UNIVERSIDADE DE LISBOA
Faculdade de Medicina
Lesão da Via Corticoespinhal e Variabilidade
no Recrutamento de Unidades Motoras
Artiom Viatcheslavovitch Poliakov
Orientador: Professor Doutor Mamede de Carvalho
Dissertação especialmente elaborada para obtenção do grau
de Mestre em Neurociências
2017
UNIVERSIDADE DE LISBOA
Faculdade de Medicina
Lesão da Via Corticoespinhal e Variabilidade
no Recrutamento de Unidades Motoras
Artiom Viatcheslavovitch Poliakov
Orientador: Professor Doutor Mamede de Carvalho
Dissertação especialmente elaborada para obtenção do grau
de Mestre em Neurociências
2017
A impressão desta dissertação foi aprovada pelo Conselho Científico da Faculdade de Medicina de Lisboa em reunião de 26 de setembro de 2017.
Abstract
The immense repertoire of movements that the human being is able to perform originates from the activity of several hundreds of skeletal muscles, which are under the control of the central nervous system. After processing the information regarding the spatial position of the body and the external environment, the motor centers of the brain and spinal cord produce the volitional commands responsible for the desired movements. This electrical neurotransference is done by the motor units.
Through recent electromyographic techniques, the study of the control of motor functioning by the nervous system, through the analysis of the action potentials of the motor units, becomes relatively simple. The force exerted by a muscle during a voluntary contraction is the result of the recruitment of motor units and the modulation of the frequency of its recruitment, which are altered in clinical situations associated with
corticospinal tract damage.
It is speculated that patients with amyotrophic lateral sclerosis (ALS) have a long preclinical phase before clinical manifestation; however, the current methods for detecting corticospinal tract involvement are complex and costly. The objective of this study was to evaluate the usefulness of a new method developed in our laboratory, which allows the analysis of the motor unit recruitment variability, in order to detect signs of corticospinal tract dysfunction.
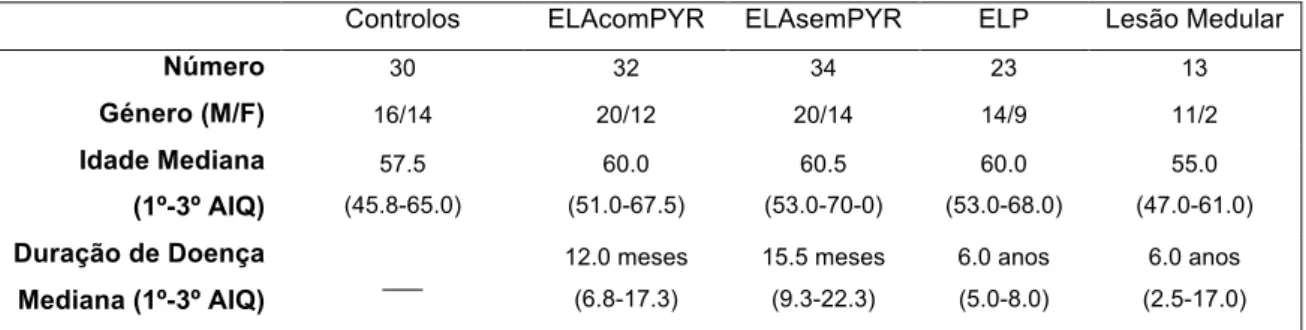
The study population consisted of 4 groups: healthy controls (30), patients with spinal cord injury (13), patients with primary lateral sclerosis (23) and patients suffering from amyotrophic lateral sclerosis (66). ALS group patients was split in patients with (32) and without upper motor neuron signs in lower limbs (34). The study of the recruitment of motor units was carried out using concentric needle electromyography in the tibialis anterior muscle, and advanced analysis of the motor unit recruitment
through the calculation of Mean Consecutive Difference of consecutive pairs.
It was found that in patients with lower limb spasticity, the first recruited motor units generally had lower recruitment frequencies when compared to healthy subjects; in addition, in these patients the frequencies of the second recruited motor units were closer to the frequencies of the first motor units compared to healthy individuals.
Although this observation is in agreement with the literature, the proposed method represents an original approach to investigate this topic. However, this novel technique does not present the necessary sensitivity to be applied in the clinical practice for diagnosis, unless further refinements are developed.
Key-‐words: Amyotrophic lateral sclerosis; Firing rate; Motor unit recruitment; Primary lateral sclerosis; Spinal cord injury.
Resumo
O imenso repertório de movimentos que o ser humano é capaz de realizar deriva da atividade de algumas centenas de músculos esqueléticos, estando estes sob o controlo do sistema nervoso central. Após o processamento das informações em relação à posição espacial do corpo e do meio exterior, os centros motores do cérebro e da medula espinhal emitem os comandos volicionais responsáveis pelos movimentos desejados. Esta neuro-‐transdução elétrica é mediada pelas unidades motoras.
Através de técnicas eletromiográficas recentes, o estudo do controlo motor pelo sistema nervoso, através da análise dos potenciais de ação das unidades motoras, torna-‐se relativamente mais simples.
