PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
ANATOMIA ÓSSEA DA CINTURA PELVINA, ESTILOPÓDIO E
ZEUGOPÓDIO E MUSCULAR DA CINTURA PELVINA E
ESTILOPÓDIO DE
Caiman
crocodilus
MARILUCE FERREIRA ROMÃO EDUCADORA FÍSICA
UNIVERSIDADE FEDERAL DE UBERLÂNDIA FACULDADE DE MEDICINA VETERINÁRIA
ANATOMIA ÓSSEA DA CINTURA PELVINA, ESTILOPÓDIO E
ZEUGOPÓDIO E MUSCULAR DA CINTURA PELVINA E
ESTILOPÓDIO DE
Caiman
crocodilus
MARILUCE FERREIRA ROMÃO Orientador: Professor Dr. André Luiz Quagliatto Santos
Tese apresentada à Faculdade de Medicina Veterinária – UFU, como parte das exigências para obtenção de título de Doutor(a) em Ciências Veterinárias (Saúde Animal).
Dados Internacionais de Catalogação na Publicação (CIP) Sistema de Bibliotecas da UFU, MG, Brasil.
761a
015
Romão, Mariluce Ferreira, 1974-
Anatomia óssea da cintura pelvina, estilopódio e zeugopódio e muscular da cintura pelvina e estilopódio de Caiman crocodilus / Mariluce Ferreira Romão. - 2015.
42 f.
Orientador: André Luiz Quagliatto Santos.
Tese (doutorado) - Universidade Federal de Uberlândia, Programa de Pós-Graduação em Ciências Veterinárias.
Inclui bibliografia.
1. Veterinária - Teses. 2. Jacaretinga - Teses. 3. Anatomia veterinária - Teses. 4. Músculos - Teses. I. Santos, André Luiz Quagliatto. II. Universidade Federal de Uberlândia. Programa de Pós-Graduação em Ciências Veterinárias. III. Título.
“JESUS ENSINOU COMO UM CANTO É UMA ORAÇÃO”
“Vamos acender essa luz, chamar os nossos filhos, ou que estejam acordados ou que estejam dormindo” “Vou acender essa luz, vou acordar meu filho, ou que esteja acordado, ou que esteja dormindo...”
***
AGRADECIMENTOS
A menção honrosa na obtenção de um título acadêmico, não se limita
simplesmente aos resultados demonstrados e alcançados, e sim, em todo o caminho
percorrido, considerando cada percalço, subsídios e pessoas envolvidas.
Ao professor Dr. Zenon Silva, na condição de grande idealizador da
minha progressão acadêmica, o meu muito obrigada pela motivação, que foi
essencial nas minhas decisões iniciais. Incomensurável é a minha gratidão pelo
conhecimento compartilhado, e a admiração pelo profissional e ser humano que
representa.
Ao professor Dr. André Luiz Quagliatto Santos, torna-se pleno o meu
respeito e admiração pela acolhida sem preconceitos, tendo em vista a minha área
de formação acadêmica, e a forma impecável na condução de todas as orientações,
entre Mestrado e Doutorado. Muito obrigada por tudo!
Aos professores, Dra. Daniela Cristina Silva, Dra. Lucélia Gonçalves
Vieira, Dr. Gilmar da Cunha Sousa, Dr. Júlio Roquete Cardoso, agradeço a
participação na avaliação, e, por conseguinte, contribuição no nosso trabalho.
Ao meu esposo, Emerito Ferreira Júnior, que sem restrições esteve ao
meu lado, em toda a minha trajetória acadêmica, sempre com uma palavra ou gesto
de apoio, os meus sinceros agradecimentos. O esforço foi nosso, e o desfecho é
consequência da nossa perseverança, o que reitera continuarmos seguindo juntos
pela vida, até que as nossas missões sejam finalizadas, e portanto, se eternizem.
À minha filha, Camila Ferreira Romão, em primeiro lugar me desculpo
pelas inevitáveis ausências, e, sobretudo, agradeço a compreensão e o apoio
À Faustina Bemfica dos Santos (In Memorian), saudosa sogra, a quem
agradeço as orações que foram feitas, ainda em vida, na intenção de que tudo
sempre fosse bem assistido e encaminhado por Deus, fez toda a diferença no meu
equilíbrio espiritual. Os meus agradecimentos e gratidão são extensivos aos
cuidados especiais com a minha filha, a nossa Camila, que a teve e eterniza uma
referência também materna. Sem outra alternativa, seguimos pesarosos, com a sua
ausência...
Ao meu pai, Roberto Romão das Dores, e à minha mãe Maria dos Anjos
Romão, agradeço sempre pelo incentivo.
Ao Fabiano Campos Lima, agradeço a indicação ao ingresso no
Programa de Pós graduação em Ciências Veterinárias, e, sobretudo, à credibilidade
sempre mantida, bem como, as inúmeras contribuições.
À equipe do Laboratório de Ensino e Pesquisa em Animais Silvestres,
SUMÁRIO
Página
CAPÍTULO 1: CONSIDERAÇÕES GERAIS...
CAPÍTULO 2: Anatomia óssea da Cintura Pelvina, Estilopódio e Zeugopódio de Caiman crocodilus...
1
6
CAPÍTULO 3: Anatomia Muscular da cintura pelvina e estilopódio de Caiman crocodilus...
CAPÍTULO 1 - CONSIDERAÇÕES GERAIS
A era Mesozoica, referida como era dos répteis, teve duração em média de 180 milhões de anos, à partir do Paleozoico, há 245 milhões de anos, até o começo do Cenozoico, há 66 milhões de anos. Neste período, os animais associados à ampla disseminação dos tetrápodes, foram os Archosauria, com destaque para os dinossauros e pterossauros. Estes, morfologicamente, evidenciavam gradações relevantes na postura dos membros, entre quadrúpedes e bípedes, modificações bruscas na dimensão corporal, como também, evolução para voo (POUGH et al., 2006). A distinção desses animais faz referência à existência de uma incisura rostro-orbital, formato da órbita ocular em triângulo reverso, dentes alongados lateralmente, com o padrão locomotor, em geral, tendenciosamente bípede, e com uma proeminência afilada na face ventral e proximal da diáfise femoral, identificada como quarto trocanter, ou trocanter interno (GATESY, 1997).
Uma vez dominantes no período mesozoico, os dinossauros foram subdivididos em duas ordens: Saurisquios, e Ornitísquios. Os saurísquios foram constituídos, em sua maioria, por répteis bípedes, carnívoros, e com três elementos radiais na cintura pelvina. Os ornitísquios apresentavam uma cintura pelvina, com projeção cranial e caudal dos ossos púbicos, conforme identificado nas aves, sendo herbívoros, e mistos, entre bípedes e quadrúpedes (ORR, 1986).
Os saurísquios (sauropodas), na representação dos maiores tetrápodes que já viveram na terra, tiveram uma evolução locomotora com inúmeras adaptações osteológicas para resistir à sua grande massa corporal. O hábito locomotor destes animais incluiu os membros colunares com ossos muito resistentes e alongados, bem como, redução de flexão dos membros, de locais de inserção musculares e das falanges distais laterais do membro pelvino (CARRANO, 1999; YATES, 2004). A cintura pelvina se articula com a coluna vertebral através das vértebras sacrais, servindo como ponto de transmissão de cargas axiais (RASSKIN-GUTMAN, BUSCALIONI, 2001). Nos Dinosauromorpha, estima-se de que em torno de 80% da massa corporal tenha sido suportada pela cintura pelvina e membros pelvinos (ALEXANDER, 1985).
ter tido um importante efeito sobre a organização muscular destes animais. (SERENO, 1991).
Os únicos remanescentes dos Archosauria são os aligátores, os crocodilos, e os gaviais (Gavialis gangeticus), que por sua vez, pertencem à ordem Crocodylia. Apesar destes animais apresentarem similaridades estruturais aos dinossauros bípedes, filogeneticamente, estão distanciados da árvore genealógica dos répteis. Em geral, não há tipicidade reptiliana em suas características anatômicas. Tais características são observadas semelhantes, em grande escala, às aves, que possuem seus ancestrais comuns aos Crocodylia (ROMER, PARSONS, 1985). As aves, uma vez considerados os Dinossauria mais avançados, com reconhecimento entre 1860 e 1870, possuem pescoço sinuoso, alongado e móvel, crânio e pescoço articulados através de um único côndilo occipital, articulação intertarsal no tornozelo, e ossos pneumáticos, bem como, as principais diferenças estão nas prática de voo e endotermia, ambos, associados às penas (POUGH et al., 2006).
Os Crocodylia, em geral, possuem ossos fortes e músculos potentes, que movimentam a mandíbula, permitindo uma ampla abertura e um fechamento rápido, com uma válvula palatal, que contribui com a respiração eficiente, mesmo enquanto a boca contém água, alimentos ou ambos (AZEVEDO, 2003; HICKMAN et al., 2006). Há possibilidades de certos níveis de salinidade serem suportados, devido às glândulas dessalinizadoras, que facilitam a permanência em águas salgadas e salobras, como mangues ou estuários (RUEDA-ALMONACID et al., 2007).
Na condição de animais ectotérmicos aquáticos, os Crocodylia são capazes de se adaptarem a diversas condições climáticas. A temperatura desses animais é controlada pelas trocas de energia com o meio ambiente, estando diretamente associadas aos eventos reprodutivos, alimentares, digestórios, e de crescimento (POUGH et al., 2003).
