PATTERNS OF DISTRIBUTION
IN INTERTIDAL ROCKY SHORES:
THE ROLE OF GRAZING AND COMPETITION
IN STRUCTURING COMMUNITIES
PATTERNS OF DISTRIBUTION
IN INTERTIDAL ROCKY SHORES:
THE ROLE OF GRAZING AND COMPETITION
IN STRUCTURING COMMUNITIES
Dissertação apresentada à Universidade do Algarve para obtenção do grau de Doutor
FARO
2000
I would like to thank FCT- Fundação para a Ciência e Tecnologia for funding the scholarship under which this research was done (PRAXIS XXI/BD/5721/95). To my supervisors, Prof. S.J. Hawkins and Luís Cancela da Fonseca, I would like to express my gratitude for their help and guidance throughout the period of this study. I admire them both for their incredible knowledge on a variety of ecological subjects and I was always very pleased and lucky to discuss my work with them. In particular, I would like to thank Steve for his encouragement since the beginning of this work. He has always showed a continuous interest and enthusiasm in the results, which definitely help me to carry on the work on rocky shores even ‘under’ huge waves. I also would like to thank him for providing all the facilities in the University of Southampton and, more recently, in the Marine Biological Association of the UK. Although most of the work was done in Portugal, we always kept contact ‘via email’ and he was there every time I needed, answering to any doubts and making his wise comments. Finally, I would like to thank his hospitality in Britain. Thanks for introducing me to a lot of British traditions (like cream teas, Indian and Chinese restaurants, fruit machines)! I am indebted to Luís Fonseca for accepting the supervision of this study in the University of Algarve and for his kind support during this period, and in particular, for his critical review of the various manuscripts. All the work in Portugal was done in Guia Marine Laboratory, for what I would like to specially thank the late Prof. Luiz Saldanha for accepting and providing the laboratory facilities. Thanks are also due to Prof. Pedro Ré, the actual scientific co-ordinator of the laboratory for having the patience to follow with interest the progress of this study and for always making sure that everything was going fine. All the staff members helped in a variety of occasions. Thank you to Enid, from the University of Southampton, and to Maria do Carmo, Valentina e Guilhermina, from the University of the Algarve, for always kindly answering my queries.
This work would not have been possible without the great help in the field from various people. In Britain, thanks are due to Paolo Della Santina, Matthew Alexander, Nick Smith, Malcom Roberts, Ross Coleman, Adam Jory for helping me on the shore and for monitoring the experiments during the time I was in Portugal (and I know this was a hard and boring task!). Thank you Paolo for also helping setting up the experiments in the UK and for teaching me all the Italian slang! I would also like to thank Pedro Ré, João Gil, Ricardo Beldade, João Gago, Verena, Claudia Raedig, Sevgi Yaman, Milton Alfredo, Pedro Fidalgo and Ana Colaço for helping in Portuguese rocky shores. In particular, a special thank you to Ricardo e João Gago for going so many times to Cabo Raso and Moledo do Minho and for always putting up with my stress on the shore! I am also grateful to Teresa Cruz and João Castro for kindly providing accommodation in Laboratório de Ciências do Mar in Sines during field trips to the south west of Portugal.
I am indebted to Prof. A.J. Underwood for his constructive comments and suggestions on the experimental designs of competition experiments.
I was very lucky with the friendly environment in the University of Southampton. The ‘rocky shore team’ of the Biodiversity and Ecology Division was notable. Thanks are due to Richard Thompson for all the suggestions on microbial film analysis, to Tasman Crowe for statistical advice and a critical review of the manuscript in chapter 3, and to Mark Johnson for such interesting discussions. It was also a pleasure to be in Southampton with António Múrias and Débora Borges, which were always so supportive. Thank you António for helping sorting out doubts with the experimental designs on several occasions and for many interesting discussions. Débora thanks for helping with bibliography, for ‘limpet conversation’ and for cheering me up. Thanks to all the other friends from Boldrewood Building. All of them have made my life in Southampton good fun and with loads of nights out! Thanks go also to my flat mates Doina and Andrei, Yu May and Markus for such a great time!
Pedro Fidalgo, Gil, Nani, Guca, Joao, Milton, Range, Abel, Ricardo for their support, advices and help. Thanks go also to the all lab ‘seniors’ and to past and present students for making such a nice working environment.
A special thank you goes to Pedro for his endless support throughout this work. Thanks also to him and Ana for patiently waiting for me to come back from the shore at the weekends and for so many good moments. At least I hope they enjoyed painting some limpets…
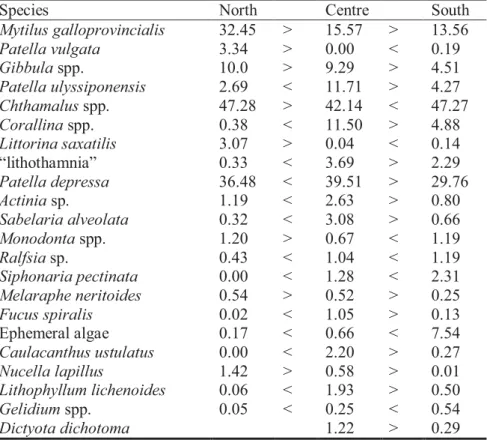
Patterns of distribution on rocky shores and the role of grazing and competition as structuring agents of intertidal communities were studied in the present work. The intertidal rocky shore communities of the Portuguese coast have been studied in a scattered way by different authors. Taking into account that Portugal is thought to be a zone of overlap of boreal Atlantic species at their southern limits and more subtropical and Mediterranean species at their northern limits, it is important to study the whole littoral coast. A general description of zonation patterns along the coast and the comparison of distribution patterns of mid-shore organisms were made in this work. In order to test if there is any variation in species distribution and abundance in upper and lower mid-shore zone along the intertidal Portuguese coast, three main regions were studied (north, centre and south). Nine shores were visited in each region and a total of 27 shores were sampled along the Portuguese coast. In each one of the 27 locations, the shore was levelled and a general qualitative description was made, which also aided stratification of subsequent quantitative sampling. Non-destructive quantitative methods were used within upper and lower mid-shore zones. Five replicate quadrats were counted in each of the two mid-shore areas using a quadrat of 50 x 50cm with 49 intersection points. Zonation patterns in Portugal may be seen as mixture of the patterns described for the north east Atlantic coasts and those described for the Mediterranean. The results of this study also confirm the latitudinal variations from north to southern Europe, with animal dominated communities extending further into shelter at more southerly locations. Similarly, southwards, dominance by large brown algae declines low on the shore and red algal turves become more important. Multidimensional analysis and the ANOSIM test have revealed clear differences in the structure of upper and lower mid-shore zone in part due to the number of species found. In general, the upper level exhibited a lower diversity and a higher dominance. Despite the decrease in abundance of several species along the regional gradient (from both North to South and South to North) the differences in the community structure were not as clear as for the vertical gradient. A possible explanation for this is that the increase in complexity of the community from high to low shore is superimposed on the latitudinal changes. Within each level, however, the northern region was considerably different from the south and central regions. SIMPER analysis revealed the species which contributed to the geographical separation of the northern region in relation to the centre and south. It has also given important information on the geographical decline in abundance of species.
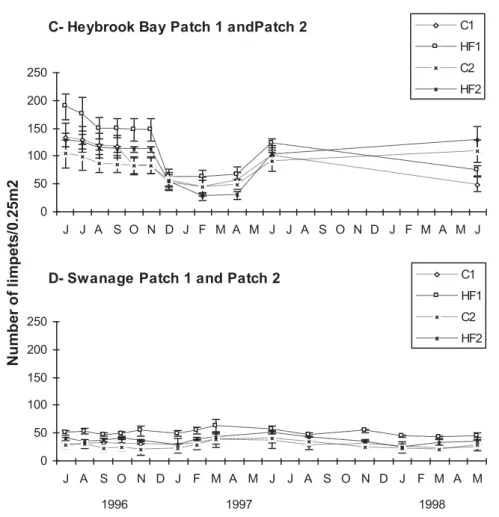
A great part of the work in this thesis was devoted to experimental ecology studies on causes of distribution patterns, essentially focussed on the importance of grazing and competition by limpets in intertidal communities. A red algal turf is often found just below the barnacle/limpet zone of many European shores, especially on steep shores of moderate exposure. The hypothesis that grazing by limpets determines the upper limit of distribution of the red algal turf was tested in Portugal and Britain. We also aimed to assess whether the grazing effect is modified by different factors operating at various spatial scales. Grazers were excluded by fences, and there were half-fenced and unfenced controls. Exclusion plots were rapidly colonised by green ephemeral algae in the months immediately after the beginning of the experiment (summer); these algae were later replaced by perennial algae. The percentage cover of turf forming macroalgae showed a significant increase at both locations. Their upper limit of distribution extended more than 50 cm in most of the shore areas. In contrast, control and half-fenced plots remained devoid of algae. After two years, ungrazed areas were mainly colonised by red algal turf (e.g. Caulacanthus ustulatus, Gelidium spp., Laurencia pinnatifida) in Portugal, while canopy cover (Fucus serratus and Himanthalia elongata) dominated in Britain. Physical factors acting at both local and geographical scales are likely to explain these differences. However, although physical factors probably have an important influence on the size and abundance of sublittoral fringe macroalgae, the results from this experiment indicate that grazers seem to play a major role in directly setting its upper limits. The community converged rapidly to undisturbed conditions after fence removal. When the algal turf was cleared beneath the “usual” limit of distribution, limpets extended down shore
higher levels on the shore and the rapidity of colonisation and growth of the algae at lower levels, being probably modified by physical factors which favour algal growth or grazing efficiency.
Inter- and intraspecific competitive interactions among the species Patella depressa and Patella vulgata were analysed on the northern coast of Portugal, where both species co-occur in similar proportions. Increased (x 2, x 4), decreased (x 1/2) and normal densities of limpets were used to test the effects of competition on the growth and mortality of the limpets. Fenced plots of 25 x 25 cm enclosing marked limpets at various densities in plastic coated wire fences (25 x 25 x 4 cm high) were set at a mid tidal level on the shore. Twelve treatments with different combination of densities and species were assigned to the plots, with three replicates of each. Mortality was recorded every fifteen days and growth was measured monthly during the five moths of the experiment. At the end of the experiment limpets were collected for biometry, sex determination and gonad stage evaluation. The availability of food was indirectly assessed by determination of chlorophyll concentration with spectrophotometric analysis of rock chips. Both species of grazing molluscs showed increased mortality and reduced growth and weight in increased density treatments. Limpets in decreased density treatments showed lower mortality and higher growth rate. Although both species could affect negatively the other, there were no significant differences between the effect of P. vulgata on P. depressa and the effect of P. depressa on P. vulgata on mortality and length. Results from tissue weight, however, revealed that intraspecific increases in density resulted in a more accentuated reduction of weight than when both species were present. The results were compared with previous studies of competition in grazing molluscs and discussed in relation to the limits of distribution of P. vulgata on the Portuguese coast. Competitive interactions within and between size-classes of the limpet Patella depressa were investigated in central Portugal. Experimental enclosures of 25 x 25 cm containing marked limpets were set at a mid tidal level on the shore. Twelve treatments with different combination of densities and size-classes were assigned to the plots, with three replicates of each. Decreased density treatments showed very low mortality and substantially increased growth. Both size-classes of limpets showed increased mortality and reduced growth in increased density treatments. This effect was, however, more accentuated for large than small limpets when fenced separately. Although both size-classes could negatively affect each other, the effect of large limpets on small was greater than the reverse. Large limpets are superior competitors that may modulate the abundance of small limpets on the shore. It is unlikely, however, that they will totally exclude small limpets due to intensity of competition within the large size-class. Niche differentiation and high recruitment at low shore levels are other possible factors that can contribute to reduce the effects of this asymmetric competition between the size-classes.
The present work has revealed that both grazing and competition are major biological factors determining the structure of intertidal rocky communities. This study has also contributed to refute old ideas on the causes of distribution patterns. Notwithstanding the obtained results on grazing and competition further experimentation is needed on other aspects of the ecology of intertidal communities.
A presente dissertação teve por objectivo estudar os padrões de distribuição em povoamentos intertidais de substrato rochoso e a influência da herbivoria e competição na estrutura das comunidades. Em Portugal pouca atenção tem sido dedicada ao estudo das comunidades intertidais de substrato rochoso. A costa portuguesa é uma zona de fronteira biogeográfica para espécies de afinidades atlântico-boreais e temperadas-quentes, sendo por isso fundamental o estudo dos povoamentos ao longo de toda a costa. No presente trabalho foi feita a descrição geral dos padrões de zonação ao longo da costa continental portuguesa e a comparação quantitativa dos padrões de distribuição na zona eulitoral. Com o objectivo de testar se existem variações na distribuição e abundância dos organismos na zona eulitoral superior e inferior ao longo de toda a costa, foram consideradas três regiões principais: norte, centro e sul. Nove estações foram visitadas em cada região, correspondendo a um total de 27 praias amostradas. Em cada uma destas estações efectuou-se uma descrição geral dos povoamentos e o levantamento dos padrões de zonação em relação ao zero hidrográfico. A amostragem quantitativa recorreu ao uso de metodologias não-destrutivas e foi efectuada na zona eulitoral superior e inferior, tendo sido amostrados 5 replicados em cada uma destas zonas. Os padrões de zonação ao longo da costa Portuguesa possuem características descritas para costas Norte Atlânticas e Mediterrânicas. Os resultados do presente estudo confirmam também as variações latitudinais do norte para o sul da Europa, com a decrescente cobertura de algas na zona eulitoral mesmo em locais abrigados. De igual modo também a abundância de algas castanhas diminui na franja sublitoral, sendo substituídas por uma zona de algas vermelhas nas regiões mais a sul. Os resultados da análise multidimensional e do teste ANOSIM revelaram diferenças significativas na estrutura das comunidades da zona eulitoral superior e inferior, relacionadas em parte com o número de espécies existentes. De um modo geral, registou-se uma menor diversidade e maior dominância no nível superior. As diferenças na estrutura das comunidades ao longo do gradiente regional não se revelaram tão evidentes como as referidas para o gradiente vertical, apesar do decréscimo na abundância de variadas espécies ao longo da costa (tanto de norte para sul como no sentido inverso). Uma explicação possível para este resultado é o facto do aumento de complexidade das comunidades do nível superior para o nível inferior prevalecer em relação ao gradiente regional. Considerando cada nível separadamente a região do Norte destacou-se das regiões do centro e sul. A análise SIMPER revelou as espécies que mais contribuíram para esta separação bem como aquelas que apresentam um gradiente de abundância ao longo da costa.
Na presente dissertação estudaram-se ainda as causas de determinados padrões de distribuição recorrendo à ecologia experimental, e focando essencialmente aspectos relacionados com a influência da herbivoria e competição nas comunidades intertidais. Em diversas regiões da Europa é frequentemente encontrado na zona intertidal inferior um “tapete” de algas vermelhas, em especial nas zonas de hidrodinamismo moderado. A hipótese de o limite superior deste tapete de algas vermelhas ser determinado por moluscos herbívoros foi testada em Portugal e Inglaterra. Também se procurou avaliar se o efeito provocado por moluscos herbívoros seria alterado por diversos factores que operam a diferentes escalas espaciais. Os gastrópodes foram excluídos recorrendo ao uso de redes, e utilizaram-se ainda controlos para testar o uso da rede (“meia-rede”) e controlos sem qualquer manipulação. As áreas experimentais sem gastrópodes foram rapidamente colonizadas por algas efémeras nos primeiros meses da experiência. Estas algas foram mais tarde substituídas por algas perenes. A percentagem de cobertura de algas vermelhas aumentou significativamente nas duas regiões. O limite superior de distribuição estendeu-se mais de 50 cm na maioria das áreas experimentais. O controlo e “meias-redes” permaneceram desprovidos de algas. Após dois anos do início da experiência, as áreas sem herbívoros eram essencialmente colonizadas por algas vermelhas (e.g. Caulacanthus ustulatus, Gelidium spp., Laurencia pinnatifida) em Portugal, enquanto que em Inglaterra dominavam algas castanhas (Fucus serratus e Himanthalia elongata). É provável que a acção de factores físicos à escala local e geográfica esteja relacionada com estas diferenças. No entanto, apesar dos factores físicos poderem ser
algas. Após a remoção das redes a comunidade retornou rapidamente às condições iniciais antes da manipulação. Quando áreas de algas vermelhas foram removidas imediatamente abaixo do limite normal de distribuição os herbívoros dirigiram-se para estas áreas mas não impediram a posterior colonização por macroalgas. O limite superior do tapete de algas vermelhas parece estar em permanente equilíbrio dinâmico entre a actividade dos herbívoros nas zonas imediatamente acima do limite e a rapidez de colonização e crescimento das algas nas zonas inferiores, sendo provavelmente alterado por factores físicos que favorecem o crescimento das algas ou a eficácia dos herbívoros.
A competição inter- e intraespecífica entre as espécies Patella depressa e Patella vulgata foi analisada na costa norte de Portugal. Densidades aumentadas (x 2, x4), reduzidas (x 1/2) e normais foram utilizadas para testar os efeitos da competição no crescimento e mortalidade das lapas. Na zona eulitoral estabeleceram-se áreas experimentais de 25 x 25 cm vedadas com rede plastificada. Doze tratamentos com diferentes combinações de densidades e espécies foram atribuídos a estas áreas, com três replicados por tratamento. A mortalidade foi registada quinzenalmente e o comprimento foi medido mensalmente durante um período experimental de cinco meses. No final da experiência as lapas foram colhidas tendo em vista o estudo biométrico, a determinação do sexo e a avaliação do estado de maturação da gónada. A quantidade de alimento disponível foi avaliada indirectamente através da determinação da concentração de clorofila existente em pequenas amostras de substrato recorrendo à espectrofotometria de absorção. Ambas as espécies sofreram uma maior mortalidade e redução no crescimento e peso nos tratamentos de densidade aumentada. As lapas nos tratamentos de densidade reduzida exibiram uma baixa mortalidade e um maior crescimento. Apesar de ambas as espécies serem capazes de afectar negativamente a outra, não se registaram diferenças significativas entre o efeito de P. vulgata em P. depressa e o efeito de P. depressa em P. vulgata na mortalidade e crescimento. No entanto, os resultados do peso revelaram que o aumento intraespecífico de densidade levou a uma redução mais acentuada de peso do que quando ambas as espécies estavam presentes. Os resultados obtidos foram comparados com estudos prévios de competição entre moluscos herbívoros e discutidos em relação ao limite de distribuição de P. vulgata na costa portuguesa.
As interacções competitivas entre classes de tamanho, e dentro das mesmas, foram investigadas na espécie Patella depressa, na costa central de Portugal. Foram utilizados doze tratamentos com diferentes combinações de densidades e classes de tamanho, com três replicados de cada. Os tratamentos de densidade reduzida mostraram baixa mortalidade e um aumento substancial no crescimento. Ambas as classes de tamanho sofreram um aumento na mortalidade e crescimento reduzido nos tratamentos de densidade aumentada. Contudo, este efeito foi mais acentuado nas lapas de maiores dimensões do que nas menores, nos tratamentos em que as classes de tamanho foram separadas. Por outro lado, apesar de ambas as classes de tamanho afectarem negativamente a outra, o efeito das lapas de maiores dimensões nas menores foi mais forte que o oposto. As lapas de maiores dimensões revelaram ser competidores superiores que podem moderar a abundância de lapas menores na região intertidal. No entanto, a exclusão total de lapas de menores dimensões é pouco provável devido à alta competição existente dentro da classe de tamanho superior. Outros factores como a ocupação de nichos diferentes e o elevado recrutamento em zonas inferiores poderão também contribuir para minimizar os efeitos da competição assimétrica entre classes de tamanho.
O presente trabalho revelou que a herbivoria e a competição são factores biológicos que desempenham um papel muito importante na estrutura das comunidades intertidais. Este estudo contribuiu também para refutar ideias antigas sobre as causas dos padrões de distribuição. Apesar dos resultados obtidos, é necessário prosseguir com o trabalho experimental relativamente a outros aspectos da ecologia das comunidades intertidais.
TABLE OF CONTENTS
Chapter 1: General introduction 1
Chapter 2: Intertidal rocky shore communities of the continental Portuguese coast: analysis
of distribution patterns 19
Chapter 3: The effects of grazing on the distribution and composition of low shore algal
communities on the central coast of Portugal and on the southern coast of Britain 51
Chapter 4: Analysis of competitive interactions between the limpets Patella depressa
Pennant and Patella vulgata L. in the northern coast of Portugal 87
Chapter 5: Intra- and inter-size-class competition in the limpet Patella depressa Pennant in
the central coast of Portugal 109
CHAPTER 1
CONTENTS
GENERAL INTRODUCTION 3
Why study rocky shores? 3
The shore environment 4
Tides 4
Environmental conditions and adaptations of intertidal organisms 5
Patterns of distribution on intertidal rocky shores 7
Factors determining distribution of species on the shore 8
Rationale and aims 12
GENERAL INTRODUCTION
Why study rocky shores?
The intertidal zone has been of interest for biologists since many years. It can be considered a boundary zone where the transition from an aquatic marine habitat to terrestrial conditions occurs within a restricted area. Therefore any shore is a sharp environmental gradient which is ideal for ecological studies. Additionally it has a great diversity of species, which may be as great or greater than that found in the more extensive subtidal habitats. It should be stressed that this area is an extension of the marine environment and is inhabited almost exclusively by marine organisms. Although plant life is restricted to algae and a few higher plants, examples of most of the major animal groups can be found on the shore. Access to the shore is generally easy and aquatic organisms can be directly observed during low tide without special equipment. Due to all of these features the intertidal zone provides an ideal system for research and education in systematics and taxonomy, ecophysiology, population and community ecology (Connell, 1972; Hawkins & Jones, 1992; Paine, 1994; Raffaelli & Hawkins, 1996; Nybakken, 1997).
Marine intertidal communities are highly productive and have been of interest and use for humans since prehistoric time. Shores are susceptible to a variety of human impacts such as the exploitation of food resources, human settlement, pollution, introduction of new species, coastal zone change or tourism. Coastal development and global climatic changes can have important consequences for shore habitats. Sea level rise as a result of the global warming is an important issue and application of intertidal ecology. Hence, because shores are affected by human impacts and are also more accessible and easier to investigate than other marine systems, they are frequently the subject of biological monitoring and conservation programmes (reviewed in Raffaelli & Hawkins, 1996).
Intertidal rocky shores are particularly easy to study. Many species are sessile and even the mobile animals are generally slow moving. Organisms are taxonomically tractable and relatively short-lived. Their abundance can be easily estimated as percentage cover or density in a non-destructive way, allowing the dynamics of populations or communities to be studied without too much interference. Most importantly, because they are easy to manipulate, they are particularly suitable for an experimental approach. This enables a better understanding of the functioning of rocky shore communities (Paine, 1966, 1994; Connell, 1972; Little & Kitching, 1996; Raffaelli & Hawkins, 1996).
In summary, the richness, the diversity of environmental factors, and the ease of access attracted to this area a considerable amount of scientific attention. This has proved important to produce unifying concepts and models regarding the organisation of marine communities and community ecology in general.
The shore environment
Tides
The study of marine intertidal communities involves the comprehension of the shore environmental conditions, particularly the tides. Tides are certainly an over-riding physical feature influencing intertidal communities by modification of the gradient between the sea and land (see discussion in Raffaelli & Hawkins, 1996).
The periodic, predictable rise and fall of the level of the sea over a given time is called a tide. This is actually a wave of very long wavelength. Its astronomical origin is due to the interaction of the gravitational attraction of the Sun and the Moon on the Earth and the forces generated by the rotating Earth and Moon system. As a result of these forces, the water in ocean basins is pulled into bulges- “two high tides”- in line with the Moon. As the Moon rotates around its common axis with the Earth these tides circle the Earth. The simplest and most common tidal pattern is a semidiurnal one, with two high and low tides each day. The two tides can be of different sizes, which is partially due to the varying declination of the Moon. The effect of the Sun is seen in spring and neap tides. When the gravitational forces of the Sun and the Moon act together, i.e. when Sun, Moon and Earth are directly aligned, tide-raising forces are at a maximum. This occurs at the times of new and full Moon and results in tides of very large amplitude called spring tides. Conversely, when the Moon and Sun are at right angles to the Earth, the gravitational attractions of Sun and Moon act in opposition. Tide-raising forces on the Earth are at minimum resulting in tides of small amplitude called neap tides. Seasonal differences in the amplitude of spring tides are caused by the changing declination of the Sun. The highest spring tides occur at equinoxes, while at the solstices spring tides are at their lowest amplitude (Duxbury & Duxbury, 1993; Little & Kitching, 1996; Raffaelli & Hawkins, 1996; Nybakken, 1997; Thurman, 1997).
Different tidal patterns such as semidiurnal, diurnal and mixed occur in various parts of the world. Variations in the number and in the height of high and low tides per day in different regions are related to complexities caused by several factors. These include, for example,
presence of land masses, peculiarities of the ocean basins or the Coriolis force (Raffaelli & Hawkins, 1996).
Despite this enormous variation in the behaviour of tides, they are in general predictable and knowledge of the timing of tides and of mean tidal heights is essential when working on a shore. Tidal heights are measured in relation to a conventional level called chart datum (CD), which corresponds to the lowest astronomical tide. Useful mean heights above this level are the mean values for high and low water of spring and neap tides (MHWS, MHWN, MLWN, MLWS). The mean tidal level (MTL) is the average of these four heights. Extreme water of spring tides (EHWH, EHWL) can also be used for representation of the height above chart datum.
Environmental conditions and adaptations of intertidal organisms
The main feature which tidal regime imposes on the littoral is the alternation of emersion and submersion periods, as irregular in duration and frequency these might be (Lewis, 1964). It can affect the intertidal communities through the duration of exposure to air and time of the day during which it occurs. Additionally, certain rhythms of activity of intertidal organisms are also related to tidal effect (Nybakken, 1997).
Most of the intertidal species evolved from marine ancestors. The marine environment is relatively stable. Salinity is constant and temperature variations are small. Carbon dioxide, water and oxygen needed for photosynthesis and respiration are abundant, and nutrients are readily available being rarely limiting in coastal waters (Hawkins & Jones, 1992). Therefore, from low water to the spray zone the environmental gradient is of increasingly harsh physical conditions, due to greater fluctuations in environmental conditions associated with the increased proportion of time exposure to the air (Hawkins & Jones, 1992; Raffaelli & Hawkins, 1996). Similarly, adaptations of intertidal organisms are intimately related with avoidance or minimisation of the stress of air exposure (Nybakken, 1997).
Intertidal organisms show various mechanisms for avoiding water loss. The simplest mechanism is to avoid heat and air exposure and, therefore, mobile animals (e.g. crabs) may seek for refuge in moist crevices or under algae canopy (Nybakken, 1997). Another mechanism, which is shown by several intertidal algae (e.g. Porphyra, Ulva and Enteromorpha), is to be simply adapted to withstand a severe water loss from their tissues (Kanwisher, 1957). Fucoid algae also seem to lack physiological or structural means to retard water loss (Schonbeck & Norton, 1979). Many other intertidal species, however, possess
structural and behavioural mechanisms to prevent desiccation (Newell, 1979; Nybakken, 1997). Sessile animals avoid water loss by closing their shells at low tide. Mussels and other bivalves have the ability to close their valves tightly to prevent desiccation. Barnacles have lateral calcareous plates and aperture plates that when closed cut down water loss. Limpets have a “homescar” into which their shells fit, and where they generally return at low tide. It has been suggested that this close fit would reduce desiccation (Lewis, 1954), but other hypotheses for the advantages of homing behaviour have been reviewed by Underwood (1979). Other gastropods have opercula that completely seal the aperture of the shell (Nybakken, 1997). At low tide there may also exist osmotic problems. A classical example is that of rock pools where salinity may be increased by evaporation or reduced by rainfall (Lewis, 1964; Hawkins & Jones, 1992). The responses of organisms to such phenomenon are generally the same as those observed for preventing desiccation (Nybakken, 1997).
Most of intertidal species conduct their main activities when immersed or immediately after the tide recedes (Lewis, 1964). Many filter feeder animals (e.g. mussels, barnacles) can only feed when immersed. Many grazers and predators also move and forage more efficiently at high tide or nocturnal low tide, when desiccation is less intense. Large mobile predators (e.g. crabs, fishes) will also have a longer feeding time low on the shore (Lewis, 1964; Hawkins & Jones, 1992). Respiration is similarly more effective in the water than in the air. Algae need to be surrounded by water for respiration, photosynthesis and nutrient uptake. The majority of the animals have respiratory surfaces adapted to extract oxygen from the water. There are, however, many adaptations protecting exchange structures and allowing intertidal animals to respire in air (e.g. modified gills, reduction of the gills and formation of a vascularised mantle cavity, anaerobic respiration) (Newell, 1979; Nybakken, 1997). The alternation emersion and immersion periods seem to have induced of activity rhythms in intertidal organisms (Lewis, 1964, Naylor, 1985). The existence of breeding cycles synchronised with certain tides to ensure fertilisation and larval dispersion can be seen as a reproductive adaptation to tidal fluctuations (Nybakken, 1997).
Exposure to wave action is another major environmental gradient in the intertidal zone. It is essentially a horizontal gradient but stress does not occur in a clearly defined direction (Raffaelli & Hawkins, 1996). In general, intertidal organisms can live higher up on the shore in exposed than sheltered conditions. This is mainly due to wave spray on exposed shores, which can minimise desiccation problems, particularly for high shore species. Exposed conditions are favourable for filter-feeders because water movement will provide plenty of suspended food particles. For mobile species, however, problems of dislodgement increase with wave action and foraging may become more difficult. Conversely, siltation can be a
problem in sheltered conditions as it may clog the gills of some animals. In extreme shelter conditions the lack of water movement may also restrict the supply of oxygen for both plants and animals, dissolved nutrients for seaweeds and reduce larval supply (Hawkins & Jones, 1992; Little & Kitching, 1996; Raffaelli & Hawkins, 1996).
Both vertical and horizontal gradients can be modified by several factors including the topography and geology of the shore and a variety of biotic influences (Lewis, 1964; Raffaelli & Hawkins, 1996).
Patterns of distribution on intertidal rocky shores
Patterns of distribution of intertidal rocky shore communities have long been studied by marine biologists. Classical descriptive works have focussed on the characteristic distribution of species in horizontal bands- zonation patterns (e.g. Stephenson & Stephenson, 1949, 1972; Lewis, 1964; Pérès & Picard, 1964). This type of distribution pattern is not exclusive of marine ecosystems and may be observed, for example, on terrestrial vegetation on mountain slopes. However, due to the sharp environmental gradient on intertidal rocky shores, zonation patterns are particularly striking and spatial condensed, and have attracted a considerable amount of attention (Russell, 1991). The recognition that shores from different parts of the world had certain biological features in common led Stephenson & Stephenson (1949) to propose an “universal” scheme of zonation for intertidal rocky shores. According to this classification scheme the shore could be divided into three main zones: i) a high shore zone to which they called supralittoral fringe, characterised by encrusting lichens, cyanobacteria, small littorinid gastropods and isopods; ii) a mid-shore zone called midlittoral zone, dominated by filter-feeders such as barnacles and mussels and; iii) a lower shore zone called
infralittoral fringe, dominated by red algae and large brown kelps.
In his the extensive study of zonation patterns on the British Isles, Lewis (1961, 1964) used a slightly different terminology. The populations of the coast and shallow seas were divided into littoral and sublittoral zones. The former corresponds to marine organisms which are adapted to or need alternating emersion and submersion periods or spray. Within the littoral zone Lewis (1964) called littoral fringe to the top zone and eulittoral to the mid-shore zone. The part of sublittoral zone that can be exposed at low tide was called sublittoral fringe. The main difference in the two classifications systems is the used terminology and the extension of the littoral zone. Stephenson & Stephenson (1972) littoral zone corresponded to the zone between extreme high and low water of spring tides, including the infralittoral fringe
and part of the supralittoral fringe. They were both, however, based upon biological characteristics and the different zones were not established according to tide levels, though they may be related to them.
A similar approach was adopted for the Mediterranean by Pérès & Picard (1964). These authors proposed a more complex zonation system from coastal to deep oceanic marine zones. None the less, the main zones described for littoral communities were essentially the same as those established in the Stephenson-Lewis approach. Three primary zones each characterised by particular kinds of organisms.
The reference to zonation patterns may be advantageous for comparison of work carried on different parts of the world, or to define the studied area in terms of zone or habitat instead of the precise tidal level (Raffaelli & Hawkins, 1996). The terminology used by Lewis (1964) was adopted in the present work to refer the three major intertidal zones: littoral fringe (high-shore), eulittoral zone (mid-shore) and sublittoral fringe (low-shore).
Factors determining distribution of species on the shore
Interested by the studies on zonation patterns (e.g. Lewis, 1964; Brättstrom, 1980; Norton, 1985; Russell, 1991) marine ecologists tried to explain the causes of the observed distribution patterns. Consequently, they started to analyse the influence of physical and biological factors on marine intertidal communities (e.g. Southward, 1958; Ballantine, 1961; Connell, 1972; Saldanha, 1974; McQuaid & Branch 1984; Hawkins & Hartnoll, 1985) and to perform experimental analyses to understand the functioning of rocky shore communities (e.g. Dayton 1971; Paine, 1974; Menge, 1976; Lubchenco & Menge, 1978; Benedetti-Cecchi et al., 1996). Manipulative studies of community dynamics involved the analysis of physical disturbance and succession on intertidal communities (e.g. Dayton, 1971; Emerson & Zedler, 1978; Sousa, 1979, 1984; Dethier, 1984; Farrell, 1991; Benedetti-Cecchi & Cinelli, 1993; McCook & Chapman, 1997) or experimental investigations on the effects of competition (e.g. Connell, 1961; Branch, 1976; Underwood, 1978a, 1984), grazing (e.g. Lubchenco, 1978, 1980; Hawkins, 1981; Hawkins & Hartnoll, 1983; Benedetti-Cecchi & Cinelli, 1992; Dye, 1993; Benedetti-Cecchi et al., 1996; Jenkins et al., 1999a, 1999b), predation (e.g. Paine, 1966; Menge 1978a,b, 1983; Menge & Lubchenco, 1981) and recruitment (e.g. Thorson, 1966; Denley & Underwood, 1979; Hawkins & Hartnoll, 1982; Kendall et al., 1985; Petraitis, 1990; Menge, 1991, Jenkins et al., 1999c).
Considerable attention has been devoted to the study of vertical distribution of some species on the shore. Early ideas to account for the pattern of zonation and vertical distribution of species were mainly related to physical factors. The concept that critical tidal levels were important determinants of the structure of intertidal communities was first suggested by Colman (1933) and was adopted by several authors for many years (e.g. Doty, 1946; Evans, 1947). Critical levels were calculated from predicted tide tables, based on the examination of a curve of proportion of emersion time throughout the year. These levels were apparently coincident with the upper and lower limits of several intertidal species. However, the concept of critical tidal levels was refuted by Underwood (1978b) after the experimental examination of this hypothesis. He found no evidence that the limits of distribution of species were clumped at any particular levels.
Physical factors have long been thought to be responsible for the upper limits of distribution of most intertidal species (e.g. Baker, 1909; Connell, 1972). Experimental work has shown the importance of aerial exposure and desiccation in controlling the upper limits of distribution of high- and mid-shore fucoid algae (Schonbeck & Norton, 1978). Lower on the shore evidence was provided that physical factors determined the upper limit of some red algae (Lubchenco, 1980; Carter & Anderson, 1991). In contrast, lower limits of distribution have usually been associated with biological factors like competition, grazing or predation (Schonbeck & Norton, 1980; Lubchenco, 1980; Hawkins & Hartnoll, 1985). Experimental investigations of the factors affecting algal zonation in New England rocky shores by Lubchenco (1980) led to the conclusion that competition is the primary determinant of Fucus sp. lower limits (affecting presence or absence) and herbivory is of secondary importance (affecting abundance). Interspecific algal competition has also been shown to set lower limits of fucoid algae (Pelvetia canaliculata (L.) Decaisne & Thuret and Fucus spp.) in the U.K., in several experiments involving both field transplants, exclusion areas and laboratory cultures (Schonbeck & Norton, 1980) and canopy removal (Hawkins & Hartnoll, 1985).
More recently, the importance of biological factors such as grazing and competition have been shown to set upper distribution limits of some species (Underwood 1980, 1985; Sousa et al., 1981; Underwood & Jernakoff, 1981, 1984; Hawkins & Hartnoll, 1985). After the Torrey Canyon oil spill, large-scale kills of limpets due to excessive dispersant application resulted in upward extension of some low shore species (Southward & Southward, 1978). Underwood (1980) demonstrated that upper limit of foliose macroalgae in Australian shores was primarily set by herbivorous molluscs. Thus, the upper limits of these lower algal beds would depend on the grazing of algal propagules by gastropods and only in the absence of grazers physical factors would become important in determining the upper limit of macroalgae (Jernakoff,
1983). Some canopy forming algae have also been induced to extend upwards after the removal of competitor algal species (Hawkins & Hartnoll, 1985).
The role of competitive interactions as structuring agents in marine benthic communities has been intensively discussed on intertidal rocky shores (e.g. Connell, 1961; Dayton, 1971; Haven 1973; Menge, 1976; Underwood, 1978a). Several reviews on the experimental analyses of competition in the field have been published in the last decades (e.g. Connell, 1983; Schoener, 1983; Underwood 1986, 1992). Birch’s (1957) definition of competition as “the process leading to ‘harm’ to individuals when other individuals of the same or different species use the same resources which are in short supply” has been generally adopted in the marine ecological literature (Underwood, 1979, 1992, 1997; Connell, 1983; Hawkins & Hartnoll, 1983).
According to Underwood (1978a, 1992) there are three major reasons why competitive interactions should be well documented for intertidal communities and be regarded with undoubted importance as one of the processes determining the structure of assemblages of species in this habitat. Firstly, space is a common resource for all intertidal organisms, either as a substratum on which to live, or over which to feed. Competitive interactions for space are known to be important and critical in many intertidal habitats, because space, as a two dimensional resource, is often in short supply. Secondly, the majority of the intertidal organisms have widespread dispersive stages of their life history. Since there is little opportunity for regulation of numbers in local populations, recruitment to the shore can be in unpredictably high densities, and situations in which numbers of organisms increase beyond carrying capacity of some local habitat can be widespread in organisms with dispersive propagules. Thus, competitive interactions have often been studied in such organisms in intertidal rocky shores. Thirdly, intertidal rocky shore organisms are very suitable for experimental manipulations, due to the high density and sessile characteristics (e.g. algae, barnacles) or relatively sedentary nature (e.g. limpets, chitons, starfish) of most organisms. Consequently, experimental manipulations can be done in relatively small areas and be easily controlled and replicated in space and time.
Competition is generally divided into exploitative competition, where individuals, by using resources, deprive others of benefits to be gained from those resources; and interference competition, where individuals harm one another to acquire the resources (e.g. by fighting, producing toxins) (Schoener, 1983).
Several forms of competitive interactions were also described with more detail by Schoener (1983). He considered that consumptive competition could occur when some quantity of resource (e.g. food, water, nutrient) is consumed by an individual, thereby depriving other individuals of it. Pre-emptive competition occurs when one organism already occupies space, making it unavailable for another individuals; it occurs primarily in sessile organisms. Overgrowth competition occurs when another individual or individuals grow over or upon a given individual, therefore depriving that individual of light (as in plants) or access to water-borne food (as in sessile, filter feeding animals) and possibly harming that individual by some consequence or physical contact. Chemical competition occurs when an individual produces some chemical (toxin) which diffuses into the medium or substrate and harms other individuals. Territorial competition occurs when an individual aggressively defends, or by its behaviour signals its intention to defend, a unit of space against other individuals; it occurs primarily in mobile organisms. Encounter competition occurs as the result of an interaction between mobile, non-spatially attached individuals, in which some harm comes to one or more; such harm can include energy losses, theft of food, injury, or death by predation, fighting or mere accident.
Early work on the role of competition in intertidal rocky shore communities focussed on sessile species (e.g. Connell, 1961; Dayton, 1971; Menge, 1976). Sessile organisms such as barnacles and mussels can compete for space when this is in short supply either by pre-emptive or overgrowth mechanisms. In many of these studies it has been suggested that importance of competitive interactions may very often be reduced due to factors like predation or disturbance that may reduce population densities of the competing species to such low levels that there are sufficient resources for all species (e.g. Paine, 1966, 1974; Dayton, 1971). The inferior competitors could have access to free space, and coexist with the potentially dominant species.
The recognition that competition among mobile animals on rocky shores may be fundamentally different from that among sessile organisms (Underwood, 1978a; Creese & Underwood, 1982) led to proliferation of studies on the importance of competition among mobile gastropods species (Marshall & Keough, 1994). Intertidal gastropods are often considered to be well suited to studies of competition (Underwood, 1984) and, experimental investigations in many parts of the world revealed that competition interactions are a prevalent characteristic of many species of grazing gastropods, particularly limpets (e.g. Branch, 1975a, b, 1976; Underwood, 1978a, 1979, 1984; Black, 1979; Creese, 1980; Creese and Underwood, 1982).
Both exploitative and interference competition can occur among intertidal gastropod species (Underwood, 1979; Branch, 1981). Territorial behaviour of gastropods that actively defend an unit of space over which they feed against the intrusion of other grazers were described for Lottia gigantea (Stimson, 1970) in North American coasts, and for several species of South African limpets such as Pattela longicosta and Patella tabularis (Branch 1975b, 1981). Inter- and intraspecific exploitative competition have also been experimentally demonstrated in a great number of studies (e.g. Underwood, 1978a, 1984; Creese and Underwood, 1982; Lasiak & White, 1993; Marshall & Keough, 1994). These investigations have shown that competition can have important consequences on the population dynamics and structure of marine assemblages. However, a multiplicity of processes can modify the outcome of competitive interactions (e.g. physical factors in the environment, disturbances, predators). Therefore, these studies need to be repeated at different spatial or temporal scales, and with a large number of species, so that any conclusion about the general effects of competition can be reached (Connell, 1983; Underwood, 1992).
Rationale and aims
The present work aims to analyse patterns of distribution of intertidal species on rocky shores and to investigate the role of grazing and competition in structuring communities. In Chapter 2, intertidal zonation patterns along the continental Portuguese coast are described and the quantitative comparison of distribution patterns in the mid-shore zone was made. So far, the intertidal rocky shore communities of the Portuguese coast have been studied in a scattered way by different authors and most of the information concerns a specific biological group or the communities of a portion of the coast. A study of this nature, covering the whole littoral coast, is particularly important if we consider that Portugal is a geographical limit for both boreal and subtropical and Mediterranean species. It may contribute for studies on the response of species distribution to global climatic change. There is also the necessity to provide baseline studies for future management and conservation programmes on intertidal rocky shores. Additionally, this study is fundamental as a reference for experimental work and may also provide an observational basis for testing hypotheses. Subsequently, the remaining chapters were devoted to experimental ecology studies on causes of distribution patterns, essentially focussed on the importance of grazing and competition by limpets in intertidal communities. The factors determining vertical distribution patterns on the shore, particularly the effects of grazing limpets on the upper limits of low shore algae, were investigated in Chapter 3. The hypothesis that grazing by limpets directly determines the upper limit of distribution of the red algal turf was tested. The work was undertaken in both Portugal and U.K. with a nested experimental design to assess whether the grazing effect is
modified by different factors operating at various spatial scales. The analysis of competition interactions amongst the limpets Patella depressa Pennant and Patella vulgata L. on the northern coast of Portugal, was done in Chapter 4. Little is know about competition interactions among these species. Both species are abundant grazers that co-occur in mid-littoral zone of the northernmost shores of Portugal. However, Patella vulgata is a northern species, and its density decreases along the Portuguese coast, being occasional or rare in some shores further to the south. The proximity to the southern limit of distribution of Patella vulgata, makes it an attractive subject for testing hypotheses about interspecific competition. Inter- and intraspecific interactions were analysed simultaneously through an asymmetric experimental design to estimate their relative importance. A further analysis on intra- and inter-size-class competition in the limpet Patella depressa was conducted in the central coast of Portugal. This experiment is described in Chapter 5 and tested the effects of increased and decreased densities on parameters such as mortality, growth and weight of the limpets (among others) and the influence of different size classes in competitive interactions. The major results and conclusions obtained from the various chapters are integrated in Chapter 6.
REFERENCES
Baker, S.M. (1909). On the causes of the zoning of brown seaweeds on the seashore. New Phytologist, 8: 196-202.
Ballantine, W.J. (1961). A biologically-defined exposure scale for the comparative description of rocky shores. Field Studies, 1 (3): 1-17.
Benedetti-Cecchi, L. & F. Cinelli (1992). Effects of canopy cover, herbivores and substratum type on patterns of Cystoseira spp. settlement and recruitment in littoral rockpools. Marine Ecology Progress Series, 90: 183-191.
Benedetti-Cecchi, L. & F. Cinelli (1993). Early patterns of algal succession in a midlittoral community of the Mediterranean sea: a multifactorial experiment. Journal of Experimental Marine Biology and Ecology, 169: 15-31.
Benedetti-Cecchi, L.; S. Nuti & F. Cinelli (1996). Analysis of spatial and temporal variability in interactions among algae, limpets and mussels in low-shore habitats on the west coast of Italy. Marine Ecology Progress Series, 144: 87-96.
Birch, L.C. (1957). The meanings of competition. American Naturalist, 91: 5-18.
Black, R. (1979). Competition between intertidal limpets: an intrusive niche on a steep resourse gradient. Journal of Animal Ecology, 48: 401-411.
Branch, G.M. (1975a). Intraspecific competition in Patella cochlear Born. Journal of Animal Ecology, 44: 263-282.
Branch, G.M. (1975b). Mechanisms reducing intraspecific competition in Patella spp.: migration, differentiation and territorial behaviour. Journal of Animal Ecology, 44: 575-600.
Branch, G.M. (1976). Interspecific competition experienced by South African Patella species. Journal of Animal Ecology, 45: 507-529.
Branch, G.M. (1981). The biology of limpets: physical factors, energy flow, and ecological interactions. Oceanography and Marine Biology Annual Review, 19: 235-380.
Brattström, H. (1980). Rocky shore zonation in the Santa Marta area, Colombia. Sarsia, 65: 163-226.
Carter, A.R. & R.J. Anderson (1991). Biological and physical factors controlling the spatial distribution of the intertidal alga Gelidium pristoides in the Eastern Cape, South Africa. Journal of the Marine Biological Association of the U.K., 71: 555-568.
Colman, J.S. (1933). The nature of intertidal zonation of plants and animals. Journal of the Marine Biological Association of the U.K., 61: 71-93.
Connell, J.H. (1961). The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology, 42 (4): 710-723.
Connell, J.H. (1972). Community interactions on marine rocky intertidal shores. Annual Review of Ecology and Systematics, 3: 169-192.
Connell, J.H. (1983). On the prevalence and relative importance of interspecific competition: evidence from field experiments. The American Naturalist, 122 (5): 661-696.
Creese, R.G. (1980). An analysis of distribution and abundance of populations of the high-shore limpet, Notoacmea petterdi (Tenison-Woods). Oecologia, 45: 252-260.
Creese, R.G. & A.J. Underwood (1982). Analysis of inter- and intra-specific competition amongst intertidal limpets with different methods of feeding. Oecologia, 53: 337-346. Dayton, P.K. (1971). Competition, disturbance, and community organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecological Monographs, 41 (4): 351-389.
Denley, E.J. & A.J. Underwood (1979). Experiments on factors influencing settlement, survival, and growth of two species of barnacles in New South Wales. Journal of Experimental Marine Biology and Ecology, 36: 269-293.
Dethier, M.N. (1984). Disturbance and recovery in intertidal pools: maintenance of mosaic patterns. Ecological Monographs, 54(1): 98-118.
Doty, M.S. (1946). Critical tide factors that are correlated with the vertical distribution of marine algae and other organisms along the Pacific coast. Ecology, 27: 315-328. Duxbury, A.B. & A.C. Duxbury (1993). Fundamentals of oceanography. Wm.C. Brown
Publishers, Dubuque, Iowa: 291pp.
Dye, A.H. (1993). Recolonization of intertidal macroalgae in relation to gap size and molluscan herbivory on a rocky on the east coast of southern Africa. Marine Ecology Progress Series, 95: 263-271.
Emerson, S.E. & J.B. Zedler (1978). Recolonization of intertidal algae: an experimental study. Marine Biology, 44: 315-324.
Evans, R.G. (1947). The intertidal ecology of selected localities in the Plymouth neighbourhood. Journal of the Marine Biological Association of the U.K., 17: 173-218.
Farrell, T.M. (1991). Models and mechanisms of succession: an example from a rocky intertidal community. Ecological Monographs, 61(1): 95-113.
Haven, S.B. (1973). Competition for food between the intertidal gastropods Acmaea scabra and Acmaea digitalis. Ecology, 54: 143-151.
Hawkins, S.J. (1981). The influence of Patella grazing on the fucoid/barnacle mosaic on moderately exposed rocky shores. Kieler Meeresforschungen, 5: 537-543.
Hawkins, S.J. & R.G. Hartnoll (1982). Settlement patterns of Semibalanus balanoides (L.) in the Isle of Man (1977-1981). Journal of Experimental Marine Biology and Ecology,
62: 271-283.
Hawkins, S.J. & R.G. Hartnoll (1983). Grazing of intertidal algae by marine invertebrates. Oceanography and Marine Biology Annual Review, 21: 195-282.
Hawkins, S.J. & R.G. Hartnoll (1985). Factors determining the upper limits of intertidal canopy-forming algae. Marine Ecology Progress Series, 20: 265-271.
Hawkins, S.J. & H.D. Jones (1992). Rocky shores. Marine field course guide 1. Immel Publishing, London: 144pp.
Jenkins, S.R.; S.J. Hawkins & T.A. Norton (1999a). Direct and indirect effects of a macroalgal canopy and limpet grazing in structuring a sheltered intertidal community. Marine Ecology Progress Series, 188: 81-92.
Jenkins, S.R.; S.J. Hawkins & T.A. Norton (1999b). Interaction between a fucoid canopy and limpet grazing in structuring a low shore intertidal community. Journal of Experimental Marine Biology and Ecology, 233 (1): 41-63.
Jenkins, S.R.; T.A. Norton & S.J. Hawkins (1999c). Settlement and post-settlement interactions between Semibalanus balanoides (L.) (Crustacea: Cirripedia) and three species of fucoid canopy algae. Journal of Experimental Marine Biology and Ecology, 236 (1): 49-67.
Jernakoff, P. (1983). Factors affecting the recruitment of algae in a midshore region dominated by barnacles. Journal of Experimental Marine Biology and Ecology, 67: 17-31.
Kanwisher, J. (1957). Freezing and drying in intertidal algae. Biological Bulletin of the Marine Biological Laboratory, Woods Hole, 113: 275-285.
Kendall, M.A.; R.S. Bowman; P. Williamson & J.R. Lewis (1985). Annual variation in the recruitment of Semibalanus balanoides on the North Yorkshire coast 1969-1981. Journal of the Marine Biological Association of the U.K., 65: 1009-1030.
Lasiak, T.A. & D.R.A. White (1993). Microalgal food resources and competitive interactions among the intertidal limpets Cellana capensis (Gmelin, 1791) and Siphonaria concinna Sowerby, 1824. South African Journal of Marine Science, 13: 97-108. Lewis, J.R. (1954). Observations on a high-level population of limpets. Journal of Animal
Ecology, 23: 85-100.
Lewis, J.R. (1961). The littoral zone on rocky shores- a biological or physical entity? Oikos,
12: 280-301.
Lewis, J.R. (1964). The ecology of rocky shores. English Universities Press Ltd., London: 300pp.
Little, C. & J.A. Kitching (1996). The biology of rocky shores. Oxford University Press, Oxford: 240pp.
Lubchenco, J. (1978). Plant species diversity in a marine intertidal community: importance of herbivore food preference and algal competitive abilities. The American Naturalist,
112: 23-39.
Lubchenco, J. (1980). Algal zonation in the New England rocky intertidal community: an experimental analysis. Ecology, 61 (2): 333-344.
Lubchenco, J. & B.A. Menge (1978). Community development and persistence in a low rocky intertidal zone. Ecological Monographs, 59: 67-94.
Marshall, P.A. & M.J. Keough (1994). Asymmetry in intraspecific competition in the limpet Cellana tramoserica (sowerby). Journal of Experimental Marine Biology and Ecology, 177: 121-138.
McCook, L.J. & A.R.O. Chapman (1997). Patterns and variations in natural succession following massive ice-scour of a rocky intertidal seashore. Journal of Experimental Marine Biology and Ecology, 214:121-147.
McQuaid, C.D. & G.M. Branch (1984). Influence of sea temperature, substratum and wave exposure on rocky intertidal communities: an analysis of faunal and floral biomass. Marine Ecology Progress Series, 19: 145-151.
Menge, B.A. (1976). Organisation of the New England rocky intertidal communities: role of predation, competition and environmental heterogeneity. Ecological Monographs, 46: 355-393.
Menge, B.A. (1978a). Predation intensity in a rocky intertidal community. Relation between predator foraging activity and environmental harshness. Oecologia, 34: 1-16.
Menge, B.A. (1978b). Predation intensity in a rocky intertidal community. Effect of an algal canopy, wave action and desiccation on predator feeding rates. Oecologia, 34: 17-35. Menge, B.A. (1983). Components of predation intensity in the low zone of the New England
rocky intertidal region. Oecologia, 58: 141-155.
Menge, B.A. (1991). Relative importance of recruitment and other causes of variation in rocky intertidal community structure. Journal of Experimental Marine Biology and Ecology, 146: 69-100.
Menge, B.A. & J. Lubchenco (1981). Community organization in temperate and tropical rocky intertidal habitats: prey refuges in relation to consumer pressure gradients. Ecological Monographs, 51(4): 429-450.
Naylor, E. (1985). Tidally rhythmic behaviour of marine animals. Symposia of the Society for Experimental Biology, 39: 63-93.
Newell, R.C. (1979). Biology of intertidal animals. Marine Ecological Surveys, Faversham: 781pp.
Norton, T.A. (1985). The zonation of seaweeds on rocky shores. In The ecology of rocky coasts, (ed. P.G. Moore & R. Seed), Hodder and Stoughton, London: 7-21.
Nybakken, J.W. (1997). Marine Biology. An ecological approach. Benjamin Cummings, Menlo Park, California: 481pp.
Paine, R.T. (1966). Food web complexity and species diversity. The American Naturalist,
100: 65-75.
Paine, R.T. (1974). Intertidal community structure. Experimental studies on the relationship between a dominant competitor and its principal predator. Oecologia, 15: 93-120. Paine, R.T. (1994). Marine rocky shores and community ecology: an experimentalist’s
perspective. Ecology Institute, Nordbruite: 159pp.
Pérès, J.-M. & J. Picard (1964). Nouveau manuel de bionomie benthique de la mer Mediterranée. Recueil des travaux de la station marine d'Endoume 31 (47): 1-137. Petraitis, P.S. (1990). Direct and indirect effects of predation, herbivory and surface rugosity
on mussel recruitment. Oecologia, 83: 405-413.
Raffaelli, D. & S.J. Hawkins (1996). Intertidal ecology. Chapman & Hall, London: 356pp. Russell, G. (1991). Vertical distribution. In Intertidal and littoral ecosystems, (ed. A.C.
Mathieson & P.H. Nienhuis), Elsevier, Amsterdam: 43-65.
Saldanha, L. (1974). Estudo do povoamento dos horizontes superiores da rocha litoral da costa da Arrábida (Portugal). Arquivos do Museu Bocage 2ª Série, 1: 1-382pp.
Schoener, T.W. (1983). Field experiments on interspecific competition. The American Naturalist, 122 (2): 240-285.
Schonbeck, M.W. & T.A. Norton (1978). Factors controlling the upper limits of fucoid and algae on the shore. Journal of Experimental Marine Biology and Ecology, 31: 303-313.
Schonbeck, M.W. & T.A. Norton (1979). An investigation of drought avoidance in intertidal fucoid algae. Botanica Marina, 22: 133-144.
Schonbeck, M.W. & T.A. Norton (1980). Factors controlling the lower limits of fucoid algae on the shore. Journal of Experimental Marine Biology and Ecology, 43: 131-150. Sousa, W.P. (1979). Experimental investigations of disturbance and ecological succession in a
rocky intertidal algal community. Ecological Monographs, 49 (3): 227-254.
Sousa, W.P. (1984). The role of disturbance in natural communities. Annual Review of Ecology and Sysytematics, 15: 353-391.
Sousa, W.P.; S.C. Schroeter & S.D. Gaines (1981). Latitudinal variation in intertidal algal community structure: the influence of grazing and vegetatitive propagation. Oecologia, 48: 297-307.
Southward, A.J. (1958). Note on the temperature tolerances of some intertidal animals in relation to environmental temperatures and geographical distribution. Journal of the Marine Biological Association of the U.K., 37: 49-66.
Southward, A.J. & E.C. Southward (1978). Recolonization of rocky shores in Cornwall after use of toxic dispersants to clean up the Torrey Canyon spill. Journal of Fisheries Research Board of Canada, 35: 682-706.
Stephenson, T.A. & A. Stephenson (1949). The universal features of