A força exercida por um músculo durante uma contração voluntária é o resultado do recrutamento de unidades motoras e da modulação da frequência do seu recrutamento, sendo que ambas se encontram alteradas em casos de compromisso da via corticoespinhal.
Especula-‐se que os doentes com esclerose lateral amiotrófica (ELA) têm uma longa fase pré-‐clínica antes da manifestação da doença; no entanto, os métodos atuais de deteção da disfunção da lesão corticoespinhal têm uma complexidade e custos elevados.
O objetivo deste trabalho foi o de avaliar a utilidade de um novo método por nós desenvolvido, que analisa a variabilidade do recrutamento de unidades motoras, de forma a detetar sinais neurofisiológicos da disfunção da via corticoespinhal.
A população de estudo consistiu em 4 grupos: controlos saudáveis (30), doentes com lesão medular (13), doentes com esclerose lateral primária (23) e doentes com esclerose lateral amiotrófica (66). Estes foram classificados em dois grupos, com (32) e sem sinais piramidais nos membros inferiores (34).
O estudo do recrutamento das unidades motoras foi realizado com auxílio da eletromiografia de deteção com a utilização de uma agulha concêntrica no músculo tibial anterior, com o cálculo da Mean Consecutive Difference em pares consecutivos de potenciais para explorar a variabilidade no recrutamento de unidades motoras.
Verificou-‐se que em doentes com espasticidade nos membros inferiores, as primeiras unidades motoras recrutadas tinham, geralmente, menor frequência de recrutamento, quando comparadas com aquelas registadas em indivíduos saudáveis; para além disso, nestes doentes, as frequências de recrutamento das segundas unidades motoras encontravam-‐se mais próximas das frequências das primeiras unidades motoras, em comparação com indivíduos saudáveis.
Apesar dos achados observados estarem em concordância com o descrito na literatura, este novo método oferece uma alternativa para o estudo deste tema. No entanto, o método proposto não apresenta a sensibilidade necessária para uso regular na rotina neurofisiológica para a deteção de sinais de lesão da via piramidal; no entanto, o seu refinamento pode permitir esse fim.
Palavras-‐chave: Esclerose lateral amiotrófica; Esclerose lateral primária; Frequência de recrutamento; Lesão medular; Recrutamento de unidades motoras.
Índice
INTRODUÇÃO 10
NÚCLEOS MOTORES 13
UNIDADE MUSCULAR 15
PROPRIEDADES BÁSICAS DAS UNIDADES MOTORAS 19
PAPEL DO TRONCO CEREBRAL NAS PROPRIEDADES ELÉTRICAS DOS NEURÓNIOS
MOTORES 25
RECRUTAMENTO DE UNIDADES MOTORAS E FORÇA MUSCULAR 27
SIZE PRINCIPLE 28
TEORIAS CONTRADITÓRIAS DO SIZE PRINCIPLE 31
PADRÕES DE RECRUTAMENTO – “ONION SKIN” E “REVERSE ONION SKIN” 34
ELETROMIOGRAFIA DE SUPERFÍCIE 37
ATIVIDADE ESPONTÂNEA 41
ALTERAÇÕES FISIOLÓGICAS DAS PROPRIEDADES ELÉTRICAS DOS NEURÓNIOS
MOTORES 45
ALTERAÇÕES EM CASOS DE PATOLOGIA 49
LESÃO AO NÍVEL DA MEDULA ESPINHAL 49
LESÃO AO NÍVEL DO NERVO PERIFÉRICO 52
ESCLEROSE LATERAL PRIMÁRIA 53
ESCLEROSE LATERAL AMIOTRÓFICA 56
ANÁLISE ESTATÍSTICA DOS PADRÕES DE DESCARGAS DE UNIDADES MOTORAS EM
MÚSCULO ESQUELÉTICO HUMANO 67
CÁLCULO DO JITTER ELETROMIOGRÁFICO 68
OBJETIVOS 69
POPULAÇÃO DO ESTUDO 70
MÉTODOS 73
RESULTADOS 75
DISCUSSÃO 79
CONCLUSÕES E LIMITAÇÕES DO ESTUDO 82
BIBLIOGRAFIA 83
AGRADECIMENTOS 101
Introdução
Uma unidade motora pode ser definida como um “neuro-‐transdutor mecânico”, sendo responsável pela conversão de inputs nervosos aferentes e volicionais do córtex motor, na despolarização dos neurónios motores medulares ou do tronco cerebral, de forma a resultar em movimento. O substrato anatomo-‐funcional consiste em todas as fibras musculares inervadas pelo mesmo axónio, com origem num neurónio motor α. Estruturalmente, a unidade motora pode ser dividida em dois componentes: (i) neurónio motor e (ii) fibras musculares inervadas pelo axónio, podendo este último ser designado por unidade muscular. Todo este sistema normalmente funciona como uma única entidade (Heckman et al., 2012).
Figura 1 – Representação básica de uma unidade motora, em que um axónio ramifica-‐se para inervar um determinado número de fibras
musculares. Diferentes patologias ocorrem, em função do componente afetado (Kandel et al., 2000).
