UNIVERSIDADE ESTADUAL DE MARINGÁ
DEPARTAMENTO DE BIOLOGIA CELULAR E GENÉTICA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
BIOLOGIA CELULAR E MOLECULAR
RELAÇÕES FILOGENÉTICAS EM CALLICHTHYINAE
(SILURIFORMES: CALLICHTHYIDAE): UMA CONTRIBUIÇÃO
CITOGENÉTICA AOS ESTUDOS EVOLUTIVOS.
JOCICLÉIA THUMS KONERAT
2
JOCICLÉIA THUMS KONERAT
RELAÇÕES FILOGENÉTICAS EM CALLICHTHYINAE
(SILURIFORMES: CALLICHTHYIDAE): UMA CONTRIBUIÇÃO
CITOGENÉTICA AOS ESTUDOS EVOLUTIVOS.
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Biológicas (área de concentração - Biologia Celular e Molecular) da Universidade Estadual de Maringá para obtenção do grau de Mestre em Ciências Biológicas.
3
JOCICLÉIA THUMS KONERAT
RELAÇÕES FILOGENÉTICAS EM CALLICHTHYINAE
(SILURIFORMES: CALLICHTHYIDAE): UMA CONTRIBUIÇÃO
CITOGENÉTICA AOS ESTUDOS EVOLUTIVOS.
Orientadora:
Dra. Isabel Cristina Martins dos Santos
Co-orientador
: Dr. Vladimir Pavan Margarido
Ficha catalográfica
Elaborada pela Bibliotecária Jeanine S. Barros CRB-9/1362
Konerat, Jocicléia Thums
K822r Relações filogenéticas em Callichthyinae (Siluriformes: Callichthyinae): uma contribuição citogenética aos estudos evolutivos. / Jocicléia Thums Konerat.— Maringá, PR: UEM, 2008.
29 f. ; 30 cm
Orientadora: Profa. Dra. Isabel Cristina Martins dos Santos Co-orientador: Prof. Dr. Vladimir Pavan Margarido
Dissertação (Mestrado) – Universidade Estadual de Maringá.
Programa de Pós-Graduação em Ciências Biológicas área de concentração Biologia Celular e Molecular.
Bibliografia
1. Evolução cromossômica. 2. Filogenia. 3. Marcadores citogenéticos. I. Santos, Isabel Cristina Martins dos. II. Margarido, Vladimir Pavan. III. Universidade Estadual de Maringá. IV. Título.
4
BIOGRAFIA
Jocicléia Thums Konerat nasceu em São Miguel do Oeste/SC em 14/11/1974.
Possui graduação em Ciências Biológicas com ênfase em Biotecnologia pela Universidade Paranaense – UNIPAR campus Cascavel; Pós-graduação lato sensu
em Biotecnologia e Análise da Biodiversidade pela Universidade Paranaense – UNIPAR campus Toledo; atualmente mestranda em Ciências Biológicas, área de
6
! ! ! !
" #
" #
" #
" #
$ %
$ %
$ %
$ %
&&&&
' ( & ) # *
'' ( &( & )) ## **
' ( & ) # *
7
AGRADECIMENTOS
A universidade Estadual de Maringá pela oferta do Programa de Pós-Graduação em Ciências Biológicas área de concentração Biologia Celular e Molecular.
A Universidade Estadual do Oeste do Paraná pela estrutura e condições essenciais para o desenvolvimento da parte prática desta pesquisa.
Aos professores que ministraram as disciplinas pela dedicação. Ao Nelsino pelo atendimento carinhoso em todas as horas.
A Profª. Dra. Isabel Cristina Martins dos Santos, pelo aceite da orientação; agradeço à compreensão permitindo o desenvolvimento desta pesquisa, fora das dependências da UEM.
Aos membros da banca Prof. Dr. Roberto Ferreira Artoni e Prof. Dr. Horácio Ferreira Júlio Junior.
A Dra. Carla Simone Pavanelli e Msc. Weferson Júnio da Graça pela identificação das espécies.
Ao Prof. Dr. Vladimir Pavan Margarido, pelas palavras de incentivo, confiança, e, acima de tudo por em momento tão difícil, ter aceitado co-orientar uma desconhecida. Neste ano de pesquisa o aprendizado não se ateve apenas ao tema da pesquisa, permitindo crescimento pessoal e profissional. Conviver ao lado de um grande profissional, instiga-nos a melhorar. Obrigado!!!!!
8
Joselaine e Leandra amigas que dividiram comigo, além do local para morar, as alegrias e tristezas vivenciadas neste período.
Aos colegas de laboratório de Cascavel: Maelin, Roberto, Vanessa, Eduarda, Ana Caroline, Jéssica, Darlene, Larissa, Fernandes, Profª Dra. Nereide, Profª Dra. Maria Amélia, Profª Dra. Luciana; e os colegas de laboratório de Maringá: Fernanda, Renata, Paula, Paulo, Suzana, Ana Paula, Carlos Alexandre, Maria José e Leonardo por propiciarem a troca de experiências e conhecimentos, aliados aos momentos de descontração.
