Replacement of wheat bran with spineless cactus in sugarcane-based diets
for steers
Maria Gabriela da Conceição1, Marcelo de Andrade Ferreira1, José Maurício de Souza Campos2, Janaina de Lima Silva1, Edenio Detmann2, Michelle Christina Bernardo de Siqueira1,
Leonardo José Assis de Barros1, Cleber Thiago Ferreira Costa3
1 Universidade Federal Rural de Pernambuco, Departamento de Zootecnia, Recife, PE, Brazil. 2 Universidade Federal de Viçosa, Departamento de Zootecnia, Viçosa, MG, Brazil.
3 Instituto Federal do Sertão Pernambucano, Floresta, PE, Brazil.
ABSTRACT - The effect of replacing wheat bran with spineless cactus (0, 25, 50, 75, and 100%) in sugar cane-based diets on the intake of nutrients, feeding behavior, and rumen digestibility of steers was assessed using digested samples collected from the omasum. Five crossbred steers (½ Holstein-Zebu) were fitted with cannulas in the rumen and then assigned to a 5 × 5 Latin square design. Samples of omasal digesta were collected for three days. Rumen was evacuated via the rumen cannula to determine the rumen pool of dry matter (DM), neutral detergent fiber (NDF), indigestible NDF (iNDF), intake (ki), passage (kp) and degradation (NDF) rates, and the passage of iNDF (kpi). Nutrient intake displayed a quadratic effect, with maximum intakes of DM (5.73 kg d−1), non-fiber carbohydrates (NFC, 4.63 kg d−1), and digestible organic matter (DOM, 3.45 kg d−1)
estimated at the replacement levels of 54.63, 60.00, and 43.17%, respectively. The replacement levels showed no effect on feeding, rumination, or idle times. There was a linear increase in the total apparent and ruminal digestibility of CP as the spineless cactus content in the diet was increased. There was no effect on total and partial digestibilities of OM and NDF or on intestinal digestibility of CP. No effect was observed on the rumen pool of DM, NDF, or iNDF. Furthermore, ki, kp, NDF degradation, and iNDF passage rates displayed a quadratic effect, with higher concentrations estimated for replacement levels of 56.07, 56.12, 59.00, and 55.20%, respectively. Replacing 55% wheat bran with spineless cactus in sugar cane-based diets is recommended.
Key Words: degradation rate, digestibility, fiber, passage rate, rumination
ISSN 1806-9290
www.sbz.org.br R. Bras. Zootec., 45(4):158-164, 2016
Received September 22, 2015 and accepted February 5, 2016. Corresponding author: [email protected]
http://dx.doi.org/10.1590/S1806-92902016000400003
Copyright © 2016 Sociedade Brasileira de Zootecnia. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
The semi-arid region of Northeastern Brazil is characterized by frequent droughts, an average annual rainfall lower than 800 mm, high temperatures (average 27 °C), and a Caatinga biome (SUDENE, 2011). The animal production in this region is impaired during severe droughts when there is a reduction of forage resources, requiring the supply of concentrate feed, thus increasing the cost of production systems.
The use of alternative feed sources can lower production costs. Wheat bran, a wheat processing by-product, is a viable option (Wesendonck et al., 2013) due to the availability of the product. However, being a feed rich in insoluble fiber (Esposito et al., 2005), its use at high concentrations could interfere with voluntary intake (Mariz et al., 2013) as a
result of an increased retention in the rumen and lower passage rates. In addition, the exclusive use of wheat bran tends to raise feed costs, since it is imported from large commercial centers.
The replacement of wheat bran with a local feed such as the spineless cactus could be a potential solution toward a more sustainable production system that is less dependent on grains. The main attributes that justify the importance of spineless cactus are its high non-fibrous carbohydrates (640-710 g kg−1 dry matter) and water (850-900 g kg−1) contents
(Nefzaoui and Ben Salem, 2002), which are essential in areas affected by severe drought. However, spineless cactus is often viewed negatively because it contains reduced amounts of crude protein (40-50 g kg−1 dry matter) and
neutral detergent fiber (200-300 g kg−1 dry matter) (Ferreira
When replacing a concentrate feed with an alternative feed, it is important to examine the effects on performance, evaluating different nutrient digestion sites and fiber digestion rates (Mariz et al., 2013).