Um músculo típico é controlado por algumas centenas de neurónios motores cujos corpos celulares se encontram num núcleo motor da medula espinhal ou do tronco cerebral. O axónio de cada neurónio motor projeta-‐se da medula espinhal através da raíz anterior ou, no caso do tronco encefálico, através de um nervo craniano, sendo que se continua, no caso das raízes espinhais, por um nervo periférico, para chegar ao músculo alvo. O axónio ramifica-‐se antes de atingir o músculo, de forma a inervar um número muito variável de fibras musculares, de poucas a vários milhares, na depedência de cada axónio. (Kandel et al., 2000)
Assim que o input sináptico despolariza o potencial de membrana de um neurónio motor acima do seu limiar de excitabilidade, o neurónio gera um potencial de ação que é propagado ao longo do axónio até ao seu botão terminal, a nível muscular. O potencial de ação provoca a libertação de neurotransmissores na sinapse neuromuscular, o que leva a um novo potencial de ação no sacrolema das fibras musculares. Uma fibra muscular apresenta propriedades elétricas semelhantes a axónios de grande calibre não-‐mielinizados. Assim, ocorre a propagação do potencial de ação no sarcolema, mas, devido à alta capacitância elétrica deste, esta propagação dá-‐se a uma velocidade mais lenta que no axónio. Como sucede a despolarização síncrona de todas as fibras musculares de uma mesma unidade motora, o somatório das correntes extracelulares cria um potencial de ação composto das fibras musculares ativadas. (Kandel et al., 2000)
Devido à sincronia entre os potenciais de ação gerados a partir do neurónio motor e os da respetiva unidade muscular, estudar o controlo do
sistema pela ativação das unidades motoras é um processo exequível (Heckman et al., 2012).
O sistema motor depende da função de vários recetores e das suas subclasses, cada uma codificada por um gene específico. Um bom exemplo deste princípio é o neurotransmissor serotonina (5-‐HT) e as suas 14 sub-‐ classes de recetores (Nichols e Nichols, 2008). Enquanto que umas sub-‐ classes (5-‐HT2a, b e c) são importantes para os neurónios motores por aumentarem fortemente a sua excitabilidade, outras sub-‐classes (5-‐HT1b) apresentam efeitos inibitórios em alguns interneurónios espinhais. Daí, a libertação de serotonina poder apresentar tanto um efeito excitatório como um efeito inibitório, dependendo do subtipo de recetor neuronal envolvido (Heckman et al., 2012).
Núcleos motores
Individualmente, cada músculo contém uma população de unidades motoras que controla a força exercida por este durante a contração. Como já se referiu anteriormente, os motoneurónios estão localizados no corno anterior da medula espinhal ou do tronco encefálico. No seu conjunto, os neurónios motores responsáveis pela inervação de um mesmo músculo estão dispostos em colunas na medula, conhecidas como núcleos motores (Burke et al., 1977; Routal et al., 1999). Numa secção transversal, os núcleos motores de músculos proximais tendem a estar localizados numa posição mais mediana em comparação com os núcleos motores de músculos distais (Datta et al., 1991; Jenny e Inukai, 1983).
O número de unidades motoras presentes num músculo pode variar de dezenas a várias centenas; contudo, estes números não são fáceis de determinar com confiança dado que cada um dos métodos de avaliação existentes na atualidade tem as suas limitações:
1. Um dos métodos utilizados baseia-‐se na contagem de neurónios motores marcados com o transportador retrógrado horseadish
peroxidase (HRP). Isto pode ser feito através da injeção direta no
músculo alvo ou através da exposição do nervo (Burke et al., 1977). A principal dificuldade desta técnica é o fornecimento de HRP suficiente ao músculo alvo, sem envolver os outros músculos adjacentes.
2. Outro método baseia-‐se na dissecção de cadáveres humanos e na contagem dos axónios motores dos nervos periféricos, a dificuldade reside na destrinça entre axónios motores e sensitivos; a estimativa resultante desta técnica é semelhante à da técnica anterior (Feinstein et al., 1955).
3. Um terceiro método deriva da medição da amplitude de potenciais obtidos através da estimulação de nervos periféricos (McComas,
1995; McComas et al., 1971). Normalmente a estimativa de contagem dos neurónios motores obtidos através destes métodos eletrofisiológicos é inferior aos dos métodos baseados na observação anatómica, para além de apresentar uma variabilidade considerável (Van Cutsem et al., 1997). Esta abordagem neurofisiológica tem a potencialidade de aplicação em doentes com patologia neuromuscular (Boe et al., 2010; Porter et al., 2008).
Ao longo dos anos, inúmeros métodos neurofisiológicos têm sido descritos. Contudo, a sensibilidade destes métodos para a avaliação de variações fisiológicas, como o envelhecimento, tem sido questionada.
Unidade muscular
1. Razão de Inervação
Através de técnicas de dissecção de cadáveres foi possível obter informação sobre o número médio de fibras musculares numa mesma unidade motora: a razão de inervação (número de fibras musculares inervadas por um único axónio) varia entre 5 a 10, para músculos como o reto lateral ou o músculo tensor da corda do tímpano (Blevins, 1967; Carlsoo, 1958), até cerca de 1750 para os grandes músculos dos membros inferiores como o gémeo interno ou o quadrícepete (Feinstein, 1955). A razão de inervação correlaciona-‐se com a precisão do movimento; quanto menor é este número maior é a precisão motora conseguida (Kandel et al., 2000).