A ordem Crocodylia é dividida em três famílias: Crocodylidae (rostro relativamente estreito e cabeça comprida), Alligatoridae (rostro mais largo e cabeça curta), e Gavialidae (cabeça comprida, rostro estreito e longo) (ZUG et al., 2001; POUGH et al., 2003; AZEVEDO, 2003; HICKMAN et al., 2006).
Caiman yacare (jacaré-do pantanal) e Caiman latirostris (jacaré-do-papo amarelo). O
Caiman crocodilus enquanto juvenil é amarelado com manchas pretas nas laterais do corpo até a cauda, e quando adulto passa a apresentar coloração esverdeada. Possui uma crista acima dos olhos e sobre o dorso do focinho, com uma crota ossificada (crista infra orbital) em frente aos olhos, estes com destaque para a íris amarelo limão. Sua distribuição ocorre em ambientes abertos, de águas tranquilas, incluindo quatro subespécies de C. crocodilus: C. c. fuscus, C. c. crocodilus, C. c. chiapasius, C. c. apaporiensis. Anteriormente, C. yacare que era considerado uma subespécie de C. crocodilus, passou a ser considerado uma espécie (PEREIRA, MALVÁSIO, 2014).
Em relação ao padrão locomotor dos membros, anfíbios e lagartos assumem posturas em extensão gradual, enquanto os mamíferos e dinossauros mantém-se eretos, e os Crocodylia são capazes de assumir posições semi eretas, além de ocasionais extensões graduais, e rastejantes (REILLY, ELIAS, 1998). O estudo comportamental comparativo intra e entre espécies traz informações essenciais sobre a história evolutiva dos organismos, capaz de prover princípios comuns atitudinais nos grandes grupos (SILVA, COSTA, 2005).
Conjetura-se que a diversidade morfológica entre os animais seja responsável, em grande parte, por diferenças funcionais. Partindo deste pressuposto, a postura pode ser um indicador de considerações anatômicas, cinesiológicas e biomecânicas, e vice versa. Trata-se de correlação da postura predominante, com a inferência de maiores ou menores níveis de cargas tensionais, ou de características anatômicas que podem indicar a postura predominante (BLOB, 2000).
REFERÊNCIAS
ALEXANDER, M. R. Mechanics of posture and gait of some large dinosaurs. Zoological Journal of the Linnean Society, v.83, n.1, p.25, 1985.
AZEVEDO, J. C. N. Crocodilianos: Biologia, Manejo e Conservação. João Pessoa: Arpoador, 2003. 122p.
CARRANO, M. T. What, if anything, is a cursor? Categories versus continua in determining locomotor habit in mammals and dinosaurs. Journal of Zoology, v.247, n.1, p.29-42, 1999.
GATESY, S. An Electromyographic Analysis of Hindlimb Function in Alligator during Terrestrial Locomotion. Journal of Morphology, v. 234, p.197–212, 1997.
HICKMAN, C. P; ROBERTS, L. S; LARSON, A. Princípios integrados Crocodile Specialist Group. IUCN - The World Conservation de zoologia. Rio de Janeiro: Guanabara Koogan. 2006.
ORR, R. T. Biologia dos vertebrados. Editora Roca, São paulo. 1986. 508p.
PEREIRA, A. C.; MALVASIO, A. Síntese das características da ordem Crocodylia, fatores de influência em estudos populacionais e aspectos de seleção e uso de habitat para Caiman crocodilus e Melanosuchus nigerno Estado do Tocantins, Brasil. Macapá, v. 4, p. 111-118, 2014.
POUGH, F. H.; JANIS, C. M.; HEISER, J. B. A Vida dos Vertebrados. São Paulo: Atheneu: Editora. 2003. 699p.
POUGH F. H.; JANIS C. M.; J. B. HEISER. A vida dos vertebrados. 4. ed. São Paulo: Atheneu, 2006.
RASSKIN-GUTMAN, D.; BUSCALIONI, A. D. (2001). Theoretical morphology of the Archosaur (Reptilia: Diapsida) pelvic girdle. Paleobiology, v.27, p. 59-78, 2001.
REILLY, S. M.; ELIAS, J. A. Locomotion in Alligator mississippiensis: kinematic effects of speed and posture and their relevance to the sprawling-to-erect PARADIGM. The Journal of Experimental Biology, v. 201, p.2559–2574, 1998.
ROMER S. R.; PARSONS T. S. Anatomia comparada dos vertebrados. São Paulo: Atheneu, 1985.
RUEDA-ALMONACID, J. V.; CARR, J. L.; MITTERMEIER, R. A.; RODRÍGUEZ-MAHECHA, J. V.; MAST, R. B.; VOGT, R. C.; RHODIN, A. G. J.; DE LA OSSA-VELÁSQUEZ, J.; RUEDA, J. N.; ITTERMEIERC. G. Las Tortugas y los
SERENO, P. C. Lesothosaurus, “fabrosaurids,” and the early evolution of Ornithischia. Journal of Vertebrate Paleontology, v.11, p.168–197, 1991.
SILVA, G. M., COSTA, C. M. Descrição dos padrões motores do Tupinambis merianae (Duméril & Bilbron, 1839) (Squamata, Teiidae) e do Caiman crocodilus yacare (Daudin, 1802) (Crocodylia, Alligatoridae) em cativeiro no Zoológico Municipal de Curitiba, Paraná, Brasil.Revista brasileira de Zoociências, v. 7, n. 1, p. 93-109, 2005.
YATES, A. M. Anchisaurus polyzelus (Hitchcock): The smallest known sauropod dinosaur and the 197 evolution of gigantism among sauropodomorph dinosaurs. Postilla, v.230, n.1, p.58, 2004.
Capítulo 2 - Anatomia óssea da Cintura Pelvina, Estilopódio e Zeugopódio de Caiman crocodilus1
Mariluce F. Romão2*, André L. Q. Santos2
ABSTRACT. MariluceF. Romão, André L. Q. Santos [Bone Anatomy of the Pelvic Girdle, Stylopod and Zeugopod of the Caiman crocodilus]. Anatomia óssea da Cintura Pelvina,
Estilopódio e Zeugopódio do Caiman crocodilus. Pesquisa Veterinária Brasileira
00(0):00-00. Laboratório de Ensino e Pesquisa em Animais Silvestres, Faculdade de Medicina
Veterinária, Universidade Federal de Uberlândia, UFU. Rua Piauí, bloco 4s, bairro Jardim Umuarama, Uberlândia MG, 38400-902 Brasil. E-mail: mariluce.ferreira@terra.br.
The objective was to identify the pelvic girdle, stylopod and zeugopod bone anatomy of the Caiman crocodilus, assuming that such information contributes to improve the
knowledge about the locomotor system of these animals. Four samples of the C. crocodilus,
males, measuring an average length of 1,50m, adults belonging to the didactic collection of the Laboratory of Education and Research of Wild Animals (Laboratório de Ensino e Pesquisa em Animais Silvestres - LAPAS), from the Federal University of Uberlândia
(Universidade Federal de Uberlândia), already set in 10% formalin, were used. The
dissection was performed, initiating with a longitudinal incision, along the ventral median line of the trunk, folding the skin laterally, from the cervical region to the caudal thoracolumbar level, allowing the removal of the viscera en bloc. In the following, the pelvic girdle, the stylopod and zeugopod were disarticulated from the trunk, going through a maceration process with boiling and submersion in hydrogen peroxide at 20 volumes, until satisfactory clarification, for the identification and description of the bone features. A pelvic girdle was identified with three bony constituents: ilium, ischium and pubis. The ilium, with dorsolateral localization, is constituted by a wing and a body. The ischium, localized ventrolaterally and caudal, is constituted by a body, a ramus and a slate with a larger caudal extension. The pubis, with ventral and cranial placement, is constituted by a body, a ramus and a slate with larger cranial extension. The stylopod consists of the femur, with emphasis to the fourth trochanter most prominent in fossil reptiles. The zeugopod consists of the tibia with larger diameter, located medially, and the fibula, with smaller diameter, located laterally. The C. crocodilus
presents larger similarity with living archosaurs, in view of the locomotor behavior that defines and explains, in large part, the adaptations to the evolutionary process, as well as the influence of habits from terrestrial ancestrals, and currently, the semiaquatic Crocodylia.
INDEX TERMS: Skeleton, spectacled caiman, morphology.
RESUMO.-Objetivou-se identificar a anatomia óssea da cintura pelvina estilopódio e zeugopódio do Caiman crocodilus, partindo do pressuposto, que tais informações contribuem
para aprimorar o conhecimento acerca do aparelho locomotor desses animais. Foram utilizados quatro exemplares de C. crocodilus, machos, medindo em média de 1,50 m de
comprimento, adultos, pertencentes ao acervo didático do Laboratório de Ensino e Pesquisa em Animais Silvestres (LAPAS), da Universidade Federal de Uberlândia, encontrados fixados em formol a 10%. Foi realizada a dissecação, iniciada com uma incisão longitudinal, ao longo
1 Recebido em... Aceito para publicação em...
2 Laboratório de Ensino e Pesquisa em Animais Silvestres, Faculdade de Medicina Veterinária,
da linha mediana ventral do tronco, rebatendo-se a pele lateralmente, da região cervical até o nível caudal toracolombar, possibilitando a retirada das vísceras em bloco. Na sequência, foram desarticulados do tronco, a cintura pelvina, o estilopódio e o zeugopódio, passando por processo de maceração com fervura, e submersão em peróxido de hidrogênio a 20 volumes, até a clarificação satisfatória, para a identificação e descrição dos acidentes ósseos. Foi identificada uma cintura pelvina com três constituintes ósseos: ílio, ísquio e pube. O ílio, em localização dorsolateral, é constituído por uma asa e um corpo. O ísquio, localizado ventrolateralmente e caudal, é constituído por um corpo, um ramo e uma tábula com maior extensão caudal. O pube, com localização ventral e cranial, é constituído por um corpo, um ramo, e uma tábula em maior extensão cranial. O estilopódio é constituído pelo fêmur, com destaque para o quarto trocanter mais proeminente nos répteis fósseis. O zeugopódio é constituído pela tíbia com maior diâmetro, localizada medialmente, e a fíbula, com diâmetro menor, localizada lateralmente. O C. crocodilus apresenta maior similaridade com os
arcossauros viventes, tendo em vista o comportamento locomotor que define e explica, em grande parte, as adaptações ao processo evolutivo, bem como, a influência dos hábitos ancestrais terrestres, e atualmente, os Crocodylia semi-aquáticos.
