A
ecologia
trófica do
Bufo-re
al
Bubo
bubo
e
as implicações conservacionistas
em
ecossistemas
med
iterrân
icos
t
Rui
Nascimento
Fazenda
Lourenço
Dissertação apresentada
para
obtenção do grau de Mestre em Biologia da ConservaçãoOrientador:
Prof.
João
E.
Rabaça
Universidade de Évora 2005
A
ecologia
trófica do
Bufo-real
Bubo
bubo
e
as
implicaçÕes
conservacionistas
em
ecossistem
as
m
editerrân
icos
Rui
Nascimento
Fazenda Lourenço
,l
ÇY6gs
Dissertação
apresentada para obtenção
do grau deMehe
em
Biolqia
daConservação
Orientador:
Prof.
João E. Rabaça
Universidade
de
Évora2005
\.['",
)
A ecotogia
úófrca
do Bufo+eal
Bubo
hubo e
as implicações
Gonaervacionistas
ern
ecosisternas
medltemânicos
Rui Fazenda Lourenço
lndice
Resumo.... Abstract... 1 2 3 1. Infrodução2- lníormadã|o biológica e aspectos da conservação do Bufo+eal em ecossisêmas meditenânicos
2.í.
Descrição2.2. Biogeografia, distribuição e situa@o populacional " " " " " "
"
2.4. Comportamento... 2.5. Ulilzação dos biótoPos
4 4 4 7 7
I
2.6. Biologia dareprodução...-2.7.Fadores de
ameap
e @usÍls de mortalidade-. 2.8. Critérios de conservação e instrumentoslqais
2.9. Ecologia tróÍica2.g.1.lntrodução teórica sobre a ecologia trófica---.---2.9.2. Ecologia trófica do Búo-real
4.lveade
estudo5. Os hábÍtos alimentares do Bufo-realna Península lbérica.
6.
lmplica@s
onservacionistas dos hábitos alimentares do Bufo-realnoSulde
Portugal 277. Conclusões finais 48 50 8. Referências
bibliográfies
oI
I
I
11 12 14Rui Fazenda Lourengo (2ü15)
Resumo
O Bufo-real (Bubo bubol é um dos principais predadores de topo na PenÍnsula lbárica e uma espécie classiftcada como Vulnerável em Portugal (lCN rn
pess). A
conservação desta ave em ecossistemas medÍtenânicos depende da existência de conhecimento de base sobre asula biotogia, nomeadarnente a ecologia trófica. A predação, sobretudo porespécies detopo,
é
um mecanismo fundamentaldo
funcionamento dos ecossistemas.Os
objedivos deste estudo são(í)
conhecer as estratrágias alimentares do Bufo-real na PenÍnsula lbéricae
(2) analisar as consequências da sua dieta noSulde
Portugalem termos de conservaçâo.O
estudo desenrolou-seno
Sulde
Portugal, mais especificamente nas áreasdo
Parque Naturalda
Sena
de
São
Mamede,Bacia
Médiado
Guadiana, Semade
Monfurado e Nordeste Algarvio.Os
casais
de
Bufo+eal
foram
detectados
com
recurso
a
escuta
de
vocalizaçõesespontâneas
e
reprodução
de
vocal2ações
conspecíficas.As
amostras
de
restos alimentares foram recolhidos nos ninhos e poisos utilizados pelas aves. Aidentifieção
das presasfoi
realizada como
apoiode
colecçõesde
referênciae
chaves especÍficas, tendo sido calculados: a frequência na dieta e o contributo em termos de biomassa. As estratégias alimentares na Península lbérica foram estudadas utilizando os dados obtidos para Portugale os
resultados publicados relativosa
diversas regiõesde
Espanha, com recursoa
uma análisede
componentes principais (PCA).A
relação entre as frequênciasdas
presas foi analisada reconendo à conelação de Spearman.Os principais resultados obtidos apontam para a existência de três estratégias alimentares
na
PenÍnsula lbérica: dietas baseadasem
lagomorfos; dietas baseadasem
roedores; e dietascom
maior diversidademas cuia presa
principalsão
os
lagomorfos.No
Sul
de Portugal,os
lagomorfossão
as
presas
principais, sobretudoo
coelho.Alguns
casaisalimentam-se sobretudo
de
lagomorfos,
enquanto
outros
apresentam
dietas
mais diversificadas, embora os coelhos e lebres sejam a base alimentar. As aves de médio porte, os ouriços e os roedores são asprsas
complementares mais oomuns.O Bufo-realdepende dos lagomorfos pelo que é essenciala manutenção da disponibilidade destas presas, fundamentais a diversos predadores ibéricos. O efeito reguladordo Bufo+eal
é
mal conhecido, sendo importante estudar asintera@es
com as suas presas, a maioriaespécies cinegéticas,
mas
também
meso-predadoresou
espécies
de
importância conseruacionista.A
conserva@o desta espécie pode passar, em grande medida, por uma conecta gestão dos habitats incluÍdos no tenitório dos casais, de modo a permitir uma maior disponibilidade de presas.tutr F aenda Lourenço (2(X)O
A ecdogia útrca doBticrrell &fu bufu e
a
Ímpli,cações conservacion'lstre sn eccsisteínc medÍterânícos.Abstract
The food
habits
of the
Eagle Owl
BuÔo Ôuboand
the
conservation implications inmediteranean ecosystems
The Eagle Orrvl
(Buôo
bubo) is one of the main top predators living in the lberian Peninsula and a species considered vulnerable in Porhrgal (lCN ,n pre§s).Theonservation
of thisod
in medi,tenanean ecosystems depends upon knowing its lífe history, namelly the food habitrs,onoe larga predatorc have an active role in
food
chains. The aims of thÍs study consist in understanding feeding strategies of lberian Eagleolrls
and analysing diet consequences on conservation in SouthemPortugal-The study was conduçted in Southem Portugat, in the Sena de São Mamede Natural
PaK
the Mld
Guadiana River Basinand
Monfurado(Cenfat
Alentejo), andthe
North+astem Algarve.Eagle 6xul pairs were detected by spontaneous calls and response to playback of conspeciftc calts. Feeding remains were collected in nests and perches and identified using reference collections
and specific studies.
Frequencyin
the
diet and
biomass contribution were calculated for each prey. Feeding strategies were studied based on our data for Portugal andpublished
resuttsfor
Spanish
regions,using
a
Principal ComponentsAnalysis
(PCA). Relations between prey frequencies were analysed using Spearman'sconelation-The resutts highlight the existence of three feeding strategies in the lberian Peninsula: diets based on lagomorphs; diets based on rodents; and diversified diets where lagomorphs are important preys. tn
Soúhem
Portugal, lagomorphs are the most important preys, and rabbits above alt. Some pairs feed mainly on lagomorphs while others have diversÍfied diets, basedon
rabbits and hares. lVledium sized birds, hedgehogs and rodents arethe
most common complementary preys.The Eagle Owl
is
dependentof
tagomorphsin
tberian meditenanean e@systems, reasonwhy is
essenciat maintainingthe
availabilityof
these
preys, also fundamentalfor
many predators.The
regulationefÍect of the
Eagleorrl
on its
preys hasbeen
poorly studied, although thisowlfeeds
mainly on game species, but also on meso+redators and species of conservation gonc€)m. The conservation of this predator seems to be dependent on a conect management of the habitats included in the tenitory of each pair, in order to achieve a good prey availability.2
Rui Fazetda Loueryo (2(XlO
Í.
lnüodução
A
onservação
de
uma espácie
ameaçadaassenta neessariamente nos pilares
do conhecirnento adquirido no âmbito de várias disciplinas das ciências nafurais e sociab-A
biologia
da conservaéo
ê
considerada umaárea
cientÍficade síntere
e
umadas
zuas cornponentes principais é a ecologia (MefÍe & Canoll1997)-No campo da ecologia, a dência das relaçôes entre os organismos vivos e o meio abiótico,
está
incluÍda muita
da
informação
de
base
essencial
à
biologia
da
conserva@o, nomeadamente em termos de conhecimento dos habÍrats indispenúveis à sobrevivência da espécie e estudo das interacções especÍficas e funçáo ecológica nos ecossistemas.A
elabora@ode
planos
de
conserva$o para as
espéciesque
apre§êntam situações desfavoráveisé
umadas
ferramentas básicasem
biologiada
conserva@o (Sutherland2O0O), tendo estes planos sido
já
preparados para algumas das aves mais ameaçadas da Europa (e.g. Goruález 1996, De Juana & Martínez 1997, Anoyo & Feneiro 2OO1).De
modoa
que seia possível delinearas
medidasde conserva$o
mais adequadas para uma espécie é primeiro necessário reunir informação sobre a sua biologia, nomeadamente: dimensão populacional, distribuição geográfica, tendência populacional, ecologia, factores limitantese
ameaças (Sutherland 2000,ver também
González 1996,Anoyo
&
Feneiro 2001).A
tarefade
síntesede
conhecimentosé
entáo umdos
primeiros passosem
biologia da conserva@o. No caso deste estudo, apresenta-se em seguida uma recolha da informa@o biológica existente, aplicadaà
conservação doBúo+eal
Bubo bubo (Ltnnaeu§, 1758) em ecossistemas mediterrânicos, e mais especiftcamente noSulde
Portugal.3
Rui Fazenda Lourenço (2@O
2,
lnformação biotógiea
ê
aspectoe
da
conservaçáo
do
Bufo-real
em
ecossistefl tas
meditedn
icos
2.í.
