A problemática do cancro bacteriano da actinídea
na região de Entre Douro e Minho
Dissertação de Mestrado em Engenharia Agronómica
Adriana Sofia Pontes Vaz
Orientadora: Prof.ª Doutora Ana Maria Nazaré Pereira Co-orientadora: Prof.ª Doutora Maria Luísa Moura
A problemática do cancro bacteriano da actinídea
na região de Entre Douro e Minho
Dissertação de Mestrado em Engenharia Agronómica
Adriana Sofia Pontes Vaz
Orientadora: Prof.ª Doutora Ana Maria Nazaré Pereira Co-orientadora: Prof.ª Doutora Maria Luísa Moura
Composição do júri:
Presidente: Doutor Virgílio Alexandre Cardoso e Falco da Costa, Professor Auxiliar do Departamento de Agronomia da Universidade de Trás-os-Montes e Alto Douro.
Vogais: Doutora Maria Isabel Mendes Guerra Marques Cortez, Professora Auxiliar do Departamento de Agronomia da Universidade de Trás-os-Montes e Alto Douro;
Doutora Maria Luísa Roldão Marques Moura, Professora Adjunta da Escola Superior Agrária do Instituto Politécnico de Viana do Castelo.
“As doutrinas apresentadas neste trabalho são da exclusiva responsabilidade do autor.”
i dada a este trabalho, deixo aqui os meus sinceros agradecimentos:
Em primeiro lugar gostaria de agradecer aos meus pais, Carlos e Isidra, pois sem eles isto não seria possível, por toda a paciência e confiança, e por me ouvirem, apoiarem e incentivarem, mesmo com todo o cansaço que a profissão lhes exige. À minha avó, Augusta e irmã, Catarina, pela energia que lhes é tão característica por nunca me deixarem ir abaixo.
À Professora Doutora Ana Maria Pereira, por ter aceite ser minha orientadora e por toda a colaboração e incentivo dado durante este trabalho, cujas palavras tiveram um peso significante para que não desistisse à primeira dificuldade e para que este processo tenha chegado até aqui, proporcionando-me grandes oportunidades, as quais agradeço mais uma vez.
À Professora Doutora Luísa Moura, pela prontidão em ajudar e por me ter recebido na Escola Superior Agrária de Ponte de Lima de braços abertos. Por todos os seus ensinamentos e por ajudar a tentar solucionar todos os desafios que surgiam no laboratório e por todas as oportunidades que me proporcionou.
Ao Professor Doutor Miguel Brito, pela colaboração no tratamento estatístico dos dados.
À APK por todas as oportunidades que me foram proporcionadas e pela disponibilidade em ajudar e por permitirem que colaborasse e acompanhasse as prospecções por eles realizadas. A todos os seus colaboradores, em especial, ao Presidente da APK, Avelino Luís, ao Engenheiro Luís Reis e à Engenheira Alexandra Gomes, que de uma maneira ou de outra me fizeram sentir como se fosse parte integrante da Associação, proporcionando-me muitas das vezes uma perspetiva diferente dos assuntos.
À Escola Superior Agrária do Instituto Politécnico de Viana do Castelo, pelo uso das instalações laboratoriais para execução de todos os trabalhos. Ao Engenheiro Virgílio, por toda a paciência, ensinamentos e ajuda imprescindível na execução das tarefas laboratoriais. À D. Maria Rocha, D. Susy Marinho e D. Maria Carvalhosa por toda a ajuda no laboratório e pelos bons momentos proporcionados.
Ao Engenheiro Seabra, da DRAPN, por proporcionar o acesso aos dados meteorológicos e por toda a disponibilidade demonstrada.
ii À minha amiga, Carolina Varanda, por todos os devaneios, apoio e pelo facto de estar sempre presente, mesmo à distância de uma mensagem.
A todos os meus amigos, especialmente, à Andreia e ao Zeka, por todos os momentos de descontração que me proporcionaram e por estarem sempre presentes em todas as etapas.
iii (Psa) é uma doença que está incluída na Lista A2 da OEPP devido à sua gravidade crescente na região do Mediterrâneo. As condições meteorológicas determinam a multiplicação e o padrão de dispersão de Psa, bem como a relação entre a bactéria e as operações culturais no pomar, pouco estudadas, e que podem influenciar o controlo biológico a efetuar.
Os objetivos deste estudo são relacionar as condições meteorológicas com a eficácia da aplicação de Bacillus subtilis (Serenade Max ®) no controlo da doença, determinar se a severidade da doença aumentou ao longo do ciclo cultural, monitorizar a passagem da doença de um ciclo vegetativo da actinídea para o seguinte, e ainda avaliar o efeito da aplicação de Bacillus subtilis na produtividade e qualidade dos kiwis. Dois pomares de A. deliciosa cv. Hayward, com sintomas típicos de Psa e com diferentes idades, localizados em Valença, foram selecionados para ensaios de controlo biológico, produtividade e qualidade. Trinta plantas foram selecionadas de um pomar novo (6 anos) e outras trinta de um pomar velho (30 anos). Psa foi identificada e caracterizada por testes morfológicos, bioquímicos e moleculares. A eficácia de Bacillus subtilis foi avaliada através da incidência da doença, e correlacionada com as condições meteorológicas. Foi utilizada uma escala standard para a infecção na folha (escala de sintomas de 0 a 4) para determinação da severidade da doença. A monitorização dos sintomas da Psa foi executada a cada 30 dias, de junho de 2016 a setembro de 2017. Foram avaliados parâmetros produtivos e qualitativos da fruta como o número de frutos por planta, número de frutos por ramo lateral, peso por planta em kg, teor de sólidos solúveis totais, pH, firmeza da polpa, acidez e matéria seca.
A severidade da doença diminui no verão, caraterizado por condições pouco favoráveis para a Psa, como as elevadas temperaturas e escassa precipitação. Bacillus subtilis foi parcialmente eficaz no controlo da Psa no pomar novo (6 anos), e mostrou não ser eficaz na protecção das folhas no pomar velho (30 anos). A passagem da doença de um ciclo cultural para outro ocorreu, e a aplicação de Bacillus subtilis na floração considerou-se uma solução viável para o controlo de Psa biovar 3 apenas no pomar novo (6 anos).
O acompanhamento das prospecções oficiais em Portugal, permitiu concluir que o número de freguesias com Psa continua a aumentar, evidenciando uma dispersão mais lenta do que nos anos anteriores.
Palavras-chave: Actinidea deliciosa, cancro bacteriano, produtividade, qualidade condições
iv disease included in the EPPO A2 List because currently increased in the Mediterranean region. The climatic conditions determine the multiplication and dispersion behavior of Psa and the relationship between the bacteria and cultural practices, although poorly understood, may influence its biological control (BC). However, the literature regarding the growth of the bacteria with the climatic conditions in Portugal is scarce.
The aims of this study were to correlate the climatic conditions with the efficacy of Bacillus subtilis (Serenade Max ®) in disease control, to determine if disease incidence increased over the growing season, and to monitor carry-over of disease from one season to the next and also, to evaluate the effect of Bacillus subtilis (Serenade Max ®) on yield and kiwifruit quality in infected orchards with different ages. Two “Hayward” orchards of Actinidea deliciosa with typical symptoms of Psa and with different ages, located in Valença, North of Portugal, were selected for BC, yield and quality assays. Thirty plants were randomly selected from a young orchard (6 years old) and other thirty plants from an old one (30 years old). Psa
was identified and characterized by morphological, biochemical and molecular tests. The efficacy of Bacillus subtilis was evaluated by the disease incidence, and correlated to climatic conditions. A standardized leaf infection index (0 to 4 scale of symptoms) was used to determine the disease severity. The number of fruits per plant, number of fruits per lateral branch, kg per plant, ºBrix, pH, pulp hardness, titratable acidity and dry matter obtained in plants of both orchards are discussed. Monitoring of Psa symptoms was performed every 30 days, from June 2016 to April 2017. The severity of the disease decreased in the summer season, characterized by unfavorable conditions for Psa such as high temperatures and scarce precipitation. Bacillus subtilis was partially effective in controlling Psa in the six-year-old orchard, and showed no efficacy in the protection of leaves in the thirty-year-six-year-old orchard. The disease carry-on from one season to the next, and the Bacillus subtilis applied at flowering could be considered as a suitable solution for the control of Psa biovar 3 only for the new kiwifruit orchard (6 years old).
The follow-up of the official surveys in Portugal allowed us to conclude that the number of orchards with Psa continues to increase, showing a slower dispersion than in previous years.