Apesar destes dados terem utilidade na comparação entre os músculos, são as variações encontradas entre as unidades motoras que vão possibilitar a compreensão da forma com que o sistema nervoso controla a força muscular. Estudos de depleção de glicogénio indicam que a razão de inervação varia entre os tipos de unidades motoras, sendo este número menor em unidades motoras com baixo limiar de excitabilidade (Bodine et al., 1987; Burke e Tsairis, 1973). Esta razão de inervação entre as diferentes unidades motoras pode ainda ser estimado a partir da medida da força muscular das unidades motoras (Bodine et al., 1987; Kanda e Hashizume, 1992). Utilizando esta abordagem, baseada na determinação de forças tetânicas, Enoka e Fuglevand (2001) estimaram que a razão de inervação para as 120 unidades motoras no primeiro músculo interósseo dorsal humano variava entre 21 a 1770. Consequentemente, a unidade motora mais forte neste músculo tem a capacidade de exercer uma força semelhante a uma unidade motora típica do músculo gémeo interno (Kandel et al., 2000).
Tabela 1 – Representação dos números da razão de inervação no homem (Kandel et al., 2000).
2. Território de inervação
As fibras musculares inervadas por um único neurónio motor ocupam um determinado subvolume do músculo, ao invés de se extenderem ao longo de toda a sua extensão (Hijikata e Ishikawa, 1997; Lateva e McGill, 2001; Young et al., 2000). Através de técnicas eletrofisiológicas e com o objetivo de identificar o territórios das unidades musculares do costureiro (o músculo mais comprido do corpo humano), Harris e colegas (2005) verificaram que, dos 64 potenciais de ação das unidades motoras analisados em cinco indivíduos, apenas 21 foram detetados nas extremidades mais proximal e distal do músculo. As restantes 43 unidades motoras, num mesmo fascículo, apenas ocupavam o início ou a terminação.
Figura 2 – Distribuição das unidades motoras e seus territórios ao longo do músculo costureiro. (A) Posicionamento dos elétrodos (R1-‐R5) ao longo do comprimento do músculo. (B) Localização das placas motoras (círculos preenchidos a negro) e a extensão, no músculo, sobre a qual foi possível registar os potenciais de unidades musculares singulares (cada barra cinzenta representa uma única unidade motora) (Harris et al., 2005).
Um outro estudo baseou-‐se em métodos eletromiográficos (registo intramuscular e superficial) do músculo gémeo interno. Os resultados demonstraram que a área dos territórios de uma unidade motora era inferior a 2,5 cm, o que equivale a aproximadamente 10% do comprimento total do músculo (Vieira et al., 2011).
É importante notar que a distribuição das unidades motoras no músculo pode sofrer alterações como consequência da idade e de doenças neuromusculares. Por exemplo, as fibras musculares perdem a sua inervação com a morte do neurónio motor; no entanto, existem mecanismos que possibilitam a re-‐inervação destas fibras por terminações axonais localizadas na sua vizinhança, possibilitando assim o restabelecimento (completo ou não) da função (Kandel et al., 2000).
Em alguns músculos, os territórios das unidades musculares podem ser limitados a compartimentos pequenos, dentro do próprio, sendo cada um destes compartimentos inervado por um ramo nervoso específico (Peters, 1989; Windhorst et al., 1989). Como resultado desta organização, um único músculo pode conter várias regiões distintas, cada uma apresentando um função fisiológica diferente (Butler et al., 2005). Por exemplo, o músculo bícipite braquial é inervado por 3-‐5 ramos nervosos primários do nervo musculo-‐cutâneo, e as unidades motoras na longa porção do bicípete parecem funcionar em duas populações distintas. As unidades motoras na porção lateral deste músculo sofrem ativação quando uma força de flexão é exercida pela articulação do cotovelo; as unidades motoras da porção mediana sofrem ativação quando o músculo produz um movimento de supinação; as unidades motoras na zona intermediária sofrem ativação por ambos – flexão e supinação (Barry, 2009).
Propriedades básicas das unidades motoras
A força exercida por um músculo depende, não só do número das unidades motoras ativadas durante a sua contração, como também de 3 propriedades referentes a estas: velocidade de contração, força máxima do “twitch” e fatigabilidade. A avaliação destas propriedades é possível através da análise da força exercida por unidades motoras individuais em resposta a potenciais de ação.
A contração isolada de uma unidade motora é definida pela força (“ force of
twitch contraction”) e pela sua duração (tempo de contração) -‐ tempo
necessário para que o “twitch” exerça a força máxima. Por exemplo: unidades motoras lentas (“slow-‐twitch”) apresentam tempos de contração longos quando comparadas com unidades motoras rápidas (“fast-‐twitch”). Uma série de potenciais repetidos num curto espaço de tempo, originam um conjunto de contrações sobrepostas – contração tetânica.
