Original Contribution
ANTHOCYANIN-RICH EXTRACT DECREASES INDICES OF LIPID
PEROXIDATION AND DNA DAMAGE IN VITAMIN E-DEPLETED RATS
CARMENRAMIREZ-TORTOSA,*†ØYVINDM. ANDERSEN,‡PETERT. GARDNER,* PHILIPC. MORRICE,* SHARON G. WOOD,* SUSANJ. DUTHIE,* ANDREW R. COLLINS,*andGARRYG. DUTHIE*
*Antioxidants and DNA Damage Group, Rowett Research Institute, Aberdeen, Scotland, UK;†Instituto de Nutricion y Tecnologı´a
de Alimentos, Departamento de Bioquimica y Biologia Molecular, Universidad de Granada, Granada, Spain; and‡Department of
Chemistry, University of Bergen, Bergen, Norway
(Received21February2001;Accepted22May2001)
Abstract—Anthocyanins are secondary plant metabolites responsible for the blue, purple, and red color of many plant tissues. The phenolic structure of anthocyanins conveys marked antioxidant activity in model systems via donation of electrons or hydrogen atoms from hydroxyl moieties to free radicals. Dietary intakes of anthocyanins may exceed 200 mg/day, however, little is known about their antioxidant potency in vivo. Consequently, the aim of this study was to establish whether anthocyanins could act as putative antioxidant micronutrients. Rats were maintained on vitamin E-deficient diets for 12 weeks in order to enhance susceptibility to oxidative damage and then repleted with rations containing a highly purified anthocyanin-rich extract at a concentration of 1 g/kg diet. The extract consisted of the 3-glucopyranoside forms of delphinidin, cyanidin, petunidin, peonidin, and malvidin. Consumption of the anthocyanin-repleted diet significantly improved (p⬍ .01) plasma antioxidant capacity and decreased (p⬍.001) the vitamin E deficiency-enhanced hydroperoxides and 8-Oxo-deoxyguanosine concentrations in liver. These compounds are indices of lipid peroxidation and DNA damage, respectively. Dietary consumption of anthocyanin-rich foods may contribute to overall antioxidant status, particularly in areas of habitually low vitamin E intake. © 2001 Elsevier Science Inc.
Keywords—Anthocyanins, Polyphenols, Vitamin E deficiency, Lipid peroxides, DNA damage, Antioxidants, Free radicals
INTRODUCTION
Many polyphenolic products of the phenylpropanoid bio-synthetic pathway in plants have considerable antioxi-dant ability in vitro. This is due to the ease with which an H-atom from an aromatic hydroxyl group of polyphenols can be donated to a free radical, and the ability of the aromatic group to then support the unpaired electron via delocalization around the-electron system [1]. In ad-dition, many polyphenols can bind transition metal ions thus preventing the formation of reactive species by Fenton chemistry [2]. There are no known deficiency states resulting from lack of plant polyphenols. There-fore, even though dietary intakes may exceed 1 g/day [3], they have generally been regarded as non-nutritive.
However, epidemiological studies inversely relating in-takes of flavonols and flavones with the incidence of heart disease (for recent review, see ref. [4]) may indicate a putative role for certain polyphenols in the prevention of chronic diseases that involve oxidative processes.
Countries in Northern latitudes generally have the greatest number of premature deaths from chronic dis-eases such as coronary heart disease and strokes [5]. This is ascribed, in part, to a habitually low consumption of foods rich in antioxidant micronutrients [6]. For exam-ple, in such countries more than 95% of the population may not achieve the U.S. recommended daily allowance for vitamin E [7]. This may be due to traditional dietary patterns and the expense and lack of availability of fresh fruit and vegetables. For such populations, a potentially important source of antioxidant-rich food may be locally grown soft fruits (e.g., raspberries, blueberries, cranber-ries, and blackcurrent). These are rich in anthocyanins, which are glycosidic-linked flavonoids responsible for Address correspondence to: Dr. Garry G. Duthie, Rowett Research
Institute, Greenburn Road, Bucksburn, Aberdeen, AB21 9SB, UK; Tel: ⫹44 (1224) 716-623; Fax:⫹44 (1224) 716-622; E-Mail: ggd@rri. sari.ac.uk.
