SILVIA BEATRIZ FONSECA DE MELO
O GATO DOMÉSTICO (Felis catus) RESPONDE Á SINAIS GESTUAIS? POSSÍVEIS IMPLICAÇÕES DO CONVÍVIO SOCIAL.
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, como requisito parcial para obtenção do título de Mestre no Programa de Pós-Graduação em Psicobiologia.
SILVIA BEATRIZ FONSECA DE MELO
O GATO DOMÉSTICO (Felis catus) RESPONDE Á SINAIS GESTUAIS? POSSÍVEIS IMPLICAÇÕES DO CONVÍVIO SOCIAL.
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre no Programa de Pós-Graduação em Psicobiologia.
Orientação: Daniel Marques de Almeida Pessoa
Título: O GATO DOMÉSTICO (Felis catus) RESPONDE Á SINAIS GESTUAIS? POSSÍVEIS IMPLICAÇÕES DO CONVÍVIO SOCIAL.
Autor: SILVIA BEATRIZ FONSECA DE MELO
silviabeatriz28@gmail.com Data da defesa: 24 DE NOVEMBRO DE 2008
Banca examinadora:
_____________________________________________________ PROFESSORA. MARIA CLOTILDE HENRIQUES TAVARES
UNIVERSIDADE DE BRASÍLIA
mchtavares@unb.br
_____________________________________________________ PROFESSORA.FÍVIA DE ARAÚJO LOPES CAVALCANTI
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
fivialopes@yahoo.com.br
_____________________________________________________ PROFESSOR. DANIEL MARQUES DE ALMEIDA PESSOA
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
AGRADECIMENTOS
Agradeço ao meu orientador, o querido “Chefinho”, Professor Dr. Daniel Pessoa, pela coragem de pegar o barco andando, no meu momento de angústia e incertezas, tendo me aceitado como mestranda. Pelo incentivo para continuar sempre e palavras amigas nos momentos certos.
Obrigada querido chefinho!
Á professora Dra. Fívia Lopes, que me deu GRANDE ajuda, de longa data, sempre que precisei e mesmo até quando não podia.
Obrigada Filó, você é demais!
Agradeço ao meu companheiro de todas as horas, Carlos Meton, pelo incentivo de prosseguir em busca do que eu realmente queria e gostava.
Obrigada Benhê!
Aos familiares e amigos, que contribuíram de alguma forma nessa longa jornada. Meu muito obrigada!
Agradeço a todos os donos dos gatinhos que me foram cedidos para realização desse estudo, pela paciência e colaboração para com uma estranha em suas casas.
Obrigada, vocês foram muito gentis!
Tenho que agradecer também a todos os gatinhos que participaram dos testes, sem eles esse trabalho não seria possível e a todos os gatos que fizeram parte da minha vida, fonte inspiradora desse trabalho.
Vocês são lindos!
E por fim, agradecer ao maior e mais lindo de todos os gatos: Tchuthcuco. Pelas horas de análise de seu comportamento e por fazer dos meus dias os mais felizes, em sua companhia.
RESUMO
Os gatos (Felis catus) foram domesticados há cerca de 9.500 anos devido à agricultura, onde
eram utilizados no combate às pragas que assolavam os alimentos colhidos. Esses animais passaram por uma seleção artificial e ao longo das gerações e milênios tiveram seus comportamentos e morfologia modificadas pelos humanos. O processo de domesticação pelo homem fez surgir uma habilidade em especial, a compreensão de sinais gestuais humanos, que é bem observada nos momentos em que alimentamos nossos animais. Nosso objetivo neste estudo foi testar a resposta à sinalização gestual (comportamento de apontar) em gatos, emitida por humanos e também verificar a influência do convívio social sobre o desenvolvimento desta habilidade. Observamos que os sujeitos experimentais de ambos os grupos, animais solitários e de convívio em grupo, foram capazes de seguir os sinais de indicação humana para localizar o alimento escondido. Porém, a forma de convívio social não influenciou no desempenho dos gatos. A habilidade aqui testada, possivelmente evoluiu durante o processo de domesticação dessa espécie, e a interação social parece exercer pouca ou nenhuma influência sobre a sua expressão.
ABSTRACT
The cats (Felis catus) were domesticated about 9,500 years ago due to the advent of agriculture,
being used to control the pests that devastated the food harvested. These animals went through an artificial selection and over generations and millennia had their behavior and morphology changed by humans. This process of domestication by man gave rise to a special ability, the understanding of human pointing gestures, clearly noticed while we feed our pets. Our goal in this study was to assess the comprehension of pointing gestures by cats and also verify the influence that social interactions exerts on the development of this ability. We found that experimental subjects from both groups, solitary animals and social animals, were able to follow human indication in order to find hidden food. However, social interaction had no effect on cats’ performances. The ability tested here probably evolved during the process of domestication of this species, and social interaction seems to exert little or no influence upon its expression.
SUMÁRIO
AGRADECIMENTOS...iv
RESUMO... v
ABSTRACT...vi
LISTA DE ILUSTRAÇÕES...viii
1. INTRODUÇÃO... 1
2. OBJETIVOS... 9
3. HIPÓTESES E PREDIÇÕES... 10
4. MATERIAIS E MÉTODOS... 11
5. RESULTADOS... 19
6. DISCUSSÃO... 27
7. CONCLUSÕES... 34
REFERÊNCIAS... 35
LISTA DE ILUSTRAÇÕES
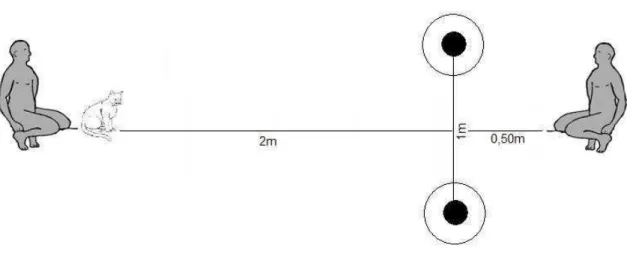
Figura 1a. Aparato experimental ...13
Figura 1b: Disposição do aparato e posicionamento do animal, dono e experimentador ...14
Figura 2: Resposta dos animais solitários (Felis catus) em relação à sinalização
humana. ...19
Figura 3: Resposta dos animais de convívio em grupo (Felis catus) em relação à sinalização
humana ...20
Figura 4: Influência do convívio social sobre a resposta dos animais em tarefas sinalizadas...21
Figura 5: Influência do convívio social sobre a resposta dos animais em tarefas não sinalizadas...22 Figura 6: Latência média de acertos de animais solitários (Felis catus) em resposta a sinalização
humana ...23 Figura 7: Latência média de acertos de animais de convívio em grupo (Felis catus) em resposta a
INTRODUÇÃO
A origem do gato doméstico e sua relação com o homem
Os gatos (Felis catus, Linnaeus, 1758) são animais dotados de uma notória capacidade
cognitiva para realização de tarefas que vêm sendo analisadas e testadas em laboratórios há algum tempo (Miklósi & Soproni, 2006). Mostrando o desenvolvimento de certas habilidades cognitivas, eles passaram a chamar a atenção de pesquisadores de todo o mundo, pois desde a antiguidade esses animais vêm sendo utilizados como modelos animais para pesquisas neurológicas, permitindo assim o melhor entendimento de nossa fisiologia (Beaver, 2005).
