Universidade de Trás-os-Montes e Alto Douro
CARACTERIZAÇÃO DO LEITE DE CERVA IBÉRICA
(Cervus elaphus hispanicus). Efeito da idade, número de parto e estado
da lactação na produção e composição do leite.
Dissertação de Mestrado em Engenharia Zootécnica
Sílvia Emanuela Pereira Braga
Orientador
Prof. Doutor Victor Manuel de Carvalho Pinheiro CECAV- UTAD
Coorientador
Prof. Doutor Andrés José García Días IDR- UCLM
iii
Universidade de Trás-os-Montes e Alto Douro
CARACTERIZAÇÃO DO LEITE DE CERVA IBÉRICA
(Cervus elaphus hispanicus). Efeito da idade, número de parto e estado
da lactação na produção e composição do leite.
Dissertação de Mestrado em Engenharia Zootécnica
Sílvia Emanuela Pereira Braga
Orientador: Prof. Doutor Victor Manuel de Carvalho e Pinheiro Coorientador: Prof. Doutor Andrés José García Días
Composição do Júri:
v
“As doutrinas apresentadas neste trabalho são da exclusiva responsabilidade do autor”
vii
Aos meus pais, que são os melhores do mundo, por terem feito de mim quem sou, por alimentarem os meus sonhos, apostarem na minha formação e me fazerem acreditar que o impossível não existe, dedico esta tese.
ix
Agradecimentos
‘’Cada pessoa que passa na nossa vida, passa sozinha. Porque cada pessoa é única e nenhuma substitui a outra. Cada pessoa que passa na nossa vida passa sozinha e não nos deixa só, porque deixa um pouco de si e leva um pouco de nós. Essa é a mais bonita responsabilidade da vida e a prova de que as pessoas não se encontram por acaso” – Charles Chaplin. Assim, não poderia deixar de agradecer a todos aqueles que com um bocadinho de si, contribuíram para a realização deste trabalho.
Ao professor Victor Pinheiro por ter aceitado a orientação desta tese, pelos seus contactos, através dos quais tive oportunidade de integrar a equipa de investigação de Veado ibérico de Albacete e, por toda a ajuda que permitiu a realização deste trabalho.
Ao Professor Andrés García por me ter aceitado e integrado na sua equipa, pelos dados fornecidos para a elaboração desta tese e pela oportunidade de trabalhar nos variados projectos de investigação de Veado ibérico. Ao Professor Laureano Gallego por toda a preocupação. Ao professor Bernardo Albiñana pela paciência e ajuda.
Ao Jamil Capelli, Isabel Tagarroso, Juan Carlos, Giovani e Maria Francesca por terem sido mais do que colegas de trabalho, amigos, e por terem tornado a estadia em Albacete mais fácil, mais rica e emocionante.
Aos meus amigos, minha claque, que sempre torceram por mim e que, mesmo longe estiveram sempre presentes. À Mafalda pela presença constante, pelas compartilhas de desespero, pelas crises de riso e por ser uma leal espectadora desta novela. À Ana Filipa por ser a minha calma, a minha paz e o meu apoio. À Inês Santos, pelo carinho incondicional, pela amizade verdadeira, pelos abraços e colos. À Taís, por ser a maior e pelas histórias e memórias que vão ficar para sempre. À Cláudia e Ana Lima por serem os meus anjos da guarda, as minhas princesas. À equipa masculina do 16 e toda a restante malta da colheita de 2009 pelas farras, abraços e amizades sinceras.
Por fim, à minha família, meu tesouro: aos meus tios e prima, Lena, Jorge e Filipa, por serem tão indispensáveis, tão presentes, tão meus. À minha avó pelo amor interminável, pelas suas orações e preocupações. À minha irmã pelas discussões e contradições naturais sinónimas de um carinho inigualável, pela força e incentivo. E à minha sobrinha pelo amor que ‘é tão grande que não cabe em mil salas’.
xi
Resumo
Este estudo pretende descrever a produção e a composição do leite de veado ibérico (Cervus elaphus hispanicus). Ambiciona descrever a curva de produção e estudar os efeitos da semana de recolha, idade e número de lactação sobre a composição do leite.
Foram controladas 77 fêmeas (entre 2005-2013) criadas em cativeiro e instaladas na exploração de veados da Universidade de Castilha la Mancha. O leite foi recolhido nas semanas 2, 4, 6, 10, 14 e 18, por ordenha mecânica e manual num conjunto de 240 lactações com a duração de 18 semanas. As amostras recolhidas foram analisadas laboratorialmente e assim, determinado o seu teor de proteína, gordura, lactose, extracto seco e células somáticas.
Durante a lactação, as cervas produziram, em média, um total 261,95 L de leite nas 18 semanas e 2,10 L por dia. A composição química média do leite obtida foi de 22,86 % de extracto seco, 9,61 % de gordura, 6,86 % de proteína e 4,47 % de lactose. A produção e composição média do leite evoluíram de forma inversa ao longo das semanas de lactação (P <0,001). Assim, à medida que a lactação se desenvolveu, a produção de leite diminuiu, descrevendo uma curva do tipo II e a concentração dos seus principais componentes aumentou.
O número de células somáticas média do leite foi de 84,79 x 103 cel/mL e verificou-se mais elevada nas primeiras semanas de ordenha. Esta descreveu uma variação decrescente entre a 2ª e 10ª semana e a partir de aí crescente até ao final da lactação (P <0,001).
A idade influenciou significativamente (P <0,05) a produção média de leite, os seus teores em proteína e lactose e o número de células somáticas. As menores produções de leite verificam-se em animais mais precoces e mais velhos e o auge produtivo ocorre por volta dos 5 anos de idade
O número de lactação exerceu uma influência significativa na produção média e total de leite (P <0,01), verificando-se que a produção média de leite foi aumentando ao longo da vida produtiva das cervas até atingir um máximo na 4ª lactação, a partir da qual decresce.
Palavras-chave: Veado ibérico, Leite, Produção, Composição, Células somáticas, Idade, Número de lactação
xiii
Abstract
This study aims to describe the production and composition of milk in the red deer (Cervus elaphus hispanicus). It attempts to describe the milk production curve and to study the effect of the sampling week, age and lactation number on the milk composition.
For this, in the years between 2005 and 2013, 77 females were bred in captivity and installed in the deer exploration facilities of the University of Castilla La Mancha in a controlled environment. Milk was sampled in the 2nd, 4th, 6th, 10th, 14th and 18th week of lactation by mechanical and manual milking totaling 240 lactations during 18 weeks. The gathered samples were analyzed in the laboratory were the protein, fat, lactose, dry extract and somatic cells contents were determined.
During lactation, females produced, in average, 261.95 liters of milk in the milking’s 18 weeks and 2.10 liters per day. The milk average chemical composition obtained was 22.86% of dry extract, 9.61% of fat, 6.86% of protein and 4.47% of lactose. Milk average production and composition evolved in an inverse manner during the lactation weeks (P <0.001). For instance, during lactation, a decrease in milk production was observed, describing a type II curve while the concentration in the principal milk components increased.
The average count of somatic cells in the milk was 84.79 x 103 cel/mL and was higher during the first weeks of milking. The variation of this parameter decreases between the 2nd and 10th week and then increases until the end of lactation (P <0.001).
Age significantly influences (P <0.05) milk production, protein and lactose content and number of somatic cells. Furthermore, older and precocious animals showed the smallest milk production and moreover the peak in production is observed when animals are around 5 years of age.
Lactation number exerts a significant influence in the average and total milk production (P <0.01), and it is possible to observe that the average milk production increases during the females life time until it reaches a maximum in the 4th lactation, after what it decreases.