Thus, the aim of this study was to evaluate the intake, feeding behavior, digestibility, and rumen dynamics of fiber in crossbred steers fed diets that contain different levels of replacement of wheat bran with spineless cactus.
Material and Methods
All procedures involving animals were approved by the Brazilian Committee for Animal Care and Experimentation (License no. 009/2015) (Brasil, 2000).
This study was carried out in Recife, Pernambuco State, Brazil.
Five rumen-fistulated crossbred steers (Holstein × Zebu) with an average initial body weight (BW) of 160.0±5.3 kg were used in the experiment, distributed in a 5 × 5 Latin square design. The experiment lasted 80 days, corresponding to five 16-day periods. The first seven days were allocated to the adaptation of the animals to the experimental diets, followed by nine days of sample collections. The animals were weighed, identified, and dewormed prior to the experiment and housed in individual pens equipped with feeders and drinkers.
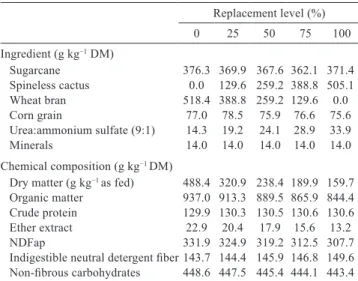
A total of five diets were formulated containing 38:62, 49:51, 63:37, 75:25, and 87:13 forage:concentrate ratios based on dry matter (DM) and containing fresh sugarcane (Saccharum officinarum L.), fresh spineless cactus (Nopalea cochenillifera Salm-Dyck), corn grain, wheat bran, and urea. Dietary treatments consisted of the replacement of the total of wheat bran in basal diet by spineless cactus using the following proportions: 0% for basal diet, and 25, 50, 75, and 100% spineless cactus replacing wheat bran. Urea was added to the diets to adjust the crude protein (CP) content to 130 g kg−1 DM.
The fresh sugar cane and spineless cactus were cut and chopped daily and then provided to animals. The DM content of sugarcane and spineless cactus was evaluated weekly to adjust the amount of feed allowed to the animals.
The diets were supplied ad libitum, allowing approximately 100 g kg−1 as orts. The animals were fed
twice daily in equal portions at 06.00 h and 18.00 h. Forage offered and orts were sampled daily during the collection period and oven-dried (60 ºC). The ingredients that composed the concentrate were sampled from the 8th to the 10th day of every period. All samples were processed in a Wiley mill to pass through a 2-mm screen sieve. After that, each sample was homogenized and divided into two
portions. Half of each sample was processed again in the same mill to pass through a 1-mm screen sieve.
Omasal digesta were collected six times at 12-h intervals between day 11 and day 13. On day 11, samples were collected at 10.00 h and 22.00 h; on day 12, at 08.00 h and 20.00 h; and on day 13, at 06.00 h and 18:00 h. At these same sample collection times, approximately 200 g of feces were collected from the rectum of each animal. Fecal samples were frozen (−20 ºC) for subsequent analysis.
Briefly, to collect the omasal digesta, the technique reported by Huhtanen et al. (1997) was adapted as follows: omasal digesta were collected by introducing the end of a collection tube into the rumen and passing it into the reticulum-omasal orifice until the first part of the tube passed into the orifice, where it was secured by hand during the collection period. The other end of the collection tube was fitted to one of the openings of a kitassato flask, and a vacuum pump was attached to the other opening. The vacuum pump was subsequently turned on to begin collection, and the digesta was collected through the tube via suction until it reached the kitassato flask. Approximately 800 mL of digesta were obtained per collection and frozen (−20º C).
To estimate the omasal flow, a double-marker system was employed in which cobalt-EDTA (6 g/day divided in four doses were infused in rumen of each animal at 00.00 h, 06.00 h, 12.00 h, and 18.00 h, beginning three days before omasal digesta sampling) was used as the liquid phase and a small particle marker and indigestible neutral detergent fiber (iNDF) were used as particulate phase.