Existem relatos de que os gatos já conviviam com os seres-humanos há pelo menos 4 mil anos, no Egito antigo (Clutton-Brock, 1993; Yamaguchiet al., 2004). Entretanto, a afirmação de
que esses animais já estavam presentes na região do Crescente Fértil (Nordeste da África) e na ilha mediterrânea do Chipre sugere uma coabitação mais antiga entre gatos e seres-humanos, entre 8 e 10 mil anos atrás (O`Brien & Johnson, 2007). É possível que o início da domesticação de felinos tenha ocorrido através da agricultura uma vez que o armazenamento de grãos facilitava o aparecimento de roedores (Driscoll et al., 2007). Visando a diminuição dos estragos produzidos
Após a constatação de que esses animais tinham muito a contribuir para o extermínio de pragas, passaram a ter a liberdade de transitar dentro das casas e assim, possivelmente, começou a sua proximidade e interação com o homem. Desde então, esses felinos vêm evoluindo juntamente com a nossa espécie, sendo selecionados, domesticados e moldados para o convívio social com seres-humanos. Ainda assim, com todas as evidências sobre seus antepassados e interações com os humanos, a sua origem e seu processo de domesticação continuam incertos (Clutton-Brock, 1999).
Domesticação e seleção artificial
Alguns estudos sugerem que os canídeos foram selecionados pelo homem ao longo de sua evolução para que se tornassem animais mais dóceis e companheiros. Um deles, descreve as mudanças físicas e comportamentais ocorridas com gerações de raposas criadas e domesticadas por humanos (Trut, 1999). No caso dos cães, esse processo de criação e convívio com os homens possivelmente acabou conferindo-lhes uma alta inteligência social (Hare et al., 2002; Hare &
Tomasello, 2005; Miklósi et al., 2004).
foram se diferenciando e sofrendo modificações genéticas além de adaptações comportamentais ao novo estilo de vida em meio a nossa espécie (Bradshaw & Brown,1990; Morris, 1986; Serpell, 1995).
Da mesma forma que os cães, os gatos também sofreram um processo de seleção artificial com base em características morfológicas e comportamentais com possíveis implicações em suas habilidades cognitivas. Essas habilidades possivelmente emergiram a partir da convivência e observação dos atos de nossa espécie e é possível dizer que o processo de domesticação tornou tais animais aptos ao convívio humano (Miklósi et al., 2005). Baseando-se nisso, é possível que
ao serem inseridos na sociedade humana, os gatos tenham precisado se adaptar a uma nova forma de sobrevivência pois, embora tenham conservado a maior parte dos seus instintos, precisaram aprender novas habilidades para a interação com nossa espécie. É possível que esses animais tenham apresentado dificuldades para manipulação de objetos, que não eram comuns ao seu meio ou até mesmo o aprendizado para a vivência dentro das habitações humanas pode ter contribuído para modificar certas habilidades cognitivas, como a capacidade de reagir de maneira adequada a novas situações impostas pelo convívio com os humanos. De fato, existem indícios de que a seleção desses animais com base na compreensão dos sinais comunicativos humanos, incluindo o gesto de apontar, parece ter modificado sua inteligência social (Miklósi & Soproni, 2006).
Baseados em sua organização social, os gatos são considerados por alguns pesquisadores como uma espécie predominantemente solitária (Jones & Coman, 1982; Parcker 1986; Sandell 1989). Mas, segundo Izawa e colaboradores (1982), esses felinos também podem ser considerados animais sociais, formando colônias e alianças de acordo com a disponibilidade de alimentos (Crowell-Davis et al., 2004). Dentro ou fora dessas alianças, esses animais apresentam
diferentes tipos de vocalizações, e apresentam também diversas maneiras de comunicação intra-especifica de contato, como: to nose touch (toque de nariz), o allogroming (comportamento de
lamber outro gato), o allorub (comportamento de se esfregar em outro animal, pessoas ou
objetos), além de movimentos do rabo e das orelhas (Crowell-Davis et al., 2004).
Em relação à organização social dos cães, estes apresentam diferenças e também semelhanças com os gatos. Exemplo disto é que, como precisavam viver em grupos, desenvolveram uma hierarquia social bem definida. Na matilha, há sempre um líder que conduz, impondo regras ao grupo por meio de sinais e atitudes, como por exemplo, a marcação do território com urina (Beaver, 2001).
Comunicação e sinalizações gestuais
Estudos realizados com espécies domésticas (e.g. cães e gatos) sobre a habilidade de
entendimento e comunicação através de gestos humanos demonstram que esses animais são capazes de entender e desempenhar com sucesso tarefas relacionadas à procura por alimento guiada por informações gestuais (Miklósi et al., 1998; Miklósi et al., 2005). Como já amplamente
humanos, muito úteis na aquisição de comida. Ainda assim, quando comparado aos estudos com chimpanzés (Call et al., 1998), cães (Hare et al., 1998; Kubinyi et al., 2003; Miklosi et al., 1998;
Morey, 1994; Virányi et al., 2004,), golfinhos (Herman et al., 1999) e pássaros sociais (Emery &
Clayton, 2001; Pepperberg, 1991), o estudo da cognição social em gatos ainda é muito recente (Miklósi et al., 2005), restando muitas questões em aberto.
Parece haver uma relação forte entre a compreensão de sinais gestuais humanos e o processo de domesticação (Miklósi & Soproni, 2006). De fato, essas habilidades cognitivas não costumam ser observadas em espécies não domesticadas, mesmo que estas apresentem elevados graus de cognição. Trabalhos que compararam o desempenho de chimpanzés ao de cães (Hare et
al., 2005; Tomasello et al., 1998) mostraram diferenças quanto à detecção de pistas oferecidas
realizados testes com um grupo controle de raposas e observou-se que essas foram menos habilidosas do que as raposas domesticadas. Em conjunto, todos esses achados fortalecem a idéia de que a compreensão do comportamento social comunicativo dos humanos evoluiu durante o processo de domesticação dessas espécies (Hare et al., 2005).