Free Radical Biology & Medicine, Vol. 31, No. 9, pp. 1033–1037, 2001 Copyright © 2001 Elsevier Science Inc. Printed in the USA. All rights reserved 0891-5849/01/$–see front matter
PII S0891-5849(01)00618-9
the red, violet, purple, and blue colors of many plants [8]. As with other plant polyphenols, many anthocyanins have marked antioxidant activity in vitro [9,10]. Some anthocyanins are bioavailable, as they have been de-tected in human plasma, however, little is known about their potential antioxidant properties in vivo [11,12]. Consequently, in order to ascertain whether anthocyanins can act as putative antioxidant nutrients, we have re-pleted rats previously maintained on vitamin E deficient rations with either a highly purified anthocyanin extract or d␣-tocopherol. The aim was to assess the ability of the anthocyanin extract to modify the elevated indices of oxidative damage induced by vitamin E deficiency.
MATERIALS AND METHODS
Anthocyanin extract
To prepare the anthocyanin extract, the scraped sur-face material from the cones ofAbies koreana (800 g) was extracted with 1l of 90% MeOH containing 0.1% HCl 0.5 M citric acid. The procedure was repeated five times and the combined extracts were then filtered, con-centrated under reduced pressure at 25°C, partitioned against ethyl acetate, and applied to an Amberlite XAD-7 column [13]. The column was washed with water and the
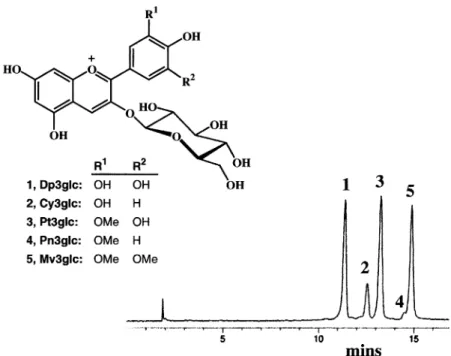
sample was eluted with MeOH 0.1% HCl. The eluant was dried under reduced pressure. The purity of the sample (10.9 g) was checked by HPLC using simulta-neous detection at 520 ⫾ 20 and 280 ⫾ 10 nm (for conditions, see legend of Fig. 1). Both chromatograms revealed only previously identified anthocyanins [14], which were the 3-glucosides of delphinidin, cyanidin, petunidin, peonidin, and malvidin, respectively (Fig. 1), their relative proportions in the extract being 30, 10, 32, 2, and 26%, respectively.
Animals and diets
Individually housed weanling male rats of the Rowett Hooded Lister strain were offered, ad libitum, a semi-synthetic diet containing less than 0.5 mg/kg vitamin E [15]. After 10 weeks, the rats were then offered the diet containing either d␣-tocopherol or the dried anthocyanin extract at a concentration of 1 g/kg diet for a further 2 weeks (six rats in each group). The extract was finely ground and mixed to an even distribution in the diets. Six rats continued to be offered the vitamin E deficient ration and six the vitamin E adequate ration throughout the trial. Diets were stored at⫺40°C until use.
Rats were anesthetized with halothane and blood re-Fig. 1. HPLC profile of the purified anthocyanin sample fromAbies procera detected at 520 ⫾20 nm. The structures of the anthocyanins are also shown: 1–5. Dp⫽delphinidin; Cy⫽cyanidin; Pt⫽petunidin; Pn⫽peonidin; Mv⫽malvidin and glc⫽ glucose. Measurement was performed on a HP-1050 module system (Hewlett Packard) using an ODS Hypersil column (200⫻4.6 mm, 5m) and diode array detection. The two solvents used for elution were: (A) HCO2H-H2O (1: 9, v/v) and (B) MeOH-HCO2H-H2O
(5: 1: 4, v/v). The elution profile was 0 – 4 min, 10% B in A (isocratic); 4 –21 min, 10 –100% B in A (linear gradient), 100% B (isocratic) for the next 4 min followed by linear gradient back to 10% B in 1 min. The flow rate was 1.2 ml min⫺l, and aliquots of 15l were
moved by cardiac puncture into heparinized evacuated tubes (Becton Dickinson, Oxford, UK). Plasma was ob-tained by centrifugation (2500⫻g, 10 min, 4°C) and red cells were washed twice prior to being resuspended to the original volume in NaCl (0.9%). Livers were per-fused in situ with chilled 0.15 M KCl and hepatic mi-crosomes were prepared by the method of Lim et al. [16]. Plasma and the remaining liver tissue were immediately snap frozen in liquid nitrogen and stored at⫺70°C.