After collection, the omasal digesta sample was frozen (−20 ºC) until processing. At the end of each experimental period, the samples were thawed at room temperature and filtered through a 100-µm nylon filter with pores covering 44% of the surface to generate two phases: the filtrate, which corresponded to the liquid phase and small particles, and the residue, which corresponded to the large particle phase. Subsequently, these samples were oven-dried (60 ºC) and ground as previously described for forage and ort samples.
separate the solid and liquid phases. A representative sample of both phases was collected and frozen (−20 ºC) for later evaluation of DM, neutral detergent fiber (NDF), and iNDF contents. After sampling, the phases were again mixed, and the remaining digesta was returned to the rumen.
At the end of the sampling period, the fecal, omasal digesta, and ruminal content samples were oven-dried (60 ºC) and ground as previously described for forage and ort samples. At the end of each period, a composite sample was prepared from the orts, feces, and omasal digesta based on the air-dried weight of the samples for each animal, and the samples were then properly identified and stored in plastic containers for further analysis.
Feeding behavior of all steers was evaluated five times during the experiment during two consecutive days at the seventh day of each adaptation period. Steers were observed every 5 min for 24 h each day starting immediately after the morning feeding, totaling 72 h of observation (three days of observation in each period). The activity of each steer was recorded as rumination, feeding, and idling. Each observation was assumed to last 5 min, which was the interval between observations. A meal was defined as at least one observation of feeding activity occurring after at least 20 min without feeding activity. This is based on the definition of feeding used by Wangsness et al. (1976), who defined a meal as at least 1 min of eating activity after at least 20 min without eating activity. A period of rumination was defined as at least 5 min of rumination occurring after at least 5 min without ruminating activity.
The feed and rumination efficiencies (kg h−1) of DM
and NDF, were calculated by dividing the intake of each of these nutrients by the total feeding time (feed efficiency) or rumination time (rumination efficiency).
Samples of feeds, orts, feces, ruminal content, and omasal digesta were analyzed for DM content by oven-drying at 105 ºC for 16 h (Tables 1 and 2). Samples were also analyzed for ash, ether extract (EE), and CP contents according to the methods of AOAC (2000). Analysis of NDF
followed the method described by Mertens (2002), using heat-stable alpha amylase, without using sodium sulfite, and corrected for residual ash. The NDF was also corrected for the nitrogenous compounds contents by using the method described by Licitra et al. (1996). The omasal digesta sample was analyzed to estimate cobalt concentrations using an atomic absorption spectrophotometer. All of these chemical analyses were performed in samples processed to pass through a 1-mm screen sieve.
Non-fibrous carbohydrates (NFC) contents were quantified according to Detmann and Valadares Filho (2010) as follows:
NFC = 1000 − [(CP − CPu + U) + NDFap + EE + MM], in which CPu = CP content from urea; U = urea content; NDFap = NDF corrected for residual ash and protein; and MM = mineral matter. The other terms were previously defined and all of them are expressed as g kg−1 DM.
The iNDF content was analyzed using the fecal, omasal digesta, feed, and ort samples processed through a 2-mm screen sieve, by using a 288 h rumen incubation procedure as described by Valente et al. (2011).
The fecal excretion was estimated using the iNDF as internal marker. The omasal flow, on the other hand, was calculated as described by France and Siddons (1986) using a double marker system in which cobalt-EDTA was used as the liquid phase and small particles marker and iNDF was used as the large particles marker.
The rates of intake (ki), passage (kp), and degradation of NDF (kd) and iNDF (kpi) were calculated by dividing the daily intake and omasal flow by their respective rumen pools (Allen and Linton, 2007).