A Leonardo e Edilson, pela compreensão, carinho, incentivo em todos os sentidos. Amo vocês!!!
9
SUMÁRIO
APRESENTAÇÃO...10
RESUMO ...11
ABSTRACT ...12
Relações filogenéticas em Callichthyinae (Siluriformes: Callichthyidae): uma contribuição citogenética aos estudos evolutivos....13
RESUMO ...13
INTRODUÇÃO ...14
MATERIAL E MÉTODOS...15
RESULTADOS...16
DISCUSSÃO ...17
Figura 1. Cariótipos corados por Giemsa. ...22
Figura 2. Cromossomos portadores das RONs ...22
Figura 3. Cariótipos C-bandados ...23
Figura 4. Evolução de caracteres citogenéticos em Callichthyinae. . ...23
Tabela I – Sumário dos dados citogenéticos em Callichthyinae. ...24
10
APRESENTAÇÃO
Esta dissertação é composta por um artigo, o qual foi redigido a partir de estudos citogenéticos em três espécies, Callichthys callichthys, Hoplosternum littorale e Lepthoplosternum pectorale, pertencentes à subfamília Callichthyinae. Os
espécimens foram coletados no Rio Paraná – Guaíra (PR). O referido artigo, em consonância com as regras do Programa de Pós-graduação em Ciências Biológicas, segue as normas da revista a qual será submetido para publicação.
11
RESUMO
Siluriformes é a ordem mais especiosa, diversificada e amplamente distribuída entre os peixes de água doce, possui 36 famílias e cerca de 3.000 espécies. Callichthyidae, uma das maiores famílias entre os Siluriformes neotropicais, é constituída por duas subfamílias, Corydoradinae e Callichthyinae, e compreende 194 espécies. A escassez de estudos anatômicos comparativos dificulta a proposição de hipóteses de relacionamento filogenético na família.
O presente trabalho foi realizado em três espécies de Callichthyinae:
Callichthys callichthys, Hoplosternum littorale e Lepthoplosternum pectorale. As
espécies foram coletadas no alto rio Paraná (Guaíra/PR) e submetidas ao estudo citogenético pela coloração convencional por Giemsa, impregnação pela prata e bandamento-C, com o objetivo de caracterizar os cariótipos destas espécies e correlacionar os mesmos de forma evolutiva, buscando confirmar as relações filogenéticas nesta subfamília. Ainda, neste trabalho foi realizada a primeira descrição citogenética para o gênero Lepthoplosternum, além do primeiro registro de L. pectorale para a bacia do Alto rio Paraná acima do reservatório de Itaipu.
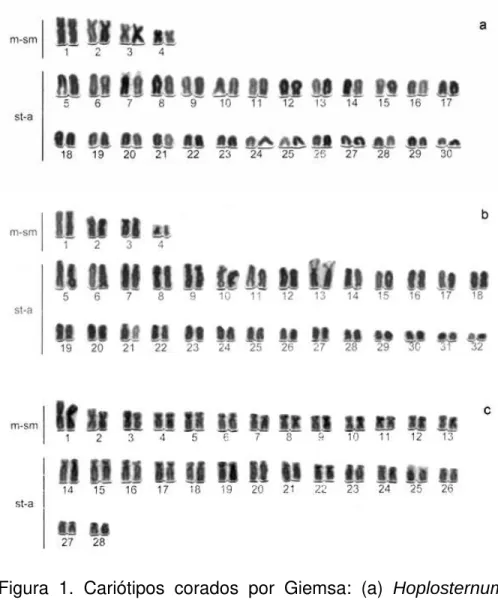
A análise dos resultados mostrou em C. callichthys 2n = 56 cromossomos (26 m-sm + 30 st-a); em L. pectorale 2n = 64 cromossomos (08 m-sm + 56 st-a); e em H. littorale 2n = 60 cromossomos (8 m-sm + 52 st-a). O padrão de distribuição da
heterocromatina mostrou-se comum às três espécies, com marcações conspícuas na região centromérica dos cromossomos, sendo que em H. littorale também foram
verificadas marcações intersticiais. Quanto às regiões organizadoras de nucléolos (RONs), diferentes situações foram verificadas: RONs simples em H. littorale e C. callichthys, e múltiplas em L. pectorale, com variação numérica intra e
inter-individual.
Embora corroborando a hipótese de Reis (1998) como Callichthys sendo o
12
ABSTRACT
Siluriformes is the most specious, diversified and widely distributed order among the freshwater fishes, possesses 36 families and around 3.000 species. Callichthyidae, one of the biggest families among the neotropical Siluriformes, is composed by two subfamily: Corydoradinae and Callichthyinae, and comprising 194 species. The lack of comparatives anatomical studies difficulties the proposition of phylogenetical relationship in the family.