Biochemical analysis
Vitamin E concentrations in plasma and liver were determined by reverse phase HPLC with fluorimetric detection [17] under the auspices of the U.S. National Institute of Standards Quality Assurance Scheme for fat-soluble vitamins. The antioxidant capacity of plasma was estimated by its ability to donate a hydrogen atom or electron to the synthetic free radical, potassium nitroso-disulphonate (Fremy’s salt) detected by electron spin resonance (ESR) spectroscopy [18]. In brief, 50l of a 10-fold dilution of plasma was mixed with an equal volume of Fremy’s salt [0.05 mM in 10 ml phosphate-buffered saline(PBS)] and the spectrum of the low field resonance of the Fremy’s radical was recorded after 5 min. Spectra were obtained at 21°C on a Bruker ECS 106 spectrometer working at ca. 9.5 GHz (X-band frequency) and equipped with a cylindrical (TM110 mode) cavity. The microwave power and modulation amplitude were set at 2 mW and 0.01 mT, respectively. The number of Fremy’s radicals reduced by the plasma was compared with that of a control reaction using PBS instead of diluted plasma. Endogenous hydroperoxide concentra-tions in liver homogenates were determined by the fer-rous orange xylenol method as described in Jiang et al. [19]. DNA damage in the liver was estimated from the amount of 8-Oxo-deoxyguanosine (8-Oxo-dG) present. This was determined by HPLC using electrochemical detection following extraction procedures designed to minimize artifactual oxidation of the DNA [20]. The susceptibility of hepatic microsomal preparations to
ox-idation in vitro was assessed by the production of thio-barbituric acid reactive substances (TBARS) following incubation (4 min, 37°C) with 30 M FeSO4.7H2O,
2mM ADP and 0.5 mM L-ascorbic acid [21]. Suscepti-bility of washed erythrocytes to peroxidation was also estimated by the formation of TBARS following 20 min incubation at 37°C with 0.1 M hydrogen peroxide [21]. Results are presented as mean ⫾ SEM. Data was initially analyzed by one-way analysis of variance and comparison of groups was made using the Bonferroni Multiple Comparison test. Apvalue of less than .05 was considered significant.
RESULTS
Final rat weights in all groups were similar and un-affected by the tocopherol or anthocyanin contents of the diets. Following consumption of vitamin E-deficient di-ets for 10 weeks, plasma and liver d␣-tocopherol con-centrations were below detectable levels (⬍ 0.5g/ml and⬍5g/g protein for plasma and liver, respectively). In the rats repleted for 2 weeks with diet containing d␣-tocopherol, concentrations were similar to those an-imals maintained throughout the trial on the vitamin E-sufficient rations (Table 1). Vitamin E deficiency was also associated with a 30% decrease (p⬍.001) in plasma antioxidant capacity. Repletion with diet containing the anthocyanin-rich extract significantly enhanced (p⬍.01) plasma antioxidant capacity, but to a lesser extent than in those rats repleted with vitamin E (Table 1). Consump-tion of vitamin E deficient raConsump-tions for 10 weeks was also associated with an increase of hydroperoxides (p⬍.001) and 8-Oxo-dG (p⬍ .001) in liver, an effect which was markedly less in rats repleted with either the anthocya-nin-rich extract or d␣-tocopherol (Fig. 2). However, analogous protection against oxidative damage was not observed when microsomes and red cells from rats con-suming the anthocyanin-rich extract were incubated with Fe/ADP and H2O2, respectively. This contrasted with the highly protective effect observed as a result of repletion with d␣-tocopherol (Table 2).
