DEPARTAMENTO DE BIOLOGIA VEGETAL
On the effect of internal tidal waves on
near-surface chlorophyll concentration and productivity
around Nazaré Canyon and in the central region of
the Bay of Biscay
Sérgio Muacho Alves
DOUTORAMENTO EM CIÊNCIAS DO MAR
DEPARTAMENTO DE BIOLOGIA VEGETAL
On the effect of internal tidal waves on
near-surface chlorophyll concentration and productivity
around Nazaré Canyon and in the central region of
the Bay of Biscay
Sérgio Muacho Alves
Tese orientada por:
Prof. Doutora Vanda Costa Brotas Gonçalves Prof. Doutor José Carlos Pinto Bastos Teixeira da Silva,
especialmente elaborada para a obtenção do grau de Doutor em Ciências do Mar
Dissertação apresentada à
Universidade de Lisboa para obtenção
do grau de Doutor em Ciências do Mar
___________________________
Sérgio Muacho Alves
Declaração
Para efeitos do disposto nº2 do Art. 8º do Dec-Lei 388/70, o autor da
dissertação declara que interveio na conceção do trabalho experimental, na
interpretação dos resultados e na redação dos manuscritos publicados e
submetidos para publicação.
Sérgio Muacho Alves
dezembro de 2012
Este trabalho foi financiado através de uma Bolsa de Doutoramento
(SFRH/BD/22561/2005) concedida pela Fundação para a Ciência e a
Tecnologia.
com enorme satisfação que noto que não foram poucas as pessoas que, direta ou indiretamente, contribuíram para este documento final. Esses contributos tanto foram de índole cientifica, como de companheirismo, amizade e amor.
Agradeço à professora Vanda toda a confiança que confiou na minha pessoa, ao importante suporte que me facultou em todos os temas da área da Biologia e à amizade que tive a oportunidade de registar por diversas vezes. É sempre um enorme estímulo trabalhar com alguém que tem uma mentalidade empreendedora.
De igual forma, agradeço ao professor José da Silva o apoio que em mim depositou. Os seus conselhos e incentivos, de alguém que se rege por um exímio rigor científico, foram decisivos para a minha formação profissional. Apesar de já o conhecer há uma década, foi com enorme prazer que registei a sua elevada dedicação ao assunto aqui trazido, em particular na reta final, onde excedeu as minhas expectativas iniciais.
Ao Paulo Oliveira, pela sua objetividade, pragmatismo e competências nesta abrangente área que é a Oceanografia, particularmente úteis em determinadas etapas deste trabalho.
Ao incansável Jorge Magalhães pelo forte apoio e pela pronta disponibilidade que sempre mostrou na elaboração de algumas figuras aqui apresentadas.
Agradeço à minha namorada, Maria da Luz Calado, por todo o apoio, incentivo, dedicação e entrega total na fase final da tese. O trabalho aqui exposto é igualmente o resultado de todo esse empenho, amor e sacrifício familiar. Mais, todos os momentos vividos em conjunto constituíram uma fonte de inspiração importantíssima à minha felicidade diária (através da sua presença, da sua simplicidade e do seu constante sorriso) e, consequentemente, a este trabalho.
Aos meus amigos João Rego e André Valente, cujos conhecimentos em Oceanografia Física permitiram trazer a este trabalho significativas mais-valias. Todas as trocas de ideias (e não só) ao longo destes últimos anos constituíram um importante tónico para mim.
Agradeço igualmente a preciosa colaboração e amizade dos meus ex-colegas Carolina Sá (pela sua simpatia e prazer pelo trabalho em equipa), Ana Sousa (pelo espírito positivo e pelos constantes incentivos) e Carlos Rafael bem como do David Ávila, meu grande amigo de longa data.
Por fim, quero deixar uma palavra de muito apreço aos meus pais e restantes amigos.
RESUMO 15
Referências 20
CAPÍTULO I – Introdução 21
1.1. Enquadramento teórico 22
1.1.1. Ondas e marés internas 22
1.1.2. Ondas internas solitárias e sua teledeteção 24 1.1.3. Fitoplâncton, produção primária e suas estimativas 27
1.2. Estado da arte 31
1.2.1. Ondas internas na margem Oeste Ibérica 31 1.2.2. Ondas internas no Golfo da Biscaia 35 1.2.3. Efeito de ondas internas no fitoplâncton e na produção primária 39
1.3. Objetivos 42
1.4. Estrutura da tese 43
Referências 44
CAPÍTULO II – Effect of Internal Waves on near-surface chlorophyll concentration and primary production in the Nazaré Canyon (West of
Iberian Peninsula) 49
Abstract 50
2.1. Introduction 51
2.2. Data and methods 55
2.2.1. In-situ data 55
2.2.2. Remote-sensing data 56
2.2.3. Primary production model 57
2.3. Results 58
2.3.1. IW effects on chlorophyll vertical distribution and detection in ocean
colour remote sensing 58
2.3.2. Multi-sensor analysis 61
2.3.3. Primary production model results 65
2.4. Discussion 67
2.5. Conclusions 71
Acknowledgements 71
Abstract 77
3.1. Introduction 78
3.2. SAR analysis 81
3.3. Ocean colour observations 87
3.4. Comparison with model results 91
3.5. Discussion 94 3.6. Conclusions 98 Acknowledgements 98 References 99 CAPÍTULO IV – Conclusões 101 4.1. Conclusões 102 Referências 106
13
ABSTRACT
The present study describes the effect of large-amplitude internal tidal waves on near-surface chlorophyll concentration in the central region of Bay of Biscay and around the Nazaré Canyon (western Portugal), and investigates its ecological impact.
Internal wave propagation introduces vertical motions within the water column, especially near the thermocline, forcing water particles to undergo upward and downward motions. Since neutrally buoyant phytoplankton cells are usually passive, these cells can be significantly displaced in the vertical. Because light intensity decreases exponentially with depth, internal waves will expose phytoplankton cells to a larger light intensity on average.
The main objective of this study was to investigate if this light enhancement in the euphotic zone can induce a biomass increase and affect positively primary production in the two regions of interest.
SAR data was used to characterize the signatures of internal solitary waves in the central Bay of Biscay, which are locally generated by internal tidal beams, which first propagate downwards from the shelf break and reflect from the sea floor. Two similar episodes of internal wave propagation in the Bay of Biscay were investigated and it was shown the existence of two different axes of wave beam propagation, which result in internal solitary wave generation and propagation in two distinct directions.
SAR and Ocean Colour satellite synergy support the occurrence of enhanced levels of chlorophyll spatially correlated with internal solitary waves observed around the Nazaré Canyon (generated by the barotropic tides at the Estremadura Promontory). In addition, the results from the central Bay of Biscay (near 46° N, 7° W) show evidence of the confluence of internal tidal waves, which produce chlorophyll patterns visible in ocean colour images.
14
The results derived from satellite climatological chlorophyll concentrations (2002-2011) and from a productivity model, suggest that high internal wave activity is an important mechanism playing a significant role on biomass growth, and thus have a relevant regional ecological impact.
Key-words: Internal tidal waves; Internal solitary waves; Chlorophyll concentration; Nazaré Canyon (Portugal); Central Bay of Biscay (46° N, 7° W); SAR and Ocean Colour satellite synergy; Primary production.
15
RESUMO
O presente estudo incidiu sobre as ondas internas e, mais concretamente, caracterizou as ondas solitárias e a maré interna na região central do Golfo da Biscaia e na região do Canhão submarino da Nazaré e, avaliou o efeito das ondas de maré interna de grande amplitude em padrões superficiais de clorofila e o eventual impacto destas ondas na ecologia das duas regiões.
As ondas internas são ondas gravíticas que se propagam no interior de um fluido estratificado, ocorrendo no oceano e na atmosfera, e que correspondem a modos de oscilação baroclínicos.
No oceano estas ondas são ubíquas, surgindo sempre que existam combinações adequadas entre o gradiente vertical de densidade, o fluxo periódico de corrente de maré e a variação da topografia, sendo possível observar uma diversidade de ondas internas com diferentes características (amplitudes, comprimentos de onda, períodos). Estas ondas imprimem oscilações verticais na coluna de água, em particular na região termoclina (de origem sazonal a médias latitudes), podendo propagar-se por centenas de quilómetros e envolvendo o transporte de energia e de momento linear, conferindo ao interior do oceano uma dinâmica muito energética.