The present study was carried out in three Callichthyinae species: Callichthys callichthys, Hoplosternum littorale and Lepthoplosternum pectorale. All the species
were collected in upper Paraná river (Guaíra/PR), and cytogenetically studied through Giemsa staining, silver staining and C-banding, with the aim to characterize the karyotype and to correlate them in a evolutive approach, trying to confirm the phylogenetic relationship in this subfamily. Moreover, this is the first cytogenetical description for Lepthoplosternum genus, beyond the first register of L. pectorale for
the upper Paraná river basin above of the Itaipu Reservoir.
The analysis showed in C. callichthys 2n = 56 chromosomes (26 m-sm + 30 st-a); in L. pectorale 2n = 64 chromosomes (8 m-sm + 56 st-a); in H. littorale 2n = 60
chromosomes (8 m-sm + 52 st-a). The heterochromatin distribution pattern showed a
common feature among the three species, with conspicuous blocks in the centromeric region of the chromosomes; in H. littorale it had been also verified
interstitial blocks. In relation to the NORs, different situations had been verified: single NORs in H. littorale and C. callichthys, whereas multiple NORs in L. pectorale,
with intra and inter-individual number variation.
Although corroborating Reis (1998) hypothesis that points Callichthys as the
13
Relações filogenéticas em Callichthyinae (Siluriformes: Callichthyidae): uma
contribuição citogenética aos estudos evolutivos.
Jocicléia Thums Konerat*, Isabel Cristina Martins dos Santos* e Vladimir Pavan Margarido**
*Universidade Estadual de Maringá, Departamento de Biologia Celular e Genética. Avenida Colombo, 5790, 87020-900, Maringá, PR – Brasil, ** Universidade Estadual
do Oeste do Paraná, Centro de Ciências Biológicas e da Saúde, Cascavel, PR, Brasil.
RESUMO
No presente trabalho foram analisadas, através de parâmetros citogenéticos, três espécies pertencentes à subfamília Callichthyinae coletadas no Rio Paraná (Guaíra – PR). A análise dos resultados mostrou em Callichthys callichthys 2n = 56
cromossomos (26 m-sm + 30 st-a), em Lepthoplosternum pectorale, 2n = 64
cromossomos (8 m-sm + 56 st-a), e em Hoplosternum littorale, 2n = 60
cromossomos (8 m-sm + 52 st-a). Quanto à localização das RONs pela impregnação
por prata, diferentes situações foram verificadas: RONs simples em H. littorale e C.
callichthys, enquanto múltiplas em L. pectorale, com variação numérica intra e
inter-individual. A heterocromatina foi verificada na região centromérica dos cromossomos nas três espécies, sendo que em H. littorale também foram verificadas marcações
intersticiais equilocais. Através de parâmetros citogenéticos (número diplóide, fórmula cromossômica e Ag-RONs), uma nova hipótese de relacionamento filogenético em Callichthyinae é apresentada.
14
INTRODUÇÃO
A ictiofauna neotropical de água doce é a mais diversificada e rica; das 13.000 espécies existentes no mundo, aproximadamente 6.000 espécies encontram-se na América Central e do Sul, correspondendo a uma grande parcela da diversidade de peixes do mundo (Reis, 2003). Frente a esta diversidade, o número de espécies citogeneticamente investigadas é bastante diminuto; contudo, as informações cariotípicas existentes, por sua relevância, contribuem para o conhecimento da biologia, sistemática e evolução desse importante grupo de vertebrados (Artoni et al., 2000).
A ordem Siluriformes compreende 36 famílias, 478 gêneros e 3.093 espécies, sendo uma das mais especiosas, diversificadas e amplamente distribuídas dentre os peixes de água doce. A família Callichthyidae, uma das maiores entre os Siluriformes neotropicais, é constituída por 194 espécies agrupadas em 8 gêneros (Ferraris, 2007) Seus representantes possuem vasta distribuição geográfica, e este fator associado à escassez de estudos anatômicos comparativos dificulta hipóteses de relacionamento filogenético, sugerindo novas revisões taxonômicas (Reis, 1998). Segundo estudos baseados em caracteres morfológicos e moleculares, Callichthyidae é considerada monofilética, compreendendo dois grupos naturais: Corydoradinae (Aspidoras, Scleromystax e Corydoras) e Callichthyinae (Callichthys,
Dianema, Hoplosternum, Lepthoplosternum e Megalechis) (Reis, 1998; Britto, 2003;
Shimabukuro-Dias et al., 2004).
15
al., 2005; Fernandes & Martins-Santos, 2005) e citossistemáticos (Oliveira et al.,
1993; Margarido & Galetti Jr., 1999; Artoni & Bertollo, 2001). Estudos desta natureza em Callichthyinae têm mostrado número diplóide variando de 52 a 66 cromossomos e apresentando variação na estrutura cariotípica (Martins-Santos et al., 1990;
Marcon et al., 1992; Porto & Feldberg,1992; Porto et al., 1992; Oliveira et al., 1993;
Porto & Feldberg,1993; Shimabukuro-Dias et al., 2005).