Table 1. Effect of Vitamin E Deficiency and then 2 Weeks Repletion with the Anthocyanin Extract or Vitamin E on Rat Weight, Plasma, and Liver d␣-Tocopherol Concentrations and Plasma Antioxidant Capacity
Parameter
Vitamin E-sufficient diet
Vitamin E-deficient diet
Vitamin E-deficient diet⫹anthocyanin extract
Vitamin E-deficient diet⫹d␣-tocopherol
Weight (g) 382⫾7 389⫾8 394⫾10 398⫾6
Plasma d␣-tocopherol (g/ml) 8.7⫾0.3 nd nd 8.1⫾0.5
Liver d␣-tocopherol (g/g protein) 370⫾34 nd nd 334⫾62
Plasma antioxidant capacity (% radicals reduced)
46⫾2 32⫾2a 38
⫾1c 46
⫾1b
aSignificant effect of vitamin E deficiency,p
⬍.001;bsignificant effect of repletion with d␣-tocopherol,p
⬍.001;csignificant effect of repletion
with anthocyanin,p⬍.01. Data as mean⫾SEM, 6 rats/group. nd⫽not detectable.
DISCUSSION
It has long been known that vitamin E deficiency results in increased indices of lipid peroxidation in blood and tissues of animals. Moreover, repletion with vitamin E can rapidly reverse the biochemical and clinical symp-toms of vitamin E deficiency, provided that the defi-ciency state is not excessively prolonged. However, as far as we can ascertain, this is the first report to demon-strate that vitamin E deficiency also increases oxidative damage to DNA as estimated by concentrations of
8-Oxo-dG, and that basal levels can be subsequently restored by reintroduction of vitamin E to the diet. Such damage to DNA may produce mutations that cause per-manent alterations in the genetic message when the cell replicates its DNA and thus increases risk of carcinogen-esis. These results therefore support recent epidemiolog-ical studies implicating low intakes or blood levels of vitamin E with the subsequent risk of cancers at various sites [22–25].
Consumption of the anthocyanin-rich extract also moderated elevated indices of oxidative DNA damage in liver. Whether this effect is ascribed to free radical scavenging, upregulation of DNA repair mechanisms and antioxidant enzymes, or the formation of protective anthocyanin-DNA co-pigmentation complexes [26] is unclear. However, analogous effects on concentration of hydroperoxides in liver and on plasma antioxidant ca-pacity also suggest that the anthocyanin-rich extract pos-sesses antioxidant activity in vivo. The apparent ability of the strongly polar anthocyanins to substitute for the lipophilic antioxidant, vitamin E, may be analogous to the ability of vitamin C to protect biomembranes from peroxidation by efficiently trapping peroxyl radicals in the cytosol [27]. Alternatively, the antioxidant effect may reflect, in part, the ability of polyphenols, including anthocyanins, to chelate transition metal ions involved in radical-forming processes such as Fenton reactions [28]. Direct intestinal absorption of anthocyanins in the intact glycoside form and subsequent biotransformation to methylated derivatives in the liver has been observed in the rat [11]. This suggests that the anthocyanins in the highly purified extract are also bioavailable. However, tissue concentrations of anthocyanins and related metab-olites were not measured in the present study and there-fore it is unclear which of the compounds in the extract has the most marked antioxidant effect in vivo.
The possibility that the marked in vivo antioxidant effects of this highly-purified extract are due to the presence of other unidentified compounds apart from the anthocyanins is unlikely but cannot be excluded. The HPLC profile (Fig. 1) is detected at 520⫾20 nm. The HPLC profile detected simultaneously at 280⫾ 10 nm Fig. 2. Effect of vitamin E depletion and 2 weeks repletion with the
anthocyanin extract or vitamin E on liver concentrations of 8-Oxo-dG and lipid hydroperoxides.
Table 2. Susceptibility of Red Cells and Hepatic Microsomal Preparations from Rats Maintained on Vitamin E-Deficient Rations, then Repleted for 2 Weeks with the Anthocyanin Extract or Vitamin E or Maintained on a Vitamin E-Sufficient Ration throughout the Trial
Parameter
Vitamin E-sufficient diet
Vitamin E-deficient diet
Vitamin E-deficient diet⫹anthocyanin extract
Vitamin E-deficient diet⫹d␣-tocopherol
Red cells (nmol TBARS/g Hb) 83⫾7 809⫾33a 801
⫾40 79⫾7b
Hepatic microsomes (nmoles TBARS/mg protein)
1.3⫾0.4 25.8⫾0.4a 23.1
⫾0.5 0.9⫾0.2b
aSignificant effect of vitamin E deficiency,p
⬍.001;bsignificant effect of repletion with d␣-tocopherol,p
shows exactly the same pattern. This suggests that there are no compounds present other than the anthocyanins, although there is the possibility that there are compounds which are not absorbing light in the region between 270 and 600 nm. However, the extract did not contain de-tectable amounts of vitamin E, vitamin C, or carotenoids, which are generally recognized as important antioxidant micronutrients.