Keywords: Actinidea deliciosa, bacterial canker, kiwifruit yield, kiwifruit quality, climatic
v
Resumo ... iii
Abstract ... iv
Índice de figuras ... vii
Índice de quadros ... ix
Lista de abreviaturas ... x
Revisão Bibliográfica ... 1
1. A cultura da actinídea ...1
2. O cancro bacteriano da actinídea ...8
2.1. História ... 8
2.2. Agente Causal ... 9
2.3. Sintomatologia ... 10
2.4. Disseminação ... 11
2.5. Meios de protecção... 13
2.5.1. Meios de protecção cultural ... 13
2.5.2. Meios de protecção biológica ... 13
2.5.2.1. Bacteriófagos ... 13
2.5.2.2. Bacillus subtilis ... 14
2.5.2.3. Leveduras ... 15
2.5.3. Meios de protecção química ... 16
2.5.3.1. Cobre ... 16
2.5.3.2. Estreptomicina ... 17
2.5.3.3. Terpenos ... 17
2.5.3.4. Quitosana ... 18
2.5.3.5. Péptidos antimicrobianos ... 18
2.5.3.6. Indutores das defesas das plantas ... 19
Material e Métodos ... 22
1. Acompanhamento das prospecções oficiais de Psa em pomares na Região de Entre Douro e Minho nos anos de 2016 e 2017 ... 22
2. Estudo do cancro bacteriano da actinídea num pomar na Região do Entre Douro e Minho. ... 23
2.1. Caraterização do pomar ... 23
2.2. Parâmetros meteorológicos registados no período de maio de 2016 a setembro de 2017 ... 24
vi
2.5. Avaliação da produção e qualidade dos frutos em 2016 e 2017 ... 26
3. Isolamento e identificação de Pseudomonas syringae pv. actinidiae (Psa) ... 28
4. Caraterização morfológica e bioquímica das estirpes obtidas ... 29
5. Identificação e Caraterização molecular das estirpes obtidas ... 30
5.1. Preparação de culturas bacterianas e extracção de ADN ... 30
5.2. Amplificação por Duplex-PCR utilizando os primers KN-F/KN-R e AvrDdpx-F/AvrDdpx-R ... 30
5.3. Caraterização molecular por BOX-PCR ... 31
5.4. Electroforese de fragmentos de ADN obtidos por Duplex-PCR e BOX-PCR ... 31
Resultados e Discussão ... 33
1. Acompanhamento das prospecções oficiais de Psa em pomares na Região de Entre Douro e Minho nos anos de 2016 e 2017 ... 33
2. Estudo de cancro bacteriano da actinídea num pomar da região do Entre Douro e Minho. ... 36
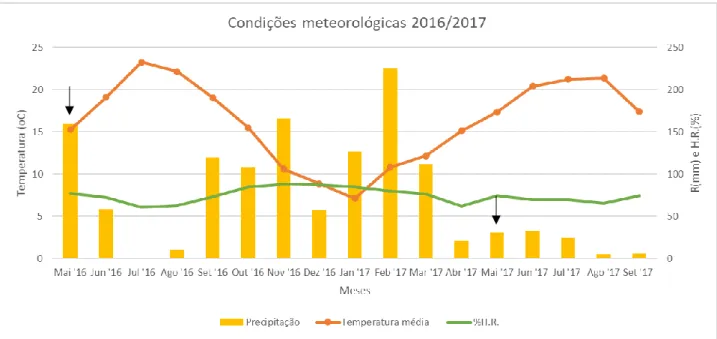
2.1. Parâmetros meteorológicos obtidos no ano de 2016 e 2017... 36
2.2. Evolução sintomatológica da doença no pomar durante 2016 e 2017... 37
2.3. Aplicação de um produto comercial à base de Bacillus subtilis (Serenade Max®).. ... 39
2.4. Avaliação da produção e qualidade dos frutos em 2016 e 2017... 44
3. Isolamento de uma colecção de isolados de Pseudomonas syringae pv. actinidiae (Psa) ... 48
4. Caraterização bioquímica das estirpes obtidas ... 49
5. Identificação e caraterização molecular das estirpes de Psa ... 52
5.1. Identificação por Duplex-PCR utilizando os primers KN-F/KN-R e avrdDpx-F/R 52 5.2. Identificação e caraterização por BOX-PCR ... 53
Conclusões ... 55
Referências Bibliográficas ... 57 ANEXOS ... lxiii
vii Figura 2. Produção mundial de kiwi entre 1977 a 2014. Fonte: FAOSTAT, 2015 ... 4 Figura 3. Produção de kiwi em Portugal desde 2012 até 2016. Fonte: INE, 2017 ... 5 Figura 4. Presença de uma cigarrinha na folha da actinídea. Foto: do autor. ... 7 Figura 5. Mapa de distribuição de Pseudomonas syringae pv. actinidiae em julho de 2017. Fonte: https://gd.eppo.int/taxon/PSDMAK/distribution ... 9 Figura 6. Ciclo de vida de Pseudomonas syringae pv. actinidiae, adaptado de Kim et al., 2017 ... 12 Figura 7. Localização do pomar no mapa de Portugal através de vista aérea. ... 23 Figura 8. Escala de severidade utilizada para avaliar os sintomas no pomar ... 26 Figura 9. Mapa das prospecções do ano de 2016 com os concelhos reconhecidos como área não livre de Psa (azul) e as freguesias positivas para a presença de Psa (vermelho) (adaptado de DRAPN, 2016). ... 34 Figura 10. Condições meteorológicas relativas ao ano de 2016 e 2017. Estação
meteorológica de Ganfei. Aplicações de Serenade Max ®. ... 36 Figura 11. Diferentes sintomas presentes nos botões florais e nas flores observados no pomar. Valença do Minho, maio 2016 e 2017. Fotos: do autor ... 37 Figura 12. Sintomas típicos nas folhas (A, B, D - pontuações necróticas nas folhas; C, F - necroses mais extensas nas folhas; E- pormenor das pontuações necróticas; G- pormenor das necroses nas nervuras da folha. Valença do Minho, junho, julho e agosto de 2016 e 2017. Fotos: do autor ... 38 Figura 13. Imagens dos diferentes exsudados observados no pomar. Valença do Minho, fevereiro e março de 2017. Fotos: do autor ... 38 Figura 14. Sintomas em primórdios florais e até mesmo em rebentos jovens. Valença do Minho, março de 2017. Fotos: do autor ... 39 Figura 15. Valor médio de necroses nas folhas nos dois pomares (A) e nas duas aplicações (B). Letras diferentes indicam diferenças significativas entre os tratamentos (p<0,05). ... 41 Figura 16. Efeito da interação entre a idade do pomar e a aplicação de Serenade Max® no nº de ramos laterais por planta (A) e na produção total por planta (B) em 2016. Letras
diferentes indicam diferenças significativas entre os tratamentos (p<0,05). ... 44 Figura 17. Efeito da interação entre a idade do pomar e a aplicação de Serenade Max® no nº de varas por planta (C) e no nº de ramos laterais por planta (D) em 2017. Letras
viii tratamentos (p <0,05)... 46 Figura 19. Características típicas das colónias de Pseudomonas syringae pv. actinidiae. Fotos: do autor. ... 49 Figura 20. Diferenças nos pigmentos fluorescentes nas diferentes estirpes isoladas. Placa da esquerda pigmentos não fluorescentes; Placa da direita com pigmentos fluorescentes. Foto: do autor. ... 49 Figura 21. Dendrograma das distâncias fenotípicas entre 19 estirpes testadas com
microplacas GEN III (Biolog). ... 50 Figura 22. Aspeto de uma microplaca GEN III Biolog já utilizada. Foto: do autor. ... 51 Figura 23. Fragmentos de amplificação obtidos com os primers KN-F/R e AvrDdpx-F/R numa reação de Duplex- PCR, obtidos com as estirpes de P.s. pv. actinidiae. Colunas 1: controlo negativo; 2: CFBP 7286; 3 a 9: Estirpes Portuguesas. M: Marcador 100 bp ADN Ladder (Bioron) ... 52 Figura 24. Perfis electroforéticos obtidos pela amplificação de fragmentos de ADN de
isolados de P.s. pv. actinidiae (Psa), através de PCR utilizando o primer BOXA1R. Coluna 1: CFBP 7286; 2-P84; 3 a 7: estirpes EDM 2016/17;8: H2O (controlo negativo); M1: 100 bp DNA Ladder (Bioron)M2: 1kb DNA Ladder (Bioron); ... 54
ix anos de 2015 e 2016. Fonte: INE, 2016; INE,2017 ... 6 Quadro 2. Produtos fitofarmacêuticos autorizados para combate/controlo da Psa em
Portugal (Associação Portuguesa de Kiwicultores, 2017b). ... 20 Quadro 3. Calendarização das operações culturais realizadas no pomar em 2016 e 2017. . 24 Quadro 4. Origem das estirpes de Psa isoladas noutros países e noutros estudos. ... 30 Quadro 5. Características dos pomares visitados para efeitos de prospeção nos anos de 2016 (Pomar A até I) e 2017 (Pomar J, K e L). ... 35 Quadro 6. Índice médio da doença para os 2 factores principais em 2016 e 2017. ... 42 Quadro 7. Efeito da idade do pomar e da aplicação de Serenade Max® nos parâmetros produtivos em 2016. ... 45 Quadro 8. Efeito da idade do pomar e da aplicação de Serenade Max® no parâmetro do número de frutos por ramo lateral para os dois pomares e para os dois tratamentos para o ano de 2017. ... 46 Quadro 9. Parâmetros qualitativos do fruto avaliados em 2016. ... 47 Quadro 10. Estirpes isoladas em 2016 e 2017 de plantas infectadas cv. Hayward. ... 48 Quadro 11. Resultados das amplificações obtidas por Duplex-PCR utilizando os primers KN-F/KN-R e AvrDdpx-F/AvrDdpx-R e identificação das estirpes estudadas. ... 53
x FAOSTAT – Food and Agricultural Organization – Statistics division
FAO – Food and Agricultural Organization APK – Associação Portuguesa de Kiwicultores
GPP – Gabinete de Planeamento, Políticas e Administração Geral INE – Instituto Nacional de Estatística
EDM – Região do Entre Douro e Minho KVH – Kiwi Vine Health
Psa – Pseudomonas syringae pv. actinidiae
EPPO/OEPP – European Plant Protection Organization/Organização Europeia para a Protecção das Plantas
DGAV – Direcção Geral de Agricultura e Veterinária UV – ultravioleta
CFBP – Colecção Francesa de bactérias fitopatogénicas ADN – ácido desoxirribonucleico
ASM – acibenzolar-S-metil
DRAP – Direcção Regional da Agricultura e Pescas
INIAV – Instituto Nacional de Investigação Agrária e Veterinária IPVC – Instituto Politécnico de Viana do Castelo
PCR – Reação em cadeia de polimerase (“polymerase chain reaction”) kb – kilobases
1
Revisão Bibliográfica
1. A cultura da actinídea
O género Actinidia pertence à família Actinidiaceae e estão identificadas perto de 55 espécies e 20 variedades das quais Actinidia deliciosa (A. Chev.) e A. chinensis (Planch.) têm maior importância comercial, para além de Actinidia arguta cuja importância tem vindo a aumentar nos últimos anos (Ferguson, 2013).