The experiment was analyzed according to a 5 × 5 Latin square design using the MIXED procedure of SAS
Replacement level (%)
0 25 50 75 100
Ingredient (g kg−1 DM)
Sugarcane 376.3 369.9 367.6 362.1 371.4
Spineless cactus 0.0 129.6 259.2 388.8 505.1
Wheat bran 518.4 388.8 259.2 129.6 0.0
Corn grain 77.0 78.5 75.9 76.6 75.6
Urea:ammonium sulfate (9:1) 14.3 19.2 24.1 28.9 33.9
Minerals 14.0 14.0 14.0 14.0 14.0
Chemical composition (g kg−1 DM)
Dry matter (g kg−1 as fed) 488.4 320.9 238.4 189.9 159.7
Organic matter 937.0 913.3 889.5 865.9 844.4 Crude protein 129.9 130.3 130.5 130.6 130.6
Ether extract 22.9 20.4 17.9 15.6 13.2
NDFap 331.9 324.9 319.2 312.5 307.7
Indigestible neutral detergent fiber 143.7 144.4 145.9 146.8 149.6 Non-fibrous carbohydrates 448.6 447.5 445.4 444.1 443.4 DM - dry matter; NDFap - neutral detergent fiber corrected for ash and protein.
Table 2 - Ingredients and chemical composition of diets
Table 1 - Chemical composition of feedstuffs
Parameter
Ingredient (g kg−1 DM)
Spineless
cactus Sugarcane Wheat bran Corn Dry matter (g kg−1 as fed) 105.5 282.5 867.3 864.8
Organic matter 802.5 980.2 949.0 986.2
Crude protein 55.5 18.7 152.4 77.0
Ether extract 12.1 11.2 29.8 39.3
NDFap 291.6 410.1 326.6 107.9
(Statistical Analysis System, version 9.4) according to the following model:
Yijk = µ + Di + aj + pk + εijk,
in which µ = overall constant; Di = fixed effect of diet i (i = 1,
2, 3, 4, 5); aj = random effect of animal j (j = 1, 2, 3, 4, 5);
pk = random effect of experimental period k (k = 1, 2, 3, 4, 5);
and εijk = unobserved random error.
After analysis of variance, the significance of the linear and quadratic effects of the replacement of the total of wheat bran in basal diet by spineless cactus was evaluated. A significance value of 0.05 was adopted as the critical value of the probability of type-I error.
Results
A quadratic effect (P<0.05) was observed for the voluntary intake of DM and other components (Table 3). The quadratic pattern can be described by an increased intake up to 50% replacement of wheat bran with spineless cactus. At higher replacement levels, the voluntary intake decreased.
The times spent feeding, ruminating, and idle were not affected (P<0.05) by the replacement levels, with average results of 310.9, 443.9, and 684.6 min d−1 (Table 4).
However, there was a quadratic effect (P<0.05) for feeding and rumination efficiencies of DM, with maximum and minimum values estimated at 50 and 25% replacement of wheat bran by spineless cactus, respectively. The rumination efficiency of NDF was not affected (P<0.05), averaging 243.6 g NDF h−1.
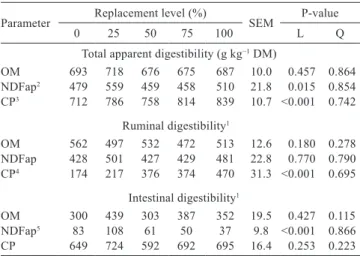
There was a linear increase (P<0.05) in the total apparent and ruminal digestibility of CP as the spineless cactus content in the diet was increased (Table 5). There was no effect (P>0.05) on total and partial digestibilities of OM and NDF and on intestinal digestibility of CP.
No effects of replacement levels on the rumen pools of DM, NDF, and iNDF were observed (P<0.05). Ingestion (ki), passage (kp), and degradation (kd) rates of NDF, and iNDF passage rate (kpi) displayed a quadratic effect (P<0.05), presenting the same pattern observed for voluntary intake, with maximum values found at 50% replacement of wheat bran with spineless cactus.