Key words: Red deer, Milk, Production, Composition, Somatic Cells, Age, Lactation Number.
xv
ÍNDICE
I- INTRODUÇÃO ... 1
II- REVISÃO BIBLIOGRÁFICA ... 3
1. Características gerais do Veado vermelho (Cervus elaphus) ... 5
1.1. Filogenia e sistemática ... 5
1.2. Distribuição mundial ... 6
1.3. Características do veado ibérico (Cervus elaphus hispanicus) ... 8
1.3.1. Taxonomia e distribuição ... 8
1.3.2. Características morfológicas ... 9
1.3.3. Comportamento ... 10
2. Lactação da cerva ibérica ... 12
2.1. Características da lactação ... 12
2.2. Métodos de estimação da produção de leite ... 13
2.2.1. Pesagem dupla ... 14
2.2.2. Ordenha ... 15
2.2.3. Observação dos aleitamentos ... 15
2.2.4. Diluição de isótopos ... 16
2.2.5. Transferência de isótopos ... 17
2.3. Curvas de lactação da cerva ibérica ... 17
2.4. Quantidade de leite ... 20
2.5. Composição do leite ... 22
2.5.1. Comparação com outras espécies ... 23
2.6. Factores que afectam a produção e composição do leite ... 24
2.6.1. Subespécies do animal ... 24
2.6.2. Idade, número de lactação e peso do animal ... 27
2.6.3. Estado da lactação ... 28
xvi
2.6.5. Época do parto ... 30
2.6.6. Peso e sexo das crias ... 31
2.6.7. Número de células somáticas ... 32
III- TRABALHO EXPERIMENTAL ... 35
1. Objectivos ... 35 2. Material e métodos ... 37 2.2. Instalações ... 38 2.3. Animais e maneio ... 44 2.3.2. Partos e identificação ... 45 2.3.3. Alimentação ... 47 2.3.4. Tratamentos sanitários ... 48 2.4. Delineamento experimental ... 48 2.4.1. Nota ética ... 48
2.5. Métodos de recolha de leite ... 49
2.6. Análises laboratoriais ... 52
2.7. Edição dos dados e análise estatística ... 53
IV- RESULTADOS E DISCUSSÃO ... 55
1. Caracterização geral da lactação da cerva ibérica... 57
1.1. Efeito da semana de lactação ... 59
1.2. Efeito da idade ... 65
1.3. Efeito do número de lactações ... 71
V- CONCLUSÕES ... 75
xvii
ÍNDICE DE FIGURAS
Figura 1: Distribuição mundial de veado vermelho, Cervus elaphus, segundo a
União Internacional para Conservação da Natureza ... 7
Figura 2: Características morfológicas do Veado Ibérico: dimorfismo sexual. ... 9
Figura 3: Grupo de fêmeas e crias ... 10
Figura 4: Grupo de machos ... 11
Figura 5: Curvas de lactação de cerva ibérica obtidas a partir dos pesos médios das crias antes e após amamentação. ... 18
Figura 6: Curvas da produção de leite e ingestão, obtidas através de ordenha manual e pesagem dupla, em dois grupos onde a amamentação ocorreu em grupo (A) ou isolamento (B). ... 19
Figura 7: Efeito da subespécie na curva de lactação de fêmea de veado vermelho. Comparação entre C. e. hispanicus e C. e. scoticus. ... 26
Figura 8: Diferenças entre a evolução da composição em gordura, proteína e proteína:gordura nos leites de cerva ibérica e cerva escocesa ao longo da lactação. ... 26
Figura 9: Curvas de lactação de dois grupos de fêmeas submetidas a dois planos nutricionais: dieta padrão e dieta com restrição.. ... 30
Figura 10: Efeito da época de parto na produção de leite. Diferenças entre grupos (P <0,05) são indicadas por asterisco. ... 31
Figura 11: Evolução do NCS nos leites de cabra (gráfico 1), vaca (gráfico 2) e ovelha (gráfico 3) de acordo com o estado da lactação (dias). Comparação entre sucessivas lactações. ... 34
Figura 12: Localização geográfica de Albacete e instalações experimentais de Cervídeos da UCLM (vista via satélite). ... 37
Figura 13: Desenho das Instalações experimentais de Cervídeos da UCLM ... 39
Figura 14: Fotografia via satélite das instalações. Delimitação a vermelho da área correspondente. ... 40
Figura 15: Fotografias dos parques segundo os diferentes grupos. A: Machos reprodutores. B: Animais jovens. C: Fêmeas e crias. ... 40
Figura 16: Parque e vedação. ... 41
Figura 17: Zonas de alimentação: comedouros portáteis e corredor de alimentação…… ... 41
xviii
Figura 18: Mangas de maneio. ... 42 Figura 19: Planta da nave de maneio. ... 43 Figura 20: Pormenores da nave de maneio.. ... 43 Figura 21: Distribuição temporal do número de animais nos anos objectos de
estudo ... 44 Figura 22: Observação da cria e progenitora e sua identificação através
colocação de brinco. ... 46 Figura 23: Pastagem artificial de um parque ... 47 Figura 24: Ficha de registo de partos 2014. ... 50 Figura 25: Mapa da planificação das ordenhas para a época de partos de 2014. . 50 Figura 26: Ficha controlo de ordenha. ... 51 Figura 27: Procedimentos da edição dos dados ... 54
ÍNDICE DE TABELAS E GRÁFICOS
Tabela 1: Subespécies de veado vermelho, Cervus elaphus segundo Geist (1999) ... 7 Tabela 2: Composição bruta de leites de cervídeos em diferentes estados da
lactação e sua comparação com os de algumas espécies de bovídeos. ... 23 Tabela 3: Fontes de variação na performance dos animais durante a lactação. .... 24 Tabela 4: Composição do leite de diferentes subespécies de veado vermelho
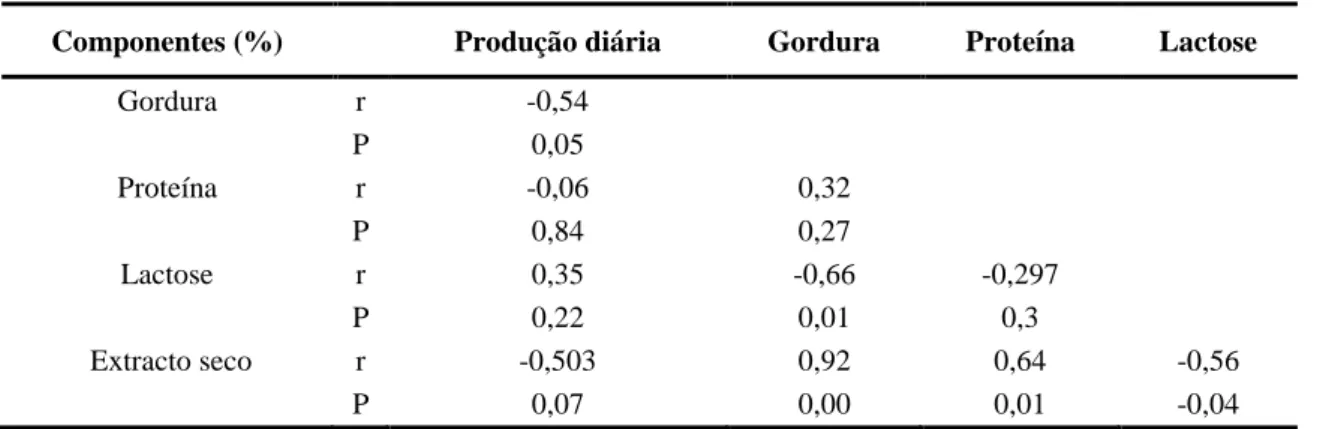
(Cervus elaphus) segundo diferentes durações e etapas de lactação. . 25 Tabela 5: Correlações (r) entre a produção média diária e os distintos
componentes do leite. ... 28 Tabela 6: Dados climáticos (2005 - 2012) da estação meteorológica ''Los
Llanos'' ... 38 Tabela 7: Distribuição temporal dos animais controlados em função da idade e
ano correspondente em estudo. ... 45 Tabela 8: Cor do colar segundo o tipo de animais. ... 46 Tabela 9: Características gerais (valores médios) da lactação da cerva ibérica. ... 59
xix
Tabela 10: Efeito da semana de lactação na produção média diária (PL) e no seu teor de gordura (G), proteína (P), lactose (L), extracto seco (ES) e número de células somáticas (CSS). ... 63 Tabela 11: Efeito da idade na produção média diária (PL) e no seu teor de
gordura (G), proteína (P), lactose (Lact), extracto seco (ES) e número de células somáticas (CSS). ... 69 Tabela 12: Efeito da idade na produção total de leite (PL), de gordura (Gt),
proteína (Pt), lactose (Lt) e extracto seco (ESt). ... 70 Tabela 13: Efeito do número de lactação na produção média diária de leite
(PL) e no seu teor de gordura (G), proteína (P), lactose (L), extracto seco (ES) e número de células somáticas (CSS) ... 73 Tabela 14: Efeito do número de lactação na produção total de leite (PL), de
gordura (Gt), proteína (Pt), Lactose (Lt) e extracto seco (ESt). ... 74
Gráfico 1: Produções totais de leite (PLt; L) obtidas nos objectos de estudo ... 57 Gráfico 2: Evolução da produção média diária de leite (mL) de cerva ibérica e
sua composição química média ao longo da lactação ... ….60 Gráfico 3: Evolução da produção média diária de leite, seus constituintes (%
gordura, proteína, lactose e extracto seco) e número de células somáticas de acordo com a idade. ... 65 Gráfico 4: Evolução da produção média diária de leite e seus constituintes de
xxi
LISTA DE ABREVIATURAS
ANPC Associação Nacional de proprietários e produtores de caça IUCN União Internacional Conservação da Natureza
UCLM Universidad Castilla-La Mancha
C. e. Cervus elaphus 2 H2O Óxido de Deutério 3 H2O Óxido de Tritio L Litro mL Mililitro Ca Cálcio K Potássio Mg Magnésio P Fosforo Mn Manganês Na Sódio Zn Zinco Fe Ferro
r Coeficiente de correlação de pearson NCS Número de células somáticas (1000/mL)
km Quilometro
m Metro
mm Milímetros
cm3 Centímetros cúbicos
h Hora
E.U.A Estados Unidos da América
PL Produção de leite diária
%G, P, L, ES Percentagens de gordura, proteína, lactose e extracto seco PLt Produção de leite total (L) produzida durante toda lactação
Gt, Pt, Lt, ESt Quantidade de gordura, proteína, lactose e extracto seco total (kg)
s Semana de lactação
N Número de amostra
1
I- INTRODUÇÃO
A caça é uma actividade com importante impacto tanto a nível do desenvolvimento sócio-económico das zonas agro-rurais como a nível de gestão de território, da paisagem e dos recursos naturais em geral.
