selective cultivation and strain isolation, thereby negating the potential biases inherent in such procedures. Moreover, studies which have employed such direct anal-yses have repeatedly demonstrated considerable variance between cultivated and naturally occurring species, thereby dramatically altering the perception of the true microbial diversity present in various habitats (Head et al. 1998; Hugenholtz et al. 1998; Cocolin et al. 2002a). Dena-turing gradient gel electrophoresis (DGGE) is perhaps the most commonly used among the nonculture-dependent fingerprinting techniques. It is based on the separation of PCR amplicons of the same size according to their differ-ent melting temperatures. PCR-DGGE of ribosomal DNA was introduced into microbial ecology by Muyzer et al. (1993). Despite the wide range of applications, this tech-nique has been introduced into food microbiology no longer than 10 years. The great potential shown in analys-ing samples from natural environments without emplo-ying a culture-based isolation, has stimulated food microbiologists to investigate the suitability of PCR-DGGE to study microbial fermentations in food and food-related ecosystems. In the last few years, several papers have been published reporting the use of DGGE for LAB strain identification (Cocolin et al. 2000; Ercolini et al. 2001a; Ogier et al. 2002), for the study of the eco-logy of fermented foods and the microbial changes in LAB populations during these food fermentations and to profile pathogens directly in food samples (Cocolin et al. 2002b). In particular, direct PCR amplification of differ-ent regions of the 16S rRNA gene and subsequdiffer-ent analysis by DGGE has been used to study the ecology of the microbial processes involved in the production of Mexican fermented maize dough (ben Omar and Ampe 2000), Mozzarella cheese (Coppola et al. 2001), fermented sausages (Cocolin et al. 2001a,b, 2004; Fontana et al. 2005; Rantsiou et al. 2005; Aquilanti et al. 2007), malt whisky (van Beek and Priest 2002), artisanal Sicilian cheese (Randazzo et al. 2002), buffalo Mozzarella and Stilton cheese (Ercolini et al. 2001b, 2003) and sourdough
(Meroth et al. 2003). The use of DGGE in food microbi-ology was reviewed by Ercolini (2004).
In this work, the microbial ecology of ‘Alheiras’, produced in six different plants in North-East Portugal, was studied by using culture-dependent and -independent methods. ‘Alheiras’ were subjected to traditional micro-biological analysis in order to enumerate specific bacterial groups. The aim of this study was also to evaluate if PCR-DGGE could be useful to identify specific motifs in the bacterial community profiles of these products. To accomplish this aim, DNA was extracted directly from the same samples and 16S DNA fragments were amplified by PCR, using ‘universal’ bacterial primers. DGGE analysis allowed fingerprinting of the microbial populations pres-ent in ‘Alheira de Mirandela’ and determination of the DNA sequence of relevant bands permitted the establish-ment of the phylogenetic affiliation of bacteria present.
Materials and methods
Sampling
‘Alheiras’ from six different producers (two or three batches of each), were purchased from retail stores, at storage time, during 2005. Samples were labelled as Ef, Tp, Tx, Gr, Ag and Am to represent the six different producers.
Microbiological analyses by cultural methods
Samples of ‘Alheiras’ (25 g) were added to 225 ml of ste-rile buffered peptone water (Merck, Darmstadt, Germany), and homogenized in a stomacher for 2 min. Appropriate decimal dilutions were prepared in sterile Ringer’s solu-tion (LabM, Bury, UK) for microbial enumerasolu-tion: LAB on de Man, Rogosa Sharpe Agar (MRS, LabM) and on M17 (LabM), incubated at 30!C for 72 h; total counts at 30!C according to ISO Standard 4833 (ISO 2003); Entero-coccaceae on bile esculin azide agar (Biokar Diagnostics, Beauvais, France), incubated at 30!C for 72 h; Micrococca-ceae on mannitol salt agar (Biokar Diagnostics), incubated at 37!C for 48 h; Enterobacteriaceae according to the Portuguese Standard NP 4137 (IPQ 1991). Enumeration of Listeria spp. was performed on Palcam Agar (Merck) medium and incubated at 30!C for 72 h. Three inde-pendent analyses, using randomly selected pieces, were performed for each sample.