Table 3 - Mean values for intake (kg d−1) of crossbred steers fed sugarcane-based diets containing different levels of replacement of wheat bran with spineless cactus
Parameter Replacement level (%) SEM P-value
0 25 50 75 100 L Q
DM1 4.08 5.47 5.88 5.12 4.84 0.39 0.145 <0.001
DM (g kg−1 BW)2 23.7 30.1 32.9 27.6 26.3 1.5 0.563 0.001
OM3 4.03 4.96 5.30 4.55 4.28 0.18 0.905 0.002
CP4 0.51 0.74 0.82 0.77 0.73 0.05 0.001 <0.001
NDFap5 1.27 1.63 1.73 1.53 1.44 0.08 0.355 0.001
NFC6 1.98 2.60 2.73 2.30 2.12 0.15 0.966 <0.001
DOM7 2.79 3.56 3.58 3.07 2.94 0.16 0.783 <0.001
SEM - standard error of the mean; L - linear; Q - quadratic.
DM - dry matter; BW - body weight; OM - organic matter; CP - crude protein; NDFap - neutral detergent fiber corrected for ash and protein; NFC - non-fibrous carbohydrates; DOM - digestible organic matter.
1 Ŷ = 4.19796 + 0.05620X – 0.00051439X2. 2 Ŷ = 24.2377 + 0.2778X – 0.0026669X2. 3 Ŷ = 4.10992 + 0.03997 – 0.00039602X2. 4 Ŷ = 0.52843 + 0.00932X – 0.00007495X2. 5 Ŷ = 1.29813 + 0.01469X – 0.00013721X2. 6 Ŷ = 1.96503 + 0.0409X – 0.00084151X2. 7 Ŷ = 2.77337 + 0.03823X – 0.00038976X2.
SEM - standard error of the mean; L - linear; Q - quadratic. DM - dry matter; NDF - neutral detergent fiber.
1 Ŷ = 806.2 + 12.45X – 0.116X2. 2 Ŷ = 632.2 + 5.177X – 0.052X2.
Table 4 - Mean values for feeding behavior in crossbred steers fed sugarcane-based diets containing different levels of replacement of wheat bran with spineless cactus
Parameter Replacement level (%) SEM P-value
0 25 50 75 100 L Q
Feeding time (min d−1) 314 316 290 316 320 2.70 0.822 0.388
Idle time (min d−1) 710 716 662 668 668 5.19 0.225 0.703
Rumination time (min d−1) 416 408 488 456 452 6.49 0.190 0.331
Feed efficiency (g DM h−1)1 790 1040 1250 940 940 33.92 0.380 0.003
Rumination efficiency (g DM h−1)2 600 800 740 680 650 15.59 0.787 0.012
Discussion
The observed increase in intake up to the level of 50% replacement (Table 3) is due to the inclusion of spineless cactus, which has a better ruminal degradation pattern of fiber compared with wheat bran and sugarcane (Lins, 2014). This faster degradation helps to increase the digesta flow to the gastrointestinal tract, consequently increasing feed intake. This pattern can be also supported by the increased degradation rate of fiber up to the level of 50% replacement (Table 6). In spite of this, the ruminal digestibility was not modified by spineless cactus in the diets (Table 5). However, it must be taken into account that both fiber degradation and passage were similarly modified. From this, it can be stated that ruminal NDF digestibility was not increased because the residence time in the rumen was decreased. Such simultaneous increase in both rates may leave NDF unchanged (Detmann et al., 2009). However, the increase in degradation and passage rates up to the level of 50% replacement brings into evidence that improvements in the rumen dynamics of fiber were observed when spineless cactus was included in the diet. On the other hand, the fiber quality of the feed used in the diet directly influences the voluntary intake. Both wheat bran and sugar cane are foods with low fiber degradation, and the use of these components at high concentrations can reduce voluntary intake (Freitas et al.,
2008; Mariz et al., 2013). The treatment without spineless cactus provided the lowest voluntary intake (Table 3), and this was likely due to the larger amount of wheat bran present in the diet, in addition to sugar cane, which was used as a staple feed in the treatments (Table 3).
Another possible factor to positively influence voluntary intake up to the level of 50% replacement is the form of feed processing. According to Ferreira et al. (2011), spineless cactus processed in fodder machines may encourage intake, since other feeds, such as urea and wheat bran particles, may adhere to exposed mucilage, thus avoiding selection by animals.