Em Portugal, tem havido uma melhoria na gestão e ordenamento dos recursos cinegéticos mas ainda insuficientemente geradora de receitas se a compararmos com as potencialidades por explorar. Segundo Associação Nacional de Proprietários e Produtores de Caça, o nosso país apresenta um potencial que poderia gerar cerca de 1000 milhões de euros, contudo este sector gera apenas um valor económico entre os 400 e os 500 milhões de euros por ano (ANPC, 2007).
Em Espanha, a caça tem uma importância económica mais elevada e, segundo dados do director geral da Real Federação espanhola de caça esta gera, actualmente, cerca de 3,6 mil milhões de euros (Martín, 2012).
O veado ibérico (Cervus elaphus hispanicus) é uma das principais espécies cinegéticas de caça maior da Península Ibérica. Encontra-se mais amplamente distribuído por Espanha, onde a sua exploração consiste principalmente na obtenção de troféus cuja valorização tem vindo a causar a expansão deste sector.
Como resultado da expansão do sector cinegético privado, aumentou a procura dos produtores pela tomada de medidas objectivas sustentadas por resultados científicos de forma a implementar uma gestão mais rigorosa. Assim, foram surgindo equipas de investigação, com objectivo de incrementar o conhecimento do veado ibérico e factores que afectam a sua produção.
A linha de trabalho seguida até ao momento nas instalações experimentais do departamento de ciência e tecnologia agroflorestal da escola técnica de engenheiros agrónomos da Universidade de Castilla-La Mancha, em Albacete, tem centrado os seus estudos nos diferentes aspectos reprodutivos e produtivos do veado ibérico. Parte destes estudos têm compreendido a produção de leite das cervas e o crescimento dos gabatos nas primeiras fases da sua vida, devido ao facto de que este período engloba o seu crescimento mais rápido e que as variáveis que afectam a lactação podem afectar o peso adulto e consequentemente a qualidade e desenvolvimento das hastes. Assim, alguns resultados já obtidos indicam uma importante influência da produção e composição do leite sobre o crescimento do gabato. Por exemplo, no que diz respeito à influência da composição do leite de cerva ibérica, o teor em proteína parece ser o factor com o efeito
2
mais importante no desenvolvimento das crias (Landete-Castillejos et al., 2001; Landete-Castillejos et al., 2003b; Ceacero, 2010). Os partos tardios (que ocorrem mais tarde na época de partos) afectam a produção e composição do leite, principalmente através da diminuição da quantidade da proteína disponível o que afecta negativamente o crescimento dos gabatos (Landete-Castillejos et al., 2000b). Além disso, foi provado que diferenças de peso que se estabelecem durante a primeira fase da vida destes animais podem manter-se até aos 18 meses afectando assim todo desenvolvimento ao longo da sua vida produtiva (Landete-Castillejos et al., 2001).
Assim, com o objectivo de aprofundar o conhecimento das características da lactação de cerva ibérica, conhecer de que forma evoluem a produção e a composição do leite ao longo da lactação e da sua vida produtiva e estudar alguns factores que influenciam a sua variação, foram analisadas as lactações compreendidas entre 2005 e 2013, de 72 fêmeas de veado ibérico das instalações experimentais da UCLM.
3
II- REVISÃO BIBLIOGRÁFICA
5
1. Características gerais do Veado vermelho (Cervus elaphus)
1.1. Filogenia e sistemática
Ruminantia é uma das quatro secções nas quais está englobada a ordem Artiodactyla (animais mamíferos ungulados com um número par de dedos nas patas) que desde o Neógeno representa o grupo mais diverso ecológica e geograficamente de ungulados vivos no mundo (Janis e Scott, 1987). Dentro desta subordem encontra-se a infra-ordem Pécora, pela qual se distribuem as famílias Cervidae, Moschidae, Giraffidae e Bovidae (Brooke, 1878). Com mais de quarenta espécies de veado, a Cervidae constitui a segunda maior família no que diz respeito ao agrupamento de espécies, logo a seguir à família Bovidae (Gilbert et al., 2006).
Apesar de ser altamente divergente no que diz respeito ao tamanho, habitat e comportamento dos animais que a constituem, as espécies da família Cervidae estão unidas por uma série de sinapomorfias (caracteres que, surgindo ao longo da evolução se mantêm em diversos grupos taxonómicos) que as distinguem das outras famílias. Entre as principais características próprias da família Cervidae, encontram-se formações cilíndricas ósseas, de crescimento descontínuo, que se ramificam a partir do crânio nos machos (à excepção das espécies do género Rangifer, onde estas se podem encontrar também nas fêmeas), a presença de molares braquiodontos, a presença de dois orifícios à saída dos ductos lacrimais situados dentro da órbita, na fossa lacrimal ou no orifício anterorbital, e por fim a existência da primeira e segunda falanges dos dedos laterais (Brooke, 1878; Janis and Scott, 1987; Gilbert et al., 2006).
A família Cervidae encontra-se por sua vez dividida em quatro subfamílias, Muntiacinae, Hydropotinae, Odocoileinae e por fim Cervinae, onde se encontram os cervídeos do género Cervus e espécie C. elaphus conhecida em todo o mundo como veado vermelho (Gilbert et al., 2006).
García (2000) considera a contabilização das subespécies do género C. elaphus ainda um pouco controversa e variável entre autores, podendo considerar-se 12, 20 ou 23 subespécies, segundo Clutton-brock. et al. (1982), Brelurut et al. (1990) e Whitehead (1993), respectivamente.