Apesar do género Actinidia ter sido trazido, mais tarde, para a Europa e Estados Unidos da América e sendo a sua origem e presença maioritária nas montanhas da China Central e do Sul (Ferguson, 2013), as primeiras produções comerciais foram obtidas na Nova Zelândia nos anos 30. Por esta razão, a Nova Zelândia iniciou o comércio externo de kiwi e também de plantas, suplementando assim países como a Itália, França e Grécia de variedades comercialmente interessantes (Hayward, Bruno, Abbot e Monty) permitindo que nos anos 70 a maior parte desses países tivessem plantas de Actinidea para a instalação de pomares (Ward & Courtney, 2013) aumentando assim a produção e o comércio internacional na fileira do kiwi.
Na China, de onde é originária a Actinidia, esta pode ser apelidada de várias maneiras, sendo que um dos nomes mais utilizados no início da dinastia de Tang foi mihoutao. Na região do rio Yangtzé, onde esta planta é mais abundante, a expressão mais utilizada para caracterizar Actinidea é yangtao. Na Nova Zelândia o nome mais utilizado é groselha chinesa ou “chinese gooseberry” em inglês, alusivo à origem da planta e ao aspecto que possui (Ferguson, 2013).
As plantas do género Actinidia são perenes de folha caduca e com hábito de crescimento trepador (Ferguson, 2013). Relativamente a outras plantas trepadoras, no género Actinidia as diferentes espécies têm comportamento muito variável entre si em termos de estrutura vegetativa (Ferguson, 2013).
Actinídea sendo uma planta dióica, tem flores femininas apenas em plantas fêmeas e flores masculinas em plantas macho, apresentando diferenças bastante visíveis (Figura 1).
2 Os frutos são uma baga de formato oblongo, dependendo o seu comprimento da cultivar a que pertencem. A epiderme do fruto varia consoante a espécie, e no caso de A. deliciosa é de cor castanha, estando coberta, em algumas cultivares, por uma camada de tricodermas densos. Nesta espécie a polpa é de cor verde, com uma columela de cor branca, aspectos característicos da cv. Hayward. Podem ser observadas também algumas fiadas de sementes de cor negra na maior parte das cultivares de A. deliciosa e A. chinensis (Associação Portuguesa de Kiwicultores, 2007).
As plantas de Actinidea têm uma morfologia que pode ser dividida em duas partes, as estruturas perenes e as estruturas produtivas. As estruturas perenes são estruturas que são formadas e definidas desde a sua condução inicial, ou seja, o tronco e os braços da planta. Nas plantas mais jovens, é necessário utilizar um tutor, durante os primeiros tempos, para condução da planta na vertical. Ao longo do tempo, as estruturas perenes vão ganhando lenhosidade permitindo assim prescindir do tutor e formar a planta definitiva (Associação Portuguesa de Kiwicultores, 2007).
Quanto às estruturas produtivas, estas podem ainda ser divididas em lançamentos do ano e laterais produtivos. Os primeiros, provêm de gomos existentes nos lançamentos do ano anterior. Estes lançamentos do ano podem adquirir dois tipos de crescimento, um crescimento determinado, em que o meristema apical não se desenvolve quando o lançamento já possui entre 5 a 10 entrenós, ou então um crescimento indeterminado, em que o meristema apical está sempre em desenvolvimento, podendo chegar a um comprimento de 10 metros (Associação Portuguesa de Kiwicultores, 2007).
Quanto aos ramos laterais produtivos ou varas, podem ter duas origens a partir de lançamentos indeterminados ou determinados. As que provêm de lançamentos indeterminados possuem 10 a 20 entrenós, sendo longos, tendo a capacidade para originar rebentos laterais frutíferos ou rebentos laterais vegetativos. As varas originárias de Figura 1. Flor feminina (esquerda) e flor masculina (direita). Foto: do autor.
3 lançamentos determinados, têm entrenós muitos curtos com elevada densidade de gomos, do tipo esporão e com uma capacidade produtiva elevada (Associação Portuguesa de Kiwicultores, 2007).
Todas estas estruturas produtivas necessitam de manutenção ao longo do ciclo vegetativo, o que leva a que o produtor tenha que integrar na gestão do pomar algumas das operações culturais tais como, a poda de inverno, a empa e atado, a monda de frutos e a poda em verde, necessitando de muita mão-de-obra e tempo, o que muitas das vezes é ignorado ou desvalorizado por parte do produtor, comprometendo por vezes a produção final e a qualidade do fruto.
A poda de inverno é efectuada entre o final do mês de dezembro e o início do mês de fevereiro, fase em que a planta se encontra em repouso vegetativo, e tem como objetivo definir a arquitectura da planta, tendo sempre presente o equilíbrio entre a parte vegetativa e produtiva da planta, para que o arejamento e a iluminação não sejam comprometidos pelas posições das varas. Nesta poda, são eliminadas também todas as varas sem potencial produtivo, muito vigorosas, com entrenós muito longos, permitindo assim que a distribuição de varas seja mais equilibrada (Associação Portuguesa de Kiwicultores, 2017).
A empa e o atado são operações que se podem considerar parte integrante da poda de inverno. A empa permite alterar o comportamento vegetativo das varas, permitindo dispô-las de forma ordenada e fixando-as através do atado. Estas operações são fundamentais para manter uma boa distribuição das varas permitindo a entrada de luz da planta, favorecendo a diferenciação dos gomos, evitando a sobreposição das varas, já que estas são atadas aos arames laterais (Associação Portuguesa de Kiwicultores, 2017).
A monda de frutos exige bastante mão de obra e efectua-se entre meados de junho e finais de julho (seis semanas a seguir ao vingamento), e tem como objetivo melhorar a qualidade dos frutos eliminando os frutos deformados, com defeitos de epiderme, pequenos e em excesso (Associação Portuguesa de Kiwicultores, 2007).
A poda em verde permite promover o arejamento e entrada de luz nas plantas, controlar o vigor vegetativo e melhorar as condições do pomar durante a fase de floração e crescimento do fruto. Nesta operação são retirados todos os ramos laterais que não têm fruto, ou seja, são infrutíferos e que estão a consumir assimilados e reservas que são fundamentais para os frutos (Associação Portuguesa de Kiwicultores, 2007).
4 Estas operações culturais são algumas das principais a efetuar num pomar, no entanto podem variar em termos de adaptação consoante o modo de condução do pomar e cultivar que esteja instalada.