TERMOS DE INDEXAÇÃO: Esqueleto, jacaré tinga, morfologia.
INTRODUÇÃO
Os arcossauros possuem uma cintura pelvina diferenciada, tendo em vista o processo evolutivo correlato ao desenvolvimento dos hábitos bípedes. Nestes répteis, o acetábulo passou a apresentar uma abertura em sua base, para melhor coaptação da cabeça do fêmur (Romer & Parsons 1985).
A evolução postural dos Crocodylia é considerada uma reflexão secundária, tendo em vista a adaptação ao estilo de vida semi aquático. Apesar da sua progressão funcional, em bases gerais, a estrutura óssea dos Crocodylia é correspondente à dos arcossauros, entretanto, o crânio e a pelve são referidos como especializados. Trata-se da incisura rostro-orbital e acetabular (Hutchinson & Gatesy 2006; Pough, Janis & Heiser 2006).
Há evidências de que a cintura pelvina contribua ativamente com a ventilação pulmonar em Crocodylia, considerando a associação de elementos locomotores apendiculares à respiração diafragmática, sem que ocorra prejuízo de uma, em detrimento à outra (Carrier & Farmer 2000a).
O fêmur nos Crocodylia é considerado tanto elemento de apoio estrutural, como área para de origem e inserção muscular. Em conformação primitiva, a sua diáfise apresentava-se retilínea, priorizando a contribuição na protração do membro. Conforme evolução postural, a tíbia passou a ser o principal ponto de apoio no zeugopódio, em relação à fíbula, que nos anfíbios, era relativamente mais desenvolvida (Romer 1923).
O Caiman crocodilus (Linnaeus, 1758) (Crocodylia: Alligatoridae), conhecido como
jacaré tinga, é um crocodiliano de porte médio, chegando a medir, em média dois metros e meio, quando adulto. Pode ser encontrado no Sul do México, América Central, América do Sul, distinguido pelo corpo esverdeado, com manchas amarelas e cinza entremeadas (Brazaitis et al. 1998).
Considera-se que estudos sobre a Anatomia da cintura pelvina e membro pelvino dos Crocodylia não só possibilitam associações entre forma e função, como também, correlaciona implicações evolutivas e locomotoras acerca dos Archosauria (Hutchinson & Gatesy 2000).
Algumas pesquisas sobre Anatomia óssea da cintura pelvina, e membro pelvino de
(Liparini 2011), Rauisuchia (Raugust 2014), e Alligator mississipiensis (Blob 2000), além de
literatura clássica (Reese 1915, Romer 1923, Rowe 1986) foram considerados como parâmetros comparativos.
No presente estudo, objetivou-se estudar a Anatomia óssea da cintura pelvina, estilopódio e zeugopódio do C. crocodilus, partindo do pressuposto, que tais informações
contribuem para aprimorar o conhecimento acerca do aparelho locomotor desses animais. Vale ressaltar a escassez de informações correlatas reunidas (Stannius 1854, Reese 1915, Romer 1923, Rowe 1986, Wilhite 2003).
MATERIAL E MÉTODO
Foram utilizados quatro exemplares de C. crocodilus, machos, medindo em média 1,50
m de comprimento, adultos, pertencentes ao acervo didático do Laboratório de Ensino e Pesquisa em Animais Silvestres (LAPAS), da Universidade Federal de Uberlândia, fixados em formol a 10%. Os animais são referentes ao SISBIO nº. 13159-1, e com parecer de ética aprovado pela CEUA/UNIPAM.
Foi realizada a dissecação, iniciada com uma incisão longitudinal, ao longo da linha mediana ventral do tronco, rebatendo-se a pele lateralmente, da região cervical até o nível caudal toracolombar, possibilitando a retirada das vísceras em bloco. Na sequência, foram desarticulados do tronco, a cintura pelvina, o estilopódio e o zeugopódio, que por sua vez passaram por descarne prévio, bem como, foram submetidos ao processo de maceração com fervura, e submersão em solução de peróxido de hidrogênio a 20 volumes. Uma vez atingida a clarificação satisfatória foram identificados e descritos os acidentes ósseos (Brandão, Iamaguti & Figueiredo, 2002; Silveira, Teixeira & Oliveira 2008). Os registros fotográficos foram feitos com câmera D300S.
Os termos anatômicos e direcionais utilizados foram embasados na literatura consultada: P. gracilis (Gauthier et al. 2011); H. foxii (Hulke 2013); L. chanarensis (Fechner
2009); M. niger (Vieira 2011); P. Chiniquensis (Liparini 2011); Rauisuchia (Raugust 2014), e A. mississipiensis (Blob 2000).
RESULTADOS Cintura pelvina
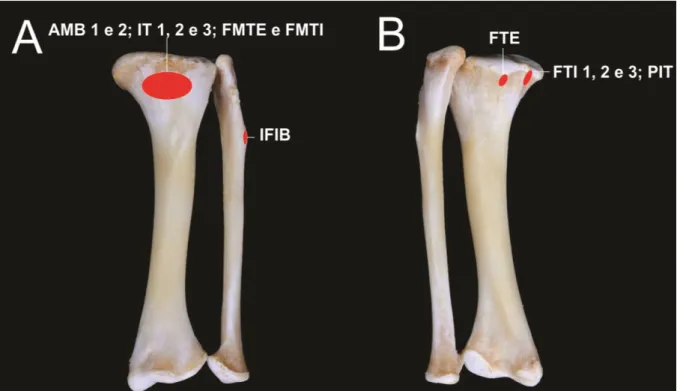
A cintura pelvina do C. crocodilus (figura 1A), é constituída por três ossos, em cada
antímero: ílio, ísquio e pube.
Ílio
O ílio do C. crocodilus (figura 1B, C e D) com localização dorsolateral na cintura
pelvina, é constituído por um corpo, com dois processos lateralizados. O processo isquiádico
cranial é mais curto, com uma área rugosa para fixação do ligamento acetabular. O processo isquiádico caudal é mais amplo, com uma face articular trilátera. Os dois processos são entremeados por uma incisura, que contribui para a formação da incisura acetabular.
Em vista dorsal do ílio, é observada uma ampla extensão laminar horizontalizada, reconhecida como asa ilíaca, com uma profundidade, identificada como fossa ilíaca, que por sua vez, possui um processo cranial curto, e um processo caudal alongado e sinuoso, lateralmente.
proeminente e alongada, e uma crista supra acetabular limítrofe, entre uma fossa acetabular, e uma fossa caudal mais rasa.
Ísquio
O ísquio do C. crocodilus (figura 2B, C, D e E) tem localização ventrolateral e caudal
na cintura pelvina.
Em vista lateral, observa-se um corpo com um processo cranial com uma face articular que contata o pube, e uma área rugosa para inserção do ligamento acetabular, que se fixa no processo isquiádico cranial do ílio. O processo ilíaco caudal é mais amplo, e trilátero. Os processos cranial e caudal são entremeados por uma incisura, que contribui para a formação do acetábulo e da incisura acetabular.
Em projeção intermediária observa-se um ramo alongado e afilado. Trata-se de uma extensão plana em vista dorsal, e em vista ventral, observa-se uma linha caudal para fixação de origem do músculo adutor 2.
Na extensão caudal do ísquio observa-se uma tábula ampla, com uma face articular isquiádica medial, e uma margem caudal com um tuber isquiádico afilado.
Pube
O pube do C. crocodilus (figura 3B, C e D) com localização ventral e cranial na
cintura pelvina, possui um corpo, que corresponde à superfície articular sinfisial, na margem medial, uma extensão em tábula cranial, e um ramo caudal. Possui uma superfície que se articula com o ísquio, formando a junção puboisquiádica. Por conseguinte, o pube não contribui para a formação do acetábulo.
Em localização dorsolateral, observa-se um processo para fixação de origem do músculo Ambiens 1, que possui ação extensora na coxa. E em extensão dorsal craniocaudal há uma crista para fixação de origem do músculo puboisquiofemoral externo 2, que realiza ação adutora da coxa.
Membro pelvino Fêmur
O fêmur do C. crocodilus (figura 5), ou estilopódio, possui uma epífise proximal, que
se articula com o acetábulo, uma epífise distal, que estabelece junção com a tíbia e fíbula, e uma diáfise, que conecta as duas epífises.
Na epífise proximal é observada uma cabeça alongada e elíptica, afilada lateralmente, que se articula com o acetábulo, para formar a articulação do quadril. Proximal à cabeça do fêmur observa-se o colo anatômico, mais acentuado em extensão lateral. Lateralmente, observa-se o trocanter maior, e em localização medial, o trocanter menor.