Unlike vitamin E repletion, consumption of the an-thocyanin-rich extract did not decrease the susceptibility of microsomal preparations or washed red cells to oxi-dize ex vivo. This may indicate that anthocyanins are less strongly intercalated in the cell membranes than vitamin E and were therefore lost during the preparation of the microsomes and red cells. Similar effects have been reported during the preparation of low-density lipopro-tein from plasma of polyphenol-supplemented human volunteers [29], and indicates that in intervention trials important biological effects may be lost through manip-ulation of samples ex vivo.
In conclusion, an anthocyanin-rich extract containing the 3-glucopyranoside forms of delphinidin, cyanidin, petunidin, and malvidin markedly decreased vitamin E deficiency-induced elevations in DNA damage and hy-droperoxides in liver and increased plasma antioxidant capacity. As anthocyanins are widespread in many fruit and vegetable species, both wild and horticultural culti-vars may be an important source of antioxidant phyto-chemicals in populations with habitually low intakes of the recognized antioxidant micronutrients.
Acknowledgements— We are grateful for financial support from the Scottish Executive Rural Affairs Department, the University of Granada, Spain, and the EU Framework V programme.
REFERENCES
[1] Rice-Evans, C. A.; Miller, N. J.; Paganga, G. Structure-antioxi-dant activity relationships of flavonoids and phenolic-acids.Free Radic. Biol. Med.20:933–956; 1996.
[2] Yoshino, M.; Murakami, K. Interaction of iron with polyphenolic compounds: application to antioxidant characterization. Anal. Biochem.257:40 – 44; 1998.
[3] Formica, J. V.; Regelson, W. Review of the biology of quercetin and related bioflavonoids.Food Chem. Toxicol.33:1061–1080; 1995.
[4] Duthie, G. G.; Duthie S. J.; Kyle, J. A. M. Plant polyphenols in cancer and heart disease: implications as nutritional antioxidants.
Nutr. Res. Rev.13:79 –106; 2000.
[5] Bellizzi, M. C.; Franklin, M. F.; Duthie, G. G.; James, W. P. T. Vitamin E and coronary heart disease: the European paradox.Eur. J. Clin. Nutr.48:822– 831; 1994.
[6] Diplock, A. T. Antioxidants and disease prevention.Mol. Aspects Med.15:293–376; 1994.
[7] Duthie, G. G.; Arthur, J. R.; Beattie, J. A. G.; Brown, K. B.; Morrice, P. C.; Robertson, J. D.; Shortt, C. T.; Walker, K. A.; James, W. P. T. Cigarette smoking, antioxidants, lipid peroxida-tion and coronary heart disease.Ann. N. Y. Acad. Sci.686:120 – 129; 1993.
[8] Clifford, M. N. Anthocyanins—nature, occurrence and dietary burden.J. Sci. Food Agric.80:1063–1072; 2000.
[9] Wang, H.; Cao, G. H.; Prior, R. L. Oxygen radical absorbing capacity of anthocyanins.J. Agr. Food. Chem.45:304 –309; 1997. [10] Tsuda, T.; Shiga, K.; Ohshima, K.; Kawakishi, S.; Osawa, T. Inhibition of lipid peroxidation and the active oxygen radical scavenging effect of anthocyanin pigments isolated from Phaseo-lus vulgaris. Biochem. Pharmacol.52:1033–1039; 1996. [11] Miyazawa, T.; Nakagawa, K.; Kudo, M.; Kayo, M.; Someyo, K.
Direct intestinal absorption of red fruit anthocyanins, cyanidin-3-glucoside and cyanidin-3-5-dicyanidin-3-glucoside, into rats and humans.J. Agric. Food Chem.47:1083–1091; 1999.
[12] Cao, G.; Prior, R. L. Anthocyanins are detected in human plasma after oral administration of an elderberry extract.Clin. Chem.
45:574 –576; 1999.