Por volta dos anos 40 (Ferguson, 2013), a espécie A. deliciosa, foi a mais atrativa para produção comercial de kiwi, sendo a cultivar Hayward, de polpa verde, a mais utlizada na implementação de pomares e uma das principais a iniciar o comércio de kiwis. Posteriormente em 1997, a empresa Zespri® lançou uma variedade de polpa amarela (Actinidia chinensis, cv. Hort16A – Zespri Gold®), que veio revolucionar todo o mercado da cultura (Associação Portuguesa de Kiwicultores, 2007).
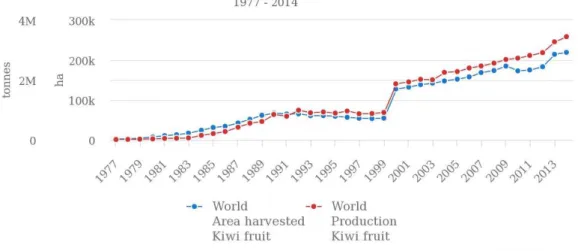
Os maiores produtores de kiwi são a China (1 227 599 toneladas), Itália (383 023 toneladas), Nova Zelândia (310 912 toneladas) e o Chile (174 346 toneladas) (FAOSTAT, 2015).
Segundo os dados estatísticos da FAO (Ward & Courtney, 2013), a produção global de Actinidea tem vindo a aumentar exponencialmente nos últimos anos (Figura 2).
A liberalização do mercado fez com que a rentabilidade da cultura tenha diminuído, dando origem a uma crise mundial do sector nos primeiros anos da década de 90 (Figura 2). Verificou-se assim uma redução da superfície da área plantada, pela falta de rentabilidade dos pomares e também por erros cometidos na plantação o que não permitiu ultrapassar a crise que se fazia sentir (Associação Portuguesa de Kiwicultores, 2007). No entanto, no ano 2000, o sector conseguiu recuperar da crise, tanto a nível mundial, como a nível nacional, e verificou-se um aumento na área de Actinidea, devido principalmente ao surgimento de novos pomares com condições ideais para uma máxima rentabilização da cultura.
5 Em Portugal Actinidea foi introduzida no início da década de 70, sendo o primeiro pomar instalado em Vila Nova de Gaia em 1973 e as regiões do Entre Douro e Minho e Beira Litoral as principais na produção de kiwis (APK, 2007). A área total de Actinidea em 2015 era de 2 305 ha, com uma produção total de 28 331 toneladas (Instituto Nacional de Estatística, 2016). A maior área de produção localiza-se na região de Entre Douro e Minho, que em 2015 totalizava 1721 ha, e uma produção de 23 206 toneladas (Instituto Nacional de Estatística, 2016).
Em termos de exportação, o kiwi é a 4ª fruta mais exportada, abaixo de frutas como a pera rocha, a framboesa e a laranja, com valores de faturação a rondar os 9,26 milhões de euros, equivalendo a 14 087 toneladas exportadas (GPP, 2015). Os principais destinos de exportação são a Espanha, Alemanha e França (GPP, 2015).
Entre janeiro e abril de 2017, houve um aumento nas exportações de 61% e um aumento nas importações de cerca de 21%, o que levou a que o valor faturado aumentasse cerca de 96% (Associação Portuguesa de Kiwicultores, 2017b). Em termos de balança comercial, esta é positiva, no entanto, passamos para valores de 7,18 milhões de euros, o que posiciona o kiwi em 3º lugar, ultrapassando apenas a laranja.
Em 2016 a produção desceu relativamente a 2015 (Figura 3), devido principalmente ao reduzido número de horas de frio que se fez sentir no Inverno e que não promoveu a diferenciação floral dos gomos, devido à deficiente polinização e à forte presença da doença do cancro bacteriano nos pomares, causado por Pseudomonas syringae pv. actinidiae (Psa), tendo-se contabilizado uma redução de cerca de 27,9% na produção (Figura 3) (Quadro 1).
6 Apesar da produção ter diminuído a área plantada aumentou devido à instalação de novos pomares, permitindo que a região de Entre Douro e Minho continuasse na liderança tanto em termos de área como em termos de produção. No ano de 2016, esta região alcançou 1 792 ha de área plantada e 16 918 toneladas de produção (INE, 2017).
Quadro 1. Valores de área e produção para a região do Entre Douro e Minho e Portugal nos anos de 2015 e 2016. Fonte: INE, 2016; INE,2017
EDM Portugal
Área (ha) Produção (ton) Área (ha) Produção (ton)
2015 1 721 23 205 2 305 28 331
2016 1 792 16 918 2 380 21 075
Estes dados posicionam a fileira do kiwi num ponto de elevada importância económica, o que significa que o valor que é atribuído ao kiwi nacional é muito superior ao kiwi importado e que a fileira pode vir a crescer ainda mais pela qualidade demonstrada. No entanto, apesar da produção por hectare estar a crescer, temos que ter em atenção que ainda não foram atingidas as médias de produção dos concorrentes europeus e, portanto, a competitividade da fileira é colocada em causa (Associação Portuguesa de Kiwicultores, 2017b).
O relativo curto período de tempo entre o estado selvagem e a domesticação e cultivo da Actinidia, e o alargamento da cultura a outros países, levou progressivamente ao aparecimento de doenças comuns a outras fruteiras. As principais doenças que afetam Actinidea, principalmente na região do Entre Douro e Minho e Beira Litoral são o cancro bacteriano, causado por Pseudomonas syringae pv. actinidiae (Psa), e que devido à sua agressividade é a doença que maiores prejuízos causa, a esca (complexo de fungos) que infetam principalmente as partes lenhosas da planta, Armillaria spp. e Phythophtora spp. que provocam podridões nas raízes (Associação Portuguesa de Kiwicultores, 2007). A podridão cinzenta, Botrytis cinerea, apresenta igualmente elevada importância, mas menos preocupante, sendo um fungo causador podridões nos frutos na pós-colheita. Actinidea é também atacada por pragas comuns a outras culturas, e que dependendo do país e região onde é cultivada o risco de incidência dessa praga varia (Hongwen, 2016). Em Portugal, existem várias pragas que atacam Actinidea, mas que têm um nível de ataque baixo (Associação Portuguesa de Kiwicultores, 2007), como é exemplo os nemátodes, cochonilhas e cigarrinhas (Figura 4).
7 Noutros países produtores de Actinidea surgem outras doenças causadas por bactérias, fungos ou até mesmo vírus que provocam prejuízos, mas que não têm importância em Portugal. Existem ainda referências a nemátodos que podem provocar também estragos na cultura com alguma importância económica dependendo da extensão com que infetam a cultura (Hongwen, 2016). Mais recentemente foi descoberto um fungo que está a provocar a morte de vários pomares no Brasil, Ceratocystis fimbriata, e também um escaravelho, que está a provocar perdas de 30% da produção em Itália, denominado, Halyomorpha halys (KVH, 2016a).
8
2. O cancro bacteriano da actinídea
2.6. História
A doença do cancro bacteriano da actinídea, é causada pela bactéria Pseudomonas syringae pv. actinidiae (Psa), afetando as diferentes espécies de Actinidea, incluindo A. deliciosa, A. chinensis, A. arguta e A. kolomikta. É uma bactéria fitopatogénica muito destrutiva, causando perdas económicas severas em todas as principais áreas de produção de kiwi no mundo (Scortichini et al., 2012; Vanneste, 2013; Fujikawa & Sawada, 2016). Este patogéneo foi isolado, identificado e descrito pela primeira vez no Japão em 1989 (Takikawa et al., 1989), onde causou danos em pomares de Actinidea.
Na Europa a doença foi primeiramente notificada na Itália em 1992, onde se manteve esporadicamente e com baixa incidência durante 15 anos (EPPO, 2009). No entanto, desde 2007/2008 a doença foi observada com gravidade no Norte de Itália (Lazio), e casos esporádicos foram reportados na região de Emilia-Romagna (outubro 2009), sendo a presença da bactéria confirmada em A. chinensis cv. ‘Hort 16A’. Prospeções realizadas em 2007/2008 no Noroeste de Itália mostraram que Psa ocorria em vários pomares das províncias de Latina e Roma (região de Lazio) e na província de Treviso (região de Veneto). A incidência da doença variou entre 50% e 80%, e em alguns casos com elevada agressividade, tendo levado à destruição total dos pomares. A maior severidade da doença estava associada à espécie A. chinensis, em particular à cultivar de polpa amarela ‘Hort 16A’ nas províncias de Latina e Roma. Na província de Treviso na cv. ‘Jin Tao’ (A. chinensis) a doença manifestou-se com menor incidência e manifestou-severidade, pois os sintomas afetaram maioritariamente as folhas, gomos e flores (EPPO, 2009).
Mais tarde a doença foi declarada noutros países da Europa como Portugal (Balestra et al., 2010), França (Vanneste et al., 2011), Espanha (Abelleira et al., 2011), Suíça (EPPO, 2011), Turquia (Bastas & Karakaya, 2012), Alemanha (EPPO, 2013). Fora da região da OEPP (Organização Europeia para a Protecção das Plantas), a doença foi reportada na Nova Zelândia (Everett et al., 2011), Chile (EPPO, 2011a), Austrália (EPPO, 2011a), Coreia (Koh et al., 2012) e China (EPPO, 2014). A distribuição atual de Psa é apresentada na Figura 5.