As ondas internas podem influenciar, de diferentes formas, determinados processos biológicos tal como a produtividade do fitoplâncton (Kahru, 1983; Holloway, 1984; Lande & Yentsch, 1988; Holloway & Denman, 1989; Wang et al., 2007; Evans et al., 2008; Pan et al., 2012).
O fitoplâncton, que se encontra na base de complexas cadeias tróficas, assume-se como o principal produtor primário dos oceanos, assume-sendo responsável por aproximadamente metade da produção primária do globo terrestre, apesar de representar menos de 1% da biomassa vegetal total. O fitoplâncton inclui os
16
microrganismos aquáticos fotoautotróficos, que se encontram nas camadas mais superficiais do oceano (i.e., na zona eufótica), onde se conjugam as condições ideais de disponibilidade luminosa e de nutrientes, determinantes para o processo fotossintético. Nesse sentido, a análise da variabilidade do fitoplâncton constitui uma ferramenta determinante no aprofundamento do conhecimento das relações tróficas e, inclusivamente, no papel ecológico do fitoplâncton na regulação de níveis de dióxido de carbono nas camadas superficiais do oceano, assim como na atmosfera.
Os movimentos verticais, ascendentes e descendentes, inerentes à atividade interna poderão favorecer a produtividade do fitoplâncton, na medida em que induzem um maior fluxo de nutrientes para a zona eufótica e expõem as células fitoplâncton, usualmente passivas, a uma maior intensidade luminosa, uma vez que esta se atenua exponencialmente com a profundidade.
Apesar de os efeitos que as ondas internas têm na produtividade primária terem sido documentados em torno do Atol de Dongsha na região norte do Mar do Sul da China (Wang et al., 2007; Pan et al., 2012), desconhece-se se este processo ocorre igualmente noutras regiões oceânicas com elevada dinâmica interna, mas com características diferentes de um Atol.
Nesse sentido, o principal objetivo deste trabalho foi investigar se a presença de ondas internas de grande amplitude (dezenas de metros) pode representar um acréscimo da biomassa de fitoplâncton e afetar positivamente a produção primária na região central do Golfo da Biscaia e do Canhão da Nazaré.
As ondas internas das regiões de estudo e especificamente, as ondas internas de maré, correspondem a oscilações internas de período semidiurno, geradas pelo fluxo barotrópico de maré sobre uma topografia irregular.
17
Na região do Canhão da Nazaré, investigações baseadas em observações in-situ e remotas (em particular, as obtidas pelo SAR) têm revelado uma complexa atividade interna em condições de estratificação (particularmente nos meses de verão), em resultado da presença de marés internas e de ondas internas solitárias de grande amplitude (Quaresma et al., 2007).
Neste trabalho, as marés internas observadas in-situ na região do Canhão da Nazaré apresentaram flutuações de grande amplitude da termoclina sazonal, atingindo 25 m. Adicionalmente, a análise de imagens SAR revelou ondas internas solitárias numa região do Canhão da Nazaré externa à plataforma continental, com uma direção de propagação para este-nordeste, tendo provavelmente emanado do Promontório da Estremadura.
Na região central do Golfo da Biscaia, as ondas internas manifestam-se como oscilações mais intensas comparativamente à região do Canhão da Nazaré, atingindo mais de 30 m (e.g., Pingree & New, 1995). Estas ondas são “geradas localmente” por raios de maré interna que emanam de diversos locais da vertente continental na região norte do Golfo, que depois de refletidos no fundo oceânico, atingem a termoclina, a cerca de 150 km do bordo da plataforma (e.g., New & Pingree, 1992).
Neste estudo, a análise de imagens SAR permitiu caraterizar ondas internas solitárias resultantes deste processo de geração, e revelar dois eixos de propagação que se intercetam na região central do Golfo da Biscaia. Esta região, centrada em torno de 46° N, 7° W, corresponde ao local onde convergem as ondas internas “geradas localmente”, constituindo portanto um local com uma significativa energética interna.
A avaliação dos efeitos das ondas internas em padrões superficiais de clorofila (usada como índice de biomassa de fitoplâncton), foi efetuada recorrendo a uma abordagem sinergética de dados de satélite - SAR e MODIS - uma vez que a ligação
18
entre a atividade destas ondas e os consequentes deslocamentos verticais de fitoplâncton encontra-se amplamente documentado (e.g., Sangra et al., 2001; Gaxiola-Castro et al., 2002).
As imagens MODIS relativas à região do Canhão da Nazaré evidenciaram uma estrutura bidimensional nos padrões superficiais de clorofila em concordância com a propagação da maré interna (estimada a partir das posições das ondas internas solitárias observadas no SAR), apresentando maiores concentrações na crista de onda.
Na região central do Golfo da Biscaia, as concentrações mais elevadas de clorofila, que se apresentam sob a forma de padrões quase-circulares, foram registadas no local onde ocorre a confluência de marés internas (perto de 46° N, 7° W), reforçadas pela presumível interferência positiva das cristas de onda.
Neste estudo pretendeu-se igualmente avaliar o impacto ecológico da elevada atividade de ondas internas registada nas duas regiões de interesse e, especificamente analisar se os fortes movimentos verticais inerentes à presença de ondas de grande amplitude nestas regiões, produzem, de facto, um aumento no crescimento do fitoplâncton, afetando positivamente a produtividade primária.
A determinação da produção primária, através de um modelo numérico, numa região no Canhão da Nazaré externa à plataforma continental, evidenciou um aumento das taxas de fotossíntese associado à presença de atividade interna, quando comparado com uma situação de inexistência de movimento ondulatório.
Na região central do Golfo da Biscaia, a análise dos dados da climatologia da concentração de clorofila obtida por satélite (no período entre 2002 e 2011), relativa ao mês de Setembro, revelou que a região centrada em torno de 46.0° N, 7.0° W apresenta uma anomalia positiva de aproximadamente de 10% na biomassa de fitoplâncton quando comparada com as regiões adjacentes, menos afetadas pela presença destas
19
ondas. Este efeito poderá resultar da conjugação das inúmeras interações entre ondas internas, provenientes de diferentes locais de geração sobre o bordo da plataforma continental, com os presumíveis efeitos da reemergência “local” que conduzem a uma intensificação dos movimentos verticais.
Os resultados obtidos através de um modelo de produtividade primária e de dados climatológicos de níveis de clorofila, confirmam portanto o impacto ecológico significativo que uma elevada atividade de ondas internas pode ter em determinados processos biológicos, tal como um aumento da biomassa de fitoplâncton e da produção primária, particularmente em regiões oceânicas onde as ondas internas dominem os processos físicos.
Palavras-chave: Ondas internas de maré; Ondas internas solitárias; Concentração de clorofila; Canhão da Nazaré (Portugal); Centro do Golfo da Biscaia (46° N, 7° W); Sinergia SAR e cor oceânica; Produção primária.
20
Referências
Evans MA, MacIntyre S, Kling GW (2008). Internal wave effects on photosynthesis: Experiments, theory, and modeling. Limnol. Oceanogr., 53, 339–353.
Gaxiola-Castro G, Alvarez-Borrego S, Najera-Martinez S, Zirino AR (2002). Internal waves effect on the Gulf of California phytoplankton. Cienc. Mar., 28: 297–309.
Holloway G (1984). Effects of velocity fluctuations on vertical distributions of phytoplankton. J. Mar. Res. 42: 559–571
Holloway G, Denman K (1989). Influence of internal waves on primary production, Journal of Plankton Research, 11, 409–413.
Kahru M (1983). Phytoplankton patchiness generated by long internal waves: a model, Marine Ecology - Progress Series, 10, 111-117.
Lande R, Yentsch CS (1988). Internal waves, primary production and the compensation depth of marine phytoplankton, Journal of Phytoplankton Research, 10, 565-571.