Descrição
O Bufo-reat
prtence
à Ordem Sfiigiformes (Voous 1977, Sibley & Monroeí996,
Howard & Moore 2OO3)e é
a
maiorave
de
rapina noc'tumado
Paleárctico Ocidentale
uma das maiores do mundo (Mihtola 1983, Burton 1W21.Os indivíduos de maiores dimensões pertencem à subespécie Bubo bubo sibiicus (Miktola
í983)
e
atingem comprimentosde
asa de 45,6qn
(machos)e
49,1 cm (fêmeas) (Crampí985),
claramente superiores aos valores referentes à subespécie que ocoÍre em Portugal, B.b-
hispanus, cujo comprimentode
asa rondaos
42,2 cm (machos) e 45,3on
(fêmeas) (Martínez et al.2OO2).O peso médio dos indivíduos da subespécie ibérica
é
1543 g para os mac{rose2027
g para asfêmeas
(Martínezet al.
2OO2), inferior aos valores médios paÍaa
subespécie centro-europeia B- b. bubo, sobretudo quando comparado com as populações da Rússia (machos:2458 g; fêmeas: 3164 g) (Cramp
í985).
A envergadura do Bufo-real, na Península lbérica, varia enhe os 147 cm (macho) e 160 cm (fêmea) (Martínez et at.2OO2), tendo uma dimensão aproximada a algumas das águias de maior dimensão que aí ocoÍrem: e.g. Águia-cobreira Circaetus gallicus (envergadura:
18$
195 cm; peso: 166l.1735 g), Águia-imperial-ibéica Aquila adalbefti (envergadura: 180-210 cm), Águia de Bonelli Hieraaetus fascraÍus (envergadura: 150-í80 cm; peso: 1721'2386 g) (Snow & Penins 1998).
O comprimento total na
subesÉcie
B-b
hispanus ronda os 63,0 cm para os machos e 67,4cm para as fêmeas
(Martínezet al.
2OO2).Ao
nível
da
plumagemesta
subespécie é bastante similarà
subespécie nominal B. b. bubo, diferindo apenas na cor de fundo maisdara e nas listras melhordefinidas na região ventral (Mikkola í983).
O dorso apresenta um padrão geral
de
manchas castanhase
pretas enquanto o ventre é amarelo-acastanhado pálidocom listas
verticais pretas bastante proeminentesno
peito (Mik'kola 1983, Crampí985).
O disco facial é castanho-acinzentado onde se destacam os olhos com íris hranja e os tufos auriculares escuros e compridos (Mikkola í983).2.2.
Biogeografia, distribuição
e
situação populacional
O
Género Bubo apresenta umadistribuiso
mundial vasta, estando apenas ausente da região Australásia(Cramp
í985).
Segundo Howard&
Moore (2003)
incluem-se neste Género 19 espécies, enquanto Sibley&
Monroe (1996) apenas consideram 15 espécies.Assim ambos estes autores
consideramas
seguintes
espéciesno
Género
Bubo:. B.Rtii F asda Lourenço (2(XlO
A ecdogh tffica do Búoreal Bub huao,es ÍmplicaçÕe consenacionbtc ern msísteÍna medÍteÍrânicc.
virginianus; B. bubo, B. ascalaphus, B. bengalensis, B. capensis, B. africanus, B. poensis, B.
vosselei,
B.
nipalensis,B.
sumatranus,B.
dtelley|
B.
ladeus,
B.
coratmndus,
B. leuoosficÍu.$B.
philippensis, diferindo em quatro espécies, induídas no Género Ketupa por Siblrgy & Monroe (1996) e no Género Bubo por Howard & Moore (2@3): Ketupa blakistani, K. zeylonensis, K. flavipes, K. ketupu.O
Bufo+ealBuba bubo
é
consi&rada uma
espéciedo tipo
paleárctico, emboraa
sua distribui@oúranja
todo
o
Paleárcticoe
parte das
regiôes lndo+nalaiae
Afro-tropical(Mikkola
í983,
Newton 2003). Trata-se
de
uma
espécie politípica
oom
diferenças morfológicas entre subespécies sobretudo ao nível das cores da plumagôm e da dimensão (Mikkola 1983, Cramp 1985).As subespécies reconhecidas no Paleárctico Ocidentalsão B. b. bubo, B.
b.
hispanus, B. b. ruthenus, B.b.
sibiricus, B.b.
interpositus, B.b.
nikolskii, B. b. turcomanus, B. b. gladkovi (Mikkola1983, Cramp
í985).
Relaüvamenteàs
subespécieeB. b.
deseftorum,B.
b. ascalaphus, B.b.
bengalensis, existem divergências entre autores que as consideram como especies distintas (e.9. Sibley & Monroe 1996, Howard & Moore 2003).O
Bufo-realocone em
grande partedo
Paleárctico Ocidental, constituindo excepçôes a lslândia,Grã€retanha,
lrlanda, França Ocidentale
várias ilhas mediterrânicas (Mik'kola 1983, Donázar&
Kalinainen 1997, Snow&
Penins
1998).No
Nortede África
ocoÍre a espécie Bubo ascalaphus (Sibley & Monroe 1996, Howard & Moore 2003) considerada por alguns autores como subespécie do Bufo-real Bubo buho (e.9. Snow & Penins í998). Durantea
primeira metadedo
SéculoXX,
o
Bufo-real sofreu um acentuado declínio em várias regiões da Europa, devido sobretudo à perseguição humana e profunda alteração dos habitats (Mikkola 1983, Donâzar & Kalinainen 1997).Segundo Mikkola
(í994), a
população reprodutorado
Bufo-real,em 34
países europeus ondeocore,
encontrava-se,no
iníciodos anos g0
entrieos
11.000e
os
40.000 casais, registando-seos
maiores valores paÍaa
Escandináviae
Rússia. Emborase
encontrasse estável ou em crescimento,a
popr.llaçáo europeia do Bufo-real, no seu global, estava em declínio, sendo por isso incluída na Categoria SPEC 3 e aúibuído o estatuto de Vulnerável (Mikikola 1994). O trabalho posterior de Donázar & Kalinainen (1997) para estes 34 países apontava, em meados da década de 90, para uma população reprodutora enlre 12.853 e 37.926 casaise
uma
tendência populacionalde
declÍnio, em @nsonância com Mikkola (1ee4).A
estimativa mais recenteda
popula@o reprodutora europeia,do
iníciodo
Século XXl,aponta para
12.OOOa
42.OO0 casais,dos quais
9.í00
a
20.O0Oem
paísesda
União Europeia, indicando uma tendência populacionalactualestável, embora a nívelpan-êuropeuRui Fzsda Lourenço (2@5)
A ecologh bófica do BtÍo-ral Bufu bub e
a
Ímptrcações consenacionistm €ín eccsist€Ína mediterÉnicos.a
situação seja maÍs desfavorável (BirdLifelntemationaEB&
2000,Birdlife
lntemational 2OA4r.A situa@o do Bgfo-real em Espanha é bem
onhecida,
estando disbibuído por praticamente todoo
país,à
excepçãodo
Litoral Cantábricoe
parte da Galiza (Marfínez &ZubrogoitÉ
2AAg). Comparativamenteà
estimativa meno§ preclsade
Punoy (19971, gue apontava a exiEtênciade
5Zoa
6&)
casais reprodr.rtoreem
Espanha,os dsdos
rnaísrealisEs
de Martínez &Zuberqoitia
(2003) referem a existência de uma populaÉo reprodutom mínimade
2345mais.
EstadiferenF
é devida à diftculdade dedete@o
daesscie,
queap6ar
da sua dimensão pode facilmente passar despercebida, sobretudo quando ocupa habitats considerad@ m6nos tÍpicos (Erclnázar&
Katinainen 1997, Martínez&
Zuberogoitia 20O3)' Ainda assim a tendência populacional parece crescente (Martínez & Zuberogoitia 2003)'A
informação disponível para Portugal, em termos da situação populacional do BuÍo+eal é redtrzidae
pouco precisa.A
estimativa de Rufino (1989) conespondea
um intervalo entre100 e 1.000 casais reprodutores, tendo desde a mesma deconido iá vários enos.
No
âmbito destatese,
foi
realzadauma oompilaÉo
de
informa@es diversassobre
aocorrência
da
espécie, muÍtas das quais não publicadas, resultando uma estimatíva dos efectivos noSulde
Portugal na ordem dos 150 aZAO casais reprodutores e para todo o Paísuma
populaçãode
200
a
400
casais.A
informação utilizadapara
a
realizaçâo destas estimativasfoi
a
seguinte: Rufino (1989), Santos (1998), EliaseÍ
at
(1998), ÁMares & Figueira (1999), Lourenço (20ü)), Lourenço et al. (2oo2), Lourenço (2OO2), Pinheiro (2003),costa
et
al.