A diáfise do fêmur é sinuosa, convexa em extensão dorsal, e predominantemente côncava ventralmente. Em vista ventral e proximal observa-se uma proeminência afilada e irregular, identificada como o quarto trocanter, com uma fossa trocantérica bem definida. Entre o quarto trocanter e o trocanter menor, é identificada a crista intertrontérica, bem como, em localização dorsal, entre o trocanter maior e o trocanter menor, a linha intertrocantérica. Uma linha áspera é identificada na diáfise, em localização dorsal, extensiva obliquamente, que serve como local de fixação de inserção do músculo iliofemoral. Ventralmente, observa-se uma linha para fixação de inobserva-serção dos músculos adutores.
articular condilar medial da tíbia, entretanto, a superfície articular condilar lateral do fêmur, contata a superfície articular condilar lateral da tíbia, também como, a cabeça da fíbula.
Tíbia
A tíbia do C. crocodilus (figura 6), tem localização medial, representando o
constituinte de maior diâmetro do zeugopódio, em relação à fíbula lateralmente. Possui junção proximal com o fêmur e a fíbula, e distal com o tarso.
A epífise proximal da tíbia apresenta uma superfície articular trilátera, com um côndilo medial, um côndilo lateral, e um processo dorsal. Entre os côndilos observa-se uma área intercondilar com um sulco central, delimitando a superfície articular medial e lateral. Tais superfícies se articulam com os côndilos medial e lateral do fêmur. Lateralmente é identificada uma face articular fibular, formando a articulação tibiofibular proximal.
Em vista ventral e proximal, a tíbia possui uma área irregular e rugosa, identificada como tuberosidade da tíbia, com um pequeno sulco evidenciado.
A epífise distal da tíbia possui um maléolo lateral, com um sulco maleolar e uma pequena incisura fibular. O maléolo lateral se articula o maléolo medial da fíbula, formando a articulação tibiofibular distal, bem como, caudalmente, se articula com o tarso.
Fíbula
A fíbula do C. crocodilus (figura 6) tem localização lateral, representando o
constituinte de menor espessura do zeugopódio, em relação à tíbia.
Na epífise proximal da fíbula, observa-se uma cabeça com um ápice dorsal, um colo, uma face articular femoral, e uma face articular tibial.
Na diáfise, em localização proximal destaca-se uma área de inserção do músculo iliofibular.
A epífise distal da fíbula possui um maléolo medial, com uma face articular maleolar, esta, se articula com a tíbia, bem como, uma face articular distal que contata o tarso.
DISCUSSÃO
No presente estudo, o ílio foi identificado possuindo uma asa, com um processo cranial curto, e um processo caudal alongado e sinuoso lateralmente. Trata-se de resultado da rotação anti horária dos ossos e músculos, que neles se inserem, em répteis referidos como semelhantes a mamíferos. Nestes, o crescimento primitivo ocorreu primeiro posteriormente, e logo em sentido anterior, até alcançar o sacro (Romer & Parsons 1985). O processo cranial curto da asa do ílio foi observado menor em P. Chiniquensis (Liparini 2011), em Rauisuchia (Raugust
2014), e em anfíbios fósseis primitivos (Romer & Parsons 1985). Entretanto, em H. foxii
(Hulke 1882), o processo cranial da asa do ílio foi identificado alongado e afilado, diferindo, totalmente, das demais espécies utilizadas para comparação. Uma maior extensão pré acetabular do ílio, que corresponde ao processo cranial da asa ilíaca, em detrimento a projeção curta pós acetabular, que corresponde ao processo caudal da asa ilíaca, é um dos indicadores de postura predominantemente aduzida. O oposto, sinaliza maiores possibilidades de postura abduzida (Figura 9) (Oliveira 2010). O processo isquiádico caudal amplo, do corpo do ílio, indica o suporte de peso do animal, que é transferido principalmente para esta área (Fechner 2009, Liparini 2011). Em relação ao diâmetro da asa ilíaca, o mesmo é observado maior nos animais quadrúpedes, como nos Crocodylia viventes, em comparação com os bípedes ancestrais (Maidment & Barrett, 2014).
O ísquio foi observado em C. crocodilus com um ramo alongado, afilado, e uma
tábula com maior extensão, com uma margem caudal com um túber afilada, esta, observada em M. niger (Vieira 2011), arredondada, e com um ramo curto. Em L. chanarensis (Fechner
com um ramo alongado. Em P. gracilis (Gauthier et al. 2011), em Rauisuchia (Raugust 2014)
e em P. Chiniquensis (Liparini 2011), o ísquio apresenta dois processos, um cranial mais
amplo, em relação ao caudal, mais estreito, com uma extensão intermediária afilada, alongada e laminar. Aspecto este, que difere no H. foxii (Hulke 1882), descrito com um processo
cranial alongado acentuado e afilado, bem como, a extensão caudal curta e circular.
Diferente do que foi observado em C. crocodilus, neste estudo, o pube, em P. gracilis
(Gauthier et al. 2011), tem formato retangular. Em H. foxii (Hulke 1882) os processos cranial
e caudal são afilados, sendo o caudal mais alongado e afilado, em relação ao cranial. L. chanarensis (Fechner 2009) apresenta um processo cranial estreito e alongado em toda a sua
projeção até o acetábulo, com um processo caudal curto e triangular. Em P. Chiniquensis
(Liparini 2011) e em Rauisuchia (Raugust 2014) observa-se um processo cranial mais amplo,
em relação ao caudal, que possui um forame identificado como obturatório. Os processos cranial e caudal são entremeados por uma extensão intermediária alongada, afilada e plana.
Em relação a não contribuição do pube na constituição do acetábulo, como identificado em C. crocodilus, em A. mississipiensis (Blob 2000), em M. niger (Vieira 2011),
e em Cayman yacare (Lima et al., 2010), este, descrito em bases gerais, é identificado o
oposto em P. gracilis (Gauthier et al. 2011), H. foxii (Hulke 1882), L. chanarensis (Fechner
2009), P. Chiniquensis (Liparini 2011), e em Rauisuchia (Raugust 2014). Nos Crocodylia, o
acetábulo é constituído pela articulação do processo isquiádico caudal do ílio, com o processo ilíaco do ísquio, e por um ligamento interposto ao processo isquiádico cranial do ílio, e o processo cranial do corpo do ísquio (Vieira 2011).
Os Crocodylia são referidos como os únicos, entre os tetrápodes viventes, com a não participação do pube na constituição do acetábulo, formando uma articulação móvel com o ísquio. Trata-se de atividade muscular hipoaxial associada aos movimentos da gastrália, que ocasionam rotação ventral do pube (figura 7). Esta ação possibilita expansão da cavidade abdominal, e espaço para o deslocamento caudal visceral, durante a inspiração (Carrier & Farmer 2000a). Conjetura-se que a principal vantagem da mobilidade púbica seja facilitar a respiração diafragmática. Na sua ausência, a contração do diafragma tenderia a pressurizar o conteúdo abdominal com deslocamento cranial, e, isto, resultaria em dificultar o retorno venoso do membro pelvino e cauda, prejudicando a função locomotora. Tem sido considerado, que o diafragma nos crocodilomorfos basais tivesse capacidade especializada de sustentação para atividade vigorosa, que evoluiu para uma condição metabólica aeróbia restrita, em razão da transição de hábitos terrestres, para um estilo de vida aquática (Carrier & Farmer 2000b). A disposição do pube e ísquio em tetrápodes não aviários, sinaliza adaptações dos arcossauros basais, que além de ajustes locomotores, facilita a respiração diagramática, considerando a representatividade do auxílio, pós evolutivo, da postura parassagital dos membros (Carrier & Farmer 2000c).
Tendo em vista o processo evolutivo, ao contrário das expansões, primeiro caudal, e logo, cranial do ílio, o pube e o ísquio se movimentaram em sentido caudal, de tal modo, que a placa ventral da cintura pelvina, estendeu-se em sentido cranial, para alcançar o acetábulo acetábulo, corroborando com a postura intermediária, em abdução, predominante nos Crocodylia viventes (Romer & Parsons 1985). Em condições contrárias, em que a placa ventral da cintura pelvina faz rotação caudal, como nos mamíferos e aves, há indicadores de postura predominantemente em adução, associada a maior eficiência muscular, em movimentos de protração, retração (Oliveira 2010).
Em C. crocodilus, observado nesta pesquisa, similar aos vertebrados terrestres
zeugopódio, ao contrário da fíbula, que sustenta pouco peso do corpo (Romer & Parsons 1985).
Em P. gracilis (Gauthier et al. 2011), H. foxii (Hulke 1882), L. chanarensis (Fechner
2009), P. Chiniquensis (Liparini 2011), e Rauisuchia (Raugust 2014), o quarto trocanter, por
vezes referido como trocanter interno, é mais proeminente em relação ao C. crocodilus, A. mississipiensis (Blob 2000) e M. niger (Vieira 2011). Trata-se de uma característica, que
tende a diminuir com o processo evolutivo, e que desaparece nos mamíferos (Romer & Parsons 1985).
A superfície articular proximal da tíbia, identificada neste estudo, em dimensão trilátera, em M. niger (Vieira 2011) foi descrita como retangular.
Na condição de resposta ao estresse exercido sobre o fêmur e a tíbia, principalmente por torção, considera-se maiores possibilidades de assumir posturas diversas, especificamente, nos Crocodylia, postura ereta, intermediária, e rastejante, sendo a intermediária atualmente predominante (Figura 6) (Liparini 2011). Por conseguinte, a diáfise sinuosa do fêmur, similar nos Crocodylia, e em seus ancestrais, indica uma melhor adaptação à torção descrita nos arcossauros, acompanhada de rotação medial na postura parassagital.