New AL, Pingree RD (1992). Local generation of internal soliton packets in the central Bay of Biscay. Deep-Sea Res. Part A 39, 1521-1534. doi:10.1016/0198- 0149(92)90045-U.
Pan X-J, Wong GTF, Shiah F-K, Ho T-Y (2012). Enhancement of biological productivity by internal waves: observations in the summertime in the northern South China Sea. Journal of Oceanography doi:10.1007/s10872-012-0107-y.
Pingree RD, New AL (1995). Structure, seasonaldevel opment and sunglint spatial coherence of the internal tide on the Celtic and Armorican shelves and in the Bay of Biscay. Deep-Sea Research Part I 42, 245–284.
Quaresma LS, Vitorino J, Oliveira A, da Silva J (2007). Evidence of sediment resuspension by nonlinear internal waves on the western Portuguese mid-shelf. Marine Geology 246, 123–143.
Sangra P, Basterretxea G, Pelegri JL, Aristegui J (2001). Chlorophyll increase due to internal waves on the shelf break of Gran Canaria (Canary Islands), Sci. Mar., 65(Suppl. 1): 89-97.
Wang YH, Dai CF, Chen YY (2007). Physical and ecological processes of internal waves on an isolated reef ecosystem in the South China Sea. Geophys Res Lett 34:L18609. doi:10.1029/2007GL030658
21
CAPÍTULO I
__________________________________________________
22
1.1 Enquadramento teórico
1.1.1 Ondas e marés internas
As ondas internas (OI) são ondas gravíticas que se propagam no interior de um fluido. Constituem modos de oscilação baroclínicos num fluido estratificado, sendo a força restauradora proporcionada pela estratificação vertical da densidade e a força da gravidade. As OI surgem então sempre que existam combinações adequadas entre o gradiente vertical de densidade, o fluxo de corrente periódica (de maré) e a variação da topografia, que tanto ocorrem no oceano como na atmosfera.
A frequência de oscilação (ω) das OI varia entre um máximo correspondente à frequência de Brunt-Väisälä e um mínimo correspondente à frequência de Coriolis. A frequência de Brunt-Väisälä ou frequência de impulsão, , é dada por
(z)
-em que representa a aceleração da gravidade, a densidade e a variação vertical da densidade); a frequência de Coriolis ou frequência inercial (também descrita como frequência natural de oscilação), é dada por , em que representa a velocidade angular da Terra em torno do seu eixo de rotação e a latitude do local. As OI situar-se-ão, portanto, em zonas onde seja satisfeita a condição <
ω
< , em particular na termoclina sazonal.No oceano, é possível observar uma multiplicidade de OI com diferentes frequências, números de onda e amplitudes. As OI dispõem-se em escalas espácio-temporais amplamente distintas apresentando períodos que variam entre minutos a horas, distâncias (comprimento de onda) que variam entre metros e dezenas de quilómetros, e amplitudes que podem ir de dezenas de centímetros até dezenas de metros.
23
Desta forma, coexistem no oceano oscilações internas que incluem as ondas inerciais, marés internas e OI não lineares de curto período (Huthnance, 1995). Estas perturbações na densidade podem propagar-se por longas distâncias (centenas de quilómetros), envolvendo transporte de energia, massa e de momento linear que conduz frequentemente à turbulência e a fenómenos de mistura (Staquet & Sommeria, 2002). A variedade de mecanismos de geração e as inumeráveis regiões oceânicas onde estes processos dominam, conferem ao interior do oceano uma dinâmica muito energética (Kantha & Tierney, 1997).
A maré interna (MI) pode ser definida como a parte do espectro das OI com frequências de maré. Corresponde a uma oscilação interna de período diurno ou semidiurno, sendo gerada pelo fluxo barotrópico de maré sobre uma topografia irregular (e.g. montanhas submarinas ou declives continentais). Estas oscilações resultam numa dissipação de 30% da energia da maré barotrópica global (e.g., Gerkema & Zimmerman, 2008). Ao deslocarem-se sobre fortes acidentes topográficos (e.g., a inclinada vertente continental), as correntes de maré vão gerar fortes movimentos verticais, particularmente através da respetiva componente barotrópica. As ondas assim geradas vão irradiar da zona de formação e podem deslocar-se ao longo da termoclina (no “plano” horizontal). Como a maré semidiurna (T=12.42h) é super-inercial (ω > f) na maioria do oceano, então a MI semidiurna pode propagar-se por quase todo o oceano, desde que existam condições de estratificação (Jeans, 2002).
Na presença de estratificação contínua, a energia das OI pode também transmitir-se ao longo de raios, correspondentes à soma de um grande número de modos de OI (ver e.g., Gerkema & Zimmerman, 2008). Estes raios apresentam uma inclinação em relação à horizontal, determinada pela expressão:
24
Habitualmente, designa-se a zona de geração como ponto “crítico” da vertente continental, que é o local onde o declive do fundo iguala o ângulo que o raio apresenta em relação à horizontal. Em seguida, a energia das OI propaga-se ao longo dos raios, com declives positivos e negativos, tanto na direção da costa como para o largo, sendo que estes se refletem no fundo oceânico e induzem oscilações (e.g., com o período da MI semidiurna) ao intersetarem a termoclina.
1.1.2 Ondas internas solitárias e sua teledeteção
Designam-se por solitões as ondas solitárias de amplitude finita, forma constante e que apresentam uma velocidade de fase que é função crescente da amplitude. No oceano, os solitões, também designados por ondas internas solitárias (OIS), podem ser observados isoladamente ou em grupo, usualmente ordenadas hierarquicamente por amplitude, velocidade de fase e comprimento de onda (Apel, 2003).
A primeira observação de uma onda solitária reporta-se a 1834, quando Scott Russell presenciou a propagação de uma perturbação elevatória solitária na superfície da água, no canal de Edimburgo (Escócia). A justificação teórica para este fenómeno foi apresentada em 1895, por Kortweg e de Vries, através de um modelo unidimensional e não-linear (hoje conhecida por equação de KdV), uma vez que a teoria linear das OI não explica a ocorrência destas ondas. Basicamente, a equação KdV determina a evolução de uma perturbação ondulatória inicial, num dado ponto, para um grupo de solitões a propagar-se e representa o princípio da conservação da amplitude da onda solitária, como o resultado do balanço entre a declividade não-linear das ondas e a sua tendência para se dispersar (Benney, 1966).
25
As OIS são frequentemente descritas como flutuações da frente de onda da MI, gerada pelo fluxo de maré sobre bancos, montes e vertentes submarinas (Gerkema et al., 1995), que se propagam em condições de estratificação. Estas assumem-se pela propagação de flutuações de curto período (alguns minutos), na região da termoclina, estando associadas às cavas das ondas de MI, propagando-se com estas à mesma velocidade (e.g., New & Pingree 1990). A geração de OIS depende claramente da intensidade local das marés barotrópicas sendo também condicionada pelos ciclos sazonais da estratificação da coluna de água. A existência de uma maior atividade de OIS no período próximo das marés-vivas está bem documentada em determinadas regiões como o Golfo da Biscaia (e.g., Pingree et al., 1986) ou a Crista das Mascarenhas (da Silva et al., 2011), embora noutras, como o Promontório da Estremadura (Magalhaes & da Silva, 2012), tal não se verifique.
Nas latitudes médias, o gradiente de temperatura na camada superior do oceano (20 – 30 m), intensifica-se durante o verão, dando origem a uma termoclina pronunciada, ao longo da qual as OI se podem propagar no “plano” horizontal. Apesar das oscilações apresentarem, em geral, amplitudes superiores a 10 m na termoclina, as OI produzem elevações muito pequenas (inferior a 10 cm) na superfície do oceano. Este processo resulta na geração de gradientes na corrente à superfície que alteram as características das ondas de capilaridade (geradas por ação do vento local, com apenas alguns mm-dm de comprimento), resultando numa assinatura superficial, sob a forma de rugosidade (resultantes da interação das regiões de convergência e de divergência com as ondas de capilaridade). Estes padrões de rugosidade produzidos na superfície do mar pelas OIS são responsáveis por torná-las visíveis para diversos sensores remotos, como os radares SAR (Synthetic Aperture Radar) e os sensores óticos de resolução elevada e moderada (e.g., MEdium Resolution Imaging Spectrometer (MERIS) ou o Moderate Resolution Imaging Spectroradiometer (MODIS)).