(2003),cEAt
(ZAf/,'), Lourenço(2W4), bem como
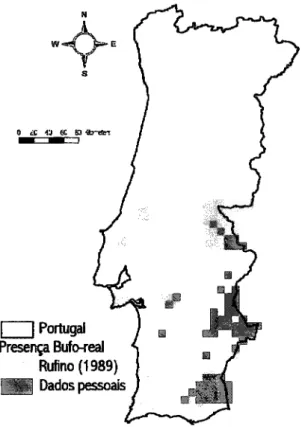
diversas observações ocasionais não incluÍdas nestestrabalhos-O Bufo-realocone de Norte a Sulde Portugal, embora a sua distribuição esteja concentrada ao longo da metade interior e bastrante associada aos vales encaixados dos rios principais
(Fig.
l;
Rtrfino1ggg,
Lourençoet
al.
2OO2,R.F.
Lourençodados não
publicados)- A tendância populacional não é conhecida uma vez que a espécie não é alvo demonitoriza$o
dos efectivosreprodutores-Esta
espécieestá incluída
na
categona
Rarodos
estatutosde
conserva@odo
lCN, apesentando uma tendência populacional regressÍva (Cabratet
al.
1ÇNO).Na
revisão doLiwo Vennelho dos Vertebrados
de
Portugal apresentaa
etegoria
Vulnerável, aÚibuÍdamm
base
no
critério D1, i.e.,
tamanho esümadoda
populaçãomenof
do
que
'l'000
indivíduos mduros (lCN ín Pre§s).6
Rui Faerda Loueryo(200O
!t
A
<F>Y
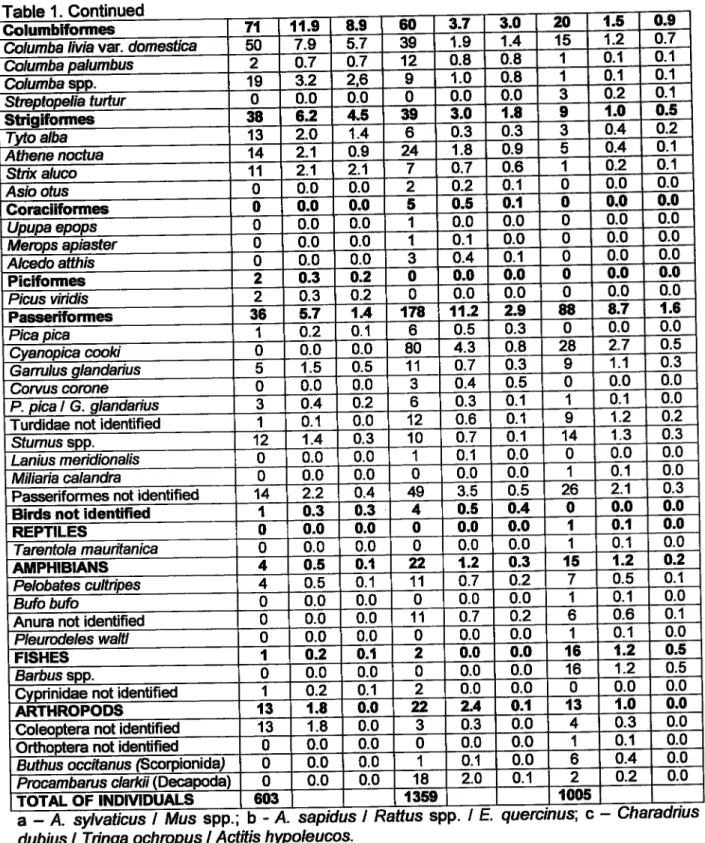
w 0 úC {:, 66 8n {rÉ r---tT-_|
Portugal PreerçfuÍo'real Rufino (1989)ffi
DadospesoabFigura
1.
Distribuiçãodo
Bufo-real Bubo bubono
Sulde
Portugal, com baseem
Rufino (1989) e em dados pessoais (1997-2OOS).2.4. Comportamento
O Bufo-realé uma espécie sedentária e bastante tenitorial ao longo de todo o ano (Mikkola
í983)- No
entanto, algumas populagõesnos
limÍtes Norteda
distributçáo(e.g.Rússia)
apresentam um comportarnento nómada tace
a
lnvemos muito rigorosos (Cramp Í985),bem como
populaçôesde zonas
montanhosas (Mikkola 1983).Os
indivíduos imaturos reatbam movimentosde
dispersâo,que
podeÍn atingiros
200 km, no entanto,a
maiorianunca
ulúapassaos 50
km
(Mikkola 1983,Cramp
í985),
Esta
espécietem
actividade crepuscular e noctuma, sendo por isso de dÍÍícil observação (Mik{<ola í983). Apresenta uma considerável actividade vo@|, uma vez queos
chamamentos constituemo
principal meio utilizado para a defesa do tenitório e atracção de parceiros (Mikkola 1983).2.5. Ulilização
dos biótopos
A dimensão dos tenÍtórios de Bufo-real na Península lbérica é mal conhecida, no entanto, a
sua
dimensâoé
relativamentepquena,
isto
é,
quasesempÍe
inferiora
10
km2 (Ruiz-Martínez et al. 1996, Lourenço 2000, V. Penteriani oom. pess.).Rui Fzerda Larenço (zfi)O
A ecologia htrca do Búqrel Bub bufu e
*
implicryões csservacionísta ern ecsbtana mediterrânicc.Em termos de
sele@o
do habitat os estudos existentes apontam para uma preferência por zonas com algumainegularidde
topográfica localEadas em regiões de altÍhrdes baixa ou média e com matagais e culturas de sequeiro (Donázarí988,
!úadínez et aL.1992, Martínez & Calvo 2(X)O, Penterianiet
al.2OA1, fr/liartínez et al.20FIJ). A heterogeneidade da paisageme a
disponibílidadede
coelho parecem §ualmente caracíerÍsticas favoráveio. Á'reas com maiordeÍlsidade populacbnal e várias estradasalcatrode
eão evitadas.No Sul
de
Portugal,as
áreas maisfavoÉveis
parao
Bufo-real sãoos
valeoripímlas
ou crisúasroc*toss
rodeadas por matagais, zonas agrícolas de sequeiro ou montadoe pouco densos, onde*
presas são abundantes, muitias vezes associado a zonas de caça especial (Santosí998,
Lourenço 2AAO, Lourençoet
al.2OA2. Pinheiro 2AO3, CEAI 2OA4,Lottenço
z0o4-).
2,6.
Biologia
da
rcprodução
Nos ambientes medÍterrânicos
o
Bufo-real iniciaa
reprodução bastante @do, deconendo em todas as suas fases entre Novembro e Maio.Como
local de
nidificação utiliza sobretudo plataformas em encostasroctosas,
embora pontualmente ocupe ninhos em árvores construídos por outras espécies (e-g. cegonhas) ounidifique
no
chão
(Mikkola
í983).
No
Sul
de
Portugal
os
ninhos
encontram-se maioritariamenteem
vales
escarpados,
cristas rochosas
e
pedreiras
abandonadas (Lourenço 2000, Pinheiro 2OO3, CEA|2004).Esta espécie epenas real;za uma postura, embora sejam frequentes posturas de reposiçáo em caso de perda de ovos ou crias (Mikkola 1933).
A
postura é geratmente de dois a três ovos (por vezes quatro), que são incubados sobretudo pela fêmea durante32 a
35 dias (Mik{<ola 1983).Após
a
eclosão,as
crias
dependemda
alimenta@odos
progenitores durante @rcz de úês meses (Crampí985,
Penteriani et al.2O04b). O sucesso reprodutor é normalmente elevado,se
na região em causa não esüverem sujeitoaa
muitos factores de perturbação.Blondel&
Badan (1979) referem paraa
Provença(Franp)
uma médiade2,l
iuvenis por casal em 13 ninhos com posfunas. Olsson(í997)
aponta um valor médio de O,Tjwenis
por casal paÍaa
Suécia. Penterianiet al.
(2OA4a) apresentam uma produtividade médiade
1,7jwenis
por casal na Provença. Em Murcia (Espanha)a
prodtrtividade média (númerode juvenis por
casal)varia
entrie0,9
e
2,4
(MartÍnezet
al-
1gg2).No Sul
de Portugal,Pinheiro (2003) refere para dois anos
@nsecutivosos
valores
de
sucesso reprodutor deí,1
e 2,2 cnas por casal com postura.Rui Fazenda Lorenço (2G)5)
A ecdogb trffica do Buforet Buôo bub e
a
impticaçÕe consenracionistm ern ecsislerno mediffiiicc.2.7. Feetaree do amêaça
ê Gausasde
mortalidade
Os principah factoree de arneaça do Bufo+eal,
a
nÍveleuropeu, estÍiomociados
à aaçao direc*a do Homam, sendo os mais relevantes:a)
PertrrbaÉo
humana devido à pÉtica de ac'ttvidades de lazernasimedia$e
dos ninhoo (Mikkola Í994);b)Abate
ilesal de avee (Mikkola 1g%, Martinez & Zuberogoitia 2003);c) Colieão e elecf,rocussão em linhas de transporte de energia (Rubolini et al.20íJ1, Sergio
eÍ
aL 2Ao4.);
d) Mortalidade por atropelamento. sobretudo de iuvenis (Mikkola 199a\;
e) Pilhaoem de ovos e crias no ninho (Mikkola 1994, Martínez & Zuberogoitia 2@3);
f) Dedínio das populações de coelho-bravo devido a epizootias (Donázar& Kalinainen 1997, Martínez & Zuberogoitia
200í,
MartÍnez & Calvo 2OO1, Martínez & Zuberogoitia2fi)3).