Segundo a União internacional para a conservação da natureza e recursos naturais (IUCN, 2014), a classificação taxonómica do veado vermelho organiza-se da seguinte forma:
6 Reino: Animalia Filo: Chordata Classe: Mammalia Ordem: Artiodactyla Família: Cervidae Género: Cervus
Espécie: Cervus elaphus (Linnaeus, 1758)
1.2. Distribuição mundial
Dentro da família Cervidae, o veado vermelho é a espécie com mais ampla distribuição no mundo. A sua localização estende-se da Europa e Norte de África, até à Ásia central, Sibéria, extremo Oriente e América do Norte, como se pode ver no mapa da figura 1. Apesar de anteriormente existir por todo o Canada e Norte da América, esta espécie está agora restrita ao Oeste da América do Norte (cujas populações são já provenientes de repovoamentos). Na Europa continental encontra-se amplamente distribuída e pode ser facilmente encontrada. A sua presença é também notada numa série de ilhas como as ilhas britânicas, com uma considerável população na Escócia, e Sardenha. Na Rússia existem várias pequenas populações (todas introduzidas em zonas de caça) de origem desconhecida, tal como na Grécia, cujas subpopulações são também resultado de introduções de animais de origem exterior. O mesmo acontece em relação a Portugal, cuja população foi proveniente da reintrodução ou expansão natural de animais provenientes de Espanha. No continente africano estes animais podem encontrar-se na Argélia e Tunísia, no médio Oriente na Turquia, Irão e Iraque e na Ásia central, numa série de países desde o Cazaquistão até à Mongólia e China (IUCN, 2014).
Como já foi dito anteriormente, a classificação das subespécies, segundo a sua origem, é ainda controversa e gera alguma discordância entre autores. Na tabela 1 encontram-se 22 subespécies conhecidas de veado vermelho, segundo Geist (1999), os seus nomes comuns e localização geográfica obtidos a partir de uma revisão de Ludt et al. (2004).
7
Figura 1: Distribuição mundial de veado vermelho, Cervus elaphus, segundo a União Internacional para Conservação da Natureza.
Fonte: IUCN em http://maps.iucnredlist.org/map.html?id=41785.
Tabela 1: Subespécies de veado vermelho, Cervus elaphus segundo Geist (1999).
Subespécies
Cervus elaphus
Nome comum
Origem geográfica
Inglês Português*
barbarus Barbary RD. Bárbaro Tunísia
corsicanus Sardinian RD. Veado-da-Córsega ou Veado-Sardo Sardenha
hippelaphus Midle-European RD. Veado da Europa central Bulgária
montanus Eastern RD. Veado do Oriente Romania
alashanicus Alashan RD. Veado Alashan* China
xanthopygus Isubra Veado Isubra Rússia
maral Maral Veado Maral Irão
nelsoni RockyMountain W. Veado Norte-Americano de montanha Norte América
canadensis American W. Veado Norte-Americano Norte América
sibericus Siberian W. Veado da Sibéria Mongólia, China
songaricus Tien Shan W. Veado Tien shan* TienShan, China
Hanglu Kashmir W. Veado-caxemira India
kansuensis Kansu RD. Veado Kansu* Dong Da Shan, China
macneilli M’Neill’s RD. Veado M'Neill's* Quinghai, China
wallichi Shou Veado do Tibete Tibete, China
bactrianus Bactrian RD. Veado Bactrian* Tadjiquistão
yarkandensis Yarkand RD. Veado Yarkande* China
atlanticus Atlanticus RD. Veado da Noruega Noruega
brauneri Krim RD. Veado Krim* Ucrânia
elaphus RD. Veado vermelho Suécia
hispanicus Iberian RD. Veado ibérico Espanha e Portugal
scoticus Scotic RD Veado escocês Escócia RD: Red deer; D: Deer; W: Wapiti; * - Por não existir a designação e Português, para estas subespécies a tradução foi feita à “letra”
8
1.3. Características do veado ibérico (Cervus elaphus hispanicus)
1.3.1. Taxonomia e distribuição
É possível que os veados nativos da Europa ocidental possam ter derivado, pelo menos parcialmente, de populações refugiadas na Península Ibérica, sobreviventes do último máximo glacial e que posteriormente se expandiram para o Norte no início do Holoceno. No entanto, a filogenia e estrutura genética das populações de veado vermelho da Península Ibérica são ainda hoje pouco conhecidas e a sua taxonomia, formalmente conhecida por Cervus elaphus hispanicus, tem vindo a ser bastante debatida. Fernández-Garcia et al. (2013) referem que Cabrera (1914), baseando-se em diferenças morfológicas juntou à subespécie C. e. hispanicus a subespécie C. e. bolivari, considerando assim que as populações da Península Ibérica seriam constituídas por duas subespécies. Contudo, hoje em dia considera-se que essas diferenças são apenas resultado de distintas dietas alimentares e biótopos (Fernández-García et al., 2013) pelo que alguns autores não as distinguem.
De acordo com alguns relatos arqueológicos e históricos, é possível que o veado vermelho possa ter estado continuamente presente na Península Ibérica a partir do final do Pleistoceno (Sommer et al., 2008).
Distribuindo-se plenamente por todo o território ibérico, o veado vermelho foi uma das espécies de ungulados mais abundantes nas matas espanholas e a sua presença foi bastante relatada desde a idade média até ao final do seculo XIX, época a partir da qual, a sua distribuição foi drasticamente reduzida devido à expansão agrícola e caça abusiva. A partir de aí, as populações de veado vermelho sobreviveram isoladas no sudoeste da península e regiões centrais de Espanha, desde os montes de Toledo até às margens do rio Guadalquivir (Perez et al., 1998; Fernández-García et al., 2013). Actualmente, a distribuição de veado ibérico é resultado de uma expansão que tem vindo a ocorrer principalmente devido a repovoações cinegéticas por todo o território peninsular, com excepção da parte mais ocidental da Galiza e da costa levantina, e com maior frequência em zonas de caça existentes em Andaluzia, Castilla-La Mancha e Extremadura (Carranza, 2011).
Em Portugal, o veado vermelho existe principalmente nas regiões transfronteiriças (Contenda Barrancos, Tejo Internacional e Montesinho) e na Serra de Lousã (Torres, 2011).
9 1.3.2. Características morfológicas
O veado ibérico é uma das quatro subespécies de veado vermelho que habitam na Europa continental, juntamente com o veado escocês, o veado da Europa Central e o veado da Noruega. Difere das outras subespécies pelo seu menor tamanho, provavelmente devido à baixa latitude e às temperaturas mais elevadas das zonas pelas quais se distribui, pela ausência de uma zona abundante de pêlo formando uma juba, no pescoço e pela menor marcação da delimitação do escudo anal, que é nesta subespécie um pouco menos visível (Carbó, 1997). A sua pelagem é curta, predominantemente castanha, com uma tonalidade que varia de acordo com a estação do ano de castanho-avermelhado no Verão para castanho-escuro no Inverno, à excepção do ventre cuja cor é sempre ligeiramente mais clara que o resto do corpo. Diferindo dos animais adultos, os gabatos apresentam uma pelagem característica, de fundo castanho, com pequenas manchas redondas brancas que se mantêm durante aproximadamente os primeiros três meses de vida (Carranza, 2011).
O cervo ibérico apresenta um grande dimorfismo sexual. Para além de terem presentes as hastes, os machos são mais robustos, mais pesados e altos que as fêmeas, com pesos entre os 130-150 kg, comprimentos entre os 180-220 cm e alturas à cernelha entre 110-140 cm, em comparação com os 80-100 kg, 160-200cm de comprimento e 90 a 120 cm de altura das fêmeas (Azorit et al., 2002). Na figura 2 podem ser observadas algumas características dos dois indivíduos, macho e fêmea, de veado ibérico.
Figura 2: Características morfológicas do Veado Ibérico: dimorfismo sexual.
10 1.3.3. Comportamento
O veado ibérico caracteriza-se por ser um animal gregário de actividade crepuscular. Os indivíduos de ambos os sexos vivem em grupos separados a maior parte do ano. As fêmeas formam pequenos grupos familiares através de relações de parentesco, geralmente constituídos por uma matriarca, mais velha, fêmeas adultas e suas crias, incluindo machos jovens até aos 2-3 anos, idade a partir da qual abandonam o grupo familiar (Ceacero et al., 2007; Carranza, 2011). Dentro dos grupos existe uma marcada hierarquia estabelecida por relações de dominância, condicionadas pelas relações de parentesco, que são visíveis através de interacções entre os indivíduos que condicionam o seu acesso aos recursos do meio ambiente. O estatuto social pode ser estabelecido de acordo com a idade, peso, grau de parentesco, sexo da cria e data de parto. As relações hierárquicas podem facilmente ser observadas através de interacções agressivas entre as fêmeas matriarcas, mais velhas e mais pesadas (dominantes) e as jovens (principalmente de outros grupos familiares), dificultando assim o seu acesso aos recursos alimentares (Ceacero et al., 2007; Carranza, 2011).