DNA extraction
Ten grams samples were homogenized in a stomacher bag with 10 ml of saline-peptone water for 1 min. After each preparation had settled for 1 min, 1 ml subsamples were
placed in 1Æ5-ml screw-cap tubes containing 0Æ3 g of glass beads. Total DNA was extracted from ‘Alheira’ following the method previously described for fermented sausages (Cocolin et al. 2001a), using treatment with vortex instead bead beader.
PCR-DGGE protocol
DNA was amplified with primers 338F (5¢-GAC-TCCTACGGGAGGCAGCAG-3¢) and 518R (5¢-ATTA-CCGCGGCTGCTGG-3¢), with a GC clamp attached to the forward primer (5), spanning the V3 region of the bacterial 16S rRNA gene (Muyzer et al. 1993). PCR was performed in a Bio-Rad iCycler Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA) as previously described (Henriques et al. 2006). Five microlitres of the product was analysed by standard agarose gel electrophoresis before DGGE analysis.
DGGE was performed on a DCodeTMUniversal Muta-tion DetecMuta-tion System (Bio-Rad Laboratories), according to Henriques et al. (2006). Briefly, samples were loaded onto 8% polyacrylamide gels (37Æ5 : 1, acrylamide: bis-acrylamide) in 0Æ5x TAE buffer (20 mmol l)1Tris-acetate, pH 7Æ4, 10 mmol l)1 sodium acetate, 0Æ5 mmol l)1 Na2EDTA) using a denaturing gradient ranging from 35–
50% (100% denaturant contains 7 mol l)1 urea and 40% formamide). Electrophoresis was performed at 60!C, ini-tially at 20 V (15 min) and then at 200 V (330 min). The gels were stained in an ethidium bromide solution (5 min) and then rinsed in distilled water (20 min). The image was acquired using a Molecular Image FX apparatus (Bio-Rad Laboratories). Every gel contained two lanes with a stan-dard of eight bands for internal and external normalization and as an indication of the quality of the analysis.
Sequencing of DGGE bands and phylogenetic analysis DGGE bands were excised with a sterile scalpel and eluted in 20ll of sterile water, overnight at 4!C. Five microlitres of the supernatant was used for re-amplification with the original primer set. The accuracy of the bands and the position in the gel were checked on DGGE gels together with the original sample. Whenever necessary, bands were processed again as described above.
For sequencing analysis, PCR products were purified with the Jetquick PCR Purification Spin Kit (Genomed, Lo¨hne, Germany) and used as template in the sequencing reactions. Those were carried out using the primer 518R and an ABI Prism"BigDyeTMTerminator Cycle Sequenc-ing Ready Reaction Kit (PE Applied Biosystems, Foster City, CA, USA). The reaction mixtures were analysed in an automatic DNA sequencer (ABI Prism" 310 Genetic Analyzer, PE Applied Biosystems).
Band sequences were compared to the GenBank nucle-otide data library using the Blast software at the National Center of Biotechnology Information website (http://www.ncbi.nlm.nih.gov/) in order to determine their closest phylogenetic relatives.
Nucleotide sequence accession numbers
The nucleotide sequences determined in the present study have been deposited in the GenBank database under the accession numbers EU555489–EU555518.
Results
‘Alheiras’ from six different producers, named Gr, Am, Ef, Tx, Ag and Tp (Fig. 1), were sampled during retail display at storage time from retail stores. From each pro-ducer, two or three batches (‘1’, ‘2’ and ‘3’ in Fig. 1) were used and, from each batch, two replicates were taken (‘a’ and ‘b’ in Fig. 1 correspond to the same batch). ‘Alheiras’ were subjected to traditional microbiological analysis and DNA, directly extracted from the samples, was subjected to DGGE analysis from which dominant bands were sequenced.