2.8.
Cnlários
de consorvação e
insúumentos
legais
O
Bufo+eal está incluÍdo na categoria SPEC 3, dado apresentar um situação desfavorável na Europa, embora a sua população mundial não esteja concentrada exdusivamente nesta região(Birdlife
lntemation al 2OO4l.Esta espécie está incluída no Anexo I da Directiva Aves (79l409lCEE;
DL
140199 de 24 deAbri[
e no Anexo ll da Convençáo de Bema (DL 95/81 de 23 de Julho; DL 316/89 de 22 de Setembro).2.9. Ecologia
frófica
2.9.1- lntroduçâo
teórica
sobre a ecologiabófica
As
interacções entre espécies são um dos temas cenhais da ecologiae
um pilar para a compreensão dos ecossisbmas, no entanto, estas apresentam uma elevada complexidade,o que na§uns
casos dificr.rltaa
sua compartimentração em categorias estanques (Brerrerí 994). Uma divisão inicial possível destas interacções poderá ser agrupá-las em
intera@s
trróficas (nasquais uma
espécie utilizaoutra
omo
alimento)e
interacç6es não troficas(Brewer
19W').Na
primeira categoriaestão
incluídas,entre outras,
a
preda$o
e
o parasitismo, enquanto quea
segunda categoria abrangeo
@mensalismo,a
compeüção, neutralismo, etc.A
complexidadedas
interacções tróficase
em particularda
predação, ponto central neste estudo, é revelada ao nívelde algumas questõesúave
paÍaa compreensão (Brewer 1994): 1) a actividade é a base da transferência de energia numa comunidade;Rui Fazeda Lourenço (2@O
A ecologia trtrca do Bufored &rb bufu e
x
implicaçõe consenmimista ern ecossistema mediltsrânicc.2)
existem diferentes técnicas de alimentação, nomeadamentede
procurae
capfura das preffis;3) pode existir uma influência da
intera@o
na dimensão dapopulafro
do predador ou da pÍesa (efeitonuméfio);
4)
existe um efeito seleciivo, ou evolutivo,da
predação sobrea
presae
da escassez de alimento sobre o predador.Na
relaçãopredadorpresa,
existem algunsfactores
mndicionantesque
determinam aincoçora$o de
uma
determlnadaespécie
na
dieta
de
um predador (Brewer
19911),desÍgnadamente:
Í)
a abundância da especie presa;2)
a
disponibilidadeda
presa em termos
de
eptur:a
(comportamento,meenismos
de defesa, etc.);3) e a preferência do predador (dependente sobretudo da relação custo captura - benefÍcio energético).
A
dieta óptima (optimal
foraging)é
consideradoum
ponto fulcral
na evoluSo
dos predadores, baseando-se numa fun@o entreo
benefício energéticodo
consumode
uma determinada presae o
custo (em termos sobretudode
tempoe
energia) dasua
captura (Brewer 1994).No entanto, na§uns
casos nãoé
apenasa
relação custo-benefÍcio que determina a dieta, mas também uma série de outros factores, como sejam a alimentação de crias, a susceptibilidade à predação, etc (Brewer í994).O efeito da predação nas populações do predador e da presa é uma questão complexa na ecologia das comunidades (Ricklefs 1990, Brewer 19€.41.
O
modelo predador-presade
Lotka-Volterra baseia-se nos princípios simplesde
que: um predadordepende da popula@o da presa, sendo o efeito expÍesso na taxa de natalidade da e§péciê predadora; e os efecfivos da presa dependem da população do predador, gue adtua sobrea
taxade
mortalidade. Em síntese,o
predadoré
limitadopela
presae
a
presa é limitada pelo predador (Breurer 1994.).De facto, este modelo revela-se demasiado simples para
explier
a complexidade inerenteàs
relaçõesenúe
predadorese
presas,não
avaliandoa
capacidadedas
presasde
se refugiaremdo
predadoç bem oomoa
capacidadedo
predador de se alimentarde
presas altemativas (Brewer 1994, Ricklefsí990).
Algumas críticas ao modelode
Lotka-Voltena apontaram sobretudopara o facto de que
a
respostados
predadoresà
densidade das presas nãoser na
generalidade uma função linear, mas poder tomar formas variadas e complexas (Ricklefs 1 9gO).Rd Fzenda Lourenço (2@5)
A ecologkt frffica do Buforeal Brb bub e
a
impfcações cons€rvacion[rtc ern eccsisterna medlterràricos.A predaSo
irúaguilde, ou Beia,a
crtfiwa
de urn animal que explora recuÍaos alimentares seneltrantes, é urn fenórneno particuhr da predaçãoe
reiÍeÇe ser rnais restrita a indivÍduasjovens
e
espéciesde
menor dimensão como prêsas;fado
devidosobtettdo ao
risco e esforço inerente ao proomso de morte de um oubo predador equiparado (Polis et al.1989)-No caso deste esúudo, são particulannente interessantes todos os fenómenos
inêfenb
à predação e ao papel dos predadores nas comunidades bióticae, uÍna vêz qüê a eepécie em çgusaé
um predadordo
topoda
cadeia úóficae
uÍna espécie relativamente @mum na regiãomeditenânie
(Donázar & Kalinainen 1997, MartÍnez & Zuberogoiüa 2003).2.9.2. Ecologia
trófica do Bufo+eal
Os
hábitos alimentaresdo
Bufo-realsão um tema
bastante estudadoa
nível europeu, existindo trabatrhos publicadospaÍe
a
maioriados
paÍses onde ocome (ver revisôes em Mikkola 1983, Penteriani 199ô). Em Portugal, o conhecimento sobre a biologia do Bufo-realé
ainda
reduzidoe
sobretudopouco
divulgado, exiEtindono entanto
alguns trabalhos pontuais (Iavares et a\.1989, Miraí995)
e poucos estudos mais aprofundados embora não publicados (Santosí998,
Lourenço 2000, Pinheiro 2003).Nesta perspectiva,
o
presente estudo pretende suprimir esta lacunado
conhecimento da ecologia tróficado
Bufo-reat, tentando aprofundara
verlenteno
âmbÍtoda
biologia da conservação.3.
Obiectivos
Os recursos alimentares são um dos principais factores limitrantes na reprodução das aves de rapina, a par com a disponibilidade de locais de nidificação (Newton 1979,lrlewton í998), sendo por isso um tópico essencial na @nservação de
predadores-Assim,
os
objectivos deste estudo consistem na análiseda
ecologia tróficado
Bufo-real,atavés
das seguintes abordagens encaradas numa perspectiva de biologia da conservação:1)
Estudo das estratfuias
atimentaresdo
Bufo-realem
ambientes mediüenânicos da Penlnsula Íbérica.2)
Estudo
da
dieta
do
Bufo-reat
no
Sul
de
Portugal
e
análise
das implie@s
conservacionistas.
Rui Fzenda Lorenço (2(ffi)
AedogiatrfficadoBrÍo,real BuúpbuE*ee implicaçôes consenacionista enr eccsisterna mediterâticc.
4-
i*eade
etrdo
Este estudo foi desenvolvido no
Sulde
Portugal, definido neste proposito ooÍno aporÉo
do tenitório continentala
suldo
rio Teio.A
árca de estudo globalempreende
guafo
áreas localizadas no Alenteloe
Algarvee
enquadradas naa baciashidrográfies dos
rios Tejo, Guadianae
§ado
(F.g-2).Estas guatro
áreas apresentama§uma
variaçãoem
temrosbiofísios e paisagÍstios,
embota em todas predominem características meditenânicas.A
região no todo
pertene
à Província Luso-Extremadurense (Costa ef aí- í998).w E 0 20 6 €0 eoFniffi I=-'l
Troço
médio do Guadiana NEAlgarvio
Fgtna
2. Locr,lbaçâo das quatro regiões estudadas no Sul de Portugal.Nordeste Alenteiano (PNSSM)
Esta área conesponde em grande parte ao Parque Naturalda
Sera
de São Mamede. Esta zonawÍrana
vana em aÍtitude entreos
300 eos
1027 m s.n.m.e os
habitats dominantes são as florestas de carvalho-negral Quercus pyrenaica, sobreiro Quercussubere
azinheira Quercus rotundifolia,os
matagais dominados por estevase
giestas,os
soutos (Castanea saÍnra), pinhais (Pinus pinastefl, olivais (Olea europaeal e cultunas agrícolas de sequeiro.S
RuÉ F aada Lourenço (200O
 ecohgb hffica & üúeted Bub hrfu e
x
Ímplicryões cmsenracionbtc ern eccsbtsÍras ÍÍteditenânÍcos.Bacia
do
PNSSM
Troço mádio da Bacia do Guadiana
Esta área looaliza-se noo concelhos deAlandroal, Reguengos de B/lorlsaraz" Mourão, Poltel
e
Moura, ondeos
principais afluentesdo Rio
Guadiana são:Luefêce,
Azevé|, Cuncos, fúcanache, Degebe,Adila
e@eare.
As planícies são dominantes, sendo cortradas polos vales acentuados dos rios e ribeiras- Os habitds dominantes são os montados e bosgues deuirtho,
os matagais escÍeróúÍtos, os olivais, as únhas e as culü.rras agrÍcolas de segueÍro.SÍtio de Monfurado (Bacia do Sado)
Esúa área destam-se da planície alentejana envolvente por um aumento da altitude até aos 40O
m.