De modo a quantificar a associação entre as propriedades histológicas de um músculo e as suas propriedades contráteis, vários fisiologistas têm vindo a desenvolver esquemas para possibilitar a identificação dos diferentes tipos de fibras musculares. Um dos critérios de classificação mais conhecido identifica três isoformas (I, IIA e IIX) da cadeia pesada da miosina, o que influencia a velocidade de contração máxima, sendo a MyHC I a mais lenta e a MyHC IIX a mais rápida (Bottinelli e Reggiani, 2000; Harridge et al., 1996).
Uma das primeiras tentativas de distinção entre unidades motoras foi proposta por Burke e colegas, que desenvolveram um método de classificação de forma a identificar os seus diferentes tipos (Burke et al., 1973; Burke et al., 1974). Os critérios de classificação consistem na determinação do perfil de uma contração tetânica incompleta, e
posteriormente na medição do tempo de declínio da força tetânica seguindo um protocolo de estimulação elétrica. O intervalo entre os estímulos numa contração tetânica incompleta foi definido como 1.25x o tempo de contração da unidade motora. As unidades motoras com decréscimo da força foram classificadas como unidades motoras de contração rápida (tipo F, de “fast”), enquanto que as unidades motoras sem redução foram classificadas como de contração lenta (tipo S, de “slow”). As unidades tipo F foram posteriormente sujeitas a 6 minutos de estimulação elétrica intermitente (40 Hz durante 330 ms, uma vez por segundo) e a redução dos picos de força tetânica foi critério na distinção entre unidades motoras de fadiga rápida (tipo FF, “fast fatigue”) e resistentes à fadiga (tipo FR, “fatigue resistant”).
No entanto, estudos subsequentes têm vindo a demonstrar que as propriedades das fibras musculares e das unidades motoras não são tão distintas como se calculava (Kernell, 2006): (1) as fibras musculares podem co-‐expressar combinações de isoformas de MyHC, como as tipo I e IIa ou IIa e IIX (Andersen e Aagaard, 2010; Bottinelli, 2001); (2) a redução da força apresenta uma elevada variabilidade (Carp et al., 1999); e (3) o protocolo usado para avaliação da resposta à fadiga utilizou a estabilidade da força durante contrações intermitentes, com base nas capacidades oxidativas enzimáticas, o que tende a desprezar outros fatores que podem levar à fadiga durante as contrações voluntárias (Enoka et al., 2011). Mesmo quando as unidades motoras humanas sofrem ativação através da estimulação intraneural, não é possível identificar os vários tipos distintos com base nos critérios propostos por Burke e colegas (Fuglevand et al., 1999; Thomas et al., 1991).
A força exercida durante uma contração tetânica depende fortemente da sincronia e somatório dos vários “twitches”, em relação com o tempo de
contração da unidade motora e com a frequência de resposta. A força máxima produzida durante uma contração tetânica pode ser caracterizada como uma função sigmóide, dependendo da frequência, sendo que a forma da curva também se relaciona com o tempo de contração da unidade motora.
Figura 3 – Traçado representando a força exercida por unidades motoras de contração rápida (fast-‐twitch) e lenta (slow-‐twitch), em resposta a um único potencial de ação (2 traçados superiores) e a múltiplos potenciais de ação de frequência variável (2 traçados inferiores). O intervalo de tempo necessário para atingir a força máxima é inferior nas unidades rápidas, sendo que com frequências de 100 Hz, esta é superior à força máxima das unidades lentas (Kandel et al., 2000).
A força máxima das unidades motoras rápidas é sempre superior à das
lentas, para qualquer frequência. No entanto, as unidades lentas
conseguem atingir forças relativas (percentagem da força máxima) superiores, em resposta a baixas frequências.
Figura 4 – Representação da força máxima e da força relativa (como percentagem da força máxima) em função da frequência de múltiplos potenciais de ação, para unidades motoras fast-‐twitch (a vermelho) e slow-‐ twitch (a azul). Com base no primeiro gráfico é possível observar a diferença considerável entre a força máxima dos 2 tipos de unidades motoras, sendo que esta é superior nas unidades rápidas, para qualquer frequência. Contudo, em termos de força relativa, o contrário acontece, verificando-‐se valores superiores, para as mesmas frequências, em unidades lentas, até se atingir os 100 Hz (frequência à qual tanto as unidades rápidas como as lentas atingem quase 100% da sua força máxima) (Kandel et al., 2000).
De modo a analisar mais profundamente o efeito da alteração da frequência na produção de força de uma unidade motora em contração isométrica, foi utilizada uma simulação recorrendo a um modelo que continha um conjunto de 200 unidades motoras (Duchateau e Enoka, 2002). Para isso foram inseridas, neste modelo, as propriedades mecânicas (força máxima e tempo para atingir a força máxima) através do método neurofisiológico denominado de “spike-‐triggered averaging” do músculo tibial anterior (Van Cutsem et al., 1998). Foi possível observar um aumento significativo da força de todas as (200) unidades motoras com o aumento da frequência de estímulo para valores de 100-‐200 Hz (figura 5). Contudo, o posterior aumento destas frequências para valores superiores a 100-‐200 Hz tem uma baixa influência na força de contração de unidades motoras mais rápidas (100 e 200); isto reflete uma diferença entre as propriedades relacionadas com a velocidade entre as unidades motoras de
Figura 5 – Simulação da relação entre a frequência de estímulo e a força relativa (em relação à MVC) para a 1ª, 100ª e 200ª unidade motora de um conjunto de 200 unidades no músculo tibial anterior. A força produzida por cada unidade motora foi simulada por 4 descargas sucessivas geradas a frequências constantes (variando de 10 a 500 Hz) (Duchateau e Baudry, 2014).