The decrease in nutrient intake followed the decrease in the passage rate at levels above 50% of replacement, since DM intake showed a higher value at the level of 50%. It is important to note that under normal feeding conditions (without a drastic nutritional unbalance), passage rate is regulated by intake (Pittroff and Kothmann, 1999), which can explain the similar quadratic pattern for voluntary intake (Table 3) and fiber passage rate (Table 6). Therefore, the rumen fill effect of the fiber will be determined mainly by the degradation rate and by the potentially digestible NDF to iNDF ratio (Detmann et al., 2014).
The probable cause of the negative effect of the higher levels of spineless cactus in the diet on voluntary intake is the high moisture content in this feed, which increases its capacity to occupy space in the rumen environment, as reported by other authors (Gebremarian et al., 2006). It must be noted that no effect with regard to the DM and NDF pool in the rumen was detected here (Table 6). The DM content of
Table 5 - Mean values for the total, ruminal and intestinal digestibility of crossbred steers fed sugarcane-based diets containing different levels of replacement of wheat bran with spineless cactus
Parameter Replacement level (%) SEM P-value
0 25 50 75 100 L Q
Total apparent digestibility (g kg−1 DM)
OM 693 718 676 675 687 10.0 0.457 0.864
NDFap2 479 559 459 458 510 21.8 0.015 0.854
CP3 712 786 758 814 839 10.7 <0.001 0.742
Ruminal digestibility1
OM 562 497 532 472 513 12.6 0.180 0.278
NDFap 428 501 427 429 481 22.8 0.770 0.790 CP4 174 217 376 374 470 31.3 <0.001 0.695
Intestinal digestibility1
OM 300 439 303 387 352 19.5 0.427 0.115
NDFap5 83 108 61 50 37 9.8 <0.001 0.866
CP 649 724 592 692 695 16.4 0.253 0.223
SEM - standard error of the mean; L - linear; Q - quadratic.
DM - dry matter; OM - organic matter; NDFap - neutral detergent fiber corrected for ash and protein; CP - crude protein.
1 Expressed as g kg−1 of the quantity that reaches each digestion site. 2 Ŷ = 500.8 – 0.156X.
3 Ŷ = 725.4 + 1.128X. 4 Ŷ = 172.4 + 2.996X. 5 Ŷ = 97.8 – 0.606X.
Table 6 - Mean values for the ruminal pool and intake, passage, and degradation rates in crossbred steers fed sugarcane-based diets containing different levels of replacement of wheat bran with spineless cactus
Parameter Replacement level (%) SEM P-value
0 25 50 75 100 L Q
Ruminal pool (kg)
DM 2.45 1.97 1.95 1.97 2.12 0.087 0.309 0.096 NDF 1.77 1.35 1.33 1.38 1.44 0.066 0.199 0.075 iNDF 0.98 0.81 0.81 0.89 0.95 0.038 0.938 0.123 (h−1)
ki1 0.0305 0.0505 0.0543 0.0493 0.0431 0.002 0.058 0.001
kp2 0.0184 0.0286 0.0294 0.0288 0.0214 0.001 0.332 <0.001
kd3 0.0121 0.0219 0.0248 0.0205 0.0216 0.001 0.035 0.023
kpi4 0.0245 0.0390 0.0403 0.0359 0.0311 0.002 0.037 0.001 SEM - standard error of the mean; L - linear; Q - quadratic.
DM dry matter; NDFap neutral detergent fiber corrected for ash and protein; iNDF -indigestible neutral detergent fiber; ki - NDFap intake rate; kp - NDFap passage rate; kd - NDFap degradation rate; kpi - iNDF passage rate.
spineless cactus was 105.5 g kg−1 (Table 1). According to
Minson (1990), water contents in forages exceeding 780 g kg−1
can compromise voluntary intake, which agrees with our previous statement.