A transmissão social de informação geralmente, ocorre entre indivíduos da mesma espécie. Entretanto, existem casos especiais em que essa transferência pode ser do tipo hetero-especifica, como mostrado para cães (Kubinyi et al., 2003). No caso dos felinos, parece existir
uma necessidade de comunicação inter-específica com seus donos, pois esse processo de comunicação os torna cada vez mais adaptados à vida doméstica. Em um estudo enfocando a interação entre os gatos e seus donos, Mertens (1991) fornece algumas evidências dessas necessidades. Segundo o autor, os gatos que eram os únicos da casa, passavam a maior parte do tempo brincando e interagindo socialmente com seus donos, diferentemente dos animais que viviam em domicílios com mais de um gato.
JUSTIFICATIVA
No trabalho realizado por McCune (1995), foi observado que gatos com um ano de idade, previamente socializados quando filhotes, ou seja, que foram manuseados por pessoas durante algumas horas por dia num período de semanas, aproximaram-se, tocaram ou se esfregaram em pessoas desconhecidas ou objetos, sendo mais exploratórios do que os animais que não foram previamente socializados. As evidências de que os gatos são animais que também apresentam uma organização social (Crowell-Davis et al., 2004) e passaram por um processo de
domesticação, modificações e seleção pelo convívio com a espécie humana (Driscoll et al.,
2007), nos levam a querer compreender um pouco mais sobre as habilidades de cognição social dos gatos. O estudo das variáveis evolutivas relacionadas ao processo de responder aos gestos humanos, a avaliação da interação e comunicação entre esses felinos e o homem, e a investigação de variáveis ontogenéticas são pontos que ainda permanecem pouco estudados.
No estudo de Miklósi & colaboradores 2003, foi investigada a diferença quanto ao desenvolvimento de respostas comunicativas entre cães e lobos quando estes se baseavam no comportamento de olhar as expressões faciais de humanos. Os autores sugerem que existe uma base filogenética e ontogenética que levou os cães a uma incontestável utilização dessa forma de comunicação com a nossa espécie. Contudo, essa mesma forma comunicativa, que é utilizada rotineiramente por nós em nossas casas, também parece exercer algum papel de comunicação com os gatos domesticados (Miklósi et al., 2005). Uma vez que cães e gatos estão
filogeneticamente distantes, a domesticação parece ser o único fator capaz de explicar o surgimento dessas habilidades em ambos os grupos. Ademais, isto sugere que o surgimento de habilidades sociais relacionadas à resposta a gestos humanos não careceu da existência de pré-adaptações relacionadas à vida em grupo, uma vez que cães e gatos diferem significativamente quanto a este parâmetro. Alguns estudos mostram que cães e gatos são espécies altamente diferentes quanto ao seu comportamento social, pois enquanto os cães, assim como seus ancestrais, são animais que vivem e caçam em bandos, os gatos em sua grande maioria são caçadores solitários por isso sendo considerados como animais de vida solitária (Bradshaw & Brown, 2006; Fox, 1971), embora outros autores sugiram que os gatos são animais com fortes padrões de sociabilidade (Crowell-Davis et al., 2004). Assim, surge a seguinte pergunta: será que
OBJETIVO GERAL:
Testar a habilidade cognitiva apresentadas por gatos domésticos com relação à resposta aos sinais gestuais emitidos por seres-humanos;
OBJETIVOS ESPECÍFICOS:
1. Testar a resposta comportamental apresentada por gatos domésticos em relação aos gestos de apontar emitidos por seres-humanos;
HIPÓTESES E PREDIÇÕES:
1. Hipótese: Os gatos são capazes de desempenhar tarefas respondendo a sinais gestuais emitidos por seres-humanos;
Predição: O animal responderá ao gesto emitido por um humano, identificando a localização da recompensa alimentar numa freqüência acima da esperada para uma situação não sinalizada.
2. Hipótese: Existe diferença de resposta à localização do alimento pelos animais que convivem apenas com humanos se comparada à resposta de animais que vivem com co-específicos e humanos.
MATERIAIS E MÉTODOS:
Sujeitos
Para o recrutamento dos animais foi utilizada uma rede de contatos próximos do experimentador. Aos donos dos animais foi pedida permissão verbal para utilização destes. A partir desses, outros donos de gatos foram indicados para recrutamento dos seus animais, a fim de que viessem fazer parte do experimento. Em seguida concordância das pessoas, o experimentador compareceu no mínimo duas vezes à casa onde habitavam os animais para que estes se habituassem com o experimentador antes do início do experimento. Ao todo foram recrutados 60 animais, entretanto a maioria destes não se interessou ou não colaborou com os testes, sendo então descartados do experimento.
Para o estudo utilizamos 20 gatos (Felis catus) adultos, sendo 10 machos e 10 fêmeas, que
foram classificados e analisados de acordo com o convívio social em que se encontravam:
A) 10 Gatos de vida solitária, que convivem exclusivamente com seres humanos, sendo cinco machos e cinco fêmeas;
B) 10 Gatos de vida em grupo, que convivem com outros gatos e com seres humanos, sendo cinco machos e cinco fêmeas.
Um questionário (Anexo 1) foi aplicado aos donos dos animais antes do início das coletas. Nesse questionário foram solicitadas informações necessárias para a triagem dos animais, a fim de que fosse possível separá-los nos dois grupos e classificá-los de acordo com a exigência do experimento. Além disso, com o questionário procuramos adquirir informações sobre as condições físicas, motoras e sensoriais dos animais, sendo dessa forma selecionados os que não apresentavam problemas de visão ou deficiência nas patas, por exemplo, que pudessem ser detectados visualmente. Nesse estudo não foram utilizadas fêmeas grávidas, fêmeas com filhotes, fêmeas que faziam uso de anticoncepcional, ou animais que fizessem uso de qualquer outro tipo de medicamento.
Piloto:
Aparato experimental
O aparato consistiu em dois pratos quadrados (17cm x 17cm) e dois copos de plástico opaco branco (8cm de diâmetro e 10,5cm de altura), conforme pode ser observado na Figura 1a. Os copos foram emborcados e posicionados acima dos pratos, de forma a esconder as recompensas alimentares, pequenos pedaços de ração para gatos, Whiskas (Fabricante Waltham) sabor atum, oferecidas aos animais durante o experimento.