As propriedades funcionais das unidades motoras apresentam variações interpessoais e intermusculares. De um lado deste amplo espectro encontram-‐se as unidades motoras de contração lenta, com baixa força de contração e resistentes à fadiga. Estas são as primeiras a ser ativadas durante contrações voluntárias. Do outro lado do espectro encontram-‐se as unidades motoras rápidas, com elevada força de contração e suscetíveis à fadiga. Estas são as últimas a ser ativadas durante contrações voluntárias.
Num estudo feito por Duchateau e colegas, com o objetivo de analisar as propriedades das unidades motoras no músculo tibial anterior, foram estimadas as propriedades contrácteis de um total de 528 unidades motoras em 10 sujeitos, aquando da realização de contração voluntária (Van Cutsem et al., 1997). Os limiares de recrutamento das unidades
motoras apresentaram uma variação entre 1% a 90% da contração voluntária máxima (MVC) para os músculos dorsiflexores, em que cerca de metade das unidades motoras apresentavam limiares de recrutamento inferiores a 20% da MVC.
Figura 6 – Representação da percentagem do número de unidades motoras total analisadas (n = 528) do músculo tibial anterior, em função da força de contração (A) e do tempo de contração (C). Verifica-‐se que a maior parte das unidades motoras apresenta baixas forças de contração (entre os 0 e os 40 mN/m) e tempos de contração intermédios (entre os 30 e os 60 ms); (B) Relação entre a força de contração e o limiar de recrutamento (em percentagem da MVC); (D) Relação entre o tempo até atingir a força máxima e o limiar de recrutamento (em percentagem da MVC) (Van Cutsem et al., 1997; Heckman e Enoka, 2012).
Devido ao facto das propriedades contrácteis de uma unidade motora dependerem das características das fibras musculares, é possível realizar a distinção entre estas. Esta distinção é baseada em aspetos estruturais e nas propriedades metabólicas das fibras musculares. Todas as fibras musculares pertencentes a uma unidade motora apresentam propriedades bioquímicas e histoquímicas semelhantes.
Papel do tronco cerebral nas propriedades elétricas dos neurónios motores
A frequência de recrutamento dos neurónios motores depende da magnitude de despolarização criada pelos inputs excitatórios e das propriedades intrínsecas da membrana dos neurónios motores na medula espinhal. Estas propriedades podem ser modificadas por inputs provenientes de neurónios monoaminérgicos do tronco cerebral, sendo que na ausência destes inputs, as dentrites dos neurónios motores transmitem passivamente uma corrente sináptica ao corpo celular, o que resulta numa despolarização. Nestas condições, observa-‐se uma relação linear entre a corrente (input) e a frequência de recrutamento. Esta relação torna-‐se não-‐linear quando as monoaminas serotonina e norepinefrina ativam os canais de cálcio tipo-‐L ao nível das dendrites dos neurónios motores. As correntes de Ca2+ resultantes destas interações têm
a capacidade de aumentar as correntes sinápticas até cerca de 10 vezes, sendo que num neurónio motor ativo esta corrente consegue sustentar uma frequência de depolarização elevada após um rápido input de despolarização – self-‐sustained firing (este mecanismo é responsável pela manutenção da postura muscular). Após um input inibitório subsequente, a frequência volta aos seus valores originais (Kandel et al., 2000).
Durante o sono, a atividade monoaminérgica sofre uma diminuição muito acentuada resultando num decréscimo de excitabilidade, favorecendo o relaxamento muscular. Assim, os inputs monoaminérgicos desempenham um importante papel na modulação da atividade dos conjuntos de neurónios motores necessários para o desempenho de diferentes tarefas do sistema locomotor. Esta “flexibilidade”, contudo, não compromete o size
principle (este irá ser analisado mais à frente) dado que os limiares de
nos neurónios motores de unidades motoras mais lentas (que são recrutadas em primeiro lugar, mesmo na ausência de input monoaminérgico) (Kandel et al., 2000).
Recrutamento de unidades motoras e força muscular
A força exercida por um determinado músculo, durante uma contração, depende do número de unidades motoras ativadas e da frequência do seu recrutamento. Esta força aumenta durante uma contração muscular através da ativação de unidades motoras adicionais, que são recrutadas progressivamente da mais “fraca” para a mais “forte”. O limiar de recrutamento de uma unidade motora é a força, durante a contração, a partir da qual a unidade motora é ativada. O decréscimo da força muscular também é gradual, inativando-‐se primeiro as unidades motoras mais “fortes” e só no final as mais “fracas” (Kandel et al., 2000).