The assessed behavioral variables showed no influence of the replacement of wheat bran with spineless cactus (Table 4). Rumination activity is stimulated by the presence of physically effective fiber (Colenbrander et al., 1991).Therefore, the lack of an effect from this parameter may be because sugar cane levels were similar for all treatments, thus showing the lack of effectiveness of the spineless cactus fiber. On the other hand, the quadratic behavior observed for feed and rumination efficiency (mass per unit of time) of DM seems to reflect the patterns of voluntary intake (Table 3) and intake rate (Table 6). Considering that no difference among treatments was observed for feeding and rumination times (Table 4), the feed and rumination efficiencies should be proportional to intake.
Sugar cane causes an increase in the retention time of the feed in the rumen, and this influences the increase in digestibility, because the feed will have a longer exposure to microorganisms, increasing the degradation of this material. The levels of sugar cane used in the treatments were similar, which may have contributed to the lack of an effect on total OM digestibility (Table 5).
The total and ruminal protein digestibility increased as more spineless cactus and urea were included in the diet (Table 5). The degradation of crude protein can be influenced by the ratio of non-protein nitrogen (NPN-urea) to true protein (wheat bran). Non-protein nitrogen is easily degraded and absorbed, while true protein has peptide bonds that hinder microbial access to protein molecules, making them more resistant to microbial degradation (Barbosa et al., 2012). Particularly, the observed increase in rumen digestibility of CP was due to the replacement of the true protein from wheat bran by urea (NPN). Urea is a non-protein nitrogen source with rapid and complete degradation in the rumen (Eustáquio Filho et al., 2008) that may have contributed to the increase in rumen digestibility of CP. Therefore, the amount of urea consumed increased as wheat bran was replaced with spineless cactus.
Conclusions
Up to 50% wheat bran can be replaced with spineless cactus in the diet of crossbred steers to increase voluntary intake. Moreover, it may also benefit the rumen dynamics of the fiber.
Acknowledgments
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq.
References
Allen, M. S. and Linton, J.A. V. 2007. In vivo methods to measure digestibility and digestion kinetics of feed fractions in the rumen. p.72-89. In: Proceedings of the Simpósio Internacional Avanços em Técnicas de Pesquisa em Nutrição de Ruminantes. Rennó, F. P.; Silva, L. F. P., eds. Pirassununga, SP, Brasil.
AOAC - Association of Official Analytical Chemists. 2000. Official methods of analysis. Arlington, VA.
Barbosa, J. G.; Costa, R. G.; Medeiros, A. N.; Queiroga, R. C. R. E.; Batista, A. M. V.; Medeiros, G. R. and Beltrão Filho, E. M. 2012. Use of different urea levels in the feeding of Alpine goats. Revista Brasileira de Zootecnia 41:1713-1719.
Brasil. 2000. Ministério da Agricultura, Pecuária e do Abastecimento (MAPA). Instrução Normativa n. 3, de 17 de janeiro de 2000. Lex: Diário Oficial da União de 24 de janeiro de 2000, Seção 1, p.14-16.
Colenbrander, V. F.; Noller, C. H. and Grant, R. J. 1991. Effect of fiber content and particle size of alfafa silage on performance and chewing behavior. Journal of Dairy Science 74:2681-2690. Detmann, E.; Paulino, M. F.; Mantovani, H. C.; Valadares Filho, S. C.;
Sampaio, C. B.; Souza, M. A.; Lazzarini, I. and Detmann, K. S. C. 2009. Parameterization of ruminal fibre degradation in low- quality tropical forage using Michaelis - Menten kinetics. Livestock Science 126:136-146.
Detmann, E. and Valadares Filho. S. C. 2010. On the estimation of non-fibrous carbohydrates in feeds and diets. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 62:980-984.
Detmann, E.; Gionbelli, M. P. and Huhtanen, P. 2014. A meta-analytical evaluation of the regulation of voluntary intake in cattle fed tropical forage-based diets. Journal of Animal Science 92:4632-4641.
Esposito, F.; Arlotti, G.; Bonifati, G. M.; Napolitano, A.; Vitale, D. and Fogliano, V. 2005. Antioxidant activity and dietary fibre in durum wheat bran by-products. Food Research International 38:1167-1173.
Eustáquio Filho, A.; Santos, P. E. F. and Yamamoto, S. M. 2008. Utilização de ureia como fonte de nitrogênio não proteico (NNP) para ruminantes. Pubvet 2:32.