Ao final de cada sessão experimental, todo o aparato foi higienizado com detergente neutro.
Os pratos foram posicionados no chão, de forma a ficarem a um metro de distância entre si, a 50 centímetros de distância do experimentador e a dois metros do animal (Figura 1b).
Figura 1b. Disposição do aparato e posicionamento do animal, dono e experimentador.
Procedimento experimental
Para a escolha do local de realização do experimento foram selecionados aqueles cômodos que apresentaram menor quantidade de estímulos visuais, como uma sala ou um quarto com poucos móveis.
Cada sessão experimental foi subdividida em duas fases, uma fase de treino e uma fase de testes. A primeira tentativa da fase de treino consistiu na colocação de uma porção da recompensa alimentar sobre um dos pratos. Neste caso, o animal pôde comer a isca durante 30 segundos. A seguir, nas cinco tentativas seguintes, a recompensa foi depositada em um dos pratos que teve os copos posicionados sobre eles, de forma que o sujeito pudesse visualizar onde a recompensa havia sido escondida. O animal deveria ser capaz de, em cinco tentativas consecutivas, indicar corretamente qual dos dois pratos continha o alimento. A fase de treino não só teve a função de assegurar ao experimentador que o sujeito estava motivado a participar do experimento, como também permitia que o animal aprendesse quais eram as regras do mesmo e qual o padrão motor necessário para a aquisição da recompensa. O animal deveria ser capaz de derrubar o copo, com a pata ou com o focinho, para adquirir o alimento escondido. O experimentador sempre ficou posicionado exatamente entre os dois pratos, evitando qualquer influência sobre a escolha do animal.
mensurada com auxílio de um cronômetro e teve o tempo máximo de 1 minuto. Caso a resposta do animal ultrapassasse o tempo de 1 minuto, a tentativa era dada como encerrada, não sendo válida, e em seguida o animal tinha a chance de responder a uma nova tentativa.
No que se refere à distância entre o dedo indicador do experimentador e o objeto apontado, o tipo de indicação utilizada foi o proximal pointing, onde uma distância de dez a vinte
centímetros separou o dedo e o aparato (Miklósi et al., 2005). Quanto à duração do gesto de
indicação, os movimentos realizados com o braço foram do tipo dynamic pointing, onde o braço
do experimentador permaneceu indicando o objeto até que o animal fosse solto e fizesse sua escolha (Miklósi et al., 2005). O animal pôde ver toda a movimentação do braço. O movimento
de indicação seria repetido caso o animal não deixasse o colo do dono quando isso fosse permitido.
Na fase de testes, a indicação de apontar descrita foi repetida quatorze vezes, pois segundo a tabela de aleatoriedade de Gellerman, a quantidade máxima de erro por tentativas em que não deixaria o teste aleatório seria de 3 erros para um total de quatorze tentativas. Em adição, quatorze tentativas controles também foram realizadas, durante as quais o experimentador permaneceu com os braços imóveis rentes ao corpo, não havendo indicação da posição da recompensa. O controle foi realizado como um fator inédito para esse tipo de experimento, pois os testes já realizados e descritos na literatura apresentam apenas as tentativas indicadas, como o trabalho de Miklósi et al., 2005. As tentativas com indicação e sem indicação (controle) foram
Durante as tentativas, o experimentador olhou diretamente para o animal, de forma a evitar que a posição correta do alimento fosse indicada por seu olhar.
O animal teve a chance de escolher apenas um copo. Caso escolhesse o copo errado no momento de indicação, não era permitido que ele comesse a comida escondida no outro copo. Caso ele escolhesse o correto, ele poderia consumir o alimento livremente até acabar. A ordem de colocação da comida sob o copo direito ou esquerdo foi realizada aleatoriamente para que não houvesse fixação de posição pelo animal.
Para evitar que os animais utilizassem pistas olfativas na procura pelo alimento, em todos os pratos e copos foi esfregado um pouco da recompensa alimentar, de forma que apresentassem o mesmo odor.
Análise de dados
Utilizamos estatística não paramétrica para a análise dos dados obtidos, comparando: tentativas sinalizadas vs não sinalizadas, animais solitários vs animais de convívio em grupo; tanto quanto ao desempenho medido em acertos, como quanto à latência média de acertos. A análise foi realizada através dos testes Wilcoxon Signed Ranks Test, para os diferentes tipos de tentativas dentro de um mesmo grupo (sinalizadas vs não sinalizadas para solitários e também para animais de convívio em grupo), e Mann Whitney, para as análises entre os diferentes grupos (solitários vs animais em grupo para cada tipo de tentativas) em uma mesma situação de sinalização. Os testes foram interpretados com base em um nível de significância de 5%
(α=0,05).
RESULTADOS:
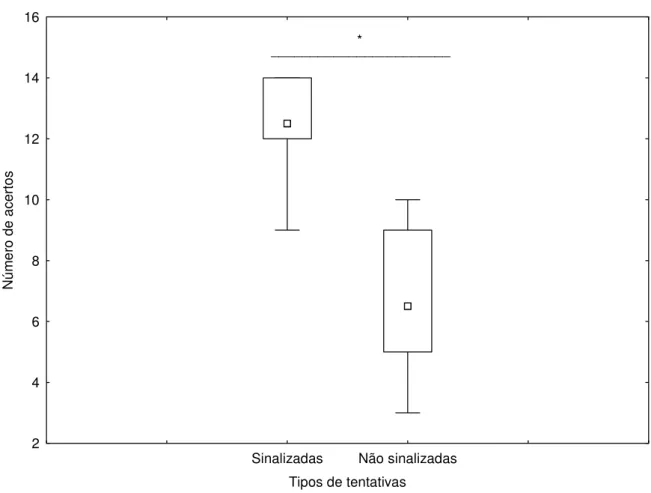
Os dados representados na Figura 2 mostram a comparação entre as tentativas sinalizadas e não sinalizadas apenas para o grupo dos animais que viviam solitariamente, isto é, sem a presença ou contato com nenhum outro gato. Observamos que os animais tiveram uma diferença de desempenho, sendo este significativamente superior na situação de tentativas sinalizadas quando comparadas às não sinalizadas (Z=2,80 e p=0,005), indicando que os animais
conseguiram responder seguindo os sinais gestuais demonstrados por um humano.
Sinalizadas Não sinalizadas
Tipos de tentativas 0 2 4 6 8 10 12 14 16 N úm er o de ac er tos ______________________* Figura 2: Resposta dos animais solitários (Felis catus) em relação à sinalização humana.