No presente estudo são analisadas citogeneticamente Callichthys callichthys
Linnaeus, 1758, Hoplosternum littorale (Hancock, 1828) e Lepthoplosternum
pectorale (Boulenger, 1895), sendo a primeira descrição citogenética para o gênero Lepthoplosternum, além do primeiro registro de L. pectorale para a bacia do Alto rio
Paraná acima do reservatório de Itaipu. Neste contexto, utilizou-se os parâmetros citogenéticos para discutir as proposições filogenéticas de Oliveira et al. (1993) e de
Reis (1998) e para este grupo.
MATERIAL E MÉTODOS
Os estudos citogenéticos foram realizados em Hoplosternum littorale (5 machos e 3
fêmeas), Lepthoplosternum pectorale (5 machos e 13 fêmeas) e Callichthys
callichthys (6 machos e 1 fêmea) coletados no Rio Paraná (Guaíra- PR). Os
espécimens analisados foram depositados no Núcleo de Pesquisas em Limnologia, Ictiologia e Aqüicultura (NUPÉLIA) da Universidade Estadual de Maringá: NUP 6095 – C. callichthys; NUP 6096 – H. littorale e NUP 6097 - L. pectorale. As células
metafásicas foram obtidas a partir do rim (Bertollo et al., 1978; Foresti et al., 1993). A
16
nitrato de prata (Howel & Black, 1980). Os tipos cromossômicos foram classificados de acordo com os critérios de relação de braços propostos por Levan et al. (1964).
Para a análise filogenética foram utilizados os seguintes caracteres: número diplóide, Ag-RONs, padrão de distribuição de heterocromatina, macroestrutura cromossômica; utilizou-se como grupo externo Loricariidae e Diplomystidae, com análise dos caracteres e confecção do clado sendo manual baseada em parcimônia.
RESULTADOS
Hoplosternum littorale
A análise das células mitóticas mostrou um número diplóide de 60 cromossomos (8
m-sm + 52 st-a) para ambos os sexos (Fig.1a). As Ag-RONs foram localizadas na
região terminal do braço curto do par 07 (Fig.2a). Blocos heterocromáticos foram verificados na posição centromérica de todos os pares cromossômicos, além de marcações equilocais em regiões intersticiais-proximais nos pares 5, 6, 7, 8, 10, 11, 12, 13, 14, 15, 19, 20, 21 e 22 (Fig.3a).
Lepthoplosternum pectorale
A análise das células mitóticas mostrou um número diplóide de 64 cromossomos (8
m-sm + 56 st-a) para ambos os sexos (Fig. 1b). As Ag-RONs foram localizadas
17
centromérica, sendo mais conspícuos nos pares 2, 3, 5, 9, 10, 11, 12, 13, 14, 15, 16, 17, 21, 23, 24, 25, 27 e 28; heterocromatina intersticial foi observada no braço longo do primeiro par de cromossomos m-sm (Fig. 3b).
Callichthys callichthys
A análise das células mitóticas mostrou um número diplóide de 56 cromossomos (26
m-sm + 30 st-a) para ambos os sexos (Fig. 1c). As Ag-RONs foram localizadas
terminalmente no braço curto do par 25 (Fig. 2c). Bandamento C revelou blocos heterocromáticos na posição centromérica dos pares cromossômicos 1, 4, 6, 7, 14, 15, 16, 18, 25, 27, 28, e marcações discretas em posição telomérica de alguns pares de cromossomos (Fig. 3c).
DISCUSSÃO
A subfamília Callichthyinae apresenta cinco gêneros (Dianema,
Hoplosternum, Lepthoplosternum, Megalechis e Callichthys), sendo que destes
apenas Lepthoplosternum não apresentava estudos citogenéticos realizados.
Lepthoplosternum possui seis espécies descritas (L. altamazonicum, L. beni, L. tordilho, L. ucamara, L. stellatum e L. pectorale), sendo que os primeiros dados
citogenéticos obtidos no presente estudo revelam número diplóide de 64 cromossomos para L. pectorale.
A análise citogenética das três espécies analisadas no presente trabalho mostra diferenças de número diplóide, variando de 56 a 64 cromossomos, com Ag-RONs localizadas no braço curto em um único par para H. littorale e C. callichthys, e
18
cariotípica e RONs para diferentes espécies da subfamília Callichthyinae são apresentados na tabela 1. Para Hoplosternum, das três espécies descritas (H.
littorale, H. magdalenae e H. punctatum), apenas H. litttorale foi estudado
citogeneticamente, apresentando 60 cromossomos (Porto & Feldberg, 1992; Shimabukuro-Dias et al., 2005). Em Megalechis, com duas espécies descritas (M.
thoracata e M. picta), o número cromossômico foi detectado apenas para M. thoracata, variando de 62 cromossomos (citado como Megalechis personata,
Shimabukuro-Dias et al., 2005) a 66 cromossomos (citado como Hoplosternum aff.