Na figura 3 pode ver-se um grupo familiar, composto por fêmeas e suas crias e na figura 4, um grupo de machos.
Figura 3: Grupo de fêmeas e crias
11 Figura 4: Grupo de machos
Fonte: IDR, 2012
Em condições de cativeiro as cervas tendem a ficar gestantes pela 1ª vez entre os 15-18 meses de idade, quando atingem normalmente 60-70% do seu desenvolvimento corporal e pesam cerca de 60-65 kg de peso vivo. Contudo, podem ficar gestantes no primeiro ano de vida se atingirem aí o desenvolvimento e peso acima descritos ou mais tarde se não reunirem essas condições (García, 2000; Carrión, 2010).
Na época reprodutiva os grupos reorganizam-se formando haréns, grupos polígamos, onde um macho adulto está presente e pode fecundar várias fêmeas. Esta espécie tem um período de cio muito marcado e característico, ao qual se dá o nome de berrea, no qual os machos emitem fortes bramidos e lutam entre si pela defesa do território e manutenção de fêmeas. Este comportamento dura aproximadamente 3 a 4 semanas e ocorre entre finais de Agosto e início de Outubro, fazendo coincidir o parto com a época do ano com maior disponibilidade alimentar, a Primavera. A gestação dura aproximadamente 230-240 dias, nascendo normalmente a cria entre os meses de Maio a Junho, onde ocorre a máxima produção de biomassa vegetal e as hipóteses de sobrevivência das crias são maiores. Normalmente nasce uma cria por parto, com um peso entre 6 a 7 kg que pode ser triplicado em apenas 4 meses. A lactação pode prolongar-se até Dezembro ou Janeiro (Soriguer et al., 1994).
12
Quanto aos seus comportamentos alimentares, o veado é um animal ruminante e dedica 64% do seu tempo activo em tarefas relacionadas com a alimentação. A sua dieta inclui culturas herbáceas e arbóreas e frutos, consoante a zona que habita e consoante a disponibilidade de alimento, que varia com a estação do ano. Contudo, sabe-se que esta espécie tem uma preferência por culturas herbáceas de leguminosas (Carranza, 2011).
2. Lactação da cerva ibérica
2.1. Características da lactação
O estudo da glândula mamária foi fundamental para o estudo da evolução e para a classificação taxonómica das espécies animais. A presença e capacidade de secreção desta glândula fornecem a base para o agrupamento taxonómico das espécies da classe Mammalia e desafia o estudo da lactação como um processo complexo aliado à teoria da evolução das espécies, cujo objectivo é assegurar a sobrevivência da descendência através do fornecimento de leite (Oftedal, 2002).
A produção de leite pelos mamíferos proporciona uma alimentação completa às suas crias desde o seu nascimento, fase em que estas são completamente dependentes da sua progenitora até ao desmame. Assim sendo, a lactação permite ao neonato continuar o seu crescimento de maneira similar à sua fase fetal, sem que este esteja anatomicamente ligado à sua mãe, libertando-a da carga de um feto excessivamente grande (Pond, 1977). Para além de satisfazer as necessidades nutricionais das crias recém-nascidas, o leite aumenta também a sua taxa de sobrevivência através da promoção da competência imunológica e da maturação do sistema endócrino (Capuco e Akers, 2009).
Ao contrário dos machos, para os quais o sucesso reprodutivo é traduzido pelo número de fêmeas que estes cobrem durante um ano (Pond, 1977), para as fêmeas este traduz-se principalmente na sua capacidade para suprimir as necessidades das suas crias, através de uma transferência de nutrientes e elementos do seu sistema imunitário, antes e após o parto (Clutton-Brock e Guinness, 1982). Em todos os mamíferos, a contribuição pós-natal maternal é, em geral, quantitativamente maior que a pré-natal, fazendo da lactação uma medida do sucesso reprodutivo (Oftedal, 1985).
A lactação é um período crucial na vida da fêmea. Os seus custos energéticos são superiores aos da gestação e constituem 43% da energia total necessária durante o
13
ano (Kelly et al., 1987 citados por Gómez et al., 2006). Esta demanda energética pode levar à deploração da condição corporal, que se não for controlada através da suplementação alimentar pode reduzir a taxa de fertilidade da fêmea no ano seguinte (Clutton-Brock et al., 1982). A baixa condição corporal e a falta de reservas para suportar a lactação resultam numa baixa produção de leite, com uma composição baixa em nutrientes afectando desta forma a taxa de sobrevivência e crescimento das crias (Landete-castillejos et al., 2003a).
Tal como todas as espécies animais da classe Mammalia, as fêmeas de veado ibérico modelam a sua produção e composição de leite segundo as necessidades das suas crias assegurando assim a sua sobrevivência e desenvolvimento pós-natal (Landete-Castillejos et al., 2000a).
A produção e a composição de leite ao longo da lactação e os factores que as influenciam como por exemplo a data de nascimento estão correlacionadas com o crescimento dos gabatos (Landete-Castillejos et al., 2000b; Landete-Castillejos et al., 2001), tendo uma notável influência no peso adulto (Landete-Castillejos et al., 2001; Gómez et al., 2006) e, consequentemente na sua eficiência reprodutiva (Clutton-Brock, 1988).
Segundo Landete-Castillejos et al. (2001), o peso ao nascimento e os ganhos médios diários durante a lactação das crias de veado vermelho estão positivamente correlacionados com a capacidade de produção de leite das suas mães durante esta etapa. Este autor afirma também que diferenças no peso no final da lactação continuam a ser notadas após 18 meses de vida. Assim, a lactação e as primeiras etapas do crescimento podem exercer, por exemplo nos machos, uma considerável influência no seu futuro peso adulto e consequentemente no tamanho das hastes, visto que existe uma forte correlação positiva entre o peso vivo e o tamanho das hastes (Gómez et al., 2006).
2.2. Métodos de estimação da produção de leite
Oftedal (1985) refere a pesagem dupla, a ordenha manual ou mecânica, a diluição de isótopos e transferência de isótopos como os quatro principais métodos que se usam para estimar a produção de leite, advertindo contudo que estes métodos constituem por si só uma fonte de variação para a estimativa. Para além desses métodos, também a observação de aleitamentos se encontra com frequência na literatura como um
14
estimador da transferência de leite da mãe para a sua cria (Clutton-Brock e Guinness, 1982; Gauthier e Barrette, 1985).
Todos os métodos interferem de alguma forma com o comportamento natural da progenitora ou da cria, podendo alterar a capacidade de produção pela primeira ou a ingestão pela segunda, assim é difícil fazer uma estimativa precisa (Van der Linden et al., 2010). Contudo, através da minimização dos factores de variação que afectam a precisão é possível minimizar os erros e ter uma estimativa aproximada das produções.
2.2.1. Pesagem dupla
A técnica da pesagem dupla tem sido amplamente utilizada na literatura como um estimador da produção de leite (Arman et al., 1974; Pettigrew e Sower, 1985; Benson et al., 1999; García et al., 1999). O procedimento consiste na pesagem da cria antes e após a amamentação de forma a verificar a variação de peso obtida após ingestão de leite. Antes da amamentação, a progenitora e sua cria são separadas por um determinado período de tempo, considerando-se que a produção de leite durante esse período é estimada como a diferença de peso do gabato antes e depois de mamar (Gomez, 2004).