Em Portugal, a doença foi detectada apenas em Março de 2010 (Balestra et al., 2010) em plantas de A. deliciosa na região do Entre Douro e Minho, mas rapidamente se espalhou pelas regiões produtoras de kiwi em Portugal (Anexo 1). A doença manifesta-se quando as condições climáticas são mais favoráveis ao seu desenvolvimento, com temperaturas entre 10-20 ºC (limitada com temperaturas superiores a 25 ºC) e alta humidade.
9
2.6. Agente Causal
A bactéria que causa o cancro bacteriano da actinídea é designada por Pseudomonas syringae pv. actinidiae, sendo uma Proteobacteria, da subdivisão Gamma, da ordem Pseudomonadales e da família Pseudomonadaceae (EPPO, 2014). Esta bactéria é gram-negativa, aeróbica, móvel com formato oval e com flagelo polar, oxidase gram-negativa, dihidrolase da arginina negativa, pectina negativa, levana positiva e hipersensibilidade em folhas de tabaco positiva (Abelleira et al., 2014).
Com base nos estudos realizados, foram isoladas um elevado número de estirpes em diversos países, compreendeu-se que as estirpes de Psa que provocaram o surto em Itália no ano de 2008, no Chile, na Nova Zelândia e na China, fazem parte de uma única linhagem de estirpes, muito idênticas entre si e que diferem daquelas que foram isoladas no Japão, Coreia e Itália em 1992 (Ferrante & Scortichini, 2010).
Foram descritas seis populações geneticamente diferentes de Psa, designadas: Psa1-4 (Chapman et al., 2012) ou biovares 1-Psa1-4 segundo Vanneste et al., (2013), biovares 5 e 6 segundo Sawada et al., (2014) e Sawada et al., (2016), respectivamente. As biovares 1, 2 e 3 são consideradas virulentas, e a biovar 4 pouco virulenta (Vanneste et al., 2012). A biovar 5 está proximamente relacionada com a biovar 2 (Cameron & Sarojini, 2014; Fujikawa & Sawada, 2016). Apesar da distribuição da biovar 6 estar extremamente limitada no Japão, atualmente, pode ter um importante papel na compreensão da origem e evolução das populações de Psa (Sawada et al., 2016).
As populações virulentas podem ser diferenciadas de várias formas, incluindo a presença ou ausência de fitotoxinas. Assim, as estirpes da biovar 1 incluem estirpes do
Figura 5. Mapa de distribuição de Pseudomonas syringae pv. actinidiae em julho de 2017. Fonte: https://gd.eppo.int/taxon/PSDMAK/distribution
10 primeiro registo da doença no Japão e em Itália antes de 2008 e têm a capacidade de produzir faseolotoxina (Chapman et al., 2012); a biovar 2, isolada na Coreia do Sul durante a epidemia dos anos 90, produz coronatina mas não faseolotoxina (Han et al., 2003); a biovar 3 inclui as estirpes isoladas na Europa, Nova Zelândia, Chile e China, associadas à pandemia de 2010, e não produzem faseolotoxina nem coronatina (Ferrante & Scortichini, 2010); a biovar 4 foi definida como um grupo particular de estirpes apresentando baixa virulência em Actinidea, causando manchas foliares mas não cancros nos ramos e troncos da planta, tendo sido isoladas primeiramente na Nova Zelândia e Austrália (Vanneste et al., 2013) e mais recentemente em França e Espanha (Cunty et al., 2015; Abelleira et al., 2015). A biovar 5 foi isolada apenas numa área local (Saga Prefecture) no Japão (Sawada et al., 2014) e está proximamente relacionada com a biovar 2, apesar de não ter capacidade de produzir a toxina coronatina (Fujikawa & Sawada, 2016). Finalmente, a biovar 6 de Psa foi relatada numa pequena região do Japão, e as estirpes produzem as duas fitotoxinas, faseolotoxina e coronatina (Sawada et al., 2016).
Após a realização de vários estudos que mostraram que a biovar 4 de Psa não causava prejuízos em plantas de Actinidia, e que eram claramente diferentes das populações virulentas de Psa (Cunty et al., 2015; Abelleira et al., 2015; Ferrante & Scortichini 2015), foram reclassificadas como uma nova patovar designada Pseudomonas syringae pv. actinidifoliorum (Cunty et al., 2015).
Em Portugal existem estudos sobre as biovares presentes nomeadamente de Garcia (2015), que caracteriza a variabilidade genética e fenotípica da Psa no Entre Douro e Minho e de Moura et al., (2015) que caracterizam também as biovares presentes na mesma região, indicando que nesta região apenas está presente a mais virulenta e destrutiva, a biovar 3.
2.6. Sintomatologia
Os sintomas do cancro bacteriano da actinídea podem ser observados em troncos, ramos, folhas, flores e frutos. Estes sintomas incluem pontuações necróticas de coloração acastanhada rodeadas por halo clorótico nas folhas, encontradas principalmente nas folhas mais jovens, podendo ser observadas no inicio da primavera e verão associadas à murchidão das folhas, que apenas ocorre quando a bactéria se movimenta sistemicamente dentro dos tecidos da planta provocando o bloqueamento dos tecidos vasculares (Vanneste et al., 2011; Hongwen, 2016). Muitas vezes essas pontuações necróticas de pequenas dimensões acabam por se agrupar e existe a formação de necroses com dimensões grandes, ocupando a totalidade da área foliar (Moura, 2013). Nos troncos e caules pode-se observar também alguma coloração castanha-avermelhada dos tecidos vasculares que se encontram por baixo
11 do córtex (EPPO, 2014) ou o aparecimento de exsudado avermelhado ou por vezes esbranquiçado nos troncos e ramos (Hongwen, 2016), sintoma característico de Psa.
Outros sintomas que são também observados nas plantas dos pomares mais afectados são as necroses visíveis nos botões florais, provocando a sua queda, levando à necrose completa e murchidão total, ou então o abortamento floral, o que impede a formação do fruto (Vanneste et al., 2011; Abelleira et al., 2011).
O sintoma de murchidão observado nos lançamentos mais jovens, pode numa fase mais avançada provocar uma coloração negra dos tecidos ficando os lançamentos espalmados, sintoma visível principalmente na primavera (Donati et al., 2014). No inverno, os sintomas apenas são visíveis no fim da estação formando-se cancros nos troncos, dos quais se libertam exsudados avermelhados ou esbranquiçados que podem constituir uma grande fonte de inóculo da bactéria (Everett et al., 2011).
As pontuações necróticas e as necroses que ocorrem nas flores podem ser confundidos com outros sintomas de bactérias patogénicas que também podem atacar Actinidea como Pseudomonas syringae pv. syringae ou Pseudomonas viridiflava, o que dificulta em muito a sua identificação em campo (Vanneste et al., 2011).
2.6. Disseminação
A disseminação da doença a pequenas distâncias pode ocorrer através da produção de exsudados nas plantas infectadas e, tendo estas uma elevada carga microbiana proliferam com bastante facilidade, pelo contacto directo entre plantas sãs e plantas infetadas, pelo vento, sendo principalmente as populações epifíticas as que são mais disseminadas por se encontrarem no exterior da planta (Vanneste et al., 2011), através da chuva que remove mais facilmente as bactérias das folhas e exsudados, pelo pólen e também através de alguns insectos, nomeadamente abelhas (Stefani & Giovanardi, 2011; Ferrante et al., 2012).
Stefani e Giovanardi (2011) defendem ainda que a humidade relativa, a ocorrência de geadas e a própria idade da planta podem ser fatores fundamentais para a disseminação da doença.
A disseminação por via humana, também é possível através dos instrumentos de poda, veículos ou maquinaria, animais e solo, podendo ser disseminada em curtas ou longas distâncias (Everett et al., 2012). A disseminação a longas distâncias pode ocorrer através de material de propagação vegetativa ou plantas infetadas, ou através do pólen infetado, proveniente de regiões muito distantes daquela onde vai ser utilizado. Em 2011, na Nova
0
12 Zelândia foram testadas amostras de pólen importado do Chile e da China e também amostras locais que mostraram estar infetadas com Psa (Vanneste et al., 2011). Os resultados de Gallelli et al. ( 2011) confirmaram que o pólen pode ficar infetado com Psa e constituir uma forma de disseminação da doença. A transmissão de Psa através do pólen, foi confirmada por Stefani & Giovanardi em 2011, quando conseguiram isolar Psa das flores e folhas, após 48 horas da aplicação de pólen inoculado com Psa.