Na Figura 3, observamos o desempenho dos animais de convívio em grupo em relação ao número de acertos de ambos os tipos de tentativas. Vemos que os animais apresentaram desempenho significativamente superior nas tentativas sinalizadas em relação às não sinalizadas (Z=2,70 e p=0,0069). Da mesma forma que no resultado anterior, percebemos que os animais
realmente conseguiram responder a sinalização produzida indicando a localização do alimento.
Sinalizadas Não sinalizadas
Tipos de tentativas 2
4 6 8 10 12 14 16
Nú
m
e
ro
de
a
c
ert
o
s
_______________________*
Figura 3: Resposta dos animais de convívio em grupo (Felis catus) em relação à sinalização
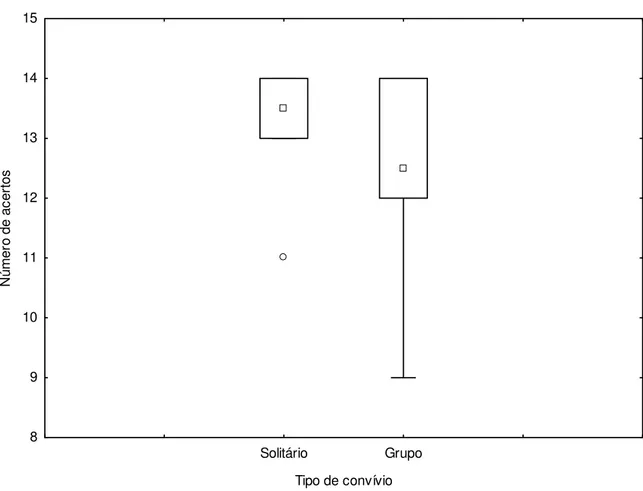
Na Figura 4, analisamos a influência do convívio social dos animais sobre o desempenho nas tentativas sinalizadas. Observamos que não existiu diferença significativa entre o desempenho dos grupos em relação ao tipo de tentativa em questão (U=36,50 e p=0,3075).
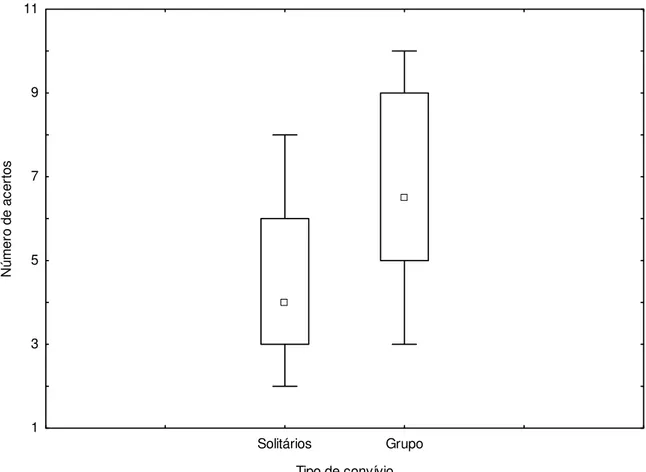
Da mesma forma para a Figura 5, a influência do convívio social relacionada ao desempenho dos animais para as tarefas não sinalizadas também não mostrou diferença significativa (U=25,50 e
p=0,0640).
Solitário Grupo
Tipo de convívio 8
9 10 11 12 13 14 15
N
ú
mer
o
de ac
er
tos
Solitários Grupo
Tipo de convívio 1
3 5 7 9 11
N
ú
mer
o
de ac
er
tos
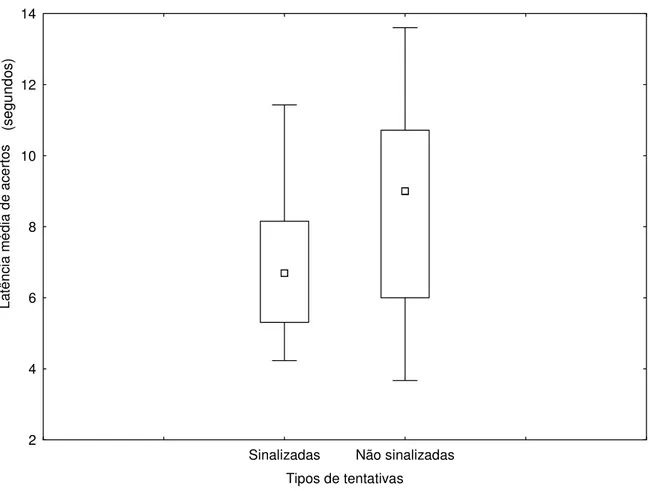
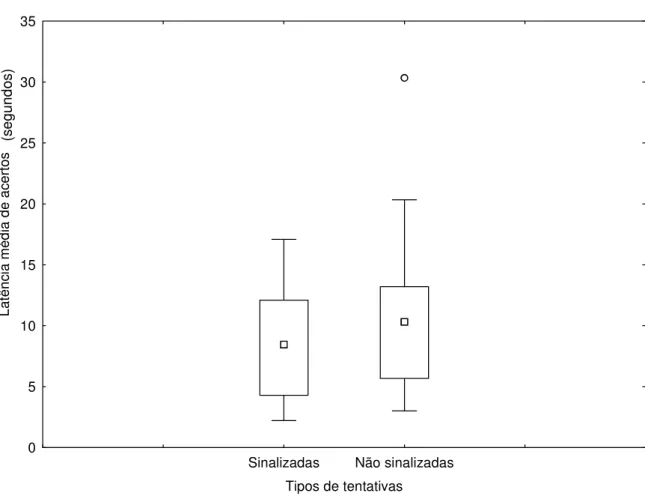
Analisamos a influência da sinalização humana sobre a latência média de acertos, medida em segundos. Não foi observada diferença significativa para as formas de indicação, tentativas sinalizadas e não sinalizadas (Figuras 6 e 7), para os animais solitários (Z=1,68 e p=0,0926), e
animais de convívio em grupo (Z=1,95 e p=0,0506), o que sugere que os animais dos diferentes
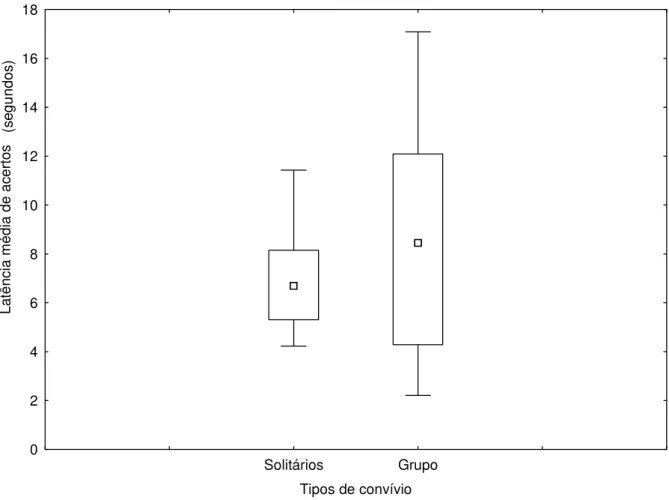
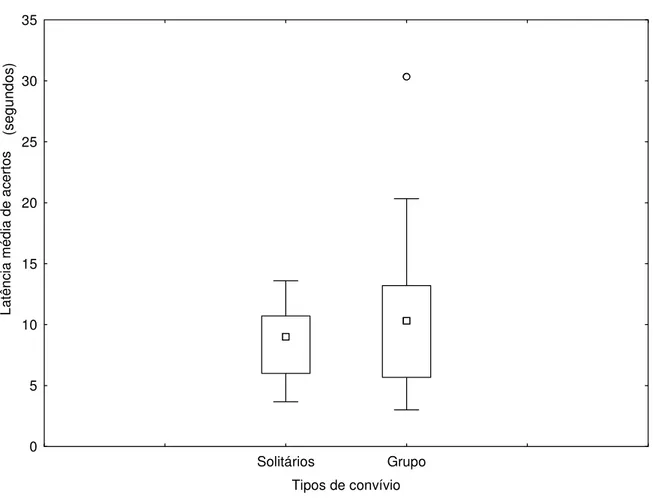
grupos não foram favorecidos pela presença dos sinais gestuais. As Figuras 8 e 9, não mostraram diferença nas respostas dos animais quanto a sinalização de indicação em relação ao tipo de convívio dos mesmos (U=42,00 e p=0,5454) e (U=34,00 e p=0,2265) respectivamente. O fato
dos animais serem mantidos em grupo ou isoladamente não influenciou no tempo gasto para a execução das diferentes tentativas do teste.
Sinalizadas Não sinalizadas
Tipos de tentativas 2 4 6 8 10 12 14 L at ê nc ia m é di a d e ac e rt os
Figura 6: Latência média de acertos de animais solitários (Felis catus) em resposta a sinalização
Sinalizadas Não sinalizadas
Tipos de tentativas 0 5 10 15 20 25 30 35 La tê nc ia m é di a de ac e rt o s
Figura 7: Latência média de acertos de animais de convívio em grupo (Felis catus) em resposta a
sinalização humana. Os símbolos representam: □ Mediana, Quartis 25%-75%, ┬ Intervalo de resultados (desconsiderando outliers), ○ Outlier.
(s
egund
os
Solitários Grupo
Tipos de convívio 0
2 4 6 8 10 12 14 16 18
La
tên
c
ia média de ac
erto
s
Figura 8: Influência do convívio social sobre a latência média de acertos em tarefas sinalizadas. Os símbolos representam: □ Mediana, Quartis 25%-75%, ┬ Intervalo de resultados (desconsiderando outliers).
(s
e
gu
ndos
Solitários Grupo
Tipos de convívio 0 5 10 15 20 25 30 35 La tê nc ia m é di a de ac e rt o s