thoracatum, Porto et al., 1992). Dianema, com descrição de duas espécies, D. longibarbis e D. urostriatum, a variação encontrada é de 60 a 62 cromossomos,
respectivamente (Marcon et al., 1992; Oliveira et al., 1993). Embora Callichthys
apresente quatro espécies descritas (C. fabricioi, C. oibaensis, C. serralabium, C.
callichthys), apenas C. callichthys possui estudos citogenéticos em diferentes
populações, apresentando variabilidade ainda maior em relação aos demais gêneros quanto ao número diplóide (52 a 58 cromossomos), além da presença de 1-8 cromossomos supranumerários (Martins-Santos et al., 1990; Porto & Feldeberg,
1993; Oliveira et al., 1993; Sanchez & Fenocchio, 1996; Shimabukuro-Dias et al.,
2005), sugerindo tratar-se de um “complexo” de espécies.
Em relação ao número diplóide de C. callichthys, 56 cromossomos foi
19
A variação de número diplóide em Callichthyinae (52 a 66 cromossomos) é pequena quando comparada à Corydoradinae, a qual apresenta número diplóide variando de 40 a 134 cromossomos (Oliveira et al., 1990; Turner et al., 1992), e cuja
diversificação cariotípica tem sido relacionada a eventos de poliploidia (Oliveira et
al., 1993). Apesar da diversificação cromossômica que leve a variação do número
diplóide dentro de cada gênero ser um evento independente, os mesmos tipos de rearranjos acima sugeridos ocorrem nos diferentes gêneros de Callichthyinae, sendo eventos recorrentes (homoplásicos). Esta situação já fora observada na origem de sistemas de cromossomos sexuais múltiplos em diferentes gêneros de Erythrinidae (Hoplias malabaricus, Bertollo et al., 1997; Erythrinus erythrinus, Bertollo et al.,
2004), ou ainda em diferentes famílias de Gymnotiformes (Eigenmannia virescens,
Almeida-Toledo et al., 2000a; Brachyhypopomus pinicaudatus, Almeida-Toledo et al.,
2000b; Gymnotus pantanal citado como Gymnotus sp., Silva & Margarido, 2005).
Com relação à evolução cromossômica em Siluriformes, Oliveira & Gosztonyi (2000) verificaram em Olivaichthys mesembrinus (citado como Diplomystes
mesembrinus) o número diplóide de 56 cromossomos, sendo que Diplomystidae é
considerada a família mais basal da ordem. De acordo com as relações filogenéticas em Siluriformes, Callichthyidae é considerado filogeneticamente próximo a Loricariidae (Pinna, 1998), e neste contexto, sugere-se que Callichthys ocuparia a
posição mais basal de Callichthyidae (Oliveira et al., 1993) e Callichthyinae (Reis,
1998). Através de estudos citogenéticos em três diferentes subfamílias de Loricariidae, Artoni & Bertollo (2001) sugerem número diplóide de 54 cromossomos como plesiomórfico para esta família; corroborando estes estudos, Alves et al.
20
em Callichthyinae (Tab.1) (Dianema, Hoplosternum, Lepthoplosternum, Megalechis e
Callichthys) sugere-se Callichthys callichthys como basal neste grupo por apresentar
número diplóide de 52-58 cromossomos, predominantemente dos tipos m-sm.
Diferentemente Dianema, Hoplosternum, Lepthoplosternum e Megalechis
apresentam número diplóide mais elevado, variando de 60-66 cromossomos, com maior proporção de cromossomos dos tipos st-a, indicando serem grupos mais
derivados, corroborando então as proposições de Oliveira et al. (1993) e Reis
(1998). Esta proposta diverge dos estudos de DNA mitocondrial realizados por Shimabukuro-Dias et al. (2004), que utilizaram as seqüências 12S rRNA, 16S rRNA,
ND4, tRNAHist e tRNASer e propõem um relacionamento diferente em Callichthyinae, tanto dos baseados em caracteres morfoanatômicos (Reis, 1998) como dos caracteres citogenéticos (presente estudo). De acordo com este estudo, Dianema e
Hoplosternum são mais basais nesta subfamília, seguido por Callichthys
considerado grupo irmão de Leptthoplosternum e Megalechis, ambos mais
derivados.
Os padrões de distribuição heterocromatina observado em C. callichthys, L.
pectorale e H. littorale, em conjunto com dados disponíveis na literatura permite
corroborar a proposta de Oliveira et al. (1993) sobre os eventos responsáveis pela
diversidade da constituição cromossômica do grupo, como facilitadores de rearranjos, além da ocorrência de eventos de dispersão e amplificação de heterocromatina. Particularmente em H. littorale e D. urostriatum (Oliveira et al.;
21
Margarido & Galetti Jr. (2000) em Leporinus desmotes. Ainda, este caráter pode
representar uma sinapomorfia para estes dois gêneros.