A colonização das plantas pela bactéria pode ocorrer em qualquer altura do ano (Figura 6), no entanto existem épocas em que pode ser mais favorecida pelas condições climáticas e operações culturais que são realizadas nessa altura, nomeadamente no fim do inverno, início da primavera e no outono. No fim do inverno, a operação cultural da poda de inverno constitui possíveis entradas da bactéria na planta facilitando a sua colonização, para além de existir produção de exsudados o que aumenta a carga microbiana que possa existir no pomar nessa altura do ano (Ferrante et al., 2012). O início da primavera e do outono são alturas do ano também muito idênticas em termos das condições climáticas (temperatura e humidade) o que faz com que o nível de colonização por parte da bactéria seja idêntico e também em maior quantidade, pois a sua multiplicação é maior e a dispersão dentro da planta e entre plantas ocorre nesta altura (Ferrante et al., 2012; Scortichini et al., 2012).
A bactéria pode entrar por aberturas naturais na planta como por exemplo as lenticelas, os tricomas danificados e as cicatrizes deixadas pela queda das folhas (Spinelli et al., 2011; Kim et al. 2017).
13
2.6. Meios de protecção
Causando esta doença perdas económicas significativas, e sabendo que não existem métodos curativos, é de toda a importância proteger ao máximo os pomares, de maneira a minimizar os prejuízos que possam ser provocados pela doença e, portanto, utilizando os vários meios de protecção como culturais, biológicos e químicos.
2.5.1. Meios de protecção cultural
Os meios de protecção cultural passam pela consciência do produtor, principalmente após todas as operações culturais que possam provocar feridas abertas na planta, que são possíveis portas de entrada da bactéria, devendo ser sempre desinfectadas e devidamente protegidas para que não haja colonização desses tecidos.
Segundo o Plano de Acção Nacional para o controlo de Pseudomonas syringae pv. actinidiae (DGAV, 2014), todas as plantas que apresentam sintomas da doença, nomeadamente aquelas que apresentam exsudados avermelhados, devem ser cortadas até um metro abaixo do ponto de libertação dos exsudados. As plantas com sintomas menos severos, nos ramos devem ser podadas até 70 cm abaixo do ponto de infecção e onde se observam os sintomas. No caso das folhas, os ramos devem ser podados no inverno, seguido de aplicações com produtos à base de cobre, cobrindo todas as feridas que forem feitas na planta, e minimizando o tamanho e o número de feridas resultantes.
Apesar de com as intervenções referidas anteriormente se remover todo o material sintomático, nunca é removida a totalidade do inóculo, pois apenas é removido o material que teve oportunidade de expressar todos os sintomas da doença (Vanneste et al., 2011). Assim, todas as plantas assintomáticas ou as que possuem populações epifíticas (bactérias no exterior da planta) constituem um grupo que a bactéria está presente nas plantas e, permitindo que se continue a espalhar pelo pomar (Vanneste et al., 2011).
Na instalação de pomares é necessário a utilização de plantas sãs, acompanhadas por um passaporte fitossanitário que garante a isenção da bactéria (DGAV, 2014).
2.5.2. Meios de protecção biológica
2.5.2.1. Bacteriófagos
Os bacteriófagos (fagos) caracterizam-se por serem entidades biológicas mais abundantes na biosfera. Os bacteriófagos são vírus específicos de bactérias que destroem o
14 metabolismo dos seus hospedeiros e se replicam provocando a lise e morte do hospedeiro (Buttimer et al., 2017).
Na cultura da actinídea os bacteriófagos são uma alternativa bastante promissora, pois existem naturalmente nos pomares e, portanto, são uma alternativa viável para o meio ambiente e também para as próprias plantas infectadas. Por outro lado, já existem alguns produtos que incluem bacteriófagos apesar de serem muito pouco utilizados devido à sua extrema sensibilidade a condições ambientais, como por exemplo, a exposição a raios UV que podem diminuir as populações de bacteriófagos (Iriarte et al., 2007). .
Frampton e os seus colaboradores (2014) conseguiram obter bacteriófagos que estavam presentes em pomares de kiwi na Nova Zelândia. Estes bacteriófagos que foram testados, mostraram capacidade de infectar as duas populações de Psa existentes na Nova Zelândia, Psa 3 e Psa 4 (P. syringae pv. actinidifoliorum pv. nov.).
Também na Coreia, Yu e colaboradores, (2015), conseguiram isolar bacteriófagos de solos de pomares que actuavam contra P. syringae pv. actinidiae, mostrando ser resistentes a situações adversas, o que é mais uma garantia de que esta é uma opção tecnicamente viável, tendo em conta o local onde são aplicados, e as condições meteorológicas que podem provocar alterações no seu efeito (Yu et al., 2015).
Existem relatórios da existência de resistência aos bacteriófagos, devido principalmente a mutações ou mudanças no sítio no qual o bacteriófago iniciou a infecção. O uso de uma mistura de bacteriófagos com diferentes métodos de infecção pode ser uma alternativa à ocorrência da resistência (Woodcock, 2016).
Também em Portugal existem referências sobre este assunto, tendo sido demonstrado em 2015 (Garcia, 2015), que em alguns pomares do Entre Douro e Minho foram isolados bacteriófagos de Psa. Estes apresentaram a capacidade de causar a morte de bactérias da população portuguesa estudada e também da estirpe italiana CFBP 7286.
2.5.2.2. Bacillus subtilis
Bacillus subtilis é a bactéria mais estudada do grupo das Gram positivas. É sabido que este organismo é capaz de produzir cerca de 20 antibióticos, para além de produzir compostos activos anti-microbianos predominantemente péptidos que podem estar envolvidos na formação de biofilmes, o que permite uma proteção avançada da doença (Stein et al., 2005). Na formação de biofilmes pode estar envolvida a secreção por parte de Bacillus subtilis, da substância surfactina (lipopéptido) conhecida pela sua acção biosurfactante (bio-tensioactivo)
15 (Bais et al., 2004). Os biofilmes podem ser caracterizados pela sua comunidade de células estruturadas que aderem com facilidade a uma superfície e que cobrem uma matriz extracelular (Bais et al., 2004).
O biofungicida comercial, Serenade Max®, possui na sua constituição uma estirpe de Bacillus subtilis, QST 713, que possui ação antibacteriana através da formação de um biofilme que previne e evita a fixação de agentes patogénicos no hospedeiro, prevenindo também a sua penetração (Bayer® Crop Science, 2009). Pode induzir a resposta sistémica da planta favorecendo ainda mais a sua resistência a infecções de agentes patogénicos, sendo, por esta razão, uma opção bastante viável para o controlo biológico de Psa. A utilização deste produto permite uma indução da resistência sistémica por parte das plantas. O produto tem uma acção bacteriostática e actua por contacto possuindo um largo espectro de acção, tendo uma persistência biológica de 7 dias e um intervalo de segurança de 3 dias.
2.5.2.3. Leveduras
As leveduras são agentes de controlo biológico bastante efectivos que conseguem colonizar com rapidez as superfícies das folhas e podem sobreviver por longos períodos de tempo independentemente das condições ambientais, para além de não serem afectados pelos fungicidas comerciais (Rosa-Magri et al., 2011). O seu modo de acção passa principalmente pela competição por nutrientes, prevenção da colonização de Psa na planta e também pela produção de compostos antibacterianos (Hatoum et al., 2012). No entanto, a sua interação com a Psa ainda não é conhecida.
Hoyte e os seus colaboradores (2013), isolaram leveduras da superfície de várias culturas, incluindo Actinidea. As culturas de leveduras codificadas como YM1 e YM2, mostraram uma elevada actividade contra Psa, mas apenas a YM2 foi selecionada para as experiências em campo. Elmer e os seus colaboradores (2014), mostraram o potencial de controlo biológico da levedura YM2, no entanto os seus componentes, YCom1 e YCom2 mostraram, respectivamente, moderada e elevada susceptibilidade ao hidróxido de cobre e, portanto, esta levedura não foi utilizada comercialmente. .
Elmer et al. (2014) realizaram novos estudos com as leveduras, YBCA4 e YBCA5, no entanto apenas avançaram com YBCA5 por ser mais barata, exibir elevada tolerância ao cobre e apresentar elevada sobrevivência na superfície da folha. No entanto ainda não foi registada para a proteção contra Psa, pois ainda são necessários mais estudos.
16
2.5.3. Meios de protecção química
2.5.3.1. Cobre
O cobre é um dos micronutrientes mais utilizados na agricultura, tanto como nutriente essencial às plantas, mas também como meio de luta contra agentes fitopatogénicos e permitido no modo de produção biológico. Os iões de cobre interrompem os processos celulares que ocorrem em conjunto com a bactéria, levando à morte desta (Woodcock, 2016).
