Analysis of viral and cellular parameters which affect the fusion process of influenza viruses
Texto
Imagem
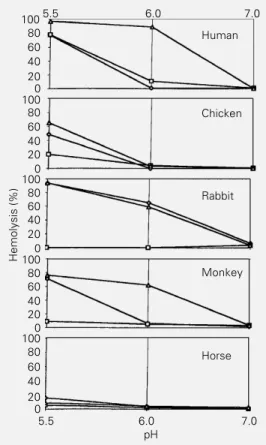
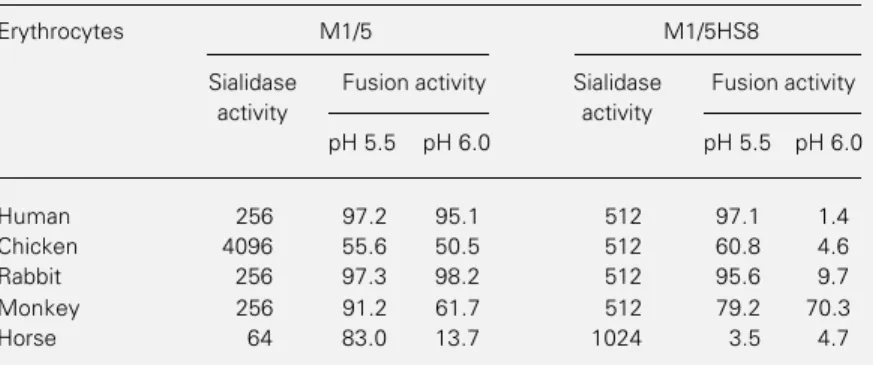
Documentos relacionados
Capoeira dança e arte Capoeira dança e arte – A arte se faz presente através da música, ritmo, canto, instrumento, expressão corporal, criatividade de movimentos, assim como
Sequence analysis of the HA gene of influenza B strains indicated that the viruses were closely related to a B/Brisbane/60/2008- like virus belonging to the Victoria lineage,
ao texto constitucional, havendo ainda que realçar tanto as fontes internacionais como as fontes legais. No plano internacional, Angola encontra-se vinculada aos sistemas de
O primeiro objectivo do presente trabalho é analisar as diferenças entre os resultados obtidos com base nas inspecções do ano de 2003 em (Almeida, 2003) e nas realizadas pelo
The circulation of influenza C viruses in Rio de Janeiro, Brazil, was studied when significant levels of antibodies were detected (56.7%) with hemagglutination inhibi- tion test,
Objective: Detection of the eight most common respiratory viruses: Human respiratory syncytial virus (HRSV), influenza virus A and B (IA and IB), parainfluenza viruses 1, 2 and
The objectives of this study were to determine the incidence of infection by respiratory viruses (RSV; parainfluenza viruses 1, 2 and 3; influenza A and B viruses; and adenovirus)
Taking into account the impact of influenza A(H1N1)pdm09, the unique charac- teristics of influenza viruses circulating worldwide and the lack of published data on molecular