Em relação às RONs foram verificadas em C. callichthys e H. littorale RONs
simples enquanto L. pectorale apresentou RONs múltiplas, com ocorrência de
polimorfismo intra- e inter-individual no número e localização das RONs. D.
uriostriatum (Oliveira et al., 1993) e M. thoracata (Shimabukuro-Dias et al., 2005)
possuem RONs telómericas simples e múltiplas, respectivamente. A ocorrência do caráter RONs simples em Olivaichthys mesembrinus (Oliveira & Gosztonyi, 2000),
considerado basal em Siluriformes, fortalece a hipótese de C. callichthys como basal
e L. pectorale, M. thoracata, como mais derivados na subfamília Callichthyinae
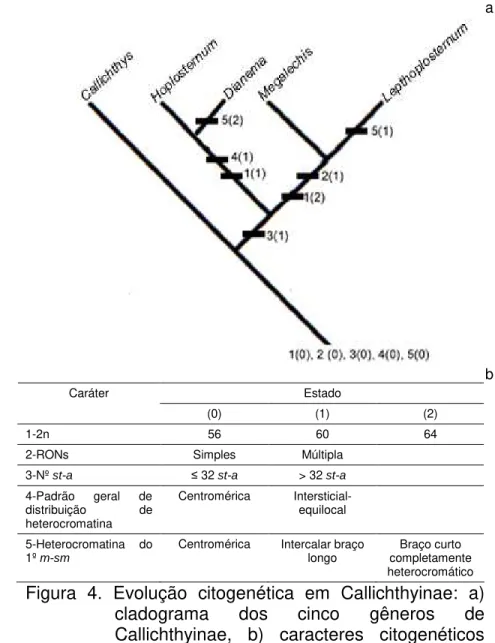
(Fig.4).
Embora corroborando a hipótese de Reis (1998) como Callichthys sendo o
gênero mais primitivo no grupo, uma nova proposta de relacionamento entre os gêneros é apresentada (Fig. 4), utilizando-se os dados de número diplóide, fórmula cromossômica, padrão de distribuição de heterocromatina e número de RONs apresentados no presente trabalho em conjunto com os dados citogenéticos disponíveis na literatura para a subfamília (Tab. 1). Nesta nova proposta, Callichthys
permanece em posição mais basal, seguido por Hoplosternum e Dianema, todos os
três gêneros apresentando RONs simples, e número diplóide mais baixo (52-58, 60 e 60-62 cromossomos, respectivamente). Megalechis e Lepthoplosternum
22
Figura 1. Cariótipos corados por Giemsa: (a) Hoplosternum littorale, (b) Lepthoplosternum pectorale e (c) Callichthys callichthys.
23
Figura 3. Cariótipos C-bandados: (a) Hoplosternum littorale, (b)
Lepthoplosternum pectorale e (c) Callichthys callichthys.
a
b
Estado Caráter
(0) (1) (2)
1-2n 56 60 64
2-RONs Simples Múltipla
3-Nº st-a ≤ 32 st-a > 32 st-a
4-Padrão geral de
distribuição de
heterocromatina
Centromérica
Intersticial-equilocal
5-Heterocromatina do
1º m-sm
Centromérica Intercalar braço
longo completamente Braço curto
heterocromático Figura 4. Evolução citogenética em Callichthyinae: a)
24
Tabela I – Sumário dos dados citogenéticos em Callichthyinae.
Espécies Localidade 2n Fórmula Cromossômica RONs Referências
Callichthys callichthys Candirú - AM 52 44 m-sm + 8 st-a Simples Porto & Feldberg (1993) C. callichthys Marchantaria - AM 54 46 m-sm + 8 st-a * Múltiplas Porto & Feldberg (1993)
C. callichthys Alto RIo Paraná - PR 54 - - Martins-Santos et al. (1990)
C. callichthys Alto Rio Paraná - PR 56 26 m-sm + 30 st-a Simples Presente estudo
C. callichthys Córrego do Pombo - SP 56 22 m + 16 sm + 18 st Simples Shimabukuro-Dias et al. (2005)
C. callichthys Reconquista - ARG. 56 14 m + 10 sm + 32 st-a Simples Sanchez & Fenocchio (1996) C. callichthys Universidade da Amazônia - AM 56-58 - Simples Porto & Feldberg (1993) C. callichthys Itanhaén - SP 58 22 m + 22 sm + 14 st Simples Oliveira et al. (1993)
C. callichthys Guarulhos - SP 58 22 m + 22 sm + 14 st Simples Oliveira et al. (1993)
C. callichthys Ribeirão Santa Rita - SP 58 18 m + 16 sm + 24 st Simples Shimabukuro-Dias et al. (2005) C. callichthys Ribeirão do Convento - SP 58 18 m + 14 sm + 26 st Simples Shimabukuro-Dias et al. (2005) C. callichthys Corumbá - MT 56/57 20 m + 16 sm + 20 st Simples Shimabukuro-Dias et al. (2005)
Dianema longibarbis Rio Amazonas - AM 60 6 m + 54 a Simples Marcon et al. (1992) D. urostriatum Rio Amazonas - AM 62 6 m + 6 st + 50 a Simples Marcon et al. (1992) D. urostriatum Estação Experimental de Aqüicultura - SP 62 8 m + 4 sm + 4st + 46a Simples Oliveira et al. (1993)
Lepthoplosternum pectorale Alto Paraná - PR 64 8 m-sm + 56 st-a Múltiplas Presente estudo Hoplosternum littorale Rio Amazonas - AM 60 4 m + 4 sm + 52 a Simples Porto & Feldberg (1992) H. littorale Alto Rio Paraná - PR 60 8 m-sm + 52 st-a Simples Presente Estudo