A eficácia do cobre é determinada pelo tamanho das partículas, pela retenção e pela solubilidade. O tamanho das partículas está diretamente relacionado com a retenção, tempo que as partículas permanecem na superfície da planta. Quando as partículas têm tamanho reduzido resulta numa área de superfície maior, o que aumenta a presença de partículas e consequentemente a quantidade de iões de cobre. Este aumento da densidade das partículas permite que aumente a eficácia da atividade antibacteriana do cobre (Woodcock, 2016). Assim, se a área de superfície da planta for maior permite que as partículas de cobre se mantenham durante mais tempo sob a superfície. As partículas de maiores dimensões, não são adequadas, pois estão sujeitas a maiores lavagens e, portanto, a não permanecerem tanto tempo na superfície da planta (Woodcock, 2016). A solubilidade determina a duração da protecção que o cobre pode fornecer, ou seja, formas mais solúveis desaparecem mais rápido e terão que ser aplicadas mais frequentemente, enquanto que formas mais insolúveis libertam iões de cobre durante mais tempo e requerem aplicações menos frequentes (Woodcock, 2016).
Outro factor que pode ser determinante na eficácia do cobre é o desenvolvimento de novas folhas e lançamentos após uma aplicação de cobre, pois existe crescimento da planta e, consequentemente, aumenta a área vegetal desprotegida e, portanto, a suscetibilidade à bactéria é maior. Portanto, uma nova aplicação de cobre deve ser controlada no tempo e no espaço de forma a conceder à planta a defesa necessária perante a bactéria (Woodcock, 2016).
No entanto, este composto, sendo usado em grandes quantidades pode ter efeitos fitotóxicos provocando danos nas plantas, desde descoloração dos tecidos, danos na superfície das folhas, danos nos frutos e também redução do vigor da planta. Estes efeitos podem ser causados por variadas condições como por exemplo, a concentração do cobre, o pH da solução, humidade da planta, temperatura do produto, humidade relativa do ar e o estado de crescimento da planta (KVH, 2016). Apesar desta fitotoxicidade ocorrer, existem também efeitos de resistência por parte dos agentes patogénicos, questão que está a ocorrer com Psa para determinados compostos de cobre, como é explicado por Lindow (2017). Esta resistência acontece principalmente pela expressão dos genes cop, que codificam proteínas
17 que permitem que Psa tolere as altas concentrações de cobre (Masami et al., 2004). Estes genes também podem ser adquiridos pela bactéria através de outras bactérias que possam existir no ambiente, transferindo esta capacidade de resistência a Psa e conferindo-lhe assim resistência ao cobre (Colombi et al., 2017).
Sendo as pulverizações com compostos de cobre as mais utilizadas em termos de protecção da cultura da actinídea, estas devem ser efectuadas nas fases do ciclo vegetativo com maior risco de infecção tendo sempre em consideração as consequências do uso do cobre a longo termo, nomeadamente o aumento de genes de resistência de bactérias do solo e também o aumento dos genes de resistência aos antibióticos (Hu et al., 2016).
2.5.3.2. Estreptomicina
A estreptomicina é um antibiótico que é utilizado para o controlo de doenças de plantas e é considerado o tratamento mais eficaz para Psa (KVH, 2014). A sua capacidade antimicrobiana deve-se ao facto de interferir na tradução do mRNA das proteínas o que leva à morte celular (Cameron & Sarojini, 2014). O uso da estreptomicina foi eficaz contra Psa tanto em laboratório como no campo (KVH, 2014). A resistência à estreptomicina pode ocorrer de duas maneiras. A primeira pode ser provocada por uma mudança no ADN, ou seja, uma mutação do gene rpsL, o qual confere a resistência através do não funcionamento do antibiótico (Springer et al., 2001). A segunda forma de resistência é adquirida através da aquisição dos genes codificados como StrA e StrB, obtidos através de outras bactérias e que podem tornar o antibiótico inerte (Chiou & Jones, 1995).
Este antibiótico não é permitido nos países da União Europeia e apenas é permitido na Nova Zelândia, em casos extremos, devido principalmente ao facto da facilidade com que determinadas bactérias podem manifestar resistência, não só em usos na agricultura, como também em tratamentos com humanos.
2.5.3.3. Terpenos
Os terpenos mais conhecidos são o geraniol e o citronelol sendo encontrados principalmente em óleos essenciais ou nos seus derivados. O geraniol é encontrado em quase todos os óleos essenciais contendo terpenos, como o óleo de palmarosa, o gerânio e óleo de rosas. O citronelol pode ser encontrado em plantas como o eucalipto-limão (Eucalyptus citriodora). Segundo Cameron e Sarojini (2014), os dois terpenos têm efeitos inibitórios contra Psa, sendo o geraniol mais eficaz que o citronelol (Ferrante & Scortichini, 2010).
18 Estas substâncias não são utilizadas para aplicações em campo por serem susceptíveis de gerarem resistências e portanto, a sua capacidade antibacteriana acaba por ser ainda mais diminuta (Cameron & Sarojini, 2014).
2.5.3.4. Quitosana
A quitosana é obtida a partir de cascas de crustáceos, sendo o camarão o mais utilizado, e possui actividade antimicrobiana contra a Psa, tendo mostrado resultados positivos nesse sentido apenas in vitro (Ferrante & Scortichini, 2010). Este composto tem variadas vantagens na sua utilização, sendo a biocompatibilidade e a biodegradabilidade as que mais se sobressaem. O seu baixo grau de acetilação resulta em maiores propriedades antimicrobianas, para além de que a quitosana foi delineada para interagir com as membranas dos patogéneos levando à libertação de compostos intracelulares destes (Cameron & Sarojini, 2014).
A quitosana é conhecida também por inibir o crescimento microbiano e a libertação de enzimas bacterianas que inibem a produção de toxinas por parte dos patogéneos. A quitosana ativa também os sistemas de defesa natural da planta e potencia o crescimento da mesma (Cameron & Sarojini, 2014).
Segundo Scortichini (2014), a quitosana mostrou melhores resultados que os compostos de cobre, em termos de redução dos sintomas da doença e na redução dos exsudados nos troncos presentes no fim do Inverno.
Esta substância mostrou ter bastantes benefícios e, portanto, pode ser aplicada em campo, sendo que pode ser adquirida através de laboratórios registados para tal na Europa, como o ChitosanLab, que está presente em França e cujas encomendas dos produtos podem ser feitas através do website.
2.5.3.5. Péptidos antimicrobianos
Os péptidos antimicrobianos conservados no sistema imunitário inato do complexo multicelular dos organismos e pode afetar bactérias no seu geral. Estes péptidos podem ter menos de 50 aminoácidos, sendo estes anfipáticos na natureza e de carga positiva (catiónica), permitindo-lhes serem solúveis em meios aquosos o que aumenta a sua capacidade antimicrobiana (Cameron & Sarojini, 2014). Os péptidos antimicrobianos atingem a membrana da bicamada lipídica, apesar de ainda não serem bem conhecidos os seus métodos de acção, danificando-a e provocando a lise total. No caso das bactérias Gram negativas, como é o caso
19 de Psa, tanto a camada externa como a interna contêm grupos aniónicos, o que permite a ligação dos péptidos antimicrobianos, pois estes têm carga catiónica (Cameron & Sarojini, 2014).
Tendo em conta a falta de informação sobre o modo de ação/controlo biológico destes péptidos, e como ainda são testes laboratoriais, a eficácia da aplicação em campo ainda não é conhecida, sendo por isso uma opção viável, mas que requer mais estudos e confirmações de eficácia.
2.5.3.6. Indutores das defesas das plantas
Os indutores das defesas das plantas são substâncias que ativam o sistema natural de defesa das plantas, o que permite que estas sejam menos susceptíveis à infeção por bactérias, como é o caso da Psa.
Acibenzolar-S-metil (ASM) é um dos indutores com melhores resultados em ensaios em estufa com A. chinensis e A. deliciosa, tendo como nome comercial Bion® na Europa e Actigard® na Nova Zelândia, sendo produtos da empresa Syngenta® (Reglinski et al., 2013).
O ASM é análogo funcional da hormona ácido salicílico que é produzido durante a infecção provocada por Psa e coordena também a resposta da planta à infeção causada pela bactéria (Reglinski et al., 2013). Quando o ASM é aplicado às folhas dActinidea, esta prepara os tecidos para uma possível infeção por parte de Psa e para que estes possam reconhecer a bactéria o mais eficientemente possível, fazendo com que a bactéria não seja tão eficaz na colonização de folhas tratadas com ASM (Reglinski et al., 2013). Sabe-se que estes compostos são muito eficazes em condições controladas, no entanto, em campo, o hospedeiro e até a própria bactéria podem ter alterações inesperadas, o que pode levar a questões na utilização deste indutor de defesas da planta.
Este indutor, porém, mostrou ser significantemente eficaz na prevenção da infecção da doença, comparativamente com outros produtos análogos de hormonas (KVH, 2015).