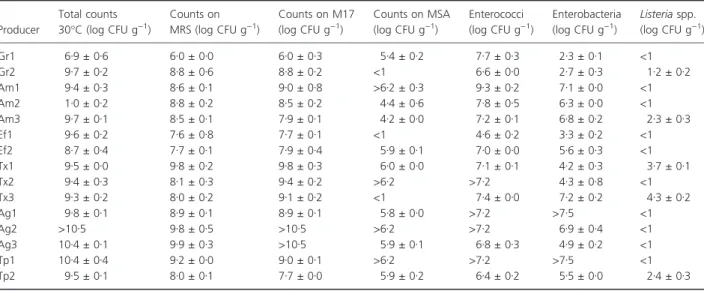
Microbiological characterization by cultural methods The results of the plate counts obtained from the six pro-ducers and their samples in this study are reported in Table 1. It is clear that there is variability in counts between producers and even between different lots from the same producer. The values of total counts at 30!C were greater than 8Æ7 log CFU g)1 in most samples with LAB being the dominant microflora. Counts on MRS and M17 were, in most cases, >7Æ5 log CFU g)1, with the exception of one sample from producer Gr (Table 1). In addition, enterococci counts were >6Æ5 log CFU g)1 in most samples. With the exception of one of the samples from producers Gr, Ef and Tx, counts on mannitol salt agar were >log 4Æ44 log CFU g)1, although values varied between 4Æ4 log CFU g)1and >6Æ20 log CFU g)1. Entero-bacteriaceae counts were >4 log CFU g)1in most samples, with the exception of producer Gr. Listeria spp. was iso-lated in countable quantities in some samples, the highest values coming from two samples from the producer Tx (3Æ8 CFU g)1and 4Æ3 CFU g)1).
DGGE analysis
A sample-specific fingerprint was obtained from total DNA directly extracted from ‘Alheiras’, after amplification of the V3 region of 16S rRNA gene followed by DGGE analysis (Fig. 1). Reproducibility of PCR amplification
and DGGE was confirmed by performing three replicate runs, the profiles proving to be highly reproducible between different gels and PCR runs. The total number of band positions detected in the three gels was 30 and the number of DGGE bands per sample varied between 2 and 10 indicating significant differences between samples in terms of their microbial diversity. Samples Gr1, Am3, Tx2, Ag1 and Tp2 displayed the most complex profiles, whereas samples Am1, Am2, Ef1 and Ef3 displayed simpler profiles.
In general, products from the same batch displayed identical profiles, with the exception of sample Tx1,
where products from the same batch displayed different DGGE profiles. On the other hand, for some producers, consistent profiles were evident within different batches of ‘Alheira’; certainly this was the case for producers Gr, Am, Ag and Tx. Producers Ef and Tp produced batches which were much less consistent in terms of the microbial ecology present.
Sequencing and identification of DGGE fragments Thirty-one bands were excised from the three DGGE gels and re-amplified with the primers 338F and 518R
Gr1a Gr1b Gr2a Gr2b Am1a Am1b Am2a Am2b Am3a Am3b Ef1a Ef1b Ef2a Ef2b Tx1a Tx1b Tx2a Tx2b Tx3a Tx3b Ag1a Ag1b Ag2a Ag2b Ag3a Ag3b Tp1a Tp1b Tp2a Tp2b
1 2 3 4 5 6 7 8 9 10 1211 3 13 15 16 17 18 19 2 21 3 23 24 25 26 27 2 29 21 31 32 33 2 35 36 37
Figure 1 DGGE gels showing PCR-amplified bacterial 16S rRNA gene profiles from ‘Alheiras’ from six different producers (Ag, Am, Ef, Gr, Tp, Tx; a and b correspond to the same batch; indicated on top of the lanes). Lane M: DGGE marker constructed using previously characterized 16S rRNA gene clones from environmental libraries (Henriques et al. 2004). Bands that were excised for sequence analysis are labelled with the same number as in Table 2 and indicated with an arrow.