Figura 9: Influência do convívio social sobre a latência média de acertos em tarefas não sinalizadas. Os símbolos representam: □ Mediana, Quartis 25%-75%, ┬ Intervalo de resultados (desconsiderando outliers), ○ Outlier.
DISCUSSÃO
também um grupo controle de tentativas não-sinalizadas, o que não foi realizado pelo estudo de Miklósi.
Ademais, esse fato também foi verificado no trabalho de Soproni & colaboradores (2002), no qual eles sugerem que cães apresentam responsividade a novos gestos comunicativos de humanos, quando verificaram que estes foram capazes de seguir formas gestuais de indicação e de certa forma capazes de compreender a natureza indicativa do gesto. Em 2001, Soproni & colaboradores, também compararam as habilidades de cães para o uso de pistas sociais para localizar alimentos com um outro estudo publicado feito com chimpanzés e crianças (Povinelli, et
al., 1999), no qual observaram que os cães tiveram um desempenho mais próximo ao das
crianças do que os chimpanzés o fizeram. Essa semelhança pode ser atribuída ao fato de que os cães gastam mais tempo em interação com os seres humanos do que os macacos, o que de certa forma, também corrobora a nossa idéia de que os gatos de nosso estudo desempenharam satisfatoriamente bem as tarefas a eles impostas, pois são animais de estreito contato com humanos e que gastam boa parte do seu tempo interagindo e observando os comportamentos sociais e gestuais de nossa espécie (Beaver, 2005). A experiência social é uma condição importante no reconhecimento dos gestos humanos, no caso dos cães. Podemos pensar que é possível que os gatos que foram selecionados para o convívio com a espécie humana tenham usado essa habilidade como uma vantagem seletiva, utilizando as pistas fornecidas pelos humanos tornando mais fácil a obtenção de comida.
Durante as sessões experimentais não foi fornecida qualquer tipo de pista além da sinalização gestual de indicação aos animais, pois alguns estudos indicam que alguns animais são capazes de se orientar através de pistas faciais, movimento de cabeça, ou olhar indicativo (Call et
traria um resultado falso devido a outras indicações que não as propostas para este estudo. Pelo fato de termos sido criteriosos quanto às pistas fornecidas, podemos afirmar que nossos animais realmente seguiram a sinalização gestual de indicação.
Um exemplo de resultado dessa interação é o trabalho de Pongrácz et al., (2001), no qual
é mostrado que cães melhoraram seu desempenho em um teste de recuperar um objeto ou alimento após terem observado uma demonstração humana. Além disso, um estudo realizado por Topál et al., (2006), mostra que cães apresentam uma nítida capacidade de imitação baseada em
observação a partir de ações humanas. Esse trabalho sugere que os cães devem adquirir habilidades através da observação, o que aumenta o seu sucesso em situações complexas de comportamento social. É possível que aqueles animais que são submetidos a um maior tempo de contato apenas com os humanos, como geralmente é o caso dos animais que vivem solitariamente com seus donos, muitas vezes habitando em apartamentos, poderiam chegar a responder positivamente as sinalizações gestuais de nossa espécie por esse contato mais estrito.