A utilização desta técnica apresenta uma série de limitações que devem ser consideradas. Visto que não se estima exactamente a produção de leite pela mãe mas sim a ingestão pela cria (Landete-Castillejos et al., 2000a) esta técnica apresenta uma variabilidade maior que a ordenha (Beal et al., 1990; Benson et al., 1999). Devido ao maneio necessário para a separação, junção e separação e pesagem dos animais, o comportamento normal de produção das fêmeas e ingestão das crias pode ser alterado, tal como os resultados da estimativa, que perdem ainda mais precisão no final da lactação, dada a necessidade de medir pequenas quantidades de incrementos de peso em animais maiores e mais activos (Oftedal, 1985). Além disto, durante o aleitamento e pesagem é difícil estimar as perdas urinárias e fecais associadas à ingestão do leite (Benson et al., 1999).
15 2.2.2. Ordenha
A valorização da produção de leite mediante o uso de ordenha pode ser realizada tanto de forma manual (Mueller and Sadleir, 1977; Krzywinski et al., 1980; Castillejos et al., 2000a,b) como de forma mecânica (Arman et al., 1974; Landete-Castillejos et al., 2003a,b,c).
A ordenha de animais selvagens, como é o caso do veado ibérico, requer o uso de anestesia de forma a permitir o seu manuseamento e de oxitocina, para induzir a libertação do leite. Contudo, este tipo de maneio e a utilização destas substâncias pode alterar a composição do leite (Arman et al., 1974; Landete-Castillejos et al., 2002; Landete-Castillejos et al., 2003a,b,c).
Segundo Oftedal (1985) os resultados obtidos através dos diferentes métodos apresentam variações de acordo com o estado da lactação. No caso da ovelha a produção de leite obtida através da ordenha é maior do que a estimada pelo método da pesagem dupla, especialmente nas primeiras semanas após o parto, o que sugere que as crias não conseguem nesta etapa da lactação esvaziar completamente o úbere das suas mães. O mesmo foi encontrado por Landete-Castillejos et al. (2000a) na espécie de veado ibérico para a capacidade de ingestão da cria na primeira parte da lactação, contudo foi encontrada também com uma inversão posterior dessa capacidade, pois o gabato passa a consumir mais leite do que o produzido pela sua progenitora, o que o leva a procurar leite de outras fêmeas do grupo.
2.2.3. Observação dos aleitamentos
Como o próprio nome indica, este método implica a observação e registo de aleitamentos para desta forma estudar o número de vezes que o gabato mama e sua duração. Baseia-se principalmente na suposição de que a cria que mama mais tempo deve obter proporcionalmente mais leite, sempre e quando todas as demais variáveis que o possam afectar (sexo, idade do gabato, espécie, número de parto da mãe, entre outros) se mantenham invariáveis (Cameron, 1998).
Segundo Birgersson e Ekvall (1994) existem duas principais fontes de variação que são importantes de controlar, para que a observação de aleitamentos possa ser uma medida relativamente fiável, para estimativa da produção e consumo de leite. Estas são a duração do tempo de amamentação nas primeiras duas semanas pós-parto, pois os
16
gabatos vão aperfeiçoando a capacidade de sucção, e a experiência da progenitora para libertar o leite, visto que crias de mães primíparas mamam durante mais tempo que as de multíparas apesar de obterem a mesma quantidade de leite.
Cameron (1998) refere ainda a variedade de metodologias utilizadas para mensurar o comportamento da amamentação, a habilidade de mamar da cria, a motivação para a amamentação (fome e outros), a experiência maternal (número de lactações e capacidade leiteira), a variação da composição do leite e, por último a contabilização de falsos aleitamentos, como factores de variação difíceis de controlar e que podem tornar inválida a estimativa através deste método. Assim, este autor sugere resultados obtidos de estudos que assumem que o tempo de amamentação é proporcional à ingestão de leite apenas são válidos após esta relação ser testada empiricamente através de outros métodos, como por exemplo a transferência de isótopos.
2.2.4. Diluição de isótopos
Macfarlane et al. (1969) desenvolveram uma técnica para estimar a ingestão de leite pelas crias através da diluição de isótopos de hidrogénio. Sendo que, mais de 80% do leite é constituído por água e a combustão de hidrogénio nos sólidos do leite produz um volume de água próximo ao dos sólidos, a medição deste turnover de água nas crias oferece uma estimativa próxima da ingestão do leite.
Considerando que a principal fonte de água para o animal é o leite, a sua ingestão pode ser estimada através da medição do volume total de água no corpo utilizando a diluição de óxido de deutério (2H2O), que é um isótopo estável que se
distribui rápida e uniformemente na água corporal. Este é injectado via intravenosa nas crias e dilui-se no sangue até atingir uma equilibrada distribuição. É esperado algum tempo e são retiradas amostras sangue, através das quais é analisado o teor em 2H2O
(Van der Linden et al., 2010).
Apesar da técnica de diluição não interferir com a rotina diária dos animais (não exigindo pesagem, ordenha e outros) requer que a única fonte de água exógena das crias seja proveniente do leite, restringindo desta forma a sua aplicação quando os animais iniciam a ingestão de forragem e água (Holleman et al., 1975).
17 2.2.5. Transferência de isótopos
Devido ao facto do uso simples de isótopo óxido de deutério sobrestimar a ingestão de leite poucas semanas após o nascimento com a iniciação da ingestão de plantas forrageiras por parte dos ruminantes, foram desenvolvidos métodos aplicáveis a toda a lactação baseados na injecção de isótopos marcadores na fêmea lactante e seu seguimento e monitorização no leite e nas crias. Entre estes métodos encontra-se o uso de óxido de trítio (3H2O) e sua transferência no leite desde a mãe até à cria, sendo a
determinação do turnover desta água na cria feita mediante o uso de óxido de deutério e o uso do isótopo radiocesium cuja transferência do leite da mãe para a cria pode ser contabilizada através de técnicas de avaliação da carga corporal desta substância na cria (Holleman et al., 1975).
Estes métodos têm como inconvenientes a necessidade de trabalhar com material radiactivo, a recolha de amostras sanguíneas e o seu envio a laboratórios sofisticados para análise de isótopos, resultando em custos elevados. Contudo, são capazes de estimar de forma mais precisa a quantidade de leite ingerida do que alguns dos outros métodos, como a observação de aleitamentos e a pesagem antes e após mamada (Pettigrew e Sower, 1985; Cameron, 1998).
2.3. Curvas de lactação da cerva ibérica
A lactação, ou mais precisamente, a quantidade de leite produzida através do tempo pode ser expressa num gráfico que os relaciona e que se designa vulgarmente por curva de lactação. Nos mamíferos a curva de lactação padrão vai aumentando à medida que aumentam os requerimentos das crias até um máximo, chamado pico de lactação, a partir do qual vai decrescendo progressivamente, pois a progenitora deixa de conseguir fisiologicamente aumentar a quantidade produzida e deixa de satisfazer todas as necessidades nutricionais da sua cria, que inicia assim uma gradual substituição do leite por alimentos sólidos, que vai aumentando, até ao seu desmame (Landete-Castillejos e Gallego, 2000).
A lactação das fêmeas de veado vermelho dura aproximadamente 4 meses, mas pode estender-se até ao próximo cio, caso esta não fique gestante, durando aproximadamente 12 meses (Clutton-Brock e Guinness, 1982).
18
A fêmea pode apresentar dois tipos de curvas de lactação. A curva de tipo I e a curva de tipo II, estando ambas relacionadas com a sua capacidade de mobilização das reservas corporais para a produção de leite (figura 5). A curva de tipo I tem a forma da curva padrão das fêmeas da classe dos mamíferos e ascende com as crescentes necessidades energéticas da cria desde o parto até alcançar uma produção máxima entre as semanas dois e quatro, a partir das quais decresce até à completa secagem da fêmea e desmame da cria. A curva de tipo II é uma curva decrescente desde o início da lactação e é normalmente atribuída a animais com piores condições corporais que possam ter sido submetidas a restrições alimentares (Loudon et al., 1983; García et al., 1999; Landete-Castillejos e Gallego, 2000; Landete-Castillejos et al., 2003a).
Como foi descrito no ponto anterior, têm vindo a ser desenvolvidos vários métodos para estimar a produção de leite. E, com o intuito de monitorizar os resultados produtivos em curvas de lactação, vários modelos matemáticos têm sido formulados. Na figura 5, encontram-se representados os dois tipos de curvas de lactação de cerva ibérica, cuja produção de leite foi estimada através do método de pesagem dupla e inserida no modelo matemático desenvolvido por Wood, num estudo de García et al. (1999).