Os produtos autorizados para a aplicação no pomar com a finalidade do controlo ou protecção da cultura contra a Psa encontram-se no quadro seguinte (Quadro 2).
20 Quadro 2. Produtos fitofarmacêuticos autorizados para combate/controlo da Psa em Portugal (Associação Portuguesa de Kiwicultores, 2017b).
Produto Empresa S.a. Concentração Dose Modo de Aplicação Nº Máx.
Aplicações Cobre Nordox 75 WG Massó oxido cuproso 37,5-75g/hl 0,5-1kg/ha Pulverizar após a queda da folha e antes do abrolhamento dos gomos 4
Cobre Nordox 75 WG Massó oxido cuproso 37,5-75g/hl 375-500g/ha Estes tratamentos podem ser realizados a partir do abrolhamento (a cada 30 dias com interrupção na floração) e até 20 dias antes da colheita
3
Cupravit Bayer oxicloreto de cobre Máx 2,5 kg/
ha
1 tratamento no fim da colheita; 1 a 2 tratamentos durante a queda da folha (50% e 100%); 1 tratamento após a poda de inverno e 1 tratamento à rebentação
5
Cuprital Sapec oxicloreto de cobre 150-250ml/hl 1,5-2,5L/ha 1 tratamento no fim da colheita; 1 a 2 tratamentos durante a queda da folha (50% e 100%); 1 tratamento após a poda de inverno e 1 tratamento à rebentação
4/5
Cuprital SC Sapec oxicloreto de cobre 100-175g/hl 1-1,75L/ha Realizar aplicação de carácter preventivo, no final da colheita, à queda das folhas, após podas de inverno, à rebentação
4/5
Cuprocol Syngenta oxicloreto de cobre 150-250ml/hl - Efetuar 3 tratamentos, respectivamente, no inicio, meio e fim da queda das folhas. Pode causar fitotoxicidade se for aplicado com tempo frio e chuvoso.
3
Calda Bordalesa Vallés
Industrias Vallés sulfato de cobre e cálcio
750-833,3g/hl 3-5kg/ha Tratar durante a queda das folhas, após a poda de inverno e à rebentação 3
Calda Bordalesa Sapec
Sapec sulfato de cobre e
cálcio
1,25 kg/hl - Tratar no final da colheita, queda das folhas e após poda de inverno 3
Cuproxat Nufarm sulfato de cobre 57-76 g/hl 300-400ml/hl Tratamentos a realizar no fim da colheita, durante a queda das folhas e, após a poda de inverno e/ou rebentação.
5
Serenade Max Bayer Bacillus subtilis 0,417-1kg/hl 2,5-4kg/ha Tratar durante a floração 4
Bion 50 WG Syngenta
acibenzolar-S-methyl
100-200g/ha Tratar durante o desenvolvimento vegetativo, quando as condições ambientais forem favoráveis ao desenvolvimento da doença. (Para aplicação em sistema de fertirrigação)
21
2.6.
Prospecções oficiais de Psa em pomares na Região de Entre
Douro e Minho
Devido à perigosidade e à sua presença em vários países do sul da Europa, a bactéria Pseudomonas syringae pv. actinidiae (Psa), está incluída na lista A2 da Organização Europeia e Mediterrânica para a Proteção das Plantas (EPPO/OEPP), sendo reconhecida como um organismo nocivo para as plantas de Actinidea, desde novembro de 2009. Esta doença tem como hospedeiros as plantas do género Actinidea, de que se destacam as espécies A. deliciosa, A. chinensis, A. arguta e A. kolomikta e não são conhecidos métodos de luta curativos. Assim, face à importância da cultura, da perigosidade da doença e dos riscos a ela associados, a Direcção Geral de Alimentação e Veterinária (DGAV) implementou o Plano Nacional para o Controlo de Pseudomonas syringae pv. actinidiae (Psa) (DGAV, 2012). Para elaboração e apoio à implementação deste Plano foi constituído um Grupo de Acompanhamento composto por elementos da DGAV, representantes da DRAP Norte e da DRAP Centro, do INIAV, da Federação Nacional das Cooperativas Agrícolas de Hortofruticultores (FENAFRUTAS), da Associação de Viveiristas do Distrito de Coimbra (AVDC), da Associação Portuguesa de Kiwicultores (APK) e da Frutas Douro ao Minho, S.A. Entre outras acções, constam deste Plano a realização de prospeções oficiais de Psa realizadas anualmente pela Autoridade Fitossanitária Nacional, sendo a DGAV a principal responsável, tendo como prioridade: i) todo o território reconhecido como áreas livres da doença; ii) nos pomares onde foi detetada a doença em anos anteriores e áreas circundantes e iii) os viveiros e campos de plantas-mãe e respectivas áreas adjacentes. São ainda implementadas ações de inspeção em unidades de distribuição e comercialização de pólen vivo e de vegetais de kiwi destinados à plantação.
22
Material e Métodos
1.
Acompanhamento das prospecções oficiais de Psa em pomares
na Região de Entre Douro e Minho nos anos de 2016 e 2017
As prospecções que acompanhamos, nos anos de 2016 e 2017, durante os meses de junho e julho, foram realizadas pela Engª Alexandra Gomes, técnica da Associação Portuguesa de Kiwicultores responsável por estas ações, e incidiram apenas em pomares da região do Entre Douro e Minho e da Beira Litoral onde a doença não tinha sido ainda identificada. No total foram visitados 12 pomares, 9 em 2016 e 3 em 2017.
As amostras a colher para identificação da bactéria consistiam em material vegetal (segmentos de varas) com tamanho aproximado de 30 cm com folhas, preferencialmente sem flores e se possível com sintomas da doença. No total, a amostra de um pomar para análise continha 20 segmentos, sendo 10 destes de plantas macho e 10 de plantas fêmea.
As amostras colhidas nos pomares sem sintomas incluíam duas sub-amostras, em que o material recolhido de plantas macho era separado das plantas fêmeas.
A amostra de cada pomar era colocada em sacos pretos devidamente identificados, sendo também preenchida uma ficha com dados do pomar e do produtor.
A análise do trabalho resultante do acompanhamento destas prospecções foi realizada com base nas observações visuais efetuadas e com base nas informações das fichas preenchidas, onde constavam dados como a área dos pomares, os sintomas presentes nas plantas, os tratamentos fitossanitários e as práticas culturais efectuadas, as cultivares, idade do pomar, o sistema de condução utilizado e a localização do pomar. Todas estas informações foram importantes para caracterizar o panorama da doença na região do Entre Douro e Minho no período 2016/2017.
23
2. Estudo do cancro bacteriano da actinídea num pomar na Região do Entre
Douro e Minho
2.1. Caraterização do pomar
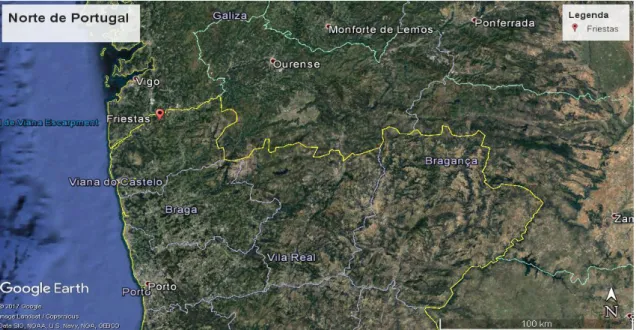
O pomar onde foi realizada a parte experimental deste trabalho, e o qual serviu para recolha de material vegetal, está a uma altitude de cerca de 30 m, tem uma área de cerca de 1,15 ha e localiza-se na região do Entre Douro e Minho (Figura 7), na freguesia de Friestas, concelho de Valença do Minho, zona onde foi detectada pela primeira vez em Portugal a doença do cancro da actinídea (Psa) (Balestra et al., 2010).
A área experimental do pomar é constituída por 2 sub-pomares, um novo, composto por plantas com cerca de 6 anos e um velho, com plantas com cerca de 30 anos. Ambos compostos por plantas da cultivar “Hayward”, (plantas fêmea), e com plantas macho das cultivares “Tomuri”, “Matua” e “Chieftain”. As plantas estão dispostas num compasso de 4 m x 3 m, perfazendo uma densidade de plantas de cerca de 400 plantas/ha. As plantas estão conduzidas em cruzeta ou “T-bar”, compostas por um poste vertical com cerca de 2,00 m de altura e com uma estrutura horizontal, a cruzeta, com dimensões que variam entre 1,50 m e 2 m e que suporta a zona central da planta, local de onde são emitidas as varas produtivas, que é composta por um tronco de cerca de 1,80 m de altura.
O sistema de rega do pomar é por microaspersão com um débito de cerca de 1L/min. As operações culturais que se realizam no pomar incidem principalmente na manutenção do pomar, assegurando que o vigor e crescimento dos ramos seja o mais adequado para se atingir os objetivos de produção e de qualidade pretendidos. No quadro