26
A deteção remota por satélite tem-se revelado como a principal ferramenta na observação de OI no oceano. As imagens obtidas a partir de sensores a bordo de satélites têm contribuído grandemente para o conhecimento desta fenomenologia, uma vez que fornecem informações que de outra forma seriam inacessíveis. Estas incluem um quadro geral da distribuição espacial das OI (e.g., a sua orientação e direção de propagação), permitindo a determinação de distâncias entre ondas individuais ou grupos de ondas.
Os sensores óticos de resolução moderada constituem uma abordagem importante para deteção de OIS pois permitem obter a estrutura bidimensional das OIS à escala sinóptica, sendo que a sua identificação surge a partir do “sunglint” (o padrão de refletância quase especular da luz solar na superfície do mar). Apesar de Jackson (2007) ter revelado que as imagens MODIS podem ser utilizadas para analisar OIS numa escala quase global, na medida em que combina uma elevada resolução espacial (250 m em dados de alta resolução) com o varrimento de vastas áreas com uma frequência diária, o SAR (com uma resolução de 25 m) continua a ser o instrumento de excelência para a observação em alta resolução de OIS de período curto.
O SAR é um sensor ativo (emite radiação eletromagnética) que opera na frequência das micro-ondas, em comprimentos de onda variam entre o milímetro e o decímetro. A vantagem deste sensor relativamente aos sensores óticos, prende-se com o facto de este emitir feixes que atravessam a atmosfera sem serem afetados pela sua estrutura ou composição. Os SAR podem assim observar OIS em condições de grande nebulosidade e durante a noite. Adicionalmente, este sensor permite detetar pequenas variações na rugosidade da superfície do mar mais facilmente que os sensores que operam na parte visível do espectro, que estão dependentes da sua inclinação relativamente ao sol para a identificação de OIS (da Silva et al., 2003).
27
Nas imagens SAR, a amplificação da rugosidade à superfície nas zonas de convergência e a atenuação da amplitude em zonas de divergência resultam na impressão de bandas claras (zonas de convergência) e escuras (zonas de divergência) (Fig.1). Consequentemente, a identificação de OIS pelo SAR é geralmente feita pela presença de extensas bandas coerentes ao longo das cavas/cristas de onda, cuja intensidade contrasta com o fundo da imagem (ver Fig. 4).
Figura 1 – Esquema que ilustra um grupo de OIS (depressionários), ordenados com amplitudes decrescentes, em que (a) é a configuração da picnoclina determinada pelas OIS; (b) padrão de rugosidade na superfície do mar induzido pelas OIS e (c) a respetiva intensidade no SAR (http://www.ifm.uni-hamburg.de/ers-sar/).
1.1.3 Fitoplâncton, produção primária e as suas estimativas
O oceano cobre 70% da superfície terrestre, apresentando uma profundidade média de 4000 m. Este vasto ambiente marinho é colonizado por inúmeras comunidades biológicas interligadas em complexas cadeias tróficas. Na base dessas cadeias tróficas encontra-se o fitoplâncton, que apesar de representar menos de 1% da
28
biomassa vegetal do planeta, é responsável por cerca de metade da produção primária na Terra (Field, 1998), assumindo-se como principal produtor primário dos oceanos.
O fitoplâncton agrega os microrganismos autotróficos aquáticos (microalgas e cianobactérias) com capacidade fotossintética, encontrando-se nas camadas mais superficiais do oceano (i.e., na zona eufótica, que se estende desde a superfície até à profundidade em que a intensidade luminosa representa apenas 1% da existente à superfície). Nas latitudes médias e, especificamente durante os meses de verão (i.e., em condições de estratificação), a clorofila a (o pigmento fotossintético que se encontra em todas as classe taxonómicas do fitoplâncton) concentra-se maioritariamente numa zona específica da coluna de água, onde se equilibram as condições ideais de luminosidade e de disponibilidade de nutrientes que favorecem o processo fotossintético (Fig. 2), sendo que a luz é o fator que mais o condiciona (Kirk, 1994). Esse máximo de clorofila a em profundidade (Deep Chlorophyll Maximum (DCM)) é gerado quando os níveis de nutrientes na camada mais superficial diminuem após os “blooms” da primavera, deixando um máximo sub-superficial de clorofila a perto da termoclina.
Os microrganismos fotoautotróficos que compõem o fitoplâncton, têm a capacidade de fixar o carbono, utilizando a luz como fonte energética, que fica acumulado na biomassa, e consequentemente, disponível para níveis tróficos superiores (Lindman, 1942). A radiação utilizada pelos microrganismos fotossintéticos - Photosynthetic Available Radiation (PAR) – corresponde ao intervalo de luz entre os 400 e os 700 nm (i.e., radiação visível) e representa cerca de 46% da radiação total solar (Gates, 1980). A taxa de fotossíntese representa, portanto, uma medida real da produtividade de um ecossistema marinho.
29 Figura 2 - Esquema que ilustra um exemplo da estrutura vertical da clorofila a, fotossíntese, temperatura, nitratos e da atenuação da luz em profundidade, em águas oceânicas. Figura adaptada de Vaulot D (2006).
Um dos modelos mais utilizados para estimar a produtividade primária, baseia-se nas relações empíricas entre a fotossíntebaseia-se e os perfis verticais de clorofila a e PAR. Neste método, as taxas de fotossíntese (PP) podem ser determinadas, a cada nível de profundidade z, usando a seguinte relação (Platt et al., 1980):
PP (z) = B (z).Pbmax [1 – exp (-α.E(z)/ Pbmax], em que PP(z) é a produtividade à
profundidade z (em mg C.m-3), B é a biomassa (dada pela concentração de clorofila a
(chl em mg.m-3)), Pbmax é a capacidade de assimilação de carbono pelo fitoplâncton (em
mg C.(mg chl)-1), α representa a inclinação inicial da curva P-E (em mg C.(mg chl) -1.(W.m-2)-1) e E (z) corresponde a PAR à profundidade z (em W.m-2); os valores dos
parâmetros α e Pbmax são medidos experimentalmente através do método do
carbono-14. Esta formulação, que não admite a fotoinibição, traduz uma correlação direta entre a produção primária e a concentração de clorofila a.
30
A variabilidade das concentrações superficiais de clorofila a (usada como índice de biomassa de fitoplâncton), pode ser observada através de dados de satélite obtidos a partir de sensores de cor do oceano (i.e., que usam a informação na gama visível do espectro). A clorofila a apresenta um pico de absorção bastante significativo e amplo na região azul do espectro eletromagnético (centrado nos 440 nm) e outro perto dos 675 nm, mas não no verde (região do espectro onde reflete mais). Consequentemente, um aumento na concentração de clorofila a fará com que a água pareça mais verde. Isto explica por que razão os algoritmos usados na estimativa da concentração de clorofila a no oceano tenham como base o rácio entre as radiâncias ou refletâncias verde/azul (presentemente, a maioria dos algoritmos são um pouco mais complexos do que este rácio, mas ainda estão intimamente relacionados com ele (Robinson, 2010)).
Uma vez que a distribuição de clorofila a varia consideradamente em profundidade (Fig.2), torna-se imprescindível aferir de que forma os sensores de cor “veem” a estrutura vertical da clorofila a. Gordon & McCluney (1975) definiram z90 como a
profundidade de penetração da luz acima do qual é originada 90% da irradiância refletida de forma difusa (excluindo a refletância especular à superfície), tendo mostrado que z90 ~ K-1 em que K é o coeficiente de atenuação difusa para a irradiância
descendente. Esta profundidade, z90, pode igualmente ser considerada como a
profundidade até onde o sensor consegue “ver”. Assim, a concentração de clorofila a medida a partir de um sensor de cor é ponderada por um fator (g(z)=exp{-2.K.z}) que conta com a atenuação exponencial da luz na água. Consequentemente, será dado um “peso” cada vez menor (exponencialmente menor) às concentrações de clorofila a à medida que a profundidade aumenta até atingir z90.