Figura 5: Curvas de lactação de cerva ibérica obtidas a partir dos pesos médios das crias antes e após amamentação.
19
Apesar da curva de tipo II ser reconhecida nas demais espécies animais como a curva associada a dietas alimentares pobres, na cerva ibérica foi encontrada, juntamente com a curva de tipo I, em animais submetidos às mesmas condições alimentares (García et al., 1999; Landete-Castillejos et al., 2000a).
Num estudo de Landete-Castillejos et al. (2000a) em que foi comparada a ingestão de leite das crias (estimada a partir da técnica da pesagem antes e após amamentação) e a produção de leite (obtida por ordenha manual) das suas progenitoras, segundo duas experiências em que a amamentação ocorreu de duas formas: em grupo ou em isolamento, foi obtida uma curva do tipo I no que diz respeito à ingestão de leite das crias e uma curva de tipo II no que diz respeito à produção de leite das fêmeas (figura 6).
Figura 6: Curvas da produção de leite e ingestão, obtidas através de ordenha manual e pesagem dupla, em dois grupos onde a amamentação ocorreu em grupo (A) ou isolamento (B).
20
Apesar da curva de produção decrescente, as fêmeas de ambas as experiências, em grupo ou em isolamento, mostraram produzir mais leite do que o ingerido pelas suas crias entre as semanas 1 e 5 (figura 6- gráfico A e B), o que não acontece a partir da semana 6, na experiência em que a amamentação ocorreu em grupo (gráfico A), onde as crias ingerem 17,2% mais leite que o produzido pelas suas mães, possivelmente através de pequenos furtos de leite a outras fêmeas do grupo, num comportamento denominado ‘allosuckling’. Assim, os autores concluíram que o excesso de leite produzido nas primeiras 5 semanas de lactação é uma forma de garantir que a cria recebe todos os nutrientes e componentes de defesa a partir do leite da sua mãe nesta fase de maior perigo de mortalidade. A partir desta fase as taxas de allosuckling tendem a aumentar devido à diminuição da produção, desempenhando também um papel importante para a produção da glândula mamária das cervas que sofreram um aborto tardio ou cuja cria morreu no início da lactação, visto que este comportamento pela parte de gabatos de outras cervas promove a continuação da produção do seu leite.
2.4. Quantidade de leite
As produções de leite de fêmea de veado vermelho tem sido estudadas por alguns autores com especial destaque para as subespécies: ibérica, Cervus elaphus hispanicus e escocesa, Cervus elaphus scoticus.
Para a subespécie escocesa, Arman et al. (1974) obtiveram a média de 136,3 L de leite, estimada através da técnica da pesagem das crias, durante 150 dias de lactação de 4 cervas. No entanto, uma das cervas desse estudo foi mantida sob restrição alimentar, podendo assim considerar-se, por exclusão da sua produção, a média de 160 L. Este mesmo autor encontrou nessas fêmeas valores máximos de produção diária variados compreendidos em intervalos de 1,5 a 2 L por dia para fêmeas com curvas de tipo I e 1,2 a 2,5 L por dia para as fêmeas com curvas de tipo II. Loudon et al. (1983) estimaram para a mesma subespécie uma média de produção total de leite maior, de 171 L em 100 dias de lactação, encontrando máximos produtivos diários entre 1,4 e 1,6 L em curvas de tipo II atribuídas a animais mantidos em pastagens de baixa qualidade e de 2,2 L em curvas de tipo I de animais com acesso a alimento de qualidade. Os valores de produção de leite obtidos por García et al. (1999), na espécie Cervus elaphus hispanicus, através do método da pesagem das crias, foram de 1,16 L de leite por dia e
21
um total de 121,4 L durante 105 dias de lactação. Um pouco mais alta se encontra a produção de leite estimada por Landete-Castillejos et al. (2000b), nesta subespécie, que mediante ordenha manual das cervas durante 34 semanas, obtiveram 224,1 L de leite, o que corresponde a aproximadamente 147,0 L num período de lactação normal de 105 dias. Neste último estudo, a produção média de leite diminui continuamente ao longo da lactação, descrevendo uma curva de tipo II, desde um máximo de 2,10 mL na semana 2 para um mínimo de 264 mL na semana 34.
Num estudo comparativo das subespécies C. e. hispanicus e C. e. scoticus, Landete-Castillejos et al. (2003b) observaram que, como seria de esperar devido às diferenças morfológicas, existem também diferenças no que diz respeito à produção e composição do leite de ambas. Nesse trabalho obtiveram uma menor produção para as fêmeas de veado ibérico, de 207 L de leite, quando comparada com a produção total de 251 L, das fêmeas de veado escocês em lactações com a duração de 126 dias.
Sadleir (1980) encontrou para a subespécie de veado norte-americano, Odocoileus hemionus columbianus, através da estimativa de pesagem da cria, antes e após a amamentação, uma produção total de 93 L de leite em 120 dias de lactação com uma média de 0,8 L por dia e um máximo produtivo de 1,3 L. Na subespécie Cervus elaphus hippelaphus, foram encontradas por Krzywinski et al. (1980) produções médias diárias de leite estimadas por ordenha manual entre 1,20 e 2,85 L.
O valor mais alto de produção de leite referido na literatura para um cervídeo foi encontrado por Robbins (1981) na subespécie Cervus elaphus nelsoni, com o valor de 410 L em 150 dias de lactação, com uma média de 2,7 L por dia e uma produção excepcional de 4,1 L no pico de lactação.
Os resultados quanto à produção de leite podem diferir de acordo com o método utilizado para a sua estimativa, como foi explicado no ponto 2.1. Landete-Castillejos et al. (2000a) encontraram resultados similares de produção de leite segundo as estimativas através da técnica da pesagem e da ordenha manual em cervas ibéricas, de 193,1 e 193,6 L de uma lactação de 34 semanas, respectivamente. Contudo, nesse mesmo estudo, verificou também que através do método da pesagem não só existe uma maior variabilidade de resultados como também ocorre uma subestimação da produção de leite quando a amamentação da cria é exclusiva da sua progenitora (o que ocorre quando estas são isoladas do grupo), obtendo-se uma estimativa de produção de 143,9 L através deste método para 200,6 L obtidos na ordenha manual.
22
2.5. Composição do leite
O leite é uma secreção líquida complexa constituída por gorduras em emulsão, proteínas em dispersão coloidal e vários componentes orgânicos e inorgânicos em solução aquosa. Diferenças interespecíficas ocorrem não só nas proporções relativas dos principais nutrientes (água, gordura, proteína, açúcar e cinzas), mas também nos componentes específicos dentro de cada categoria (Robbins et al., 1987).
A composição média do leite é um conceito teórico, visto que esta difere de espécie para espécie e dentro da mesma espécie varia de forma natural ao longo da lactação, estando sujeita a numerosos factores tais como o potencial genético, a idade, o número de lactação, as relações mãe-cria, a alimentação, tipo de ordenha e muitos outros factores (Oldham e Friggens, 1989; Capuco e Akers, 2009).
O conhecimento dos componentes detalhados do leite é restrito a poucas espécies domésticas e aos seres humanos. Entre os ruminantes, os bovinos, ovinos e caprinos tem vindo a ser estudados, mas muito pouco se sabe ainda acerca da composição do leite de cervídeos.
Devido à sua elevada concentração e valor nutricional, o leite de cervídeos tem vindo a suscitar alguma curiosidade. É dentro da família dos cervídeos que se encontra a espécie animal que produz o leite mais concentrado de todos os ungulados, a rena, Rangifer tarandus (Geist, 1999; Gjøstein et al., 2004).
Robbins et al. (1987) caracterizaram o leite de cervídeos com tendo uma composição de 19-26% matéria seca, 6-11% de gordura, 6-10% de proteína, 3-5% de açúcar e 1,1 a 2,0% de cinzas. Dentro dos limites de variação de valores estabelecido por estes autores encontra-se a composição de leite de cervo ibérico, segundo Landete-Castillejos et al. (2000b), com 9,5% gordura, 6,7% proteína, 6,0% lactose e 24% extracto seco.