A
paisagemé
dominadapor
montadosde
sobroê
ern mênorgrau
por cufturas agrÍcolas de sequeiro, vinhedos e rnatagais.Nordeste Algarvio
Esta área
c,orespondoàs bacias hidrogÉficas
do
Rio
Vascãoe
Ribeirada
Foupana, afluentes do Rio Guadiana. O relevo é ondulado, mas de caraderísticas não muito bruscas, existindo poucos desníveis muito acentuados.Os
habitats dominantessão os
matagais esclerófrtos, os montados de azinho e as culturas hortofrutÍcolas de sequeiro. Os bosques de sobro e as plantações de pinheiro-manso Pinus pinea estão presentes, embora assumam uma menor importância.Rui FazerdaLqreryo(2@O
A
@ia
trófica do Bufored Brfu bubea implicaçb conseruacionistm ern ecssisterna mditerránicc.5. Os
hábitos
alimerúare
do
Bufo-real na Península lbérica
Artigo subrnetido a: Joumal of FtoH Omithology (Asrcciation of Field Omithologists)
Rui Fazerda Loureryo (2(X)O
À ecotogia hffica Uo ÉWLrea Arrôo butu e
* impk#
cqrsenrcíqr[stc sr ecesbterns mediterrtuice.THE FOOD T.ffiEITS OF EAGLE
OIflI-g
IN THE IBERTÂSI PEHINST'I.ARui F.
Lourenço
Cenfio de Estudos da Avifauna
lbérie,
CEAI, Apartado 535,7AA2'506 ÉvoraABSTRACT: lnformation on the diet of Eagle
&vls
inhabiting the lberian Peninsula is mostly dertved fromstrdies
in Spain, and until nouí nopubtishd
data was availablefor
Portugal.Diet was
analyzedin
23
EagleGail tenitories
in
Southem Portugaland
results where compared with data ftom Spanish studies using PCA. Diet in Southem Portugal raras mainly based on Lagomorphs. Overall in the lberian Peninsula three main diets were described: (1) tagomorphs-based;(2)
rodents-based; (3) diversifted diets wÍth lagomorphsas
mainprey-There were no evident large
sele
diet
variationsdue
to
high
heterogeneityin
lberianMeditenanean landscapes. Differences
in
diet are
mainly determined
by
tenitorycharac*eristics. The results obtained point out the need to (1) recover rabbÍt populations; (2) appty habitat management measures to increase preys populations at the tenitory scale; and (3)
analpe
the influence of diet variations on the fiútess of breeding pairs.KEY-\ /ORDS: Bubo bubo, diet heterogeneity, Mediterranean habitats, Portugal, Spain
Rui Fazelda Lourenço (2(X)O
AmtogiatóficaOoÉufer€at Bufubuáoea implicaçÕes consenacionhb ern eccstslernc mediterrânicc.
Knowledge on the food habits of a predator is na essenüal feature to its conseryaüon' since diet inftuances survival (Hakkarainen et at. 2AA2l, breeding performanoe (Vrftuela arud Veiga 1992;
Anoyo 1998;
Penterianiet
al.
2ao2l and
revêals possibleconflicb wtth
human activities Clhirgood et al. 2000). Moreover, thê analysis of the diet of topprdators
may be partictrlarlyurefulfor
species conrervation vuhen intraguild predation is acting (sergio et al'zAAq.
The Eagle oad Bubo bubo is a large prcdator of Palearctic disffibution and diet of European populations
was
laryely studied
in
several
counÚies (Penteriani1996)'
Hourcver, theinformaüon on the diet of this species in the lberian Peninsula concêms only spanish regions (e.g. HÍraldo et
at.
1975;Donázar 1989; Serrano 1998; Martínezand Zuberogoitia 20O1), and no information is availab&a forPortugal-Lagomorphs,
and
more
speciftcallyrabbits
Orvctolaouscuniculus,
play
an
important ecological role in the food webs of westem Meditenanean ecosystems (Delibes and Hiraldo1981; lbona
et al.
1990). Eagle
owls
are
rabbit-specializedpredators
in the
lberianpeninsula (Hiraldo et al. 1g76; Donázar et al. 1989) that, due to the abundance of this prey,
attain
here high
population densities.
ln
the
last
decades,
myxomatosisand
rabbithemonhagic disease were responsibte for the impressive decline of rabbit populations in the tberian peninsula (Villafuerte et al.
í995).
As a consequence, two rabbitdependent species likethe
Spanish lmperial Eagle Aquila adalberti andthe
lberianLynx
Lvnx pardinus are nomadays in dramatic conservation situations (Fener and Negroz}Aq.Athough
large scaleeffeds of
rabbit diseases on lberian Eagle Ouvl populations and their diet have never beenevaluated, soÍne
aúhors
have reported negative
consequences associatedwith
the reduction in availability of this prey (Martínez and Zuberogoitia1ool; Penteriani et al- 2002)' Main obiectivesof the
present studywere:
(1)the
descripüonof the
Eagle&vl's
diet
in Southem portugal; and (2) a comparison of food hab'lts among different lberian populations to assess the possibleexistene
of diverse diet skategies in the lberian Peninsula.METHODS
§firdy
aÍee. The study areais
located in Alenteio and Algarve provinces (Southem portugal), included inthe
Mesomediteranean and Thermomeditenanean bioclimaüc belts'character2ed
by
a
hot
and
dry
Meditenaneanclimate
(Rivas-l\íartínezet
al-
2oo2}
Landsepe
is dominated by plains or low hills with etevation ranging from sea-leveltoí000
m a.s.!., and main habitats are agricultural fields (cereal steppes, olive groves, vineyards), Meditenanean scrublands,and
Holmoak
(Quercus rotundifolia)and
Cork Oak (Quercus sube4 pastoralwoodlands (lnontados). The area is cunently suffering large scale changes inRui Fzenda Loureaço (2ülO À
""aogà trmo
Oo ÉufLreaf alm bub e
8
implic@ consenracisrbta eÍn €caisteínã medÍÍenâticc'the
landsepe,
mainly due to large reseruoirs, convercion of traditÍonal agricultural fields into tnigated onês, and abandonment or afÍolestation of openfields-Tenitory
suryley. Eagúe Orul tenitories raere searched from í997 to 20&4 by combining listeningand
playbad< sesÊions duringüre
pre{ay}ng peÍiod(Novemberfebruary in
the study area), followÍng Penterianiet
al.
QOOI) and l{lartít'elz etaL
QAA3)- [/loreover, from hlarchto
June, atl suitable aÍaas were visited to searchfor
nests, roost sitea, and feeding percfies.Diet analyeis. Eagle charl's diet was studied on a sampb of 23 tenitories.
í5
tenÍtories were visited twice (in dÍfferant yearc), while the remaining eight were visited only once- Toredue
disturbance, visits took place at the end ofthe
breeding season only. During eacft visit,al
feeding materials (i.e.reent
and old pellets, prey remains, and accumulated bones)rruere collected. To reduce bias due to double-counting of a same prey, the minimum number
of
individualEwas
determined fotlowing recommendationsof
Simmonset al- (í991)
and Marchesiel
al. (20O2a). prey Ítems were identÍfied by comparison with both the collection ofthe
Laboratoryof
Archaeozoology(lPA
-
Portuguese lnstitute
of
Archaeology),
and identificationtables
of
specific studies (Gama
1957; Madureiraand
Ramalhinho 1981; Moreno 1985, 1986, 1987; Callou 1997; Prendaetal.
1997). Data are presentedas
prey Írequenciesonly
(i.e.the
numberof
itemsin
a
prey
class dividedby the
sample size),because
they best
reflect foraging efforts
(Rosenbergand Cooper 1990)-
Moreover, frequencies were also used because in the lberian diet comparisons severalstudies did not reported biomass percentages.Spanftsh
sfrrd'les. For
comparisonswith feeding
habitsof
Portugueseowls,
the inclusion of Spanish diet studies followed three main criteria. First, the diet descripüon had to be separated by localities, each conesponding to a breeding pair. Secondly, sample size per tocality hadto
be targer than50
prey individuals- Finally, only those studies in which prey groups were well dÍscriminated (i.e. species or genus) were included. So, the comparative analysis induded five studies witha
total of 68 localities (Hiraldoet al.
1975; Pérez 1980; Donázar 1989; Senano 1998; Martínez and Zuberogoitia 2001), which allowed a good cover of the Spanish Medítenanean region.Staüstical anaryses.