Figura 7 – Unidades motoras que exercem menos força são recrutadas antes do recrutamento de unidades motoras que exerçam força superior. (A) Medição de potenciais de ação de duas unidades motoras, recorrendo a um único elétrodo intramuscular, à medida que o sujeito aumenta gradualmente a força. Recrutamento da unidade motora 1 é evidente quase desde o início do movimento, notando-‐se ainda um aumento na sua frequência. O recrutamento da unidade motora 2 é apenas evidente no final da contração voluntária. (B) Representação da força de contração média das duas unidades motoras analisadas (Desmedt e Godaux, 1977).
Já desde as pesquisas pioneiras de Adrian e Bronk (1929) existe o conhecimento da relação entre a força exercida por um músculo e o número e recrutamento de unidades motoras. As unidades motoras são
recrutadas consoante uma sequência ordenada (Denny-‐Brown e Pennybacker, 1939) que depende das diferenças dos tamanhos dos neurónios motores (Henneman, 1957) e do aumento da força de contração (Thomas et al., 1987). A fadiga irá reduzir os limiares de recrutamento das unidades motoras menos excitáveis (maiores), mesmo estas tendo um envolvimento mínimo na tarefa (Farina et al., 2009). No entanto, a ordem de recrutamento não é influenciada por fatores como o envelhecimento (Fling et al., 2009), imobilização (Duchateau e Hainaut, 1990), fadiga muscular (Adam e De Lucca, 2003) ou treino físico (Van Cutsem et al., 1998).
Size Principle
A ordem pela qual as unidades motoras são recrutadas tem uma estreita relação com o tamanho dos corpos celulares dos neurónios motores, com o diâmetro e velocidade de condução axonal, e com a força exercida pelas fibras musculares. Devido ao facto do limiar de recrutamento de uma unidade motora depender da resistência da membrana do neurónio motor, o que é inversamente relacionado com a sua área da superfície, uma corrente sináptica irá produzir maiores alterações no potencial de membrana de neurónios motores de pequeno diâmetro. Assim, com o aumento gradual do input excitatório, os neurónios motores mais pequenos são recrutados primeiro e os maiores em último lugar. Este fenómeno é conhecido como “size principle”, um princípio enunciado por Elwood Henneman (1957).
Este princípio apresenta duas consequências importantes para o controlo do movimento, por parte do sistema nervoso: (1) a sequência de recrutamento de neurónios motores é determinada pelos mecanismos espinhais e não por segmentos supra-‐espinhais do sistema nervoso, ou
seja, o córtex não tem controlo seletivo sobre unidades motoras específicas; (2) as unidades motoras são ativadas por ordem crescente de fatigabilidade, sendo que as unidades motoras menos fatigáveis são as primeiras a serem ativadas de modo a produzir a força necessária para desempenhar determinada tarefa.
A força necessária para o recrutamento da última unidade motora varia entre os diferentes músculos: em alguns músculos da mão, durante uma contração lenta, todas as unidades motoras são recrutadas quando a força atinge cerca de 60% da força máxima; no entanto, em músculos como o bícipite braquial, deltóide e tibial anterior, o recrutamento só termina quando se atinge cerca de 85% da força máxima. Esta hipótese foi sugerida por Edgar Adrian (1925) e foi posteriormente comprovada.
Contudo, devido ao facto do limiar de recrutamento das unidades motoras diminuir com o aumento da velocidade de contração, durante um movimento de contração rápida grande parte das unidades motoras são recrutadas com uma carga de aproximadamente 33% do máximo. Além do recrutamento, a força muscular pode ser aumentada através da variação da frequência de recrutamento dos neurónios motores (Kandel et al., 2000).
Figura 8 – Força muscular pode ser ajustada através da variação do número de unidades motoras recrutadas e da sua frequência de recrutamento. Um aumento gradual, seguido de uma diminuição gradual da força exercida por um músculo responsável pela extensão do joelho (linha azul), teve influência da ativação de 4 (de entre muitas outras) unidades motoras (linhas vermelhas). Observa-‐se que a força muscular varia com o número das unidades motoras ativas e com a sua frequência de recrutamento (Kandel et al., 2000).
Existem vários mecanismos de feedback cujo objetivo é a regulação da frequência de recrutamento das unidades motoras. Com baixas frequências, a regularidade do recrutamento das unidades motoras são controladas pela atividade dos fusos musculares (Vallbo, 1974). Considera-‐se que as aferências provenientes dos fusos musculares reforçam a atividade dos neurónios motores α e sensibilizam o sistema motor aos inputs sensitivos. Kanda e Desmedt (1983) observaram que o
feedback aferente cutâneo tem influência na ordem de recrutamento das
unidades motoras, fazendo com que unidades com alto limiar de recrutamento apresentassem um limiar mais baixo; como consequência,
unidades motoras de baixo limiar nunca se apresenta completamente regular (Rosenfalck e Andreassen, 1980), especialmente quando apresentam uma atividade muito baixa; esta característica foi relacionada com a variação espontânea na atividade elétrica das membranas dos neurónios motores e irregulariedades (a curto prazo) na curva de despolarização neuronal (Stålberg et al., 1973). De um ponto de vista geral, as unidades motoras de músculos com inervação lombar apresentam um padrão de atividade mais lento e regular que as unidades motoras de músculos dos membros superiores (Stålberg et al., 1973). A variabilidade do recrutamento de unidades motoras de baixo limiar é superior em indivíduos de idade avançada, quando comparada com indivíduos jovens (Tracy et al., 2005), o que pode ser explicado pelos AHP (afterhyperpolarization potentials) mais longos associados ao aumento da idade (Piotrkiewicz et al., 2007). Para além disso, esta variabilidade diminui com o aumento progressivo da força muscular (de Carvalho et al., 2014).