Em postura rastejante (figura 8) o acetábulo amplo e raso, conforme observado em C. crocodilus, e seus similares viventes, é indicador de maiores amplitudes femorais de
protração, retração, adução, abdução, bem como, as rotações, estas, consideradas essenciais nesta postura (Parrish 1986). Nos arcossauros, conforme observado neste estudo, e nas espécies utilizadas para comparação, as amplas faces articulares condilares femorais, com orientação transversa, são indicadas como responsáveis pela maior amplitude de movimento do eixo longo do fêmur, em detrimento à cabeça do fêmur, que em postura rastejante, pouco se desloca, concentrando as forças recorrentes medialmente (Blob 2001).
Em postura ereta (figura 8) o eixo femoral é reto em vista cranial, indicando que as forças de torção são minimizadas, e que, nesta condição, a resistência contra flexão médio lateral torna-se reforçada (Figura 11) (Fechner 2009). A limitação de deslocamento da cabeça do fêmur, ocorre, principalmente, por mediação da crista acetabular. Restrições, também, acometem a amplitude de movimento do eixo longo do fêmur, que passa a priorizar o ajuste da posição corporal, e não maior amplitude, conforme ocorre na postura rastejante. Nesta situação as forças recorrentes ficam concentradas dorsalmente, com declínio da amplitude dos movimentos femorais. (Parrish 1986).
Em postura intermediária (figura 8) indicada como predominante nos Crocodylia atuais, ocorre redução de amplitude rotacional, principalmente lateral, em razão de restrição locomotora nos ajustes articulares dos segmentos distais dos membros pelvinos (Fechner 2009).
CONCLUSÕES
A cintura pelvina do C. crocodilus é similar à dos saurísquios, com exceção da não
contribuição do pube, na formação do acetábulo, em razão da expansão do ísquio, tendo em vista o processo evolutivo, no que se refere aos padrões ancestrais bípedes.
O C. crocodilus apresenta maior similaridade com os arcossauros viventes, tendo em
vista o comportamento locomotor que define e explica, em grande parte, as adaptações ao processo evolutivo, bem como, a influência dos hábitos ancestrais terrestres, e atualmente, os Crocodylia, semi-aquáticos.
REFERÊNCIAS
Brandão C. V. S., Iamaguti P. & Figueiredo L. M. A. 2002. Substituição do ligamento da cabeça do fêmur com auto-enxerto de fáscia lata na luxação coxofemoral em cães. Cienc. Rural, Santa Maria. 2:275-280.
Brazaitis P., C. Yamashita & G. Rebelo. 1998. The caiman trade. Scient. Americ. New York.
278:70-76.
Blob R. W. 2000. Interspeci®c scaling of the hindlimb skeleton in lizards, crocodilians, felids and canids: does limb bone shape correlate with limb posture? J. Zool., Lond. 250:507-31. Blob, R. W. & A.A. Biewener. 2001. Mechanics of limb bone loading during terrestrial
Locomotion in the Green Iguana (Iguana iguana) and American Alligator (Alligator mississippiensis). The Journal of Experimental Biology. 204:1099.
Farmer C. F. & Carrier D. R. 2000a. The evolution of pelvic aspiration in archosaurs. Paleobiol. 26:271-293
Farmer C. F. & Carrier D. R. 2000b. Pelvic aspiration in the American alligator (Alligator mississippiensis). Journal of Experimental Biology. 203:1679–1687.
Farmer C. F. & Carrier D. R. 2000c. The Integration of Ventilation and Locomotion in Archosaurs. Amer. Zool., 40:87–100.
Fechner R. 2009. Morphofunctional Evolution of the Pelvic Girdle and Hindlimb of Dinosauromorpha on the Lineage to Sauropoda. 2009. 211f. Dissertation zur Erlangung des Doktorgrades in den Naturwissenschaften. Ludwigs Maximilians niversität, München by Dipl.-geol.
Gauthier J. A., Nesbitt, S.J., Schachner, E. R., Bever, G. B. & Joyce, W.G. 2011. Bulletin of the Peabody Museum of Natural History 52:107–126.
Hulke J.W. 1882. An Attempt at a Complete Osteology of Hypsilophodon Foxii; a British Wealden Dinosaur. Philosophical Transactions of the Royal Society of London, 173:1035-1062.
Hutchinson J. R. & Gatesy, S. M. 2000. Adductors, abductors, and the evolution of rchosaur locomotion. Paleobiology26: 734–751.
Hutchinson J. R. & S. M. Gatesy. 2006. Beyond the bones. Nature. 440:292-294.
Lima F. C. 2010. Desenvolvimento do esqueleto em embriões de Caiman yacare (Daudin
1802) (Crocodylia, Alligatoridae). 123f. Dissertação de mestrado em Saúde Animal. Universidade Federal de Uberlândia, Uberlândia.
Lipari A. 2011. Aspectos biomecânicos e morfofuncionais do esqueleto apendicular de
Prestosuchus Chiniquensis (Archosauria: Pseudosuchia) e suas implicações para
locomoção. 2011. 283f. Tese (Doutorado em Geociências, para título de Doutor em Ciências) Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Oliveira T.V. 2010. Postura e locomoção em cinodontes do Triássico Sul Americano: um estudo de caso baseado em Trucidocynodon riograndendesis Oliveira, Soares e Shutz, Triássico Superior, Formação Santa Maria, Bcia do Paraná, Rio Grande do Sul, Brasil. 2010. 223f. Tese de Doutorado em Ciências, Instituto de Geociências. Universidade Federal do Rio Grande do Sul, Porto Alegre.
Otero A., Gallina, P.A. & Herrera Y. 2010. Pelvic musculature and function of Caiman latirostris. J. Herpetol. 20: 173–184.
Parrish, J. M. 1986. Locomotor adaptations in the hindlimb and the pelvis of the Thecodontia. Hunteria: 1:1-36.
Pough F. H., Janis C. M. & J. B. Heiser 2006. A vida dos vertebrados. 4. ed. São Paulo: Atheneu.
Raugust T. 2014. Descrição osteológica e análise filogenética de um novo material de
Grandense, Brasil. 2014. 383f. Tese de Doutorado em Ciências, Instituto de Geociências. Universidade Federal do Rio Grande do Sul, Porto Alegre.
Reese A. M. 1915. The Alligator and Its Allies. New York: G.P. Putnam.
Romer A. S. 1923. Crocodilian pelvic muscles and their avian and reptilian homologues. Bulletin of the American Museum of Natural History. 48: 533–551.
Romer S. R. & Parsons T. S. 1985. Anatomia comparada dos vertebrados. São Paulo: Atheneu, 1985.
Rowe T. 1986. Homology and evolution of the deep dorsal thigh musculature in birds and other Reptilia. J. Morphol.189:327–346.
Silveira M. J., Teixeira G. M. & Oliveira E. F. 2008. Análise de processos alternativos na preparação de esqueletos para uso didático. Acta Sci. Biol. Sci. Maringá. 4:465-472.
Stannius H. 1854. Handbuch der Anatomie der Wirbelthiere. Berlin: Veit.
Vieira L. G. 2011. Desenvolvimento embrionário de Melanosuchus niger (Crocodylia,
Alligatoridae): descrição de estágios e ontogenia do esqueleto. 2011. 184 f. Tese de Doutorado em Ciências Biológicas. Instituto de Ciências Biológicas Programa de Pós-Graduação em Biologia Animal, Universidade de Brasília, Brasília.
FIGURAS
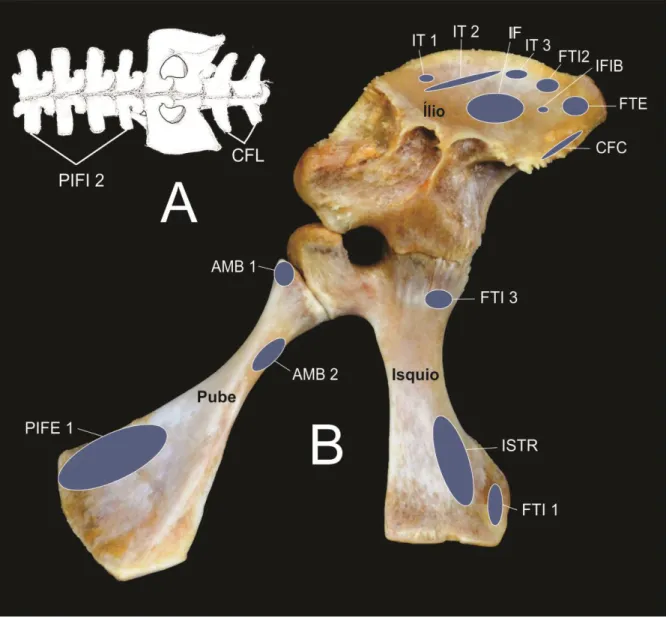
Figura 1. Fotografia da cintura pelvina do antímero direito, em vista lateral (A) e Ilio em vista lateral (B), dorsal (C) e medial (D). A - CA, Crista Acetabular; FA, Fossa Acetabular; FeA, Fenestra Acetabular; Is, Isquio; IL, ílio; Pu, Pube. B - AI, Asa Ilíaca, CI, Crista Ilíaca; CA, Crista Acetabular; FA, Fossa Acetabular; F Is, Face Isquiádica; FLA, Face para fixação do Ligamento Acetabular; PIsCa, Processo Isquiádico Caudal; PIsCr, Processo Isquiádico Cranial. C - FI, Fossa Ilíaca; In I, Incisura ilíaca; PAICa, Processo da Asa Ilíaca Caudal; PAICr, Processo da Asa Ilíaca Cranial; D – FC 1, Face Articular Costal 1; FC2, Face articular Costal 2.