Table 1 Microbiological characterization of ‘Alheira de Mirandela’ from six different producers (Ag, Am, Ef, Gr, Tp, Tx;): important microbial parameters in fermented products
Producer Total counts 30!C (log CFU g)1) Counts on MRS (log CFU g)1) Counts on M17 (log CFU g)1) Counts on MSA (log CFU g)1) Enterococci (log CFU g)1) Enterobacteria (log CFU g)1) Listeria spp. (log CFU g)1) Gr1 6Æ9 ± 0Æ6 6Æ0 ± 0Æ0 6Æ0 ± 0Æ3 5Æ4 ± 0Æ2 7Æ7 ± 0Æ3 2Æ3 ± 0Æ1 <1 Gr2 9Æ7 ± 0Æ2 8Æ8 ± 0Æ6 8Æ8 ± 0Æ2 <1 6Æ6 ± 0Æ0 2Æ7 ± 0Æ3 1Æ2 ± 0Æ2 Am1 9Æ4 ± 0Æ3 8Æ6 ± 0Æ1 9Æ0 ± 0Æ8 >6Æ2 ± 0Æ3 9Æ3 ± 0Æ2 7Æ1 ± 0Æ0 <1 Am2 1Æ0 ± 0Æ2 8Æ8 ± 0Æ2 8Æ5 ± 0Æ2 4Æ4 ± 0Æ6 7Æ8 ± 0Æ5 6Æ3 ± 0Æ0 <1 Am3 9Æ7 ± 0Æ1 8Æ5 ± 0Æ1 7Æ9 ± 0Æ1 4Æ2 ± 0Æ0 7Æ2 ± 0Æ1 6Æ8 ± 0Æ2 2Æ3 ± 0Æ3 Ef1 9Æ6 ± 0Æ2 7Æ6 ± 0Æ8 7Æ7 ± 0Æ1 <1 4Æ6 ± 0Æ2 3Æ3 ± 0Æ2 <1 Ef2 8Æ7 ± 0Æ4 7Æ7 ± 0Æ1 7Æ9 ± 0Æ4 5Æ9 ± 0Æ1 7Æ0 ± 0Æ0 5Æ6 ± 0Æ3 <1 Tx1 9Æ5 ± 0Æ0 9Æ8 ± 0Æ2 9Æ8 ± 0Æ3 6Æ0 ± 0Æ0 7Æ1 ± 0Æ1 4Æ2 ± 0Æ3 3Æ7 ± 0Æ1 Tx2 9Æ4 ± 0Æ3 8Æ1 ± 0Æ3 9Æ4 ± 0Æ2 >6Æ2 >7Æ2 4Æ3 ± 0Æ8 <1 Tx3 9Æ3 ± 0Æ2 8Æ0 ± 0Æ2 9Æ1 ± 0Æ2 <1 7Æ4 ± 0Æ0 7Æ2 ± 0Æ2 4Æ3 ± 0Æ2 Ag1 9Æ8 ± 0Æ1 8Æ9 ± 0Æ1 8Æ9 ± 0Æ1 5Æ8 ± 0Æ0 >7Æ2 >7Æ5 <1 Ag2 >10Æ5 9Æ8 ± 0Æ5 >10Æ5 >6Æ2 >7Æ2 6Æ9 ± 0Æ4 <1 Ag3 10Æ4 ± 0Æ1 9Æ9 ± 0Æ3 >10Æ5 5Æ9 ± 0Æ1 6Æ8 ± 0Æ3 4Æ9 ± 0Æ2 <1 Tp1 10Æ4 ± 0Æ4 9Æ2 ± 0Æ0 9Æ0 ± 0Æ1 >6Æ2 >7Æ2 >7Æ5 <1 Tp2 9Æ5 ± 0Æ1 8Æ0 ± 0Æ1 7Æ7 ± 0Æ0 5Æ9 ± 0Æ2 6Æ4 ± 0Æ2 5Æ5 ± 0Æ0 2Æ4 ± 0Æ3
(Table 2). Figure 1 shows the original gel from which the bands were excised together with band numbers. The sequence obtained from band 33 was of low quality and so not considered for phylogenetic analysis.
In Table 2 the closest relatives of DGGE band sequences is presented. In some cases retrieved sequences shared the same value of similarity with several closely related micro-organisms and in such cases only one example of each close relative is presented.
Most of the sequences were similar to 16S rRNA gene sequences reported for cultured bacteria present in food samples such as meat products, packaged fish, spoiled food, fermented vegetables and cheese (Table 2). How-ever, the DNA sequence of some bands affiliated to sequences with their origins in bacteria initially isolated from other sources such as soil, marine sediments, human blood, phyllosphere, animal gut, faeces and ice. Six bands (labelled in Fig. 1 as 6, 16, 21, 23, 31, 32 and 36) affili-ated with sequences corresponding to bacteria classified as ‘uncultured’ in the BLAST database. In the case of DNA sequences from bands 6 and 32, these could not be clearly affiliated to any phylogenetic group, displaying similarity
with 16S rRNA gene sequences from uncultured bacterial clones.
