Relações biogeográficas entre comunidades de aves suportam diferentes histórias na mata atlântica
92
0
0
Texto
(2)
(3)
(4)(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
Imagem


+2
Documentos relacionados
◦ Os filtros FIR implementados através de estruturas não recursivas têm menor propagação de erros. ◦ Ruído de quantificação inerente a
No latossolo (LEa), como houve cresci- mento máximo em altura e diâmetro aos seis me- ses após o plantio, foram determinados os níveis críticos de fósforo nas folhas para estes
O objetivo do curso foi oportunizar aos participantes, um contato direto com as plantas nativas do Cerrado para identificação de espécies com potencial
Cândida Fonseca Duração e local: 11/09/2017 a 03/11/2017 – Hospital São Francisco Xavier Objectivos e actividades desenvolvidas: Os meus objectivos centraram-se na
• A existência de bloqueio por devolução, seja na conta empresa ou na conta vinculada do trabalhador, impossibilita a informação da movimentação pelo empregador via
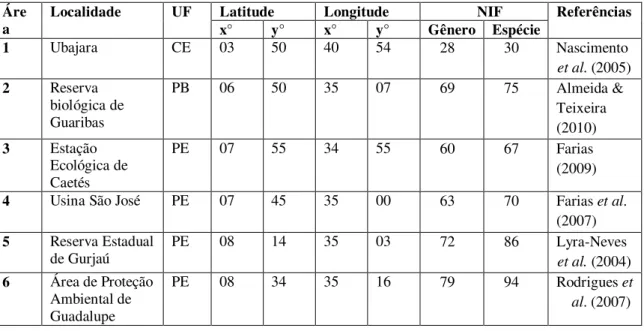
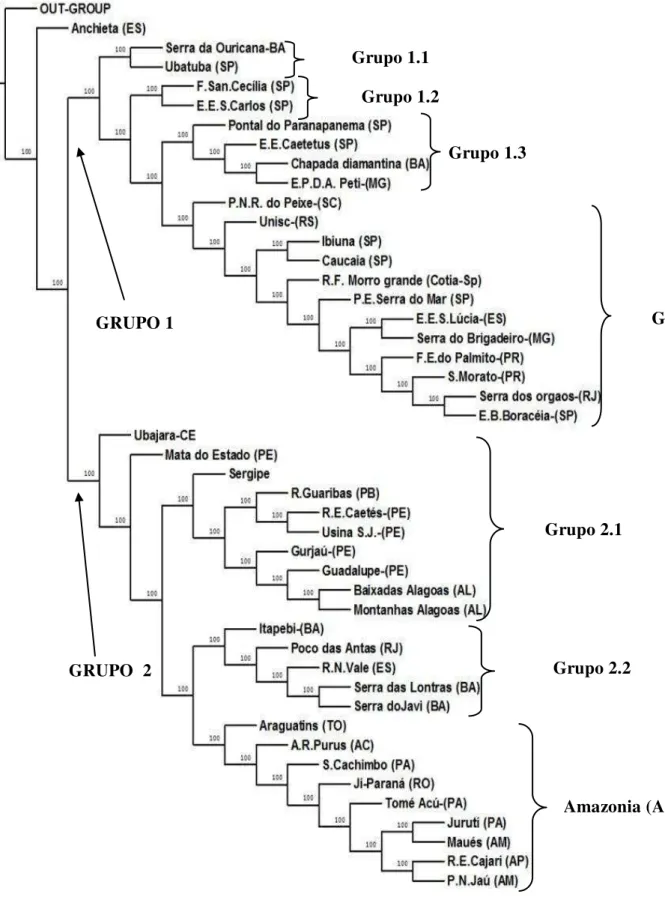
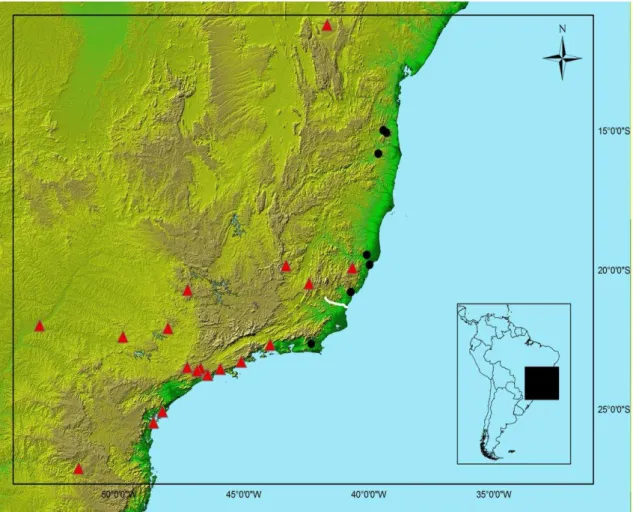
Espécies com registros apenas nas áreas limítrofes da Mata Atlântica, bem como espécies com ocorrência errante, não foram consideradas pertencentes ao
32 Figura 4 - Massa seca da parte aérea de mudas de Handroanthus ochraceus (Cham.) Mattos, produzidas com diferentes doses de adubação nitrogenada de cobertura
Em relação ao Currículo Tecnológico, McNeil (2001) afirma que a educação consiste na transmissão de conhecimentos, comportamentais éticos, práticas, sociais e