Principal ComponentsAnalysis
(PCA)was used
to
arrange localiüesin
subsets, basedon
prêy classes frequenciesin the
diet-ln this
analysis the components (axes)revealthe
conelations among variables (prey classes) and each locality is then retated with these components (tabachnickand
Fidell 2001). PCAwas
performed using 28 variables, consisting in the frequency of different prey classes in the Eagte Owl diet-Data inthe
cross-producús matrix was centeredby
columns (variane/covariance), so that the variables could contribute to the analysis in proportion to theirvariances (Jongman etal-Rui Fzenda Lqrenço(2@O
À àr"Si" hófi* Oo gufLÍêd'Auôo bub e
a
implicações consermcionistas sn eco§sisteÍn6 mediteÍrâniúc.1995). Scores
for
column variabtes were calculated by weighted averaging (McCune and Mefford 1999).Spearman's rank conelation (rs; Zar 1999) was used to test for relationships between prey class frequencies and diet diversi§ (Shannon's index; Brower et a!. 1990). All tests were two-tailed and s§nificance level was defined at
5
0.05. All statistical analysis were computed usingthe
softwarePCORD
4
(McCuneand
Mefford 1999) and SPSS 10.0 (SPSS lnc. 1eeg).RESULTS
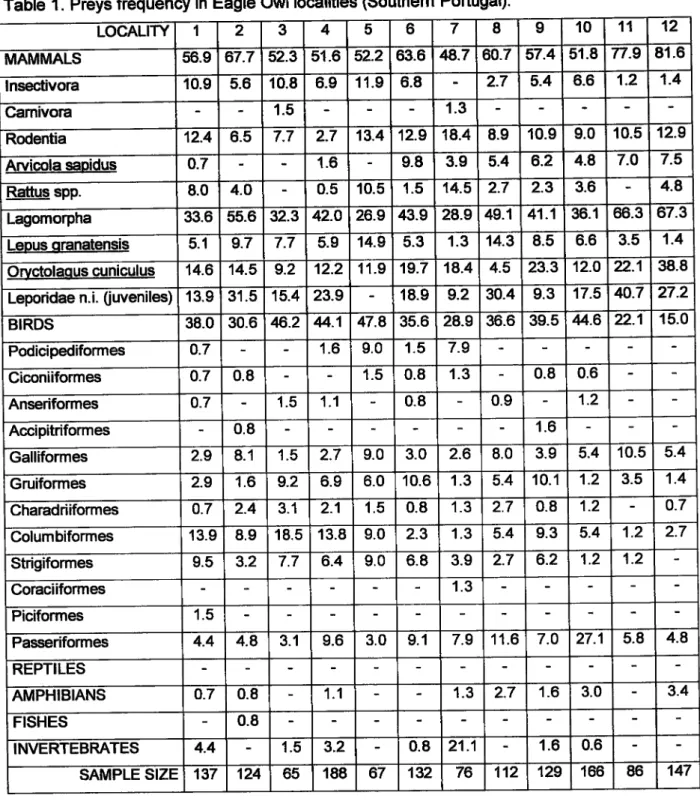
Diet in
Southern
Portugal. The 23 tenitories yielded a total of 2666 prey individuals, belongingto 64
different species(Iable
1). Lagomorphs (rabbitand
lberian hare Lepus sranatensis) werethe
most frequent preys inthe majori§ of
localities, ranging from 27 to670/o (mean frequency = 47o/o). Among the other mammal species, the most frequent preys
were
rodents (mean frequency
=
13%)
and
hedgehogs
Erinaceuseuropaeus
(mean frequency = 5%). Birds were present with frequencies ranging between 14 and 48olo (meanfrequency = 32o/ol, from which passerines (mean frequency = 9olo), pigeons and doves (mean
frequency = 4o/o\, owls (mean frequency
=
3o/o\, moorhen Gallinula chloropus (91%of
the overall Gruiformes, mean frequency = 3%), and the red-legged partridge Alectoris rufa (99% of the overall Galliformes, mean frequency = 9%) were the most frequent preys.Diet in the lberian Peninsula.
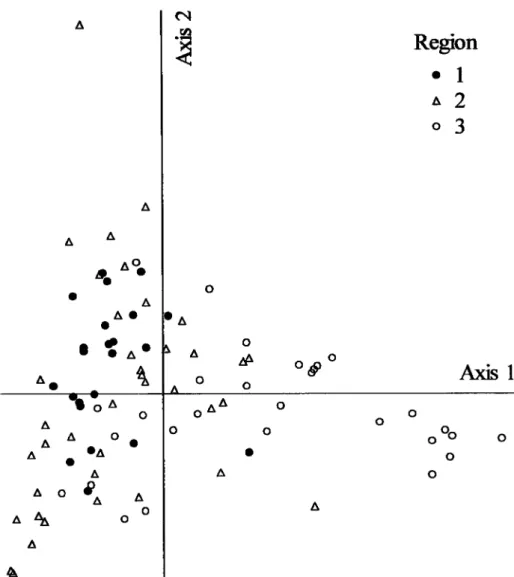
lnthe
comparative analysis,the 91
Portuguese and Spanish localities corresponded to a totalof
14251preys. First and second axes in the PCA explained cumulatively 80.5% of the variance (axis 1:
51.1o/oi axis 2: 29.4o/o). Axis 1 showeda
high
conelationwith the
frequencyof
rodents (eigenvector=
0.739),rats
Rattus spp. (eigenvector=
0.216) and mammals (eigenvector=
0.157) onthe
positive side, while the negative side was mainly conelated with the frequency of lagomorphs (eigenvector = -0.587)and
birds (eigenvector= 4.166).
Axis2 was
highly conelatedwith
birds (eigenvector = 0.559) on the positive side, and on the negative side with mammals (eigenvector ={.556)
and Iagomoçhs (eigenvector = -0.512). Therefore, the four quadrants of the PCA plot (Fig. 1) corresponded to:(í)
loelities
with diet based on lagomorphs and birds; (2) localÍties withdiet
based mainly
on
lagomorphs;
(3)
localities
with
diet
based
on
rodents
andcomplemented with birds; (4) Iocalities with diet based
on
rodents and complemented with lagomorphs. Localities from Southem Portugal belonged almost entirely to the two quadrants on the left, that is,diets
based only on lagomorphs and based on lagomorphs and birds. A locality in Salamanca (Spain) was clearly isolated on the top left quadrant, conelated with the frequency of birds. On the lower right, there was also an evident group mainly constituted by localities from north-eastem Spain, which was conelated with high frequency of rodents.Rui Fazenda Lourenço (2fi)S)
A ecologia trôÍica do Bufereal Bubo bubo e
a
implicaçõe consenracionistas em eccsistemas meditenânicos.Diet diversúy
was
negatively conelatedwith the
frequencyof
mammals (r"=
{.81;
P
= 0.000) and lagomorphs (r, ={.36;
P = 0.000) and positively conelated with the frequency ofinsectivores(r.=0.70;
P=0.000),hares(r"=0.24; P=0.020)andbirds(r"=0.64;
P=
0.000). Â
Region
e\
a
x
.l
À2
o3
À^a
D. a  a^
o ood)
Axils
I
ooÂ
o oooo
o oÂo
Âà
A A AFigure
1. PCA
scatterplot
of
91
localitieswith
EagleOwl
breeding paircin the
lberian Peninsula, compared using the frequency of 28 prey classes. Regions: 1) southem Portugal; 2) south-central Spain; 3) north-eastem Spain.DISCUSSlON
The Eagle Owl
is
considereda
generalist top-predatorwhich
specializesin
the
most advantageous food and therefore being able to occupy several biomes (Donázaret al. 1989). ln Southem Portugal, as in other Meditenanean habitats, lagomorphs are the key preys ofthis
owl, once rabbits area
very profitable prey dueto
their abundance and size.ln
the Portuguese region however, lberian hares also havean
important contribution, showing ao o Áê o A a
â
o Âoo a  A ao
a Ào o o a A o a  ooa
o O4 ÂI
  a A À^
o oRui Fazenda Lourenço (2005)
A eologia trófica do Bufqreal Bubo bubo e as implicaçÕe consenacionistm em eccsisternas mediterrânico.
higher mean frequency (9%) than most Spanish studies (e.9. Hiraldo et
al.
1975; Donázar 1g8g;Serano
1998; MartÍnez and Zuberogoitia 2001). On the other hand, some breeding pairs have more diversified diets, which include small mammals and birds,as a
probable response to lower availabilÍty of lagomorphs, typical of a generalist predator.Overall, in the lberian Peninsula, three main diets can be considered, depending mostly of the avaitability
of
rabbits: (1) low diversified diets based almost exclusively on lagomorphs; (2) diets base on rodents but also including lagomophs and birds; and (3) diversified diets in urhich lagomorphs are the main preys but birds, and sometimes small mammals, have hÍgh frequencies also.lberian localities with lagomorphs-based diets were found mainly in areas
with
high to very high abundanceof
rabbÍts (Vitlafuerte et al. 1998; Mathias í999). Accordingto
Marchesi etal. (2O02b), focusing on an optimal prey is the best Íoraging strategy and is related to higher breeding success
in
Eagle Owls. Higher proportionof
rabbitsin the diet
of
Black Kites 1g1ygs_tdorans) is also related to higher reproductive success (Vifiuela and Veiga 1992) and in this case, lberian Eagle Owls pairs specialized on capturing lagomorphs may also have a better breeding perfonnance.On the other hand, localities with rodents-based diets were found mainly in the north-eastem
lberian
Peninsula,in
areaswith low
rabbit abundance (Villafuerteet
al.
1998; but
see Donázar 1989;Senano
1998).ln
a
situationof
rabbit scarcity, Eagle Owlswere
able to capture rats as main altemative preys. Nevertheless,the
existence of effects on breeding performance hasnot
been assessed, and these localities may colTespondto
lower quality tenitories.ln their tum, one would expect diversified diets
to
be associated with mediumto
low rabbit abundance, as highlighted by Penteriani etal.
(2002) for Meditenanean France. Localities with diversified diets were widespread in the lberian Peninsula, and according to Villafuerteet al.