Sabe-se que uma das formas mais comuns de sinalização gestual, ou gestos referenciais, é a sinalização de apontar, que no estudo de Xitco & colaboradores (2001) é mostrado através de testes com golfinhos que foram capazes de reproduzir gestos referenciais ou de indicação em relação a um objeto ou alimento. Nesse caso, entretanto, deixou-se claro que esses animais tiveram uma prévia interação com seres humanos por muitos anos, corroborando com a idéia de que o contato permanente com humanos diferencia esses animais no sentido de uma melhor desenvoltura para realização de tarefas complexas baseado apenas em sinalizações gestuais.
comportamento. De toda forma, nossos dados estão de acordo com os resultados mostrados em alguns trabalhos que têm sido realizados com animais apenas socializados com a espécie humana. Por exemplo, focas, que apresentaram tempo de contato reduzido com humanos diariamente foram capazes de desempenhar teste de escolha entre objetos baseados apenas em pistas de indicação (Scheumann & Call, 2004). Além disso, no estudo realizado por Hare & colaboradores (2002), é mostrado que cães são analisados em relação à realização de tarefas envolvendo a sinalização de comida escondida e seu desempenho é comparado ao de chimpanzés, e ao de lobos, ancestrais dos cães. Nesse estudo, observa-se que os cães apresentam o melhor desempenho das três espécies. Os lobos do estudo foram criados por humanos acreditando-se que esse fato contribuiria para um desempenho favorável nessa questão. Entretanto o que foi visto é que os filhotes dos cães apresentaram desempenho superior aos dos filhotes de lobos e que essa experiência com humanos continuou sendo satisfatoriamente demonstrada da mesma forma quando se tornaram cães adultos. Os autores chegaram à conclusão de que apenas a simples exposição humana a curto prazo não seria suficiente para um bom desempenho dos animais, como no caso dos lobos. Os cães mostraram que foi necessário um fator a mais do que apenas exposição imediata à nossa espécie, ou seja, foi necessário um processo que durou milênios, chamado de domesticação. Podemos extrapolar esse fato para os nossos animais experimentais, pois estes, de longa data, também passaram por processo semelhante de domesticação pelo homem, portanto mostrando também evolução das habilidades sócio-cognitivas aqui testadas (Hare et al., 2002).
si só com a espécie humana não foi suficiente para mostrar diferença no comportamento dos nossos sujeitos experimentais quando relacionamos a forma de convívio social. Baseados nessas evidências, é possível pensar que os nossos animais já nasçam portando toda a maquinaria neural necessária para a expressão da habilidade estudada, mesmo sem a influência do ambiente. Além disso, não podemos descartar a possibilidade de que com uma maior amostra de sujeitos pudéssemos ter encontrado uma diferença significativa nos resultados referentes à comparação entre os dois grupos.
Alguns animais experimentais que conviviam em grupo, podiam estar sob influência da hierarquia, o que poderia interferir no seu desempenho durante o teste, entretanto este fator possivelmente não teve consequências sobre o resultado do experimento pelo fato de que os nossos animais foram testados separadamente dos outros membros do grupo no momento do experimento e mesmo antes das sessões quanto separados para a ambientação. Cada animal foi ambientado no recinto escolhido para os testes e com a segurança de ser um local neutro de dominância de qualquer um dos membros, a fim de evitar qualquer tipo de influência que a hierarquia pudesse exercer sobre o desempenho de cada um dos animais.
CONCLUSÕES
Podemos ressaltar que esse estudo sugere que os gatos utilizam a sinalização gestual de indicação emitidos por um humano para a localização de alimento. Esse fato observado nos resultados indica a existência de uma habilidade cognitiva que é observada em poucas espécies de animais. Em contrapartida, percebemos que a utilização dessa sinalização gestual não influenciou o desempenho dos animais bem como não apresentou diferenças relevantes quando analisamos se eles viviam em grupo ou solitariamente.
REFERÊNCIAS BIBLIOGRÁFICAS
Beaver, B. V. (2001). Comportamento canino: um guia para veterinários. São Paulo: Roca.
Beaver, B. V. G. (2005). Comportamento Felino: um guia para veterinários. 2ª ed. São Paulo: Roca.
Blanc, P. R. (2005). Larousse dos Gatos: comportamentos, cuidados, raças. São Paulo: Larousse
do Brasil.
Bradshaw, J. W. S. & Brown, S. L. (1990). Behavioral adaptations of dogs to domestication. Waltham Symposium. 20, 18-24.
Bradshaw, J. W. S., & Brown, S. L., (2006). Beahvioural adaptations of dogs to domestication. In: Pets, benefits and practice, Waltham Symposium No. 20, ed. Burger. I., H., J., Small Animal Practice. (pp. 18-24).
Call, J., Hare, B. A., & Tomasello, M. (1998). Chimpanzee gaze following in an object-choice task. Animal Cognition. 1, 89-99.
Clutton-Brock, J. (1993). The animal that walks by itself. In: Yearbook of Science and the Future. (pp. 156-177). Encyclopedia Britannica, Chicago.
Clutton-Brock, J. (1999). A Natural History of Domesticated Mammals. 2a Ed. Cambridge University Press, Cambridge., The Natural History Museum, London.
Driscoll, C.A., Menotti-Raymond, M., Roca, A.L., Johnson, W.E. & O'Brien, S.J. (2007). The Near Eastern Origin of Cat Domestication. Science. 317, 519–523.
Emery, N. J. & Clayton, N. S. (2001). Effects of experience and social context on prospective caching strategies by scrub jays. Nature. 414, 443-446.
Fox, M.W., (1971). Integrative development of brain and behavior in the dog. Chicago. University of Chicago Press.
Hare, B., Call, J., & Tomasello, M. (1998). Communication of food location between human and dog. Evolution of Communication. 2(1), 137–159.
Hare, B., & Tomasello, M., (1999). Domestic dogs (Canis familiaris) use human and conspecific
social cues to locate hidden food. Journal of Comparative Psychology. 113, 173–177.
Hare, B. & Tomasello, M. (2005). Human like social skills in dogs? Trends in cognitive sciences. 9, 439-444.
Hare, B., Brown, M., Willianson, C. & Tomasello, M. (2002). The domestication of social cognition in dogs. Science. 298, 1634-1636.
Hare, B., Plyusnina, I, Ignacio, N., Schepina, O., Stepika, A., Wrangham, R. & Trut, L. (2005). Social cognitive evolution in captive foxes is a correlated by-product of experimental domestication. Current Biology. 15, 226-230.