[13] Andersen, Ø. M. Semipreparative isolation and structure deter-mination of pelargonidin 3-O-␣-L-rhamnopyranosyl-(1-2)- -D-glucopyranoside and other anthocyanins from the tree Dacrycar-pus dacrydioides. Acta Chem. Scand.42:462– 468; 1988. [14] Andersen, Ø. M. Anthocyanins from reproductive structures in
Pinaceae.Biochem. Syst. Ecol.20:145–148; 1992.
[15] Abdel-Rahim, A. G.; Arthur, J. R.; Mills, C. F. Effects of dietary copper, cadmium, iron, molybdenum and manganese on selenium utilization by the rat. J. Nutr.116:403– 411; 1986.
[16] Lim, V. S.; Henriquez, H.; Seo, H.; Refetoff, S.; Martino, E. J. Thyroid function in an uremic rat model.J. Clin. Invest.66:946 – 954; 1980.
[17] Hess, D.; Keller, H. E.; Oberlin, B.; Bonfanti, R.; Schuep, W. Simultaneous determination of retinol, tocopherols, carotene and lycopene in plasma by means of high performance liquid chro-matography on reversed phase.Int. J. Vitam. Nutr. Res.61:232– 238; 1991.
[18] Pedersen, C. B.; Kyle, J.; Jenkinson, A. M.; Gardner, P. T.; McPhail, D. B.; Duthie, G. G. Effects of blueberry and cranberry juice consumption on the plasma antioxidant capacity of healthy female volunteers.Eur. J. Clin. Nutr.54:405– 408; 2000. [19] Jiang, Z. Y.; Woollard, A. C.; Wolff, S. P. Lipid hydroperoxide
measurement by oxidation of Fe2⫹ in the presence of xylenol
orange. Comparison with the TBA assay and an iodometric method.Lipids26:853– 856; 1991.
[20] Wood, S. G.; Gedik, C. M.; Vaughan, N. J.; Collins, A. R. Measurement of 8-oxo-deoxyguanosine in lymphocytes, cultured cells and tissue samples by HPLC with electrochemical detection.
Methods Mol. Med.38:171–178; 2000.
[21] Duthie, G. G.; Gonzalez, B. M.; Morrice, P. C.; Arthur, J. R. Inhibitory effects of isomers of tocopherol on lipid peroxidation of microsomes from vitamin E-deficient rats.Free Radic. Res. Commun.15:35– 40; 1991.
[22] Levi, F.; Pasche, C.; Lucchini, F.; La Vecchia, C. Dietary intake of selected micronutrients and breast-cancer risk.Int. J. Cancer
91:260 –263; 2001.
[23] Michaud, D. S.; Spiegelman, D.; Clinton, S. K.; Rimm, E. B.; Willett, W. C.; Giovannucci, E. Prospective study of dietary supplements, macronutrients, micronutrients, and risk of bladder cancer in US men.Am. J. Epidemiol.152:1145–1153; 2000. [24] Helzlsouer, K. J.; Huang, H. Y.; Alberg, A. J.; Hoffman, S.;
Burke, A. Norkus, E. P.; Morris, J. S.; Comstock, G. W. Association between alpha-tocopherol, gamma-tocopherol, selenium, and subse-quent prostate cancer.J. Natl. Cancer Inst.92:2018 –2023; 2000. [25] Shklar, G.; Oh, S. K. Experimental basis for cancer prevention by
vitamin E.Cancer Invest.18:214 –222; 2000.
[26] Sarma, A. D.; Sharma, R. Anthocyanin-DNA copigmentation complex: mutual protection against oxidative damage. Phyto-chemistry52:1313–1318; 1999.
[27] Seis, H.; Stahl, W.; Sundquist, A. R. Antioxidant function of vitamins: vitamin E and C, beta-carotene and other carotenoids.
Ann. N.Y. Acad. Sci.669:7–20; 1992.
[28] Dangles, D.; Dufour, C.; Fargeix, G. Inhibition of lipid peroxi-dation by quercetin and quercetin derivatives: antioxidant and pro-oxidant effects.J. Chem. Soc. [Perkin 2]2:1215–1222; 2000. [29] Rice-Evans, C.; Leake, D.; Bruckdorfer, K. R.; Diplock, A. T. Practical approaches to low density lipoprotein oxidation: whys, wherefores and pitfalls.Free Radic. Res.25:285–311; 1996.