(1998) somewere
locatedin
areaswith
high rabbit abundance.The
heterogeneity inherentto
MedÍtenanean landscapesis
responsiblefor
intrapopulation variations (see Penterianiet
al. 2OO2, 2AO4l, and is probably behind the fact that in areaswith
high rabbit abundance, both diversifiedand
lagomorphs-based diets were found.Above all this
is a maüerof
scale, oncein
areasof
high rabbit abundance, smaller patches may have lower abundance and though a lower frequency of lagomorphs which leads to diversified diets. Overall, variations in the diet of Eagle Owls in the lberian Peninsula seem mainly dependent onthe
characteristicsof
tenitories. Differences between study areas have comparatively lower importance due to landscape heterogenei§. When available, lagomorphs are the main preys, however Eagle Owls are ableto
diversifu their diet or even shiftto
rodents-based diets, when rabbits are scarce in theirtenitory. This dietary plasticity, associated with relative eclecticism inthe
choiceof
nesting site could represent favorable life-history traits helpingRui Fazenda Lourenço (2ü)5)
A eologia úúÍica do Buferel Bufu hubo e
*
implicações cúsen acioni§tã qn ecossist€rÍtm meditenânice.Eagle Owl's to survive in the lberian Peninsula betterthan other large avian predators as the Bonelli's Eagle Hieraaetus fasciatus or the Spanish lmperial Eagle. Nevertheless, a piece in
the
puzle
is süll missing,that
is, the consequencesof
eachfreding
strategy in terms of fitness.Conservaüon
implicaüons,
Rabbit populations have been suffering large decreases, associatedwith
diseases (Viftuelaand
Villafuerte2003), as well as
inadequate hunting efforts and habitat management (Moreno and Villafuerte 1995: Calvete et al. 2005), that canhave several negative
consequenceson
Eagle
Owl
populations(e.9.
Martínez
and Zuberogoitia 2001; Martínezand
Calvo 2001).Since
rabbits arethe
stapleprey of
this predator, the recovery of the populations of this lagomorph is essential to lberian owls. Conservation measures based on habitat management as a way to increase prey availability (e.g. Moreno & Villafuerte 1995; Ontiveros et al. 2005) should be, in the case ofthe
Eagle Owl, based firet on the tenitory scale.Finally,
it
is
necessaryto
assesshow tenitory
characteristics influencediet
in
lberian Meditenanean ecosystemsand which are
the
consequencesin
terms
of
survival
and breeding performance. These results can help in identifying high quality areas for Eagle Owls and primary for conservation.ACKNOWLEDGMENTS
I'm thankful to
V.
Penteriani, J.E. Rabaça, A. Mira, and M.M. Delgado for useful comments and suggestions on this paper. M. Álvaro, M. Basto, S. Chollet, C. Franco, M. Pais. l. Prego and J.P. Silva gave helpful field assistance. S. Santos gave fundamental help in most parts of this study.LITERATURE CITED
Anoyo,
B.E. 1998. Effectof
diet onthe
reproductive successof
Montagu's Hanier Circus pvqarqus. lbis 140:690€93.
Brower,
J.E., J.H. Zar,
and C.N.Ende.
1990.Field and
laboratory methodsfor
general ecology. 3rd edition. Wm.C. Brown Publishers, Dubuque.Callou, C. 1997. Diagnose différentielldes principaux éléments squelettiques du lapin (Genre Oryctolagus) et du liêvre (Genre Lepus)
en
Europe Occidentale. Centre de Recherches Archéologiques CNRS, Valbonne€ophia Antipolis.Calvete,
C.,
E. Angulo,
and R.
Estrada.2005.
Conservationof
Europeanwild
rabbit populations when hunting is age and sex selective. Biological Conservation121:623€34.
Delibes,
M., and
F.
Hiraldo. 1981.
The
rabbit
as
prey
in
the
lberian
Meditenanean ecosystem.ln:
Proceedings of theWorld
Lagomorph Conference (K. Myersand C.
D.Maclnnes, eds.), pp.
614€22.
Guelph.Rui Fazenda Lourenço (2005)
A ecologia trôÍica do Éuforeal Bubo bubo e
t
implicaçôes consermcionistas sn eccsistemas medilerÉnice.Donázar, J.
A.
1989. Variaciones geográficasy
estacionalesen
la alimentación del Búho RealBubo bubo en Navana. Ardeola 36: 25-39.Donázan, J.
4.,
F. Hiraldo, M. Delibes, and R. R. Estrella. 1989. Comparative food habitsof
the
Eagle cnd Bubo bubo andthe
Great Homed @vl Bubo virqinianus in six Palearctic and Nearctic biomes. Omis Scandinavica 20: 298-306.Fener, M., and J. J. Negro .2004. The near extinction of two large european predator§: super specialists pay a price. Conservation Biology
18:344'il9.
Gama, M.
M.
1957. MamÍferosde
Portugal: chavespara
a
sua
detenninação. Museu Zoológico da Universidade de Coimbra, Coimbra.Hakkarainefl,
H.,
E.
Korpimaki,V.
Koivunen,and R.
Ydenberg.2002
Survivalof
male Tengmalm's owts undertemporally varying food conditions. Oecologiaí31: 83€8.
Hiraldo,
F., J.
Andrada,and F. F. Panefio.
1975. Dietof the
Eagle Owl (Eubo-bube) in Meditenanean Spain. Dofiana, ActaVertebrala2:
161-177.Hiraldo, F., F. F. Panefio, V. Andrada, and F. Amores. 1976. Variations in the food habits
of
the european Eagle Owt (Bubo-bubo). Dofiana, Acta Vertebrata 3: 137-156.
lborra, O., C. P. Arthur, and P. Bayle. 1990. lmportance trophique du lapin du garenne pour les grands rapaces provençaux. Vie Milieu
4O:177-188-Jongman, R. H. G., C. J. F. Ter Braak, and O. F. R. Van Tongeren. 1995. Data analysis in community and landscape ecology. Cambridge
Univers§
Press, Cambridge.Madureira,
M.L.,
and M. G.
Ramalhinho. 1981. Notas sobrea
distribuição, diagnose e ecologia do lnsectivora e Rodentia portugueses. Arquivos do Museu Bocage 1: 165-263. Marchesi, L., P. Pedrini, and F. Sergio. 2OO2a. Biases associated with diet study methods inthe Eurasian Eagle-Owl. Joumal of Raptor Research 36: 11-í6.
Marchesi, L., F. Sergio, and
P.
Pedrini. 2O02b. Costs and benefitsof
breedingin
human-altered Iandscapes for the Eagle Owl Bubo bubo. lbis 144:E1U'8177.
Mart[nez, J.
4.,
and l. Zuberogoitia. 2001. The response of the Eagle Owl (Eubo bubo) to an outbreak of the rabbÍt haemonhagic disease. Joumal für Omithologie 142:2O4'21'l-MartÍnez,
J.
4.,
D.
Senano,
and
l.
Zuberogoitia.2003.
Predictivemodels
of
habitat preferences for the Eurasian eagle owl Bubo bubo: a multiscale approach. Ecography 26: 21-28.Mart[nez,
J.
E., and J.F.
Calvo.
2OO1.Diet
and
breeding success
of
Eagle
Owl
in Southeastem Spain: effectof
rabbit haemonhagic disease. Joumalof
Raptor Research 35:259-262.Mathias, M.
L.
1999.
MamÍÍerostenestres
de
Portugal Continental,Açores
e
Madeira. lnstituto da Conservação da Natureza, Lisboa.McGune,
8.,
and
M.J. Mefford. 1999. PC-ORD. Multivariate analysisof
ecologiel
data. Version 4. MjM Software Design, Gleneden Beach, Oregon.Rui Fazenda Lourenço (2d)Q
A ecologia úóÍica do Éuferel Buâo bubo e
a
impllcaçÕes conson acionist* ern eccsistemmMoreno,
E.
1985. Clave osteológica parala
identificaciónde
los Passeriformes ibéricos l.Aegithalidae,
Remizidae,Paridae,
Emberizidae, Passeridae, Fringillidae, Alaudidae. Ardeola 32:295-377.Moreno,
E.
1986. Claveosteolfuie
parala
identÍficación de los Passeriformesibérios
ll.Hirundinidae, Prunetlidae, Sittidae, Certhiidae, Troglodytidae, Cinclidae,
Laniidae, Oriolidae, Corvidae, Stumidae, Motacillidae. Ardeola 33: 69-129.Moreno,
E.
1987. Clave osteológica para la identÍficaciónde
los Passeriformes ibéricos lll. Muscicapidae. Ardeola 34: 243-27 3.Moreno,
S.,
and R.
Villafuerte. 1995. Traditional
managementof
scrubland
for
the conservationof
rabbits Orvctolaqus cuniculus andtheir
predatorsin
Dofiana National Park, Spain. Biological Conservation 73:8í€5.
Ontiveros, D.,
J.
M. Pteguezuelos, andJ.
Caro. 2005. Prey density, prey detectabili§ andfood habits:
the
case
of
Bonelli'seagle and
the
conservation measures. Biological Conservation 1 23: 19-25.Penteriani, V. 1996. The eagle owl. Calderini Edagricole, Bologna.