31
1.2 Estado da arte
1.2.1 Ondas internas na margem Oeste Ibérica
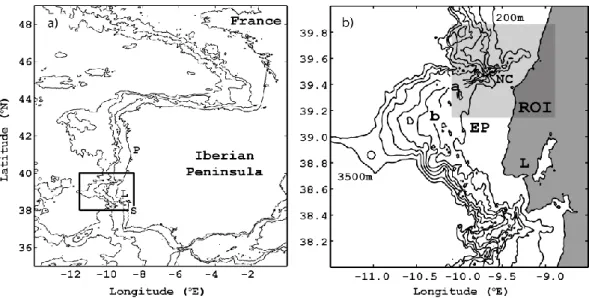
A margem Oeste Ibérica estende-se desde o Golfo de Cádis (identificado como GC na Fig. 3) até ao Banco da Galiza (identificado como GB na Fig. 3) localizado a noroeste da Península Ibérica, e apresenta uma plataforma continental com uma largura média de 45 km.
Existem várias referências que identificam a propagação de ondas de MI e de OIS ao longo da plataforma continental da margem Oeste da Península Ibérica (e.g., Alpers, 1985; da Silva et al., 1998; Jeans & Sherwin, 2001a, b; Small, 2002; Quaresma et al., 2007; da Silva et al., 2007; Magalhaes & da Silva, 2012). Estas investigações, baseadas em imagens SAR ou em observações in-situ, têm revelado uma dinâmica interna bastante significativa (pela presença de MI e de OIS de grande amplitude), particularmente nos meses de verão, quando existem condições de estratificação.
Figura 3 – Mapa da margem Oeste Ibérica relativamente à Península Ibérica. Os contornos identificam isóbatas em metros (0, 200, 1000, 2000, 3000 e 4000 m). As siglas GB, NC, EP e GC identificam, respetivamente, o Banco da Galiza, o Canhão da Nazaré, o Promontório da Estremadura e o Golfo de Cádis.
32
A campanha oceanográfica inserida no projeto MORENA (EU MAST II Multidisciplinary Oceanographic Research in the Eastern boundary of the North Atlantic), desenvolvida em agosto de 1994 na região da plataforma continental ao largo do Porto, constituiu um passo determinante para o conhecimento desta fenomenologia (nomeadamente os processos de mistura no bordo da plataforma) no litoral norte de Portugal. Os resultados desta campanha observacional revelaram MI de grande amplitude (com deslocamentos da termoclina de mais de 45 m) a propagarem-se em direção à plataforma (junto ao paralelo 41° N), sugerindo uma geração de OIS na vertente da plataforma continental por correntes de maré semidiurna (Jeans & Sherwin, 2001a). Após a sua geração, estas ondas propagam-se em direção à costa, com velocidades de fase na ordem de 0.6 m.s-1 (valores similares aos descritos noutros
trabalhos posteriormente conduzidos ao longo da plataforma continental Oeste da Península Ibérica).
A presença de MI e de OIS de grande amplitude encontra-se igualmente reportada na região centro-sul da margem Oeste Ibérica. Small (2002), através de imagens SAR e de observações in-situ, mediu OI de grande amplitude (40 m) a propagarem-se em zonas em que a profundidade é inferior a 100 m ao largo do Cabo de Sines e, mais recentemente, da Silva et al. (2007) identificaram OIS no extremo sul do Promontório da Estremadura (identificado como EP na Fig. 3), região de onde emanam estas ondas, propagando-se posteriormente para sul, num processo consistente com o mecanismo de “geração local” (explicado em detalhe na secção 1.2.2).
O Canhão submarino da Nazaré é um dos maiores do mundo e encontra-se localizado na costa Oeste Portuguesa (identificado como NC na Fig. 3). O Canhão apresenta o seu eixo com uma direção quase perpendicular à costa, estendendo-se por cerca de 270 km (Vanney & Mougenot, 1990), onde desempenha um papel preponderante na circulação regional (Vitorino et al., 2005). A análise de imagens SAR
33
permitiu a Quaresma et al. (2007) identificarem a geração de OIS sobre a cabeceira do canhão (39º35’N/009º25’W) e a sua subsequente propagação em direção à plataforma média, tendo igualmente revelado, através de observações in-situ, a importância destas grandes OIS na dinâmica sedimentar sobre a plataforma continental.
Quaresma (2006) documentou amplamente a propagação de OIS na plataforma continental média da região do Canhão da Nazaré. Ficou clara a multiplicidade de rumos de propagação de OIS, sustentando a hipótese da existência de diferentes locais de geração ao longo da vertente continental, e que se traduz numa complexa atividade de OIS nesta região. A análise de imagens SAR revelou que os processos de refração condicionam a propagação destas ondas, orientando-as perpendicularmente à batimetria. Este mecanismo origina uma constante interceção entre OIS, provenientes de diferentes locais da vertente continental, que se encontram esta região (as imagens SAR apresentadas na Fig. 4 ilustram este fenómeno de forma exemplar). Essa interceção de OIS origina interação não linear entre os solitões, podendo amplificar as ondas, e por isso, proporcionar eventos de mistura localizada (ver e.g. o recente estudo conduzido por Wang & Pawlowicz, 2012).
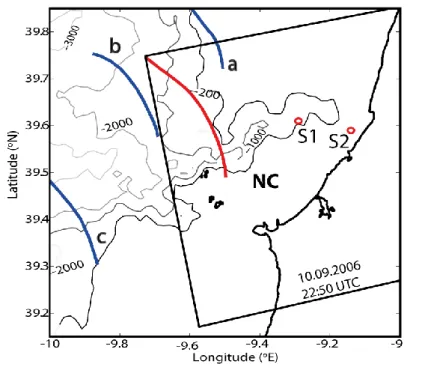
Magalhaes & da Silva (2012) basearam-se num amplo conjunto de imagens SAR para identificar o Promontório da Estremadura como um local de geração de OIS. O seu trabalho revelou a geração de OIS nas encostas norte e sul do Promontório, apresentando duas direções principais de propagação (sudeste e nordeste), sendo, que a componente oriental da sua direção de propagação intensifica-se à medida que as ondas se aproximam de costa. Esta última interpretação pode ser constatada com o mapa de observações de OIS (obtidas a partir de imagens SAR) representado na Fig. 5, que apresenta grupos de ondas, geradas no Promontório da Estremadura, e que se propagam para nordeste em direção à plataforma continental.
34 Figura 4 - Composição de imagens SAR, obtidas pelo satélite ERS2 às 22h55m UTC em 17 de agosto de 2003. Figura publicada em Quaresma (2006).
Figura 5 – Mapa composto de observações de OIS com base num conjunto de 10 imagens SAR. Os contornos identificam isóbatas em metros (200, 1000 e 2000 m). Adaptado de Magalhaes & da Silva (2012).
35
1.2.2 Ondas internas no Golfo da Biscaia
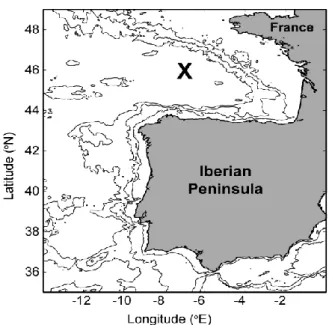
O Golfo da Biscaia está localizado na região nordeste do oceano Atlântico estando delimitado pela costa oeste de França e pela costa norte de Península Ibérica, sendo caracterizado por uma extensa plataforma continental principalmente a norte e nordeste (Fig. 6).
OI de grande amplitude têm sido extensivamente estudadas na região do Golfo da Biscaia e constituem presentemente um fenómeno amplamente documentado. As MI de período semidiurno resultam da interação de fortes correntes da maré superficial (barotrópica) com topografia íngreme do bordo da plataforma continental, tendo Baines (1982) classificado as ondas geradas na vertente da plataforma do Mar Céltico (47° - 48° N, 6° - 8° W) como as mais energéticas do mundo. Estas ondas de grande amplitude (até 50 m), geradas na região norte do bordo da plataforma continental, têm sido observadas a propagar-se tanto em direção à costa (e.g., Pingree & Mardell, 1985), como para oceano profundo (e.g., Pingree et al., 1986). Elas são particularmente intensas no período das marés-vivas, manifestando-se como oscilações de grande comprimento de onda (30 - 50 km) da termoclina sazonal, e foram observadas por Pingree et al. (1986) a propagarem-se para águas profundas com uma velocidade de 1.0 m.s-1 no verão.