H. littorale Rio Guaíba - RS 60 6 m + 2 sm + 52 a Simples Shimabukuro-Dias et al. (2005) H. littorale Represa Jurumirim - SP 60 6 m + 2 sm + 52 a Simples Shimabukuro-Dias et al. (2005) H. littorale Afluente do Rio Piraí - MT 60 6 m + 2 sm + 52 a Simples Shimabukuro-Dias et al. (2005) Megalechis thoracata Rio Amazonas - AM 64 8 m + 12 sm + 44 st-a Simples Porto & Feldberg (1992)
M. thoracata Rio Amazonas - AM 66 - - Porto et al. (1992)
M. thoracata Afluente do Rio Itiquira - MT 62 6 m + 2 sm + 54 a Simples Shimabukuro-Dias et al. (2005) M. thoracata Córrego Almoço - AC 62 8 m + 54 a Múltiplas Shimabukuro-Dias et al. (2005)
25
REFERÊNCIAS
Almeida-Toledo, L. F., Foresti, F., Daniel, M. F. Z. & Toledo-Filho, S. A. (2000a). Sex chromosome evolution in fish: the formation of the neo-Y chromosome in
Eigenmannia (Gymnotiformes). Chromosoma 109: 197–200.
Almeida-Toledo, L. F., Foresti, F., Daniel, M. F. Z., Lopes, C. E. & Toledo-Filho, S. A. (2000b). Sex chromosome evolution in fish. II. Second occurrence of an X1X2Y sex chromosome system in Gymnotiformes. Chromosome Research 8: 335– 340.
Alves, A. L., Oliveira, C., Foresti, F. (2005). Comparative cytogenetic analysis of eleven species of subfamilies Neoplecostominae and Hypostominae (Siluriformes: Loricariidae). Genetica 124: 127-136.
Artoni, R. F., Vicari, M. R. & Bertollo, L. A. C. (2000). Citogenética de Peixes Neotropicais: Métodos, Resultados e Perspectivas. PUBLICATIO UEPG: Biological Health Sciences 6(1): 43-60.
Artoni, R. F. & Bertollo, L. A. C. (2001). Trends in the karyotype evolution of Loricariidae fish (Siluriformes). Hereditas 134: 201-210.
Bellafronte, E., Margarido, V. P. & Moreira-Filho, O. (2005). Cytotaxonomy of
Parodon nasus and Parodon tortuosus (Pisces, Characiformes). A case of
synonymy confirmed by cytogenetic analyses. Genetics and Molecular Biology 28(4): 710-716.
Bertollo, L. A. C., Takahashi, C. S. & Moreira-Filho, O. (1978). Cytotaxonomic Considerations on Hoplias lacerdae (Pisces, Erythrinidae). Brazilian Journal of
26
Bertollo, L. A. C., Fontes, M. S., Fenocchio, A. S. & Cano, J. (1997). The X1X2Y sex chromosome system in the fish Hoplias malabaricus. I. G-, C-, and chromosome
replication banding. Chromosome Research 5: 493-499.
Bertollo, L. A .C., Oliveira, C., Molina, W. F., Margarido, V. P., Fontes, M.S., Pastori, M. C., Falcão, J. N. & Fenocchio, A. S. (2004). Chromosome evolution in the erythrinid fish, Erythrinus erythrinus (Teleostei: Characiformes). Heredity 93:
228–233.
Britto, M. (2003). Phylogeny of the subfamily Corydoradinae Hoedeman, 1952 (Siluriformes: Callichthyidae), with a definition of its genera. Proceedings of the Academy of Natural Sciences of Philadelphia 153: 119–154.
Fernandes, C. A. & Martins-Santos, I. C. (2005). Sympatric occurrence of three cytotypes and four morphological types of B chromosomes of Astyanax
scabripinnis (Pisces, Characiformes) in the River Ivaí Basin, state of Paraná,
Brazil. Genetica 124: 301-306.
Ferraris, C. J., Jr. (2007). Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types, pp.110-129. Zootaxa 1418: 628.
Foresti, F., Oliveira, C. & Almeida-Toledo, L. F. (1993). A method for chromosome preparations from large specimens of fishes using in vitro short treatment with colchicine. Experientia. 49: 810-813.
Howell, W. M. & Black, D. A .(1980). Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 6: 1014-1015.
27
do rio Paraná, município de Porto Rico (PR). p. 24. III Simpósio de Citogenética Evolutiva e Aplicada de Peixes Neotropicais, Brasil.