Herman, L., Abichandani, S. L., Elhajj, A. N., Herman, E. Y. K., Sanchez, J. L., & Pack, A. A. (1999). Dolphins (Tursiops truncatus) comprehend the referencial character of the human
Izawa, M., Doi, T. & Ono, Y. (1982). Grouping patterns of feral cats living on a small island is Japan. Japanese Journal of Ecology.32, 373-382.
Jones, E. & Coman B. J. (1982). Ecology of the Feral Cat Felis catus (L.) in south-eastern
Australia. Australian Wildlife Research. 9, 409-420.
Kubinyi, E., Miklósi, Á., Topál, J., Csányi, V. (2003). Dogs (Canis familiaris) Learn From Their
Owners via Observation in a Manipulation Task. Journal of Comparative Psychology, 117 (2), 156-165.
McCune, S. (1995). The impact of paternity and early socialisation on the development of cats' behaviour to people and novel objects. Applied Animal Behaviour Science. 45, 109-124.
Mertens, C. (1991). Human-cat interactions in the human setting. Anthrozoös. 4, 214-231.
Miklósi, Á., R. Polgárdi, J. Topál, & V. Csányi. (1998). Use of experimenter given cues in dogs. Animal Cognition. 1, 113-121.
Miklósi, Á., Kubinyi, E., Topál, J., Gacsi, M., Virányi, Zs. and Csáni, V. (2003). A simple reason for a big difference: wolves do not look back at humans but dogs do. Current Biology. 13, 763- 766.
Miklósi, Á., Topál, J. and Csányi, V. (2004). Comparative social cognition: What can dogs teach us? Animal behavior. 67, 995-1004.
Miklósi, Á., Pongrácz, P., Lakatos, G., Topál, J., & Csányi, V. (2005). A comparative study of the use of visual communicative signals in interactions between dogs (Canis familiaris) and
Miklósi, Á. & Soproni, K. (2006). A comparative analysis of animals' understanding of the human pointing gesture. Animal Cognition. 9, 81-93.
Miller, J. (1996). The domestic cat: perspective on the nature and diversity of cats. Journal of the American Veterinary Medical Association. 208(4), 498-502.
Morey, D. F. (1994). The early evolution of the domestic dog. American Scientist. 82, 336-47.
Morris, D. (1986). Guia Essencial do Comportamento do Cão. Portugal. Europa- America.
O’Brien, S. J. & Johnson. W. E. (2007). A Evolução dos Gatos. Scientific American.63, 56-63.
Parcker, C. (1986). The ecology of sociality in felids. In: Ecological Aspects of social Evolution (Ed. By D. I. Rubenstein & R. W. Wrangham), (pp. 429-451). Princeton, New Jersey. Princeton University Press.
Pepperberg, I. M. (1991). Learning to communicate: the effects of social interaction. In: Perspectives in Ethology. (Ed. by P. J. B. Bateson & P. H. Klopfer), (pp. 119-164). New York. Plenum.
Plyusnina, I. Z., Oskina, I. N. & Trut, L. N. (1991). An analysis of fear and aggression during early development of behaviour in silver foxes (Vulpes vulpes). Applied Animal Behaviour
Science.32, 253-268.
Pongrácz, P., Miklósi, Á.., Kubinyi, E., Gurobi, K., Topál, J., & Csányi, V. (2001). Social learning in dogs. The effect of a human demonstrator on the performance of dogs (Canis
Povinelli, D. J., Bierschwale, D. T., & Cech, C. G. (1999). Comprehension of seeing as a referential act in young children, but not juvenile chimpanzees. British Journal of Developmental Psychology. 17(1), 37-60.
Price, E. O. (1984). Behaviour aspects of animal domestication. Quaterly Review in Biology, 59, 1-32.
Sandell, M. (1989). The mating tactics and spacing patterns of solitary carnivores. In: Carnivore Behavior, Ecology and Evolution (Ed. By J. L. Gittleman), (pp. 164-182). Ithaca, New York: Comstock Publishing.
Scheumann, M., & Call, J. (2004). The use of experimenter-given cues by South African fur seals (Arctocephalus pusillus). Animal Cognition, 7, 224–231.
Serpell, J. A. (1988). The domestication and history of the cat. In Turner, D. C. & Bateson, P. “The domestic cat”. 5th reprinting. (pp. 151–158). Cambridge. Cambridge University press.
Serpell, J.A. (1995). The Domestic Dog: Its Evolution, Behavior and Interactions with People. Cambridge. Cambridge University Press.
Soproni, K., Miklosi, A., Topal, J., & Csanyi, V. (2001). Comprehension of human communicative signs in pet dogs (Canis familiaris). Journal of Comparative Psychology. 115,
122-126.
Soproni, K., Miklosi, A., Topal, J., & Csanyi, V. (2002). Dogs’ (Canis familiaris)
Tomasello, M., Call, J., & Hare, B. (1998). Five primate species follow the visual gaze of conspecifics. Animal Behaviour, 55, 1063-1069.
Topál, J., Byrne, R. W., Miklósi, Á. & Csányi, V. (2006). Reproducing human actions and action sequences: “Do as I Do!” in a dog. Animals Cognition. 9, 355 – 367.
Trut, L. (1999). Early canid domestication: the farm-fox experiment. American Scientist, 87, 160–169.
Turner, D. C., Feaver, J., Mendl, M., & Bateson, P. (1986). Variation in domestic cat behaviour towards humans: A paternal effect. Animal Behaviour, 34, 1890–1901.
Virányi, Z., Topál, J., Gácsi, M., Miklósi, A. e Csányi, V. (2004). Dogs respond appropriately to cues of humans’ attentional focus. Behavioural Processes. 66, 161-172.
Yamaguchi, N., Driscoll, C. A., Kitchener, A. C., Ward, J. M. & Macdonald, D. W. (2004). Craniological differentiation between European wildcat (Felis silvestris silvestris), African
wildcat (F. s. Lybica) and Asian wildcats (F. s. Ornata): implications for their evolution and
conservation. Biological Journal of the Linnean Society.83, 47-63.
ANEXO 1
Questionário:
1) Quantos animais há em casa e de que espécie? 2) Qual a idade do gato, raça, sexo e nome? 3) Desde que idade o animal foi adquirido?
4) Ele tem todas as patas sem nenhuma deficiência? 5) Apresenta visão perfeita?
6) O animal sai de casa por algum motivo? 7) O animal interage com algum outro?
8) Quantas refeições ele faz por dia, que tipo de alimento costuma comer e qual o alimento preferido?
9) Ele aceita com facilidade estranhos em casa?
10) Seu animal é castrado? Se sim, com que idade foi feita a castração?