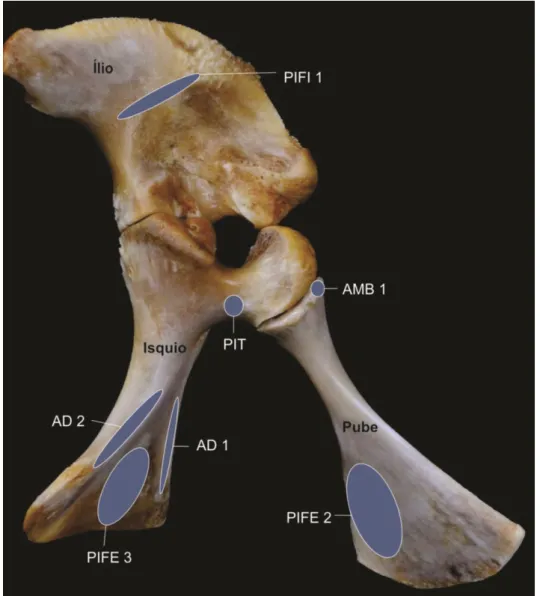
Figura 3. Fotografia da cintura pelvina do antímero direito, em vista lateral (A) e do pube em vista dorsal (B), ventral (C), e caudal (D). A - CA, Crista Acetabular; FA, Fossa Acetabular; FeA, Fenestra Acetabular; Is, Isquio; IL, ílio; Pu, Pube. B– CPIFE 2, Crista para fixação de origem do músculo Puboisquiofemoral Externo 2; FS, Face Sinfisial; P Am1, Processo para fixação de origem do músculo Ambiens 1; RP, Ramo do Pube; TaP, Tábula do Pube. C - TaP, Tábula do Pube. D – F Is, Face articular isquiádica.
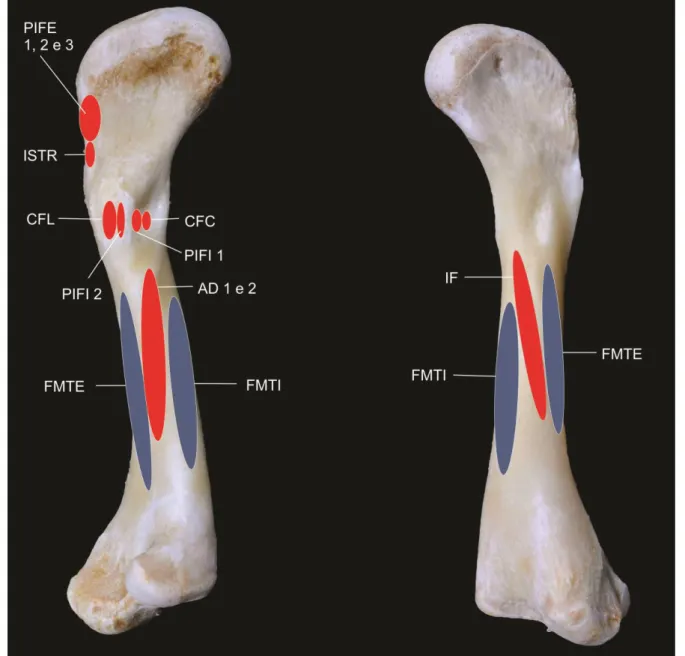
Figura 5. Fotografia da tíbia e fíbula do antímero direito em vista Cranial (A), ventral(B) e dorsal(C). A – SIc, Sulco Intercondilar; B – CaF, Cabeça da Fíbula; CoF, Colo da Fíbula; CM, Côndilo Medial; CL, Côndilo Lateral; I If, Inserção do músculo Iliofibular; ML, Maléolo Lateral; MM, Maléolo Medial; TB, Tuberosidade da Tíbia. C – ACaF, Ápice da Cabeça da Fíbula; ATFP, Articulação Tibiofibular Proximal; ATFD Articulação Tibiofibular Distal; PD, Processo Dorsal.
Figura 7. Ilustração representativa dos músculos pelvinos e abdominais do Alligator mississipiensis (Farmer & Carrier 2000b). A, vista ventral da cintura pelvina e gastralia; B,
Vista ventral: músculo Rectus abdominis e Ischiocaudalis; C, Músculo Ischiopubis; D, Ilustra
a relação entre os músculos Truncocaudalis e o músculo Ischiotruncus.
Capítulo 3 - Anatomia muscular da cintura pelvina e estilopódio de Caiman crocodilus3
Mariluce F. Romão4*, André L. Q. Santos4
ABSTRACT. Mariluce F. Romão, André L. Q. Santos [Muscular Anatomy of the Pelvic Girdle and Stylopod of the Caiman crocodilus]. Anatomia Muscular da cintura pelvina e
estilopódio do Caiman crocodilus. Pesquisa Veterinária Brasileira 00(0):00-00. Laboratório
de Ensino e Pesquisa em Animais Silvestres, Faculdade de Medicina Veterinária, Universidade Federal de Uberlândia, UFU. Rua Piauí, bloco 4s, bairro Jardim Umuarama, Uberlândia MG, 38400-902 Brasil. E-mail: mariluce.ferreira@terra.br.
The objective was to study the muscular anatomy of the pelvic girdle and stylopod of the Caiman crocodilus, augmented of kinesiological correlations, and homology in some
groups of tetrapods, under the assumption that such information can contribute to improve the knowledge about the locomotor system of these animals. Four samples of the C. crocodilus,
set in 10% formalin, males, measuring an average length of 1,50m, adults belonging to the didactic collection of the Laboratory of Education and Research of Wild Animals (Laboratório de Ensino e Pesquisa em Animais Silvestres), from the Federal University of
Uberlândia (Universidade Federal de Uberlândia), were used. The dissection was performed,
initiating with a longitudinal incision, along the ventral median line of the trunk, folding the skin laterally, until the tail. Next, the muscles of the pelvic girdle and stylopod of the C. crocodilus were individualized and identified, with verification of the respective muscular
origins and insertions. In a lateral and superficial view, there was the identification of the internal puboischiofemoral muscle 2, iliotibial group (heads 1, 2 and 3), and iliofibular muscle. In a deep lateral view (Table 2) the external femorotibial and iliofemoral muscles were identified. In a superficial and medial view, there was the identification of the ambiens group (head 1 and 2), internal puboischiofemoral muscle 1, external puboischiofemoral group (heads 1, 2 and 3), femoral adductor muscle 1, puboischiotibial, internal tibial flexor (heads 1 and 2), external tibial flexor, and caudofemoral group (long and short). In a deep medial view, the internal puboischiofemoral muscle 1, femoral adductor 2, internal tibial flexor 3, and ischiotrochanteric muscles were identified. The muscular anatomy of the pelvic girdle and stylopod of the Caiman crocodilus confirms the morphological diversity, in terms of greater
responsibility directed to functional activities, for presenting differences, including with the living Crocodylia. The external pubioischiofemoral group was considered the most similar in the comparisons, and the internal tibial flexor group, the most diverse. Functional adaptations due to evolution happen, with tendency to minimize and redirection functions.
INDEX TERMS: Stylopod, spectacled caiman, muscles.
RESUMO.-Objetivou-se estudar estudar a anatomia muscular da cintura pelvina e estilopódio do Caiman crocodilus, acrescido de correlações cinesiológicas, e homologia em alguns
grupos de tetrápodes, partindo do pressuposto, que tais informações podem contribuir para aprimorar o conhecimento acerca do aparelho locomotor destes animais. Foram utilizados quatro exemplares de C. crocodilus, fixados em formol a 10%, machos, medindo em média
1,50 m de comprimento, adultos, pertencentes ao acervo didático do Laboratório de Ensino e
3 Recebido em... Aceito para publicação em...
4 Laboratório de Ensino e Pesquisa em Animais Silvestres, Faculdade de Medicina Veterinária, Universidade
Pesquisa em Animais Silvestres, da Universidade Federal de Uberlândia. Foi realizada a dissecação, iniciada com um uma incisão longitudinal, ao longo da linha mediana ventral do tronco, rebatendo-se a pele lateralmente, até a cauda. Na sequência, foram individualizados e identificados os músculos da cintura pelvina, e estilopódio do C. crocodilus, com sequente
verificação de suas respectivas origens e inserções musculares. Em vista lateral e superficial foram identificados os músculos puboisquiofemoral interno 2, grupo iliotibial (cabeças 1, 2 e 3), e músculo iliofibular. Em vista lateral e profunda (Quadro 2) foram identificados os músculos femorotibial externo e iliofemoral. Em vista medial e superficial foram identificados o grupo ambiens (cabeça 1 e 2), músculo puboisquiofemoral interno 1, grupo puboisquiofemoral externo (cabeças 1, 2 e 3), músculo adutor femoral 1, puboisquiotibial, flexor tibial interno (cabeças 1 e 2), flexor tibial externo, e grupo caudofemoral (longo e curto). Em vista medial e profunda foram identificados o músculo puboisquiofemoral interno 1, adutor femoral 2, flexor tibial interno 3, e isquiotrocantérico. A anatomia muscular da cintura pelvina e estilopódio do Caiman crocodilus confirma a diversidade morfológica, no
que refere à maior responsabilidade direcionada para as atividades funcionais, por apresentar diferenças, inclusive com os Crocodylia viventes. O grupo puboisquiofemoral externo foi considerado com maior similaridade entre as comparações, e o grupo flexor tibial interno, o mais diverso. Adaptações funcionais devido à evolução acontecem, com tendência a minimizar e redirecionar funções
TERMOS DE INDEXAÇÃO: Estilopódio, jacaré tinga, músculos.