According to Blast results sequences affiliated with LAB, included in order Lactobacillales, with micro-organ-isms included in order Bacillales and with Gammaproteo-bacteria. Most of the profiles displayed clearly dominant bands (Fig. 1). Bands affiliated with Lactobacillus were found in samples from producers Ef, Ag and Tp; bands affiliated with Leuconostoc were found in producers Ef, Am and Tx (Band 3); Enterococcus were found only in samples form producer Tx; samples from producer Am have an intense band affiliated with Firmicutes (Band 21); samples form producer Tx have intense bands affiliated with Pediococcus and Lactococcus (Band 13 and 15, Fig. 1); band associated with Psychrobacter is intense in producer Gr. Band 2 was detected at the same position in all the six producers. To confirm that matching positions correspond to identical phylotypes, Band 2 was excised and sequenced from all samples. The same nucleotide sequence was obtained for each band. This band position was detected in most of the profiles (exceptions were samples Am3a and Am3b), although it was more
Table 2 Closest relatives of DGGE band sequences
Band no. Accession number Sample Closest relative (accession number) Similarity (%)
1 EU555489 Ef1a Lactobacillus sakei (EU081017.1) 100
2 EU555490 Ef1a Brochothrix thermosphacta (AY543029.1) 100
3 EU555491 Ef2a Leuconostoc lactis (AB295117.1) 98
4 EU555492 Ef2b Streptococcus lutetiensis (EU163503) 100
5 EU555493 Ef2b Macrococcus caseolyticus (EU048336.1) 100
6 EU555494 Tx1a Uncultured soil bacterium clone (EU052112.1) 100
7 EU555495 Tx1a Lactococcus lactis (EU080989.1) 99
8 EU555496 Tx1a Pseudomonas sp. (EF028699.1) 99
9 EU555497 Tx1b Leuconostoc mesenteroides (EU099617.1) 99
10 EU555498 Tx1b Weissella paramesenteroides (EF422380.1) 100
11 EU555499 Tx2b Enterococcus sp.(EU157915) 100
12 EU555500 Tx2b Enterococcus gallinarum (EF025908) 98
13 EU555501 Tx2a Pediococcus acidilactici (EF059987.1) 100
15 EU555502 Tx3a Lactococcus lactis BMG 125 (EU080989.1) 99
16 EU555503 Gr1a Uncultured Bacilli bacterium (EF706149.1) 100
17 EU555504 Gr1b Pseudomonas aeruginosa (DQ149582.1) 99
18 EU555505 Gr2b Psychrobacter sp. (EU075120.1) 100
19 EU555506 Gr2a Acinetobacter sp. (AB365066) 100
21 EU555507 Am2a Uncultured bacterium DGGE gel band (EU200335) 100
23 EU555508 Am3a Uncultured bacterium (AY511592.1) 99
24 EU555509 Am3a Klebsiella sp. (EU075144.1) 99
25 EU555510 Ag1a Lactobacillus curvatus (EU081014.1) 100
26 EU555511 Ag1a Bacillus sp. (DQ448756.1) 98
27 EU555512 Ag1b Psychrobacter sp. (AJ582399.1) 99
29 EU555513 Ag2a Lactobacillus brevis (EF120367.1) 98
31 EU555514 Ag3a Uncultured Bacilli bacterium (EF706149.1) 98
32 EU555515 Tp2a Uncultured soil bacterium clone G08 (EU052112.1) 99
35 EU555516 Tp2a Hafnia alvei strain (AM042710.1) 99
36 EU555517 Tp2b Uncultured Bacilli bacterium (EF706149.1) 100
pronounced in samples Gr2a, Gr2b, Tp2a and Tp2b. The sequence from this band was 100% identical to Brocho-thrix thermosphacta, isolated from modified-atmosphere-packed salmon and coalfish. In addition, Bands 7 and 15 were retrieved from different gels as presented in Fig. 1 and confirmed as identical.
Discussion
Studies on the micro-ecology of fermented sausages date back to the 1970s (Lu¨cke 1974). These earlier studies, based on traditional methods, described the microbial populations developing during production and storage. In the last few years, the possibility of using culture-independent molecular approaches has opened up areas of research. In this study we used PCR-DGGE analyses in combination with traditional microbiological analyses for studying the microbial diversity of ‘Alheira’. Microbial DNA was sampled directly in order to determine the diversity of the 16S rRNA genes of the most prominent bacteria, which may also reflect the microflora of ‘Alheira’. The most salient observation from Fig. 1 concerns the variability in the microecology between samples from dif-ferent producers, which is considerable, and that between lots from the same producer which is evident but lesser. Variability between producers is not, in itself, surprising; production methods, recipes and raw materials vary themselves. Other factors probably contributing to the detected inter-producer variability include the manner in which they were stored and distributed (‘Alheiras’ are sold loose or packaged – vacuum or modified atmo-sphere) and indeed the time elapsed since production – not all samples were labelled with shelf life information. Indeed, as a largely artisanal production process which is not initiated by starter cultures, these same factors are likely to be responsible for the intra-producer variations. The variations between batches from the same producer are less pronounced which suggests some stability of the microecology although variations in batches of raw mate-rials, process parameters and plant hygiene are likely to contribute some variation.