O mecanismo de geração da MI conduz frequentemente à formação de OIS (que se propagam na mesma direção da MI), por ação de forças não-lineares e dispersivas. Baseados num vasto conjunto de imagens SAR, New & da Silva (2002), identificaram essas OIS na região norte do bordo da plataforma continental simultaneamente com outras na região central do Golfo da Biscaia, a cerca de 150 km de distância da plataforma continental. As OIS observadas na região central do Golfo, numa região próxima de 46.0° N, 7.0° W (ver Fig. 6), apresentam uma propagação para
36
sudoeste, e terão sido “geradas localmente” pela interceção dos raios de MI com a termoclina (New & Pingree, 1990; 1992; Gerkema, 2001; New & da Silva, 2002).
Figura 6 – Mapa do Golfo da Biscaia relativamente à Península Ibérica e França. Os contornos identificam isóbatas em metros (0, 200, 2000, e 4000 m). A região central encontra-se identificada (46.0° N, 7.0° W).
New (1988) desenvolveu um modelo para explicar este processo de “geração local”. Ao deslocar-se sobre fortes declives topográficos, a maré barotrópica vai gerar uma variedade de modos baroclínicos que se combinam para criar um raio de maré (que correspondem a oscilações de grande amplitude) que emana dessa topografia “crítica” (New, 1988) (ver a definição de topografia “crítica” na secção 1.1.1. Determinados locais da vertente continental na região norte do Golfo da Biscaia (e.g., 47.0° - 47.5° N, 6.5° - 7.0° W) apresentam declives muito pronunciados em que é satisfeita a condição de igualdade (entre o ângulo do raio e o declive topográfico), sendo portanto “críticos”. Desses pontos vão emanar dois raios, sendo que um propaga-se para cima e em direção ao interior da plataforma, enquanto o outro encaminha-se para baixo e em direção ao oceano profundo. Este último vai ser refletido no fundo do
37
oceano e posteriormente atingir a termoclina, a cerca de 150 km da extremidade da plataforma continental, em concordância com os resultados de Pingree & New (1989, 1991). Admite-se então que a reemergência deste raio e a consequente interceção com a termoclina (que ocorre na região central do Golfo da Biscaia), seja responsável pela “geração local” de MI e de OIS de grande amplitude.
Azevedo et al. (2006) e da Silva et al. (2007) demonstram a existência de OIS na região central do Golfo, tendo fundamentado a sua formação identicamente pelo mecanismo de “geração local”, através de raios de MI que emanam de uma região próxima do Cabo Ortegal, no noroeste de Espanha. Ao propagarem-se para nordeste e nor-nordeste, estas ondas apresentam uma direção de propagação oposta àquelas geradas na plataforma continental Francesa, exibidas por New & da Silva (2002). As investigações produzidas na região sudoeste do Golfo da Biscaia (Azevedo et al., 2006 e da Silva et al., 2007) comprovam que este mecanismo de “geração local” desempenha um papel relevante na complexa atividade interna no centro do Golfo, uma vez que é nesta região que as MI são ampliadas, quando os raios, depois de refletidos no fundo do oceano, atingem a termoclina sazonal (Azevedo et al. 2006; Pichon & Correard 2006; Pichon et al., 2011).
Pingree & New (1995) revelaram uma característica notável no centro do Golfo. A região centrada perto de 46.0° N, 7.0° W, tem a aparência de um “olho elíptico” (de escala horizontal de cerca de 50 km) indicando a presença de MI de grande amplitude “geradas localmente”. Estes autores sugeriram que este “olho” poderia resultar da concentração de MI geradas ao longo de um arco quase circular do bordo da plataforma continental, com o “olho” no centro de curvatura da elipse a 170-200 km da vertente da plataforma (Fig. 7).
38 Figura 7 – Zona central do Golfo da Biscaia (46.0° N, 7.0° W) que corresponde à região do “olho elíptico” denominado por Pingree & New (1995) que advém da concentração confluência de OI geradas ao longo de um arco quase circular do no bordo da plataforma continental (identificado pela linha descontinua a laranja).
Adicionalmente, este “olho” encontra-se próximo da posição prevista para as reemergências dos raios das ondas “geradas localmente”, a cerca de 150 km do bordo da plataforma, constituindo assim um foco de raios que emanam de diferentes locais do arco semi-circular, os quais depois de refletidos do fundo oceânico, reemergem próximo da posição do “olho”. Baseados por observações e resultados de modelo, Pichon et al. (2011), reforçaram esta ideia ao revelar fortes interações tridimensionais, na região 45.5° - 46.5° N, 7.2° - 7.5° W, entre MI provenientes de locais de geração diferentes. Estes resultados (movimentos verticais de até 80 m) comprovam que esta região possui uma enorme atividade interna a qualquer nível de profundidade.
39
1.2.3 Efeito de ondas internas no fitoplâncton e na produção primária
As OI são um fenómeno ubíquo de oceanos (e igualmente de grandes lagos) podendo influenciar, de várias formas, os processos biológicos tal como a produtividade do fitoplâncton (e.g., Evans et al., 2008). Os mecanismos mais importantes são o aumento do fluxo de nutrientes para a zona eufótica, através da geração de turbulência e de processos de mistura na região da termoclina (Holligan et al., 1985; Lande & Yentsch, 1988; Wang et al., 2007), e a disponibilização de uma maior exposição luminosa às células de fitoplâncton (Kamykowski, 1974; Haury et al., 1983), uma vez que a intensidade da luz decai exponencialmente com a profundidade. Adicionalmente, as OI intervêm igualmente nos processos biológicos ao transportarem nutrientes para a zona eufótica quando a rebentação ocorre (MacIntyre & Jellison, 2001) e por influenciarem a distribuição desigual do fitoplâncton, fazendo com que este fique agregado em zonas acima das cavas das OI e abaixo destas nas cristas (Lennert-Cody & Franks, 2002).
O acoplamento entre a atividade de OI e descolamentos verticais de fitoplâncton encontra-se amplamente documentado (Sangra et al., 2001; Gaxiola-Castro et al., 2002). O uso de imagens SAR e de cor oceânica, obtidas a partir do Sea-viewing Wide Field-of-view Sensor (SeaWiFS), na região central do Golfo da Biscaia, conduziu da Silva et al. (2002) a revelar expressões superficiais de cristas de ondas de MI em imagens de clorofila a. A Fig. 8 apresenta frentes de OIS (representadas através de linhas contínuas brancas) sobrepostas em imagens SeaWiFS, sendo ainda identificáveis concentrações mais elevadas de clorofila a ao longo de toda a região do bordo da plataforma continental, que têm como origem os processos de mistura já mencionados. Mais a sul, na região central do Golfo da Biscaia, é possível observar estruturas coerentes com níveis de clorofila a mais elevados, que se encontram entre as posições das OIS (que assinalam os locais das cavas da MI) e se dispõem
40
perpendicularmente à direção de propagação das OIS (e da MI). Estes autores justificaram as observações argumentando que as cristas de MI terão elevado o DCM para níveis de profundidade em que o seu topo produziu uma resposta mensurável para os sensores de cor do oceano (neste caso o SeaWiFS).
A propagação de OI introduz movimentos verticais na coluna de água, particularmente pronunciados na termoclina, forçando as partículas de água a movimentos ascendentes e descendentes. Como as células de fitoplâncton são usualmente passivas, elas são deslocadas na vertical de forma significativa. Uma vez que a intensidade da luz decai exponencialmente com a profundidade, as OI expõem as células de fitoplâncton (em média durante um período de maré semidiurna) a uma maior intensidade luminosa.