INTRODUÇÃO
Os répteis descritos como dominantes no período mesozoico foram os dinossauros, subdivididos nas ordens: Saurísquios, semelhantes a répteis ou lagartos, e Ornitísquios, semelhantes às aves. A ordem Crocodylia, na qual estão inseridos os jacarés, crocodilos e gaviais, faz referência a um grupo restrito, que tem as suas origens nos arcossauros primitivos (Carrol 1969, Walker 1972; Orr 1986; Storer et al. 2005).
Inicialmente terrestres, com evolução para animais semi-aquáticos, os Crocodylia permaneceram com membros bem desenvolvidos, mantendo a possibilidade de realização de movimentos amplos em terra, o que nos seus primórdios, já lhes permitia a caça ativa de diapsidas menores. Trata-se de animais, que podem galopar, e que a partir da postura normal, com os membros lateralizados e em extensão, assumem uma postura dos mesmos, quase vertical, sob o corpo, realizando uma série de movimentos essenciais (Reilly et. al. 2004; Pough, Janis & Heiser 2006; Britton 2009).
A função do membro do animal deve atender à variação mecânica tendo em vista as demandas de locomoção. Além de gerar a força para o suporte de peso corporal, os músculos também devem produzir ou absorver energia, quando os animais se movem sobre variados graus, considerando velocidade, saltos ou qualquer outra forma de deslocamento (Daley & Biewener 2003).
O Caiman crocodilus, (Linnaeus, 1758) (Crocodylia: Alligatoridae), conhecido como
jacaré tinga, é um crocodiliano de porte médio, chegando a medir, em média dois metros e meio, quando adulto. Pode ser encontrado no Sul do México, América Central, América do Sul, distinguido pelo corpo esverdeado com manchas amarelas e cinza entremeadas (Brazaitis et al. 1998, Villela 2008).
No presente estudo, objetivou-se estudar a anatomia muscular da cintura pelvina e estilopódio do Caiman crocodilus, acrescido de correlações cinesiológicas, e homologia em
alguns grupos de tetrápodes, partindo do pressuposto, que tais informações podem contribuir para aprimorar o conhecimento acerca do aparelho locomotor destes animais (Stannius 1854, Rowe 1986, Wilhite 2003).
MATERIAL E MÉTODOS
Foram utilizados quatro exemplares de C. crocodilus, machos, medindo em média 1,50 m de
comprimento, adultos, pertencentes ao acervo didático do Laboratório de Ensino e Pesquisa em Animais Silvestres (LAPAS), da Universidade Federal de Uberlândia, fixados em formol a 10%. Os animais são referentes ao SISBIO nº. 13159-1, e com parecer de ética aprovado pela CEUA/UNIPAM.
Foi realizada a dissecação, iniciada com um uma incisão longitudinal, ao longo da linha mediana ventral do tronco, rebatendo-se a pele lateralmente, até a cauda. Os quatro exemplares de C. crocodilus foram mantidos fixados em solução de formol a 10%.
Na sequência, foi feita a individualização muscular da cintura pelvina, e membro pelvino do C. crocodilus, com sequente identificação dos músculos, bem como, das origens e
inserções musculares, e, por último foram fotografados com câmera D300S.
Os termos direcionais utilizados e as ações musculares, foram embasados no estudo de Otero et al. (2010), em descrição da cintura pelvina e membro pelvino do Caiman latirostris.
RESULTADOS
Em descrição textual apresenta-se a topografia muscular da cintura pelvina e estilopódio do C, crocodilus, e nos quadros subsequentes 1, 2, 3 e 4, são apresentadas origem, inserção e ação
sugerida (Otero et al. 2010) dos músculos da cintura pelvina e estilopódio do C. crocodilus.
As figuras 1, 2, 3, 4, apresentam as origens e inserções musculares da cintura e estilopódio do
C. crocodilus, e as figuras 5 e 6 demonstram a topografia muscular.
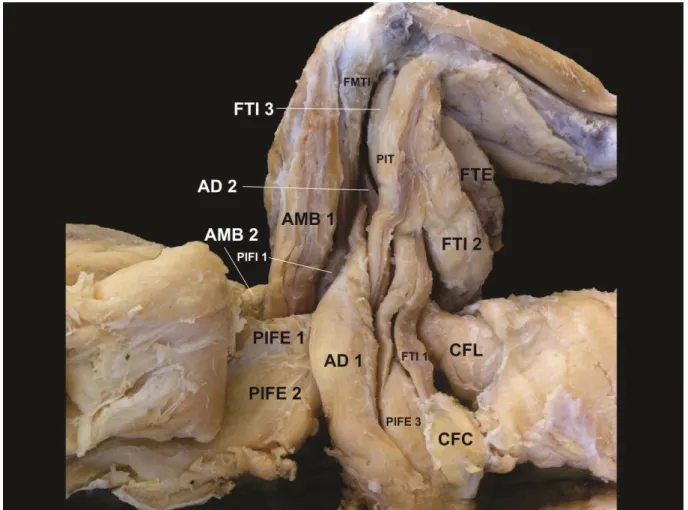
Em vista lateral e superficial (Quadro 1) foram identificados os músculos puboisquiofemoral interno 2, grupo iliotibial (cabeças 1, 2 e 3), e músculo iliofibular.
O músculo puboisquiofemoral 2 tem formato triangular, é amplo e localiza-se cranial em relação à cabeça 1 do grupo iliotibial.
O grupo iliotibial possui três cabeças. A cabeça 1, a menor do grupo, tem localização cranial, entremeando fibras tendíneas, distalmente, com a cabeça 2 do mesmo grupo. A cabeça 2, tem localização média, e é a maior do grupo, em comprimento e diâmetro. A cabeça 3 tem localização caudal, é longa e estreita.
O músculo iliofibular foi identificado em localização proximal, caudal ao iliotibial 2, com um curto percurso profundo ao iliotibial 3, contínuo distalmente à última cabeça citada, e cranial ao flexor tibial externo.
Em vista lateral e profunda (Quadro 2) foram identificados os músculos femorotibial externo e iliofemoral.
O músculo femorotibial externo tem localização profunda em relação às cabeças 2 e 3 do grupo iliotibial, com fixação de origem ao longo da diáfise femoral, lateralmente, nas faces ventral e dorsal.
O músculo iliofemoral tem localização profunda em relação às cabeças 2 e 3 do grupo iliotibial, e localização média, em relação às origens dos músculos femorotibial externo lateralmente, e interno medialmente.
O grupo ambiens foi identificado com duas cabeças. A cabeça 1, a maior do grupo, localiza-se medialmente ao longo da diáfise do fêmur, enquanto a cabeça 2, menor e mais estreita, é perpendicular e profunda distalmente à cabeça 1 do mesmo grupo.
O músculo puboisquiofemoral interno 1 é estreito e curto, com localização profunda, entre os músculos ambiens 1 e adutor 1.
O grupo puboisquiofemoral externo foi identificado com três cabeças. A cabeça 1 entremea ventre muscular com a cabeça 2, esta, a mais ampla do grupo. E a cabeça 3 tem localização caudal ao músculo adutor femoral 1.
O músculo adutor femoral 1 foi identificado com diâmetro amplo, maior em relação ao adutor femoral 2, e caudal às cabeças 1 e 2 do grupo puboisquiofemoral externo.
O músculo puboisquiotibial tem localização caudal ao músculo adutor 1 e cranial à cabeça 1 do grupo flexor tibial interno 1. Em extensão distal o puboisquiotibial se une ao grupo flexor tibial interno para inserção comum na epífise proximal da tíbia, medial ao processo dorsal.
O músculo flexor tibial interno 1 tem localização distal ao caudofemoral curto, caudal à cabeça 3 do grupo puboisquiofemoral externo, e puboisquiotibial, e cranial ao caudofemoral longo.
O músculo flexor tibial interno 2 foi identificado caudal em relação ao flexor tibial interno 1, e cranial ao flexor tibial externo.
O músculo flexor tibial externo foi identificado caudal em relação ao flexor tibial interno 2, representando o maior músculo do grupo flexor tibial, em diâmetro.
O grupo caudofemoral foi identificado com duas cabeças. A cabeça curta, a menor do grupo, tem localização caudal à cabeça 3 do grupo puboisquiofemoral externo, e cranial ao caudofemoral longo. O músculo caudofemoral longo foi identificado caudal em relação ao caudofemoral curto, com diâmetro amplo, destacado em localização proximal.
Em vista medial e profunda (Quadro 4) foram identificados o músculo puboisquiofemoral interno 1, adutor femoral 2, flexor tibial interno 3, e isquiotrocantérico.
O músculo puboisquiofemoral interno 1 foi identificado profundo, entre o ambiens 1 e adutor femoral 1, localizados superficialmente.
O músculo adutor femoral 2 foi identificado profundo ao puboisquiotibial, entre o femorotibial interno e flexor tibial interno 3, localizados superificialmente.
O músculo flexor tibial interno 3 foi identificado profundo ao puboisquiotibial e flexor tibial interno 1.
O músculo isquiotrocantérico foi identificado com diâmetro estreito e curto, profundo ao flexor tibial interno 1, em localização proximal, cruzando medialmente a articulação do quadril.