Levan, A., Fredga, K. & Sandberg, A. A. (1964). Nomenclature for centromeric position on chromosomes. Hereditas 52: 201-220.
Marcon, J. L., Feldberg, E., Porto, J. I. R. & Affonso, E. G. (1992). Estudos Fisiológicos e Citogenéticos em peixes do Gênero Dianema (Siluriformes,
Callichthyidae) da Amazônia Central. p. 34. IV Simpósio de Citogenética Evolutiva e Aplicada de Peixes Neotropicais, Brasil.
Margarido, V. P. & Galetti Jr., P. M. (1999). Heterochromatin patterns and karyotype relationships within and between the genera Brycon and Salminus (Pisces,
Characidae). Genetics and Molecular Biology 22(3): 357-361.
Margarido, V. P. & Galetti Jr., P. M. (2000). Amplification of a GC-rich heterochromatin in freshwater fish Leporinus desmotes (Characiformes,
Anostomidae). Genetics and Molecular Biology 23(3): 569-573.
Moreira-Filho, O. & Bertollo, L. A. C. (1991). Astyanax scabripinnis (Pisces,
Characidae): A species complex. Brazilian Journal of Genetics, 14(2): 331-357. Oliveira, C., Almeida-Toledo, L. F., Foresti, F., Britski, H.A. & Toledo-Filho, S. A.
(1990). Comparative cytogenetic analysis of three cytotypes of Corydoras
nattereri (Pisces, Siluriformes, Callichthyidae). Cytologia 55: 21-26.
Oliveira, C., Almeida-Toledo, L. F., Mori, L. & Toledo-Filho, S. A. (1993). Cytogenetic and DNA content in six genera of the family Callichthyidae (Pisces, Siluriformes). Caryologia 46(2-3): 171-188.
Oliveira, C. & Gosztonyi, A. E. (2000). A cytogenetic study of Diplomystes
mesembrinus (Teleostei, Siluriformes Diplomystidae) with a discussion of
28
Pinna, M. C. C. (1998). Phylogenetic relationships of Neotropical Siluriformes (Teleostei: Ostariophysi): historical overview and synthesis of hypotheses. In
Phylogeny and Classification of Neotropical Fishes (Malabarba, L. R., Reis,
R.E., Vari, R. P., Lucena, Z. M. S. & Lucena, C. A. S., eds), pp. 279-330. Porto Alegre: Edipucrs.
Porto, J. I. R., Feldberg, E., Nakayama, C. & Falcão, J. N. (1992). A checklist of chromosome number and karyotypes of Amazonian freshwater fishes. Revue D’Hydrobiologie Tropicale. 25: 287-299.
Porto, J. I. R. & Feldberg, E. (1992). Comparative cytogenetic study of the armored catfishes of the genus Hoplosternum (Siluriformes, Callichthyidae). Brazilian
Journal of Genetics 15(2): 359-367.
Porto, J. I. R. & Feldberg, E. (1993). Is Calichthys Linné (Ostariophisy, Siluriformes,
Callichthyidae) a monotypic genus? Acta Amazonica 23(2-3): 311-314.
Reis, R. E. (1998). Anatomy and phylogenetic analysis of the neotropical callichthyid catfishes (Ostariophysi, Siluriformes). Zoological Journal Linnean Society 124: 105-168.
Reis, R. E. (2003). Family Callichthyidae (Armored cattfishes). In Check List of the
Freshwater of South and Central America (Reis, R. E., Kullander, S. O &
Ferrais, C. J. Jr, eds), pp.291-309. 1st edn, Porto Alegre: Edipucrs.
Sanchez, S. & Fenocchio, A. S. (1996). Karyotypic studies and cytotaxonomic considerations on Callichthys callichthys (Pisces, Siluroidei) from Argentina.
Cytologia 61: 247-252.
29
Shimabukuro-Dias, C. K., Oliveira, C. & Foresti, F. (2005). Comparative cytogenetic studies in species of the subfamily Callichthyinae (Teleostei: Siluriformes: Callichthyidae). Caryologia 58(2): 102-111.
Silva, E. B. & Margarido, V. P. (2005). An X1X1X2X2/ X1X2Y multiple sex chromosome system in a new species of the genus Gymnotus (Pisces, Gymnotiformes).
Environmental Biology of Fishes 73: 293-297.
Souza, I. L., Galián, J., De La Rúa, P., Bertollo, L. A. C. & Moreira-Filho, O. (2001). Non-random distribution of the GC-rich heterochromatin and nucleolar rDNA sites on Astyanax scabripinnis chromosomes. Cytologia 66: 85-91.
Sumner, A. T. (1972). A simple technique for demonstrating centromeric heterocromatin. Experimental Cell Research75: 304-306.
Turner, B.J., Diffoot, N. & Rasch, E. M. (1992). The callichthyid catfish Corydoras
aeneus is an unresolved diploid-tetraploid sibling complex. Ichthyologycal