É expectável que esse aumento da radiação disponível para o fitoplâncton (e do transporte vertical de nutrientes), por ação das OI, possa desempenhar um papel importante em determinados processos biológicos, tais como, afetar positivamente a fotossíntese e, em consequência, induzir um aumento da biomassa. Existem várias referências que identificam, embora meramente através de modelos teóricos, que os movimentos da MI na termoclina conduzem a um aumento da produtividade se estes ocorrerem em profundidades em que a fotossíntese é limitada pela luz (Kahru, 1983; Holloway, 1984; Lande & Yentsch, 1988; Holloway & Denman, 1989). Evans et al. (2008) investigaram amplamente o processo fotossintético resultante do efeito combinado das OI e de flutuações na irradiância à superfície num lago oligotrófico, tendo demonstrado, que esse efeito conjunto pode causar fortes aumentos (até 200%) ou reduções (até 15%) na produtividade primária.
41 Figura 8 – (a) Concentrações de clorofila a obtidas a partir do sensor SeaWiFS às 13h00m UTC de 4 de setembro de 1999, e as posições estimadas de frentes de ondas de grupos de OIS (identificadas por linhas brancas) a partir da imagem ERS-2 SAR adquirida às 22h36m de 3 de Setembro de 1999 (deslocadas 7.4 km na sua direção de propagação (da Silva, et al. (2002) para mais detalhes)); (b) Concentrações de clorofila a obtidas a partir do sensor SeaWiFS às 12h53m UTC de 14 de agosto de 1998, e as posições estimadas de frentes de ondas de grupos de OIS (identificadas por linhas contínuas brancas) a partir da imagem ERS-2 SAR adquirida às 11h17m de 14 de agosto de 1999 (deslocadas 5.9 km na sua direção de propagação (da Silva, et al. (2002) para mais detalhes)). Figura publicada em da Silva, et al. (2002).
Muito embora este processo ainda não esteja devidamente quantificado no oceano (nomeadamente à escala global), começam a surgir resultados que suportam a ideia que as OI induzem efetivamente um aumento de biomassa de fitoplâncton (Wang et al., 2007; Pan et al., 2012) em determinadas regiões oceânicas. Investigações efetuadas recentemente na região norte do Mar do Sul da China vieram destacar a relevância destas ondas na ecologia regional. Wang et al. (2007) observaram grandes
42
variações de oxigénio dissolvido na camada sub-superficial 6 dias após a intrusão de água fria pelo efeito de OI, seguido de mais 2 dias por um aumento significativo nas concentrações de clorofila a. Os autores argumentam que este atraso de 8 dias sugere que o “bloom” do fitoplâncton ocorreu após os nutrientes terem sido libertados pela decomposição microbiana de compostos orgânicos trazidos pela atividade de OI.
A climatologia relativa aos meses de verão, obtida a partir do sensor MODIS, permitiu a Pan et al. (2012) comprovar a existência de concentrações mais elevadas de clorofila a em presença de OI. Os resultados revelaram um aumento médio de 19% nas concentrações superficiais de clorofila a e de 15% na produtividade primária numa zona amplamente afetada pela atividade de OI, quando comparadas com uma região próxima e menos perturbada pela sua presença. Esta anomalia positiva nos valores de clorofila a e de produção primária em dados climatológicos, vieram evidenciar a importância que a persistência de atividade interna tem em fenómenos biológicos, nomeadamente no crescimento do fitoplâncton, em escalas temporais amplas.
Ao demonstrarem os efeitos ecológicos que as OI têm na região norte do Mar do Sul da China (em particular em torno do Atol de Dongsha), Wang et al. (2007) e Pan et al. (2012) lançaram a questão de aferir se este impacto na ecologia regional terá igualmente efeitos noutras regiões oceânicas com características diferentes de um Atol (e.g., Canhões submarinos e em oceano aberto), embora igualmente afetadas por elevada dinâmica interna.
1.3 Objetivos
Tendo em consideração a ubiquidade das OI nos oceanos e o papel determinante do fitoplâncton nos ecossistemas marinhos, o principal objetivo deste estudo foi compreender o efeito de ondas de MI na estrutura superficial do fitoplâncton
43
e avaliar o seu eventual impacto ecológico em dois locais com uma inequívoca dinâmica interna: o Canhão submarino da Nazaré e o centro do Golfo da Biscaia.
Os objetivos específicos foram os seguintes:
1) Descrever o efeito de MI em padrões superficiais de fitoplâncton nas duas regiões de estudo, usando sinergia de sensores remotos (SAR e MODIS);
2) Determinar taxas de produção primária associadas ao efeito da MI na região do Canhão da Nazaré, através de um modelo numérico e de medições in-situ; 3) Avaliar o efeito integrado da convergência de ondas de MI na região central do
Golfo da Biscaia na biomassa de fitoplâncton numa escala temporal alargada, usando dados climatológicos.
1.4 Estrutura da tese
Os objetivos específicos desta Tese são abordados em estudos independentes que se encontram nos capítulos seguintes (II, III) sob a forma de trabalhos em processo de publicação.
Desta forma, o Capítulo I procura facultar um enquadramento teórico dos aspetos mais relevantes, bem como, descrever o estado da arte das OI nas regiões de estudo e apresentar o panorama geral do efeito destas na dinâmica do fitoplâncton e na produção primária. O Capítulo II intitulado “Effect of Internal Waves on
near-surface chlorophyll concentration and primary production in the Nazaré Canyon (West of Iberian Peninsula)” procura responder aos objetivos específicos 1 e 2 (ver
secção 1.3). O Capítulo III intitulado “Chlorophyll enhancement in the central region
of the Bay of Biscay as a result of internal (tidal) wave interaction”, atende aos
objetivos 1 e 3 (ver secção 1.3), numa tentativa de compreender o presumível efeito ecológico nesta região. O Capítulo IV corresponde à conclusão geral desta Tese.
44
Referências
Alpers W (1985). Theory of radar imaging of internal waves. Nature (London) 314, 245-247. doi:10.1038/413245a0.
Apel JR (2003). A New Analytical Model for Internal Solitons in the Ocean. Journal of Physical Oceanography, 33, 2247-2269.
Azevedo A, da Silva JCB, New AL (2006). On the generation and propagation of internal waves in the southern Bay of Biscay. Deep-Sea Res. I 53, 927941. doi:10.1016/j.dsr.2006.01.013.
Baines PG (1982). On internal tides generation models. Deep-Sea Res. Part A 29, 307-338. doi:10.1016/0198-0149(82)90098-X.
Benney DJ (1966). Long non-linear waves in fluid flows. Journal of Mathematical Physics, 45, 52-63.
da Silva JCB, Ermakov SA, Robinson IS, Jeans, DRG, Kijashko SV (1998). Role of surface films in ERS SAR signatures of internal waves on the shelf. 1. Shortperiod of internal waves. J. Geophys. Res. 103 (C4), 8009-8031. doi:10.1029/97JC02725.
da Silva JCB, New AL, Srokosz M, Smyth T (2002). On the observability of internal tidal waves in remotely-sensed ocean color data. Geophys. Res. Lett. 29 (12), 1569-1572. doi:10.1029/2001GL013888.
da Silva JCB, Correia SM, Ermakov SA, Sergievskaya IA, Robinson IS (2003). Synergy of MERIS ASAR for observing marine film slicks and small scale processes. Paper presented at Proc. MERIS User Workshop, November, Frascati, Italy. ESA, Noordwijk, The Netherlands.
da Silva JCB, New AL, Azevedo A (2007). On the role of SAR for observing “Local Generation” of internal solitary waves off the Iberian Peninsula. Can. J. Remote Sens. 33, 388-403. doi:10.5589/m07-041.
da Silva, JCB, New AL, Magalhaes JM (2011). On the structure and propagation of internal solitary waves generated at the Mascarene Plateau in the Indian Ocean. Deep-Sea Res. I 58, 229-240. doi:10.1016/j.dsr.2010.12.003.
Evans MA, MacIntyre S, Kling GW (2008). Internal wave effects on photosynthesis: Experiments, theory, and modeling. Limnol. Oceanogr., 53, 339–353.
Field CB, Behrenfeld MJ, Randerson JT, Falkowsky P (1998). Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science, Vol. 281, pp. 237–240.