DISCUSSÃO
No presente estudo, o grupo iliotibial (figura 1, 5) foi observado com três cabeças, como no
Caiman latirostris (Otero et al 2010), Alligator mississipienses (Romer 1923, Gatesy 1997), e Poposaurus gracilis (Wilhite 2003, Schachner et al. 2011). O m. iliotibial 1 foi identificado
no presente estudo com origem cranial, dorsolateral na asa ilíaca e inserção na tuberosidade da tíbia, no mesmo nível do m. iliotibial 2. Em A. mississipienses (Reilly et al. 2004) o m.
iliotibial 1, além de extensor e abdutor do joelho, como referido em C. crocodilus e C. latirostris (Otero et al. 2010), foi descrito como estabilizador da cintura pelvina e joelho.
Trata-se de um músculo com localização profunda ao m. iliotibial 2, diferente do que foi identificado no presente estudo e nas demais espécies utilizadas para comparação. A cabeça 1, a mais curta do grupo iliotibial, neste estudo, interdigita distalmente aponeurose com m. iliotibial 2. Tal descrição não corrobora com o mesmo músculo descrito em C. latirostris
nítida. Em A. mississipienses foram descritas duas cabeças do grupo iliotibial (Reese 1915), e
três cabeças, na mesma espécie, por Gatesy (1997). Em geral, nos Crocodylia são consideradas três cabeças constituindo o grupo iliotibial (Schachner, Manning & Dodson 2011). Nos ornitísquios a cabeça 1 do grupo iliotibial não é evidente, e as demais fixações musculares referentes a este grupo são referidas como inequívocas (Susannah & Barret 2011). Nas aves o grupo iliotibial apresenta quatro cabeças (Romer 1923).
O grupo ambiens (figura 1, 6) foi identificado, no presente estudo, com duas cabeças distintas, em relação à origem e disposição do ventre muscular, entretanto, com área de inserção comum, na tuberosidade da tíbia, e destaque para o tendão alongado da cabeça 2. O m. ambiens 1 foi observado superficial em relação à extensão distal do m. ambiens 2. Trata-se de aspecto similar à descrição da inserção do mesmo músculo, na tuberosidade da tíbia, referida também, como crista cnemial em C. latirostris (Otero et al. 2010), em A. mississipienses (Gatesy 1997), e em P. gracilis (Schachner et al. 2011). Em C. latirostris
(Otero et al. 2010) e em P. gracilis (Schachner et al. 2011) foi identificada somente uma
cabeça, que pela fixação de origem, corresponde ao m. ambiens 2 do presente estudo, sem evidências de uma segunda cabeça. Nos Crocodylia (Allen et al 2014) foram descritas duas cabeças, conforme observado em C. crocodilus, com similaridade de origem e inserção, e
disposição superficial do ventre muscular do m. ambiens 1, em relação à extensão distal do m. ambiens 2. Nos ornitísquios (Susannah & Barret 2011), o m. ambiens também foi descrito somente com uma cabeça, com origem cranial e ventral no pube, correspondendo à tábula em
C. crocodilus. Entretanto, Reese (1915) descreveu o grupo ambiens nos Crocodylia ancestrais,
com origem no ílio, e junção iliopúbica, correspondendo às origens das cabeças 1 e 2, respectivamente. Nas aves, a cabeça 1, que é a menor do grupo, tende a ser reduzida ou inexistente (Romer 1923).
O grupo flexor tibial (figura 1, 6) foi encontrado no presente estudo, constituído por quatro músculos superficiais, e um profundo. Trata-se do m. puboisquiotibial, m. flexor tibial interno 1, m. flexor tibial interno 2, m. flexor tibial interno 3 (profundo, em extensão média, ao m. puboisquiotibial), m. flexor tibial externo, sem evidências do m. flexor tibial interno 4. Distalmente, o m. puboisquiotibial, m. flexor tibial interno 1, 2 e 3, convergem para mesma inserção no côndilo medial da tíbia, em localização dorsal, enquanto o m. flexor tibial externo insere no processo dorsal da epífise proximal da tíbia. O grupo flexor tibial interno foi descrito com três cabeças em A. mississipienses (Reese 1915; Gatesy 1997). Em C. latirostris
(Otero et al. 2010) o m. flexor tibial interno 3, foi identificado em localização profunda ao m. flexor tibial interno 2, em toda a sua extensão, e com inserção por tendão comum com o flexor tibial externo, no processo dorsal da tíbia. Em P. gracilis (Schachner et al. 2011) foram
identificadas duas cabeças, e em A. mississipienses (Romer, 1923) quatro cabeças, sendo a 3 e
4 profundas ao m. flexor tibial interno 2 e flexor tibial externo. Nos Crocodylia, considera-se a possibilidade de existência de quatro cabeças dos mm. flexores tibiais internos, com probabilidade de variações individuais, podendo estar relacionadas à idade (Allen et al 2014). Conjetura-se, que quarta cabeça do grupo flexor tibial interno não tenha fixação osteológica, e que por isso, seja desconsiderado por muitos autores (Schachner et al. 2011; Susannah & Barret 2011).
Em C. crocodilus m. puboisquiotibial (figura 2, 6) foi identificado longo, com origem
ventral e cranial no processo púbico do ísquio, em área restrita, e inserção no côndilo medial da tíbia, com disposição de fibras caudal ao m. adutor femoral 1, e cranial ao m. puboisquiofemoral externo 3. Em extensão distal se une ao m. flexor tibial 2, para inserir em área comum. Em A. mississipienses (Gatesy 1997), e P. gracilis (Schachner et al. 2011), o m.
O m. caudofemoral curto (figura 1, 6) foi identificado no presente estudo com origem dorsomedial no processo caudal da asa, e inserção no quarto trocanter, em localização medial, diferente do que foi observado em A. mississipienses (Gatesy 1997), com fixação de origem
na costela 2 e vértebra sacral 2. Em C. latirostris (Otero et al. 2010) foi identificado com
origem na margem caudal do ílio e vértebras caudais. Portanto, considera-se não existir um consenso de origem do m. caudofemoral curto nos Crocodylia, tendo em vista a diversidade apresentada nas pesquisas disponíveis (Wilhite 2003). O m. caudofemoral longo, com auxílio do m. caudofemoral curto é considerado como o grupo de maior representatividade, tendo em vista a retração e rotação do eixo longo do fêmur, durante a locomoção (Reilly et al. 2005). Em P. gracilis (Schachner et al. 2011) a origem da cabeça curta do grupo caudofemoral, foi
identificado na superfície lateral do ílio. Na retração da cintura pelvina e membro pelvino, a contribuição muscular mais significativa é direcionada ao grupo caudofemoral, sendo a cabeça longa, agonista, e a cabeça curta, sinergista, com auxílio dos mm. adutores e puboisquiofemoral externo 3 (Hutchinson & Gatesy 2000).
No presente estudo foram identificadas três cabeças do grupo puboisquiofemoral externo (figura 1, 2, 6), conforme descrições gerais nos Crocodylia (Allen et al. 2014). Em C. crocodilus, a cabeça 1, interdigita fibras musculares com a cabeça 2, e a cabeça 3 foi
identificada caudal em relação ao m. adutor 1, com ação de adução e flexão da coxa, comum ao grupo (Otero et al 2010, Romer 1923, Gatesy 1997). Em A. mississipienses o mesmo grupo
muscular foi destacado em ação significativa de estabilização na fase de apoio (Reilly et. al. 2004). Em A. mississipienses (Gatesy 1997) a cabeça 3, foi encontrada profunda, entre os mm.
adutores 1 e 2, assim como, em C. latirostris (Otero et al. 2010). Em P. gracilis (Schachner et
al. 2011), foi observada, além das três cabeças, uma quarta cabeça. Nos ornitísquios, foram identificadas três cabeças, entretanto, com inserção comum no trocanter maior do fêmur (Susannah & Barret 2015).
A origem do m. puboisquiofemoral interno 1 (figura 2, 6) foi identificada em C. crocodilus na asa ilíaca em localização ventral e medial, e da cabeça 2, nos processos
transversos das vértebras lombares, bem como, inserção medial e lateral, respectivamente, em relação ao quarto trocanter. Em P. gracilis (Schachner et al. 2011), a origem da cabeça 1 foi
identificada no processo púbico do ílio, e da cabeça 2, na superfície medial do pube, com inserção comum no trocanter menor do fêmur, não havendo consenso para tais divergências, conforme o último autor citado.
O músculo isquiotrocantérico (figura 1) foi identificado no C. crocodilus com inserção
no trocanter maior, conforme em C. latirostris (Otero et al. 2010) e demais espécies utilizadas
para comparação.
Durante a fase de apoio na locomoção, em A. mississippiensis (Hutchinson & Gatesy,
2000), foi detectado controle primordial do mecanismo de adução e abdução sobre a articulação do quadril. O m. adutor 1 (quadro 3; quadro 5; figura 2; figura 3; figura 6) foi identificado como o principal na ação de adução, com auxílio do grupo puboisquiofemoral externo, m. puboisquiotibial, e m. adutor 2 (quadro 4; quadro 5; figura 2; figura 3; figura 6), bem como, o m. caudofemoral longo e m. femorotibial interno. Em fase de apoio, a postura fica quase perpendicular ao plano sagital do corpo, de maneira que os locais de inserção dos músculos, acima referidos, direcionam-se a uma ação mediolateral, resultando em adução da coxa. O principal abdutor indicado na fase de balanço foi o m. iliofemoral, com auxílio dos mm. iliotibiais 2, 3 e m. iliofibular. Em C. crocodilus no presente estudo e nas descrições em