Em relação à concentração em minerais, os estudos publicados são escassos e mostram diferenças e variações notáveis entre eles. Gallego et al. (2006), num estudo do leite de 54 fêmeas de veado ibérico obtivera os seguintes resultados: Ca=2330 mg/ kg, P=640 mg/kg, K=1e 385 mg/kg, Mg=138 mg/kg, Zn=12.5 mg/kg e Fe=0.65 mg/kg.
23 2.5.1. Comparação com outras espécies
A composição média do leite de cerva apresenta diferenças notáveis no seu conteúdo em gordura e proteína, consideravelmente mais alto quando comparado com o conteúdo de leites de algumas das principais espécies animais produtoras de leite. Na tabela 2, encontram-se dados de natureza fragmentária, acerca da composição química do leite de cinco espécies cervídeos (alce, veado vermelho, veado Norte-Americano, veado de cauda branca e rena) em diferentes estados da lactação e, de forma a poder compará-la, a composição de 3 espécies de bovídeos: vaca doméstica, búfalo e ovelha. Assim, pode ver-se que leite da espécie Rangifer tarandus (rena) tem o maior teor em matéria seca quando comparado com as demais espécies e que, dentro das espécies domésticas apresentadas, é com o leite de ovelha que a concentração do leite de veado vermelho, C. elaphus, mais se aproxima. Contudo, pode também ver-se que existem diferenças quanto às composições do leite de veado vermelho obtidas pelos três autores citados. Devem-se ao facto de serem provenientes de diferentes subespécies ou de terem sido obtidas em diferentes fases da lactação, que influenciam estes valores como será discutido no ponto seguinte. Krzywinski et al. (1980) referiram que quando comparado com o leite de vaca, o leite de fêmea de veado vermelho pode conter o dobro de matéria seca, duas vezes mais proteína e 4 vezes mais gordura.
Tabela 2: Composição bruta de leites de cervídeos em diferentes estados da lactação e sua comparação com os de algumas espécies de bovídeos.
Espécies N Estado Matéria
Seca Gordura Proteína Açúcar Referências Amostras Lactação
Cervidae
Alces alces 15 >2 21.5 10.0 8,4 3,0 Ivanova,1965 20 4-90 24,5 5,8 10,3 - Franzmann et al.,1975 Cervus elaphus 6 3-31 21,1 8,5 7,1 4,5 Arman et al.,1974
6 5-77 22,1 9,4 6,2 4,9 Krzywinski et al.,1980 6 +101 27,1 13,1 8,6 4,5 Arman et al.,1974 Odocoileus hemionus 12 14-35 - 12,6 7,2 4,8 Mueller e Sadleir,1977 Odocoileus virginianus 4+ 21-28 22,5 7,7 8,2 4,6 Compilação1
Rangifer tarandus 6 21-30 26,3 10,9 9,5 3,4 Luhtala et al., 1968 Luick et al.,1974
Bovidae
Bos taurus 2000+ ‘Madura’ 12,4 3,7 3,2 4,6 Macy et al., 1953 Bubalis bubalis 42 30 16,8 6,5 4,3 4,9 Eltawil et al., 1976
Ovis aries 194+ 14-35 18,2 9,9 6,3 4,4 Oftedal, 1981
1
Murphy, 1960; Silver, 1961; Ben Shaul, 1962; Youatt et al., 1965; Jennesse Sloan 1970
24
2.6. Factores que afectam a produção e composição do leite
Em geral, os factores que afectam a lactação podem dividir-se por três grandes grupos (Oldham e Friggens,1989). O grupo no qual se incluem os factores que afectam o animal (factor animal), entre os quais se inclui a espécie, o indivíduo, o status reprodutivo e outros; o grupo no qual se encontram os factores de variação determinados pelo ambiente (factor ambiente), como a nutrição, necessidades energéticas, estado sanitário e frequência de ordenha e, aquele que está relacionado com o tempo (factor tempo), que agrupa a idade, número de lactação, estado da lactação e data de parto. A influência de cada um destes factores não ocorre de forma independente, estando todos altamente relacionados. Na tabela 3 encontram-se resumidas algumas das fontes de variação na performance da lactação classificados de acordo com os factores animal, ambiente e tempo.
Tabela 3: Fontes de variação na performance dos animais durante a lactação.
Factor animal Factor ambiente Factor tempo
Entre espécies Ambiente térmico Idade da progenitora
Entre raças dentro de espécies Nutrição Número de parto
Entre indivíduos Esforço físico Estação do ano
Genótipo Estado sanitário Estado da lactação
Status reprodutivo Frequência de ordenha
Tamanho da ninhada Agentes externos
Fonte: Adaptado a partir de Oldham e Friggens (1989)
2.6.1. Subespécies do animal
Quando comparadas as produções e composições dos leites das diferentes subespécies de veado vermelho, observam-se diferenças e portanto um importante efeito do factor subespécie nas características leiteiras destes animais.
Na tabela 4 encontram-se resumidos os valores encontrados por vários autores, para a composição de leite de diferentes subespécies de veado vermelho (C. e. scoticus, C. e. hippelaphus, C. e. nelsoni e C. e. hispanicus) segundo diferentes etapas de lactação, para algumas delas.
25
Tabela 4: Composição do leite de diferentes subespécies de veado vermelho (Cervus elaphus) segundo diferentes durações e etapas de lactação.
Espécie Período lactação1 Gordura Proteína Lactose E.S. Referência
C. e. scoticus <3 11,0 18,4 4,0 34,4 Arman et al., 1974 3-30 8,5 7,1 4,5 21,1
8-28* 9,4 9,2 4,9 22,1 31-100 10,3 7,6 4,5 23,5 +101 13,1 8,6 4,5 27,1
C. e. hippelaphus 5-77 9,4 6,2 4,9 22,1 Krzywinski et al., 1980
C. e. nelsoni - 90 7,5 6,2 4,1 19,8 Robbins et al., 1981
C. e. scoticus 0-116 9,5 - 12,82 7,2-8,12 — — Loudon, 1984
C. e. hispanicus 14-105 9,5 6,7 6,0 24 (Landete-C. et al., 2000b) 14-238 11,5 7,6 5,9 26,7
*Pico da lactação
1Dias depois do nascimento 2
Em função da qualidade da pastagem (má-boa)
Quando comparada a variação dos diferentes componentes do leite entre subespécies, podemos ver que o teor de lactose parece ser o menos variável e teor em gordura o que sofre a maior variação. Contudo estas variações dependem de outros factores e apenas existem dados acerca da composição do leite em diferentes estados de lactação para a subespécie escocesa, o que limita a tomada de conclusões.
Segundo Landete-castillejos et al. (2003b) podem também ver-se diferenças entre a quantidade de leite produzida e sua composição nas subespécies ibérica e escocesa, sendo que a segunda produz uma maior quantidade de leite (211 vs. 172 L) com uma maior concentração (gordura 17% vs. 14.3%; proteína 15,1% vs. 11% e lactose 12,4% vs. 10%), como se pode ver nos gráficos das figuras 7 e 8.
Quanto à composição mineral do leite, Vergara et al. (2003) não encontraram diferenças significativas entre as composições minerais dos leites de 6 cervas ibéricas e 6 cervas escocesas, obtendo uma composição média de 1717 mg/kg de Ca, 860 mg/kg de K, 109 mg/kg de Mg, 910 mg/kg de P, 109 mg/kg de Mn, 233 mg/kg Na de 109 e 9,06 mg/kg de Zn. Contudo, estes resultados são diferentes dos obtidos por Gallego et al. (2006) e já apresentados anteriormente no ponto 2.5.1.
26
Figura 7: Efeito da subespécie na curva de lactação de fêmea de veado vermelho. Comparação entre C. e. hispanicus e C. e. scoticus.
Fonte: Adaptado a partir de Landete-Castillejos et al. (2003b).
Figura 8: Diferenças entre a evolução da composição em gordura, proteína e proteína:gordura nos leites de cerva ibérica e cerva escocesa ao longo da lactação.