45 Gaxiola-Castro G, Alvarez-Borrego S, Najera-Martinez S, Zirino AR (2002). Internal waves effect on the Gulf of California phytoplankton. Cienc. Mar., 28: 297–309.
Gerkema T (2001). Internal and interfacial tides: beam scattering and local generation of solitary waves. J. Mar. Res. 59, 227-255. doi:10.1357/002224001762882646.
Gerkema T, Zimmerman JTF (2008). An Introduction to Internal Waves. Lectures notes, Royal NIOZ, Texel, 207 pp.
Gordon HR, McCluney WR (1975). Estimation of the depth of sunlight penetration in the sea for remote sensing. Applied Optics, Vol. 14, No. 2, 413-416.
Haury LR, Wiebe PH, Orr MH, Brisco MG (1983). Tidally generated high-frequency internal wave packets and their effects on plankton in Massachusetts Bay. J. Mar. Res. 41: 65– 112
Holligan PM, Pingree RD, Mardell GT (1985). Oceanic solitons, nutrient pulses and phytoplankton growth. Nature 314, 348–350.
Holloway G (1984). Effects of velocity fluctuations on vertical distributions of phytoplankton. J. Mar. Res. 42: 559–571
Holloway G, Denman K (1989). Influence of internal waves on primary production, Journal of Plankton Research, 11, 409–413.
Huthnance JM (1995). Circulation, exchange and water masses at the ocean margin: the role of physical processes at the shelf edge. Progress in Oceanography, 35, 353- 431.
Jackson CR (2007). Internal wave detection using the moderate resolution imaging spectroradiometer (MODIS). J. Geophys. Res. 112, C11012.doi:10.1029/2007JC004220.
Jeans DRG (1995). Solitary Internal Waves in the Ocean: A Literature Review Completed as Part of the Internal Waves Contribution to Morena, University College of North Wales, Menai Bridge, United Kingdom. 64 p.
Jeans DRG, Sherwin TJ (2001a). The variability of strongly nonlinear solitary internal waves observed during an upwelling season on the Portuguese Shelf. Cont. Shelf Res. 21, 1855– 1878.
Jeans DRG, Sherwin TJ (2001b). Energetics of non-linear internal waves on the Portuguese Shelf. J. Mar. Res. 59, 327–353.
Kahru M (1983). Phytoplankton patchiness generated by long internal waves: a model, Marine Ecology - Progress Series, 10, 111-117.
46 Kamykowski D (1974). Possible interactions between plankton and semidiurnal internal tides. J. Mar. Res. 32: 67–89.
Kantha LH, Tierney CC (1997). Global baroclinic tides. Progress in Oceanography, 40, 163-178.
Kirk JTO (1996). Light and photosynthesis in aquatic ecosystems, Cambridge University Press, Cambridge, 509 pp.
Lande R, Yentsch CS (1988). Internal waves, primary production and the compensation depth of marine phytoplankton, Journal of Phytoplankton Research, 10, 565-571.
Lennert-Cody CE, Franks PJS (2002). Fluorescence patches in high-frequency internal waves. Mar. Ecol. Prog. Ser. 235: 29–42.
Lindeman RL (1942). The trophic-dynamic aspect of ecology, Ecology, Vol. 23, 399-418. MaCIntyre S, Jellison R (2001). Nutrient fluxes from upwelling and enhanced turbulence at the top of the pycnocline in Mono Lake, California. Hydrobiologia 466: 13–29.
Magalhaes JM, da Silva JCB (2012). SAR Observations of Internal Solitary Waves Generated at the Estremadura Promontory off the West Iberian Coast. Deep-Sea Res. I, 69, 12-24, http://dx.doi.org/10.1016/j.bbr.2011.03.031.
New AL (1988). Internal tidal mixing in the Bay of Biscay. Deep-Sea Research 35, 691– 709.
New AL, Pingree RD (1990). Large-amplitude internal soliton packets in the central Bay of Biscay. Deep-Sea Research 37, 513–524.
New AL, Pingree RD (1992). Local generation of internal soliton packets in the central Bay of Biscay. Deep-Sea Res. Part A 39, 1521-1534. doi:10.1016/0198- 0149(92)90045-U.
New AL, da Silva JCB (2002). Remote-sensing evidence for the local generation of internal soliton packets in the central Bay of Biscay. Deep-Sea Res. I 49, 915-934. doi:10.1016/S0967-0637(01)00082-6.
Pan X-J, Wong GTF, Shiah F-K, Ho T-Y (2012). Enhancement of biological productivity by internal waves: observations in the summertime in the northern South China Sea. Journal of Oceanography doi:10.1007/s10872-012-0107-y.
Platt T, Gallecos CL, Harrison WG (1980). Photoinhibition on photosynthesis in natural assemblages of marine phytoplankton, Journal of Marine Research, 38, 687-701.
47 Pichon A, Correard S (2006). Internal tides modelling in the Bay of Biscay. Comparisons with observations. Scientia Marina 70 (S1), 65-88. doi:10.3989/scimar.2006.70s165.
Pichon A, Morel Y, Baraille R, Quaresma LS (2011). Internal tide interactions in the Bay of Biscay: observations and modelling. J. Mar. Syst. doi:10.1016/j.jmarsys.2011.07.003.
Pingree RD, Mardell GT (1985). Solitary internal waves in the Celtic Sea. Progress in Oceanography 14, 431–441.
Pingree RD, Mardell GT, New AL (1986). Propagation of internal tides from the upper slopes of the Bay of Biscay. Nature 312, 154-158. doi:10.1038/321154a.
Pingree RD, New AL (1989). Downward propagation of internal tidal energy into the Bay of Biscay. Deep Sea Res. Part A, 36, 735-758. doi:10.1016/0198-0149(89)90148-9.
Pingree RD, New AL (1991). Abyssal penetration and bottom reflection of internal tidal energy in the Bay of Biscay. J. Phys. Oceanogr. 21, 28-39. doi:10.1175/1520- 0485(1991)021<0028:APABRO>2.0.CO;2
Pingree RD, New AL (1995). Structure, seasonaldevel opment and sunglint spatial coherence of the internal tide on the Celtic and Armorican shelves and in the Bay of Biscay. Deep-Sea Research Part I 42, 245–284.
Quaresma LS (2006). Observação de ondas internas não-lineares geradas sobre o canhão da Nazaré. M.Sc Thesis, Lisbon University, Portugal, unpublished.
Quaresma LS, Vitorino J, Oliveira A, da Silva J (2007). Evidence of sediment resuspension by nonlinear internal waves on the western Portuguese mid-shelf. Marine Geology 246, 123–143.
Robinson IS (2010). Discovering the ocean from space: the unique applications of satellite oceanography, Berlin, Germany, Springer/Praxis Publishing, 638pp.
Sangra P, Basterretxea G, Pelegri JL, Aristegui J (2001). Chlorophyll increase due to internal waves on the shelf break of Gran Canaria (Canary Islands), Sci. Mar., 65(Suppl. 1): 89-97.
Small J (2002). Internal tide transformation across a continental slope off Cape Sines, Portugal. Journal of Marine Systems, 32:43-69.
Staquet C, Sommeria J (2002). Internal Gravity Waves: From Instabilities to Turbulence. Annual Review of Fluid Mechanics, 34, 559-593.
Vanney JR, Mougenot D (1990). Un canyon sous-marin du type “gouf”: le Canhão da Nazaré (Portugal). Oceanol. Acta 13, 1–14.
48 Vaulot D (2006) Phytoplankton. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0004306]
Vitorino J, Oliveira A, Beja J (2005). The Nazare Canyon (W Portugal): Physical processes and sedimentary impacts. Geophysical Research Abstracts, EGU2005, 7, 10013.
Wang YH, Dai CF, Chen YY (2007). Physical and ecological processes of internal waves on an isolated reef ecosystem in the South China Sea. Geophys Res Lett 34:L18609. doi:10.1029/2007GL030658
Wang C, Pawlowicz R (2012). Oblique wave-wave interactions of nonlinear near-surface internal waves in the Strait of Georgia, J. Geophys. Res., 117, C06031, doi:10.1029/2012JC008022.
49