Teorias contraditórias do Size Principle
Apesar do consenso geral sobre o recrutamento das unidades motoras durante contrações voluntárias ocorrer seguindo uma ordem que tem a ver com o “size principle”, foram feitas várias sugestões que certas condições requerem um ajustamento a este esquema, não sendo este tão linear. A noção da possibilidade de recrutar uma unidade motora seletivamente e, ao mesmo tempo, inibir o recrutamento de outras unidades motoras de baixos limiares de ativação foi um aspeto importante para fortalecer a ideia anterior (Ashworth et al., 1967).
Outra hipótese que surgiu sugere que em contrações rápidas poderá haver recrutamento preferencial de unidades motoras de contração rápida (Grimby e Hannerz, 1977; Tanji e Kato, 1973). Contudo, Desmedt e Godaux
(1977) demonstraram que, apesar do limiar de recrutamento decrescer consoante a velocidade de contração (van Bolhuis, 1997), a ordem de recrutamento das unidades motoras nos músculos tibial anterior e primeiro dorsal interósseo é a mesma durante contrações lentas e rápidas. Uma terceira hipótese proposta analisa a possibilidade da alteração da ordem do recrutamento das unidades motoras em ações que envolvem a alteração do comprimento muscular (Maton, 1980). Dado que o input sináptico recebido por um núcleo motor difere para contrações isométricas e anisométricas (Tax et al., 1989; van Bolhuis, 1997), existe a possibilidade da alteração da ordem de recrutamento durante contrações de encurtamento e alongamento muscular. Foi demonstrado que as unidades motoras envolvidas numa ação de levantamento de uma carga através de contrações concêntricas e de abaixamento da mesma carga através de contrações excêntricas, por parte dos músculos posteriores da perna, eram diferentes nas duas ações (Nardone et al., 1989). Grande parte das investigações subsequentes falharam em demonstrar estas diferenças na ordem de recrutamento entre contrações concêntricas e excêntricas (Christova e Kossev, 2000; Pasquet et al., 2006). Estes resultados sugerem que, apesar da ordem de recrutamento poder diferir em algumas tarefas, esta permanece relativamente constante nos diferentes tipos de contração. Ao contrário das três condições anteriores, existe maior evidência científica de que a ordem de recrutamento das unidades motoras num determinado músculo pode diferir quando este músculo contribui para diferentes ações (Butler, 2005). Por exemplo, o músculo primeiro dorsal interósseo produz grande parte da força de abdução exercida pelo segundo dedo, mas é apenas um de vários músculos que contribui para a flexão do mesmo dedo ao nível da articulação metacarpo-‐falângica. Desmedt e Godeaux (1981) estabeleceram que cerca de 8% dos 142 pares de unidades motoras no primeiro dorsal interósseo tinham a capacidade de alterar de forma consistente a ordem de recrutamento entre as duas
ações descritas anteriormente, o que foi posteriormente confirmado durante contrações estáticas (Thomas et al., 1986) e dinâmicas (Thomas et al., 1987). Devido ao forte coeficiente de correlação entre as intensidades das forças resultantes e os limiares de recrutamento durante a tarefa de abdução, quando comparados com a tarefa de flexão, Desmedt e Godeaux concluíram que a distribuição dos inputs centrais para os núcleos motores era diferente para as duas ações. No entanto, para músculos do tronco, esta evidência científica já não se apresentava tão forte (Westgaard e De Luca, 2001).
Outra condição capaz de influenciar a ordem de recrutamento é a manipulação do feedback recebido pelos neurónios motores. Garnett e Stephens (1981) descobriram que a estimulação do nervo digital do segundo dedo da mão aumenta o limiar de recrutamento de unidades motoras de baixo limiar, e diminui o limiar de recrutamento em unidades motoras de elevado limiar, no músculo primeiro dorsal interósseo. Posteriormente, Kanda e Desmedt (1983) relataram que, os movimentos de esfregar a ponta do segundo dedo na ponta do primeiro dedo, poderiam levar a alterações tanto no limiar como na ordem de recrutamento de unidades motoras de alto limiar de ativação no primeiro dorsal interósseo, durante um movimento preciso. A interpretação que se obteve com estas observações foi a existência de uma distribuição preferencial de inputs cutâneos excitatórios para as unidades motoras de alto limiar (pelo menos no caso do músculo analisado). Algumas estimativas em relação à distribuição dos inputs sinápticos sugerem a consistência de aferentes cutâneos de baixo limiar com este esquema (Burke, 1970; LaBella et al., 1989); contudo, nem todos os estudos chegaram ao mesmo resultado (Erim et al., 1996; Jones e Lee, 2006).