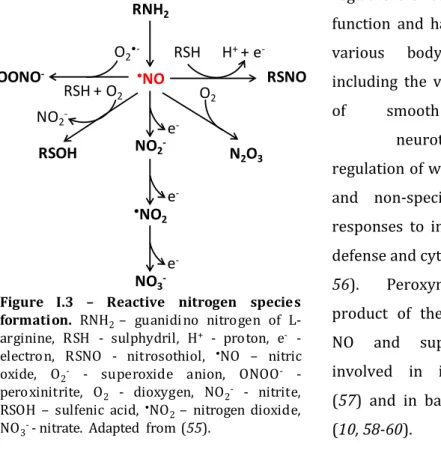
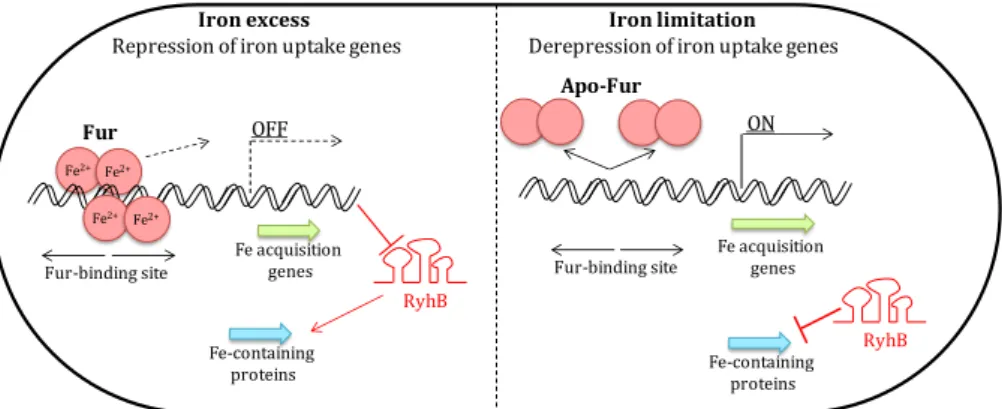
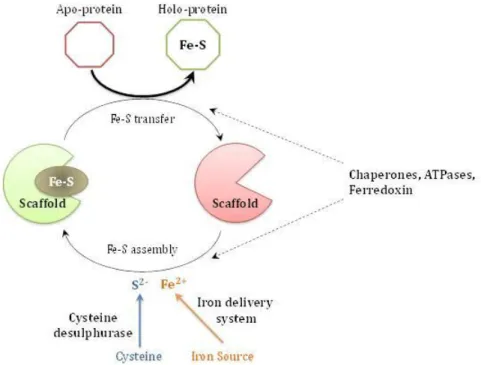
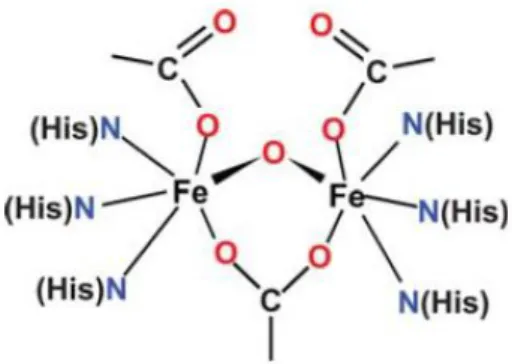
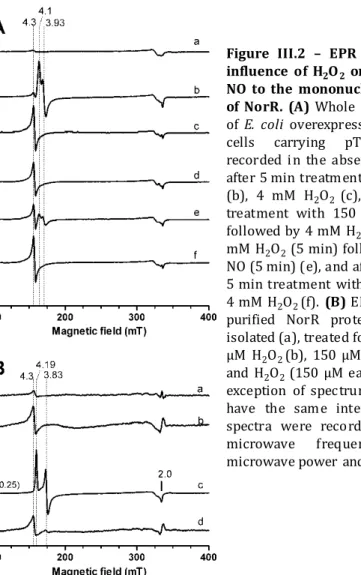
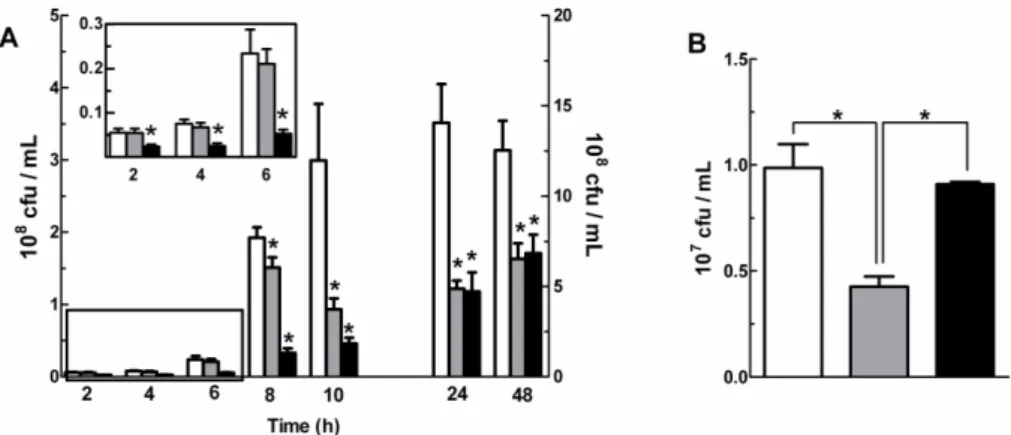
The Role of Di-iron Proteins in Pathogen Resistance
260
0
0
Texto
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)(11)
(12)
(13)
(14)(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)(29)(30)(31)
(32)(33)
(34)(35)
(36)
(37)
(38)
(39)
(40)
(41)
(42)
(43)
(44)
(46)
(47)
(48)
(49)
(50)
Imagem


+7


Documentos relacionados
redemocratização no Brasil, expressada pela Constituição de 1988 (BRASIL, 1988), estabeleceu um novo paradigma para as liberdades individuais, diferenciando-se do período de
67 Ora, é perante este conjunto de problemas, que dizem respeito aos domínios da história do Egipto, da ciência, do colonialismo, do Estado, da economia e da política
A teoria Kohlbergiana tem em linha de conta o problema das relações entre o pensamento moral e acção moral, que procura saber quando, como e em que circunstâncias é que
Relativamente à forma como homens e mulheres que passam por um ou mais tratamentos e homens e mulheres que não passaram por nenhum tratamento, caraterizam a relação
De acordo com os normativos vigentes, a avaliação é considerada como parte constitutiva da aprendizagem, um elemento integrante e regulador de toda a prática educativa,
O Estado na prossecução do interesse público e numa articulação entre contenção orçamental e necessidade de realização de grandes empreendimentos, tem optado
mulheres e os homens de forma diferente de acordo com o seu ESE - e um efeito prevalecente de sexo nos participantes do sexo masculino – avaliam mulheres e homens do mesmo ESE de
In a study performed by Barakat, Moustafa and Bikhazi (2012), it was found that 4 weeks of treatment with sodium selenite (5 ppm in drinking water) increased mRNA and protein