When the results obtained from both traditional plat-ing and DGGE are analysed (Tables 1 and 2), it becomes evident that LAB constitute the predominant microbial population of ‘Alheira’. In DGGE gels, multiple bands were visible in most of the samples, and different phylo-types, most of which related to the order Lactobacillales, were identified. In a previous, culture-based study, four-teen species of LAB were isolated from ‘Alheira’ from MRS and M17 media, such us: Lact. paraplantarum, Lact. brevis, Lact. rhamnosus, Lact. sakei, Leuc. mesenteroides, Lact. zeae, Lact. paracasei, Ped. pentosaceus, Ped. acidilac-tici, Weissella cibaria, W. viridescens and Ent. faecium
(Albano et al. 2007). Albano et al. (2007) also observed that Lact. plantarum and members of the genus Entero-coccus were isolated from all samples of ‘Alheira’ analysed. Enterococcus-related phylotypes were identified also in PCR-DGGE sequences.
In addition to LAB, Micrococcaceae counts were >4Æ2 log CFU g)1in most samples. Intriguingly, although the separation of the V1 and V3 rRNA gene amplicons by PCR-DGGE has been described as a reliable tool to inves-tigate the LAB and CNC diversity in fermented sausages (Cocolin et al. 2001a,b), only one band was identified as being part of this latter microbial group (Macrococcus caseolyticus, Band 5) in the denaturing gels generated in this present study. This finding might be due to the selec-tive amplification of the most abundant templates, a bias previously recognized when performing PCR-DGGE anal-ysis of heterogeneous DNA template mixtures (Felske et al. 1998; Ampe et al. 1999; Ercolini et al. 2001b).
According to the guidelines for the microbiological quality of fermented meats published by Gilbert et al. (2000), most of the samples tested (80%) would be con-sidered unsatisfactory, with Enterobacteriaceae counts greater than 4 log CFU g)1. The detection of Listeria spp. in some samples points to the possibility of product con-tamination with the pathogenic specie L. monocytogenes. Sequences closely related to Enterobacteriaceae were retrieved from DGGE bands. Brochothrix-related phylo-types were also detected in almost all the samples, as were close relatives of Pseudomonas; in both cases suggesting the presence of potential spoilage strains. Quantitative approaches, such as those using real-time PCR would be needed in order to quantify the presence of specific potentially problematic bacteria. As shown by Rantsiou and Cocolin (2006), the combination of cultivation strate-gies and DNA-based techniques allows the identification of the dominant microbial species in fermented sausages, since it permits the generation and comparison of charac-teristic patterns from cultivated isolates and complex microbial communities. However, Ercolini (2004) described possible pitfalls of the whole procedure when applied to food products, like sampling, DNA extraction, DNA purity, PCR conditions, formation of heteroduplex and chimeric molecules, small fragments, between others.
PCR-DGGE applied to template DNA, directly extracted from a food matrix, generates a specific profile of that product in that moment, given the conditions used. DGGE analysis and sequencing of the resultant bands allows the identification of the dominant phylo-types present in all products. The approach explored in this study allowed the description of the microbial com-munity present in ‘Alheira’ in particular the diversity of LAB. DGGE could be used in monitoring the growth kinetics during the ripening process and the effects of
these on the hygienic quality, allowing tracking of the microbial ‘typicity’ of the ‘Alheira de Mirandela’, and col-lecting of important information for the designing of autochthonous starter cultures.
Acknowledgements
This work received financial support via project POCTI⁄ AGG⁄ 39587 ⁄ 2001, Fundac¸a˜o para a Cieˆncia e Tecnolo-gia (FCT)⁄ Fundo Europeu Desenvolvimento Regional (FEDER). Financial support for author H. Albano was pro-vided by PhD fellowship, SFRH⁄ BD ⁄ 13161 ⁄ 2003 (FCT) and for I. Henriques by PhD fellowship (SFRH⁄ BPD ⁄ 21384⁄ 2005).