Ferreira, M. A.; Silva, F. M.; Bispo, S. V. and Azevedo, M. 2009. Estratégias na suplementação de vacas leiteiras no semi-árido do Brasil. Revista Brasileira de Zootecnia 38:322-329.
Ferreira, M. A.; Pessoa, R.A.S.; Silva, F. M. and Bispo, S. V. 2011. Palma forrageira e ureia na alimentação de vacas leiteiras. Editora UFRPE, Recife.
France, J. and Siddons, R. C. 1986. Determination of digesta flow by continuous marker infusion. Journal of Theoretical Biology 121:105-119.
Freitas, A. W. P.; Rocha, F. C.; Zonta, A.; Fagundes, J. L.; Fonseca, R.; Zonta, M. C. M. and Macedo, F. L. 2008. Consumo de nutrientes e desempenho de ovinos alimentados com dietas à base de cana-de-açúcar hidrolisada. Pesquisa Agropecuária Brasileira 43:1569-1574. Gebremarian, T.; Melaku, S. and Yami, A. 2006. Effect of different
Huhtanen, P.; Brotz, P. G. and Satter, L. D. 1997. Omasal sampling technique for assessing fermentative digestion in the for estomach of dairy cows. Journal of Animal Science 75:1380-1392.
Licitra, G.; Hernandes, T. M. and Van Soest, P. J. 1996. Standardization of procedures for nitrogen fractionation of ruminant feeds. Animal Feed Science and Technology 57:347-358.
Lins, S. E. B. 2014. Palma com ureia em dietas à base de cana-de-açúcar corrigida para ovinos fistulados no rúmen. Dissertação (M.Sc.). Universidade Federal Rural de Pernambuco, Recife. Mariz, L. D. S.; Valadares Filho, S. C.; Detmann, E.; Pereira, O. G.;
Pereira, L. G. R.; Marcondes, M. I.; Santos, S. A.; Villadiego, F. A. C.; Zanetti, D.; Prados, L. F. and Nunes, A. N. 2013. Intake and ruminal digestion determined using omasal and reticular digesta samples in cattle fed diets containing sugar cane in natura or ensiled sugar cane compared with maize silage. Livestock Science 155:71-76.
Mertens, D. R. 2002. Gravimetric determination of amylase treated neutral detergent fiber in feeds with refluxing in beaker or crucibles: collaborative study. Journal of AOAC International 85:1217-1240. Millen, D. D.; Pacheco, R.D. L.; Arrigoni, M. D. B.; Galyean, M. L.
and Vasconcelos, J. T. 2009. A snapshot of management practices and nutritional recommendations used by feedlot nutritionists in Brazil. Journal of Animal Science 87:3427-3439.
Minson, D. J. 1990. Forage in ruminant nutrition. Academic Press, San Diego, CA.
Nefzaoui, A. and Ben Salem, H. 2002. Forage, fodder, and animal nutrition. p.199-210. In: Cacti: Biology and uses. Nobel, P. S., ed. University of California Press, Berkeley and Los Angeles, CA. Pittroff, W. and Kothmann, M. M. 1999. Regulation of intake and
diet selection by herbivores. p.366-422. In: Nutritional ecology of herbivores. American Society of Animal Science, Savoy, Illinois.
SUDENE - Superintendência do Desenvolvimento do Nordeste. 2011. O semiárido nordestino. Available at: <http://www.sudene.gov.br/ site/extra.php>. Accessed on: Apr. 30, 2012.
Valente, T. N. P.; Detmann, E.; Valadares Filho, S. C.; Cunha, M.; Queiroz, A. C. and Sampaio, C. B. 2011. In situ estimation of indigestible compounds contents in cattle feed and feces using bags made from different textiles. Revista Brasileira de Zootecnia 40:666-675.
Wangsness, P. J.; Chase, L. E.; Peterson, A. D.; Hartsock, T. G.; Kellmel, D. J. and Baumgardt, B. R. 1976. System for monitoring feeding behavior of sheep. Journal of Animal Science 42:1544-1549.