Penteriani, V., M. Gallardo, and P. Roche. 2OO2. Landscape structure and food supply affect eagle
owl
(Bubo_bgbo) densityand
breeding performance:a
cese of
intra-population heterogeneity. Joumal of Zoology, London 257:365-372.Penteriani, V., M. Gallardo, P. Roche, and H. Cazassus. 2OO1. Effects
of
landscape spatialstructure
and
compositionon the
settlement
of
the
Eagle
Owl
Bubo
bubo
in
a Meditenanean habitat. Ardea 89: 331-340.Penteriani,
V.,
M. M. Delgado, M. Gallardo, and M. Fener. 2004. Spatial heterogenei§ and structure of bird populations: a case example with the eagle owl. Population Ecology 46: 185-192.Pérez,
V.
1980. Alimentación del Búho Real @uOo_OuOo L.) en Espafia Central. Ardeola 25:93-112.
Prenda,
J.,
D.
Freitas,M.
Santos-Reis,and M.
J.
Collares-Pereira. 1997.Guía para
laidentifieción
de
restos óseos pertenecientesa
algunos peces comunesen
las águas continentalesde
la
Penínsulalbérica para
el
estudio
de
la
dieta
de
depredadores ictiófagos. Dofi an a, Acta Vertebrata 24: 1 55-1 80Rivas-MartÍnez, S.,
T.
E. Díaz, F. Femández-González,J.
lzco,J.
Loidi, M. Lousã, and A. Penas. 2OO2. Vascular plant communities of Spain and Portugal. ltinera Geobotanica 15:5432.
Rosenberg, K.
V.,
and R.J.
Cooper. 1990. Approachesto
aviandiet
analysis. Studies in Avian Biology 13: 80-90.Sergio, F., L. Marchesi, and P. Pedrini. 2003. Spatial refugia and the coexistence of a diuma! raptor with its intraguil owl predator. Joumal of Animal Ecology
72:232-245.
Rui Fazenda Lourenço (20[)5)
A ecologia fófica do Buforeal Bub bubo e
*
ímplicações conseruacionistc eÍÍt eccsisteÍnm meditenânicos.Senano, D. 1998. Diferencias interhabitat en la alimentación del Búho Real (Bupg bubo) en
el
Valle Med6
del
Ebro (NE
de
Espafta): efecto
de la
disponibilidadde
conejo (Orvctolasus cuniculus). Ardeola 45: 35-46.Simmons, R. E., D. M. Avery, and G. Avery.
í991.
Biases in diets determined from pellets and remains: conecüon factors fora
mammal and bird-eating raptor. Joumalof
Raptor Research 25:6387.
SPSS lnc. 1999. SPSS Base 10.0 forWindows
Useis
Guide. SPSS lncorporated, Chigago. Tabachnick, B. G., and L. S. Fidetl. 2001. Using multivariate statistics. 4th edition. Allyn andBa@n, Needham Heights.
Thirgood,
S., S.
Redpath. 2000.
Raptorsand Red
Grouse: conservation conflicts and management solutions. Conservation Biology 14:9*1O4-Mllafuerte,
R.,
C.
Calvete,
J. C.
Blanco,
and
J.
Lucientes.1995. lncidence
of
viral haemonhagic disease in wild rabbit populations in Spain. Mammalia 59:651€59.
Villafuerte, R., J. Vifiuela, and J. C. Blanco. 1998. Extensive predator persecution caused by population crash in a game species: the case of red kites and rabbits in Spain. Biological Conservaüon 84: 1 81 -1 88.
Vifiuela, J., and J. P. Veiga.1992.lmportance of rabbits in the diet and reproductive §uccess of Black Kites in southwestem Spain. Omis Scandinavica23:132-138.
Vifiuela, J., and R. Villafuerte. 2003. Predators and Rabbits (Orvctolagus cuniculus) in Spain:
a
key
conflict
for
european raptor
conservation.ln:
Birds
of
prey
in
a
changing environment(D.
B.
M.
Thompson,S.
Redpath,A.
Fiefding,M.
Marquissand C.
A. Galbraith, eds.), pp. 511-526. The Stationery Office, London..Zar, J. H. 1999. BiostatisticalAnalysis. 4th edition. Prentice Hall, New Jercey.
Rui Fazenda Lourenço (2005)
A 6ologia trófica do Éufo-real Bubo buboex implicaç{B conservacionistas eÍn eccsislemas meditenânicc.
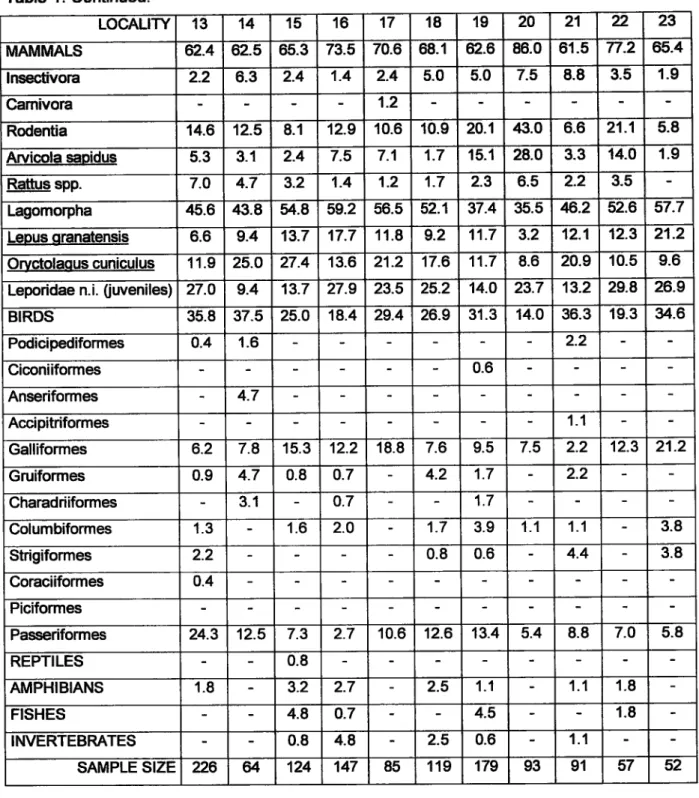
LOCALITY 1 2 3 4 5 6 7
I
I
10 11 12MAMMALS 56.9 67.7 52.3 51.6 52.2 63.6 48.7 60.7 57.4 5í.8 77.9 81.6 lnsectivora 10.9 5.6 í0.8 6.9 11.9 6.8 2.7 5.4 6-6 1.2 1.4
Camivora 1.5 1.3
Rodentia 12.4 6.5 7.7 2.7 1.3.4 12.9 18.4 8.9 10.9 9.0 10.5 12.9
Arvicola sapidus o.7 1.6 9.8 3.9 5.4 6.2 4.8 7.O 7.5
Rattus spp. 8.0 4.O 0.5 10.5 1.5 14.5 2.7 2.3 3.6 4.8
Lagomorpha 33.6 55.6 32.3 42.O 26.9 4Íi.9 28.9 49.1 41.1 36.1 66.3 67.3
Lepus qranatensis 5.1 9.7 7.7 5.9 14.9 5.3 1.3 14.3 8.5 6.6 3.5 1.4
Oryctolaous omiculus 14.6 14.5 9.2 12.2 11.9 '|'9.7 18.4 4.5 23.3 12.O »..1 38.8
Leporidae n.i. fiweniles) 13.9 31.5 15.4 23.9 18.9 9.2 30.4 9.3 17.5 40.7 27.2
BIRDS 38.0 30.6
6.2
4.1
47.8 35.6 28.9 36.6 39.54.6
?2.1 15.0 PodiciFdiformes o.7 1.6 9.0 1.5 7.9 Ciconiiformes 0.7 0.8 1.5 0.8 1.3 0.8 0.6 Anseriformes o.7 1.5 1.1 0.8 0.9 1.2 Accipitriformes 0.8 1.6 Galliformes 2.9 8.1 1.5 2.7 9.0 3.0 2.6 8.0 3.9 5.4 10.5 5.4 Gruiformes 2.9 1.6 9.2 6.9 6.0 10.6 1.3 5.4 10.1 1.2 3.5 1.4Charadriiformes o.7 2.4 3.1 2.1 1.5 0.8 1.3 2.7 0.8 1.2 o.7 Columbiformes 13.9 8.9 18.5 í3.8 9.0 2.3 1.3 5.4 9.3 5.4 1.2 2.7 Strigiformes 9.5 3.2 7.7 6.4 9.0 6.8 3.9 2.7 6.2 1.2 1.2
Coraciiformes 1.3
Piciformes 1.5
Passeriformes 4.4 4.8 3.1 9.6 3.0 9.1 7.9 11.6 7.O 27-',I 5.8 4.8 REPTILES
AMPHIBIANS o.7 0.8 1.1 1.3 2.7
í.6
3.0 3.4FISHES 0.8
INVERTEBRATES 4.4 1.5 3.2 0.8 21.1 1.6 0.6
SAMPLE SIZE 137 124 65 188 67 132 7A 112 129 166 86 147
Table 1. Preys frequency in Eagle Owl localities (Southem Portugal).
Rui Fazenda Lourenço (2fl)5)
A ecologia trófica Oo gufdrd Suôo bubo e
a
impliceçÕes consenncionistas em eco§sistsna meditenánice.,; iW-&,
- i \ .. ," )