References
Albano, H., van Reenen, C.A., Todorov, S.D., Hogg, T., Dicks, L.M.T. and Teixeira, P. (2007) Ecology of lactic acid bacte-ria in ‘‘Alheira’’, a fermented sausage traditionally pro-duced in Portugal: isolation, identification and molecular characterization. Personal communication in MICRO’07 – BIOTEC’07-XXXIII, Lisbon, Portugal, 30 November, 2007–2 December, 2007.
Ampe, F., ben Omar, N., Moizan, C., Wacher, C. and Guyot, J.-P. (1999) Polyphasic study of the spatial distribution of microorganisms in Mexican pozol, a fermented maize dough, demonstrates the need for cultivation independent methods to investigate traditional fermentations. Appl Environ Microbiol 65, 5464–5473.
Aquilanti, L., Santarelli, S., Silvestri, G., Osimani, A., Petruzzelli, A. and Clementi, F. (2007) The microbial ecology of a typical Italian salami during its natural fermentation. Int J Food Microbiol 120, 136–145.
van Beek, S. and Priest, F.G. (2002) Evolution of the lactic acid bacterial community during malt whisky fermentation: a polyphasic study. Appl Environ Microbiol 68, 297–305. Cocolin, L., Manzano, M., Cantoni, C. and Comi, G. (2000)
Development of a rapid method for the identification of Lactobacillus spp. isolated from naturally fermented Italian sausages using a polymerase chain reaction-temperature gradient gel electrophoresis. Lett Appl Microbiol 30, 126–129.
Cocolin, L., Manzano, M., Cantoni, C. and Comi, G. (2001a) Denaturing gradient gel electrophoresis analysis of the 16S rRNA gene V1 region to monitor dynamic changes in the bacterial population during fermentation of Italian sausages. Appl Environ Microbiol 67, 5113–5121. Cocolin, L., Manzano, M., Aggio, D., Cantoni, C. and Comi,
G. (2001b) A novel polymerase chain reaction (PCR) denaturing gradient gel electrophoresis (DGGE) for the identifcation of Micrococcaceae strains involved in meat fermentations. Its application to naturally fermented Italian sausages. Meat Sci 57, 59–64.
Cocolin, L., Aggio, D., Manzano, M., Cantoni, C. and Comi, G. (2002a) An application of PCR-DGGE analysis to profile the yeast populations in raw milk. Int Dairy J 12, 407–411. Cocolin, L., Rantsiou, K., Iacumin, L., Cantoni, C. and Comi, G. (2002b) Direct identification in food samples of Listeria spp. and Listeria monocytogenes by molecular methods. Appl Environ Microbiol 68, 6273–6282.
Cocolin, L., Rantsiou, K., Iacumin, L., Urso, R., Cantoni, C. and Comi, G. (2004) Study of the ecology of fresh sau-sages and characterization of populations of lactic acid bacteria by molecular methods. Appl Environ Microbiol 70, 1883–1894.
Coppola, S., Blaiotta, G., Ercolini, D. and Moschetti, G. (2001) Molecular evaluation of microbial diversity occurring in different types of Mozzarella cheese. J Appl Microbiol 90, 414–420.
Ercolini, D. (2004) PCR-DGGE fingerprinting: novel strategies for detection of microbes in food. J Microbiol Methods 56, 297–314.
Ercolini, D., Moschetti, G., Blaiotta, G. and Coppola, S. (2001a) Behavior of variable V3 region from 16S rDNA of lactic acid bacteria in denaturing gradient gel electrophore-sis. Curr Microbiol 42, 199–202.
Ercolini, D., Moschetti, G., Blaiotta, G. and Coppola, S. (2001b) The potential of a polyphasic PCR-DGGE approach in evaluating microbial diversity of natural whey cultures for water-buffalo mozzarella cheese production: bias of ‘‘culture dependent’’ and ‘‘culture independent’’ approaches. Syst Appl Microbiol 24, 610–617.
Ercolini, D., Hill, P.J. and Dodd, C.E.R. (2003) Bacterial com-munity structure and location in Stilton cheese. Appl Envi-ron Microbiol 69, 3540–3548.
Esteves, A., Patarata, L., Saraiva, C. and Martins, C. (2008) Assessment of the microbiological characteristics of indus-trially produced Alheira, with particular reference to food-born pathogens. J Food Safety 28, 88–102.
Felske, A., Akkermans, A.D. and de Vos, W.M. (1998) Quanti-fication of 16S rRNAs in complex bacterial communities by multiple competitive reverse transcription-PCR in tem-perature gradient gel electrophoresis fingerprints. Appl Environ Microbiol 64, 4581–4587.
Ferreira, V., Barbosa, J., Vendeiro, S., Mota, A., Silva, F., Monteiro, M.J., Hogg, T., Gibbs, P. et al. (2006) Chemical and microbiological characterization of alheira: a typical Portuguese fermented sausage with particular reference to factors relating to food safety. Meat Sci 73, 570–575. Fontana, C., Vignolo, G. and Cocconcelli, P.S. (2005) PCR–
DGGE analysis for the identification of microbial popula-tions from Argentinean dry fermented sausages. J Microbiol Methods 63, 254–263.
Gilbert, R.J., de Louvois, J., Donovan, T., Little, C., Nye, K., Ribeiro, C.D., Richards, J. and Roberts, D. (2000) Guide-lines for the microbiological quality of some ready-to-eat foods sampled at the point of sale. PHLS advisory
committee for food and dairy products. Communicable Dis Public Health 3, 163–167.
Head, I.M., Saunders, J.R. and Pickup, R.W. (1998) Microbial evolution, diversity and ecology: a decade of ribosomal RNA analysis of uncultivated microorganisms. Microbiol Ecol 35, 1–21.
Henriques, I.S., Almeida, A., Cunha, A. and Correia, A. (2004) Molecular sequence analysis of prokaryotic diversity in the middle and outer sections of the Portuguese estuary Ria de Aveiro. FEMS Microbiol Ecol 49, 269–279.
Henriques, I.S., Alves, A., Taca˜o, M., Almeida, A., Cunha, A. and Correia, A. (2006) Seasonal and spatial variability of free-living bacterial community composition along an estuarine gradient (Ria de Aveiro, Portugal). Estuar Coast Shelf Sci 68, 139–148.
Hugenholtz, P., Goebel, B.M. and Pace, N.R. (1998) Impact of culture-independent studies on emerging phylogenetic view of bacterial diversity. J Bacteriol 180, 4765–4774. IPQ (1991) Food Microbiology. General Guidance for the
Enu-meration of Enterobacteriacea Without Ressuscitation. MPN Technique and Colony Count Technique. Portuguese Stan-dard NP 4137.
ISO (2003) Microbioloy of Food and Animal Feeding Stuffs – Horizontal Method for the Enumeration of Microorganisms – Colony-Count Technique at 30!C. International Standard 4833:2003E.
Lu¨cke, F.K. (1974) Fermented sausages. In Microbiology of Fermented Foods ed. Wood, B.J.B. pp. 41–49 London: Applied Science Publishers.
Meroth, C.B., Walter, J., Hertel, C., Brandt, M.J. and Hammes, W.P. (2003) Monitoring the bacterial population dynamics
in sourdough fermentation process by using PCR-denatur-ing gradient gel electrophoresis. Appl Environ Microbiol 69, 475–482.
Muyzer, G., De Waal, E.D. and Uitterlinden, A.G. (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59, 695–700.
Ogier, J.C., Son, O., Gruss, A., Tailliez, P. and Delacroix-Buchet, A. (2002) Identification of the bacterial micro-flora in dairy products by temporal temperature gradient gel electrophoresis. Appl Environ Microbiol 68,
3691–3701.
ben Omar, N. and Ampe, F. (2000) Microbial community dynamics during production of the Mexican fermented maize dough pozol. Appl Environ Microbiol 66, 3664– 3673.
Randazzo, C.L., Torriani, S., Akkermans, A.D.L., de Vos, W.M. and Vaughan, E.E. (2002) Diversity, dynamics, and activity of bacterial communities during production of an artisanal Sicilian cheese as evaluated by 16S rRNA analysis. Appl Environ Microbiol 68, 1882–1892.
Rantsiou, K. and Cocolin, L. (2006) New developments in the study of the microbiota of naturally fermented sausages as determined by molecular methods: a review. Int J Food Microbiol 108, 255–267.
Rantsiou, K., Urso, R., Iacumin, L., Cantoni, C., Cattaneo, P., Comi, G. and Cocolin, L. (2005) Culture-dependent and -independent methods to investigate the microbial ecology of Italian fermented sausages. Appl Environ Microbiol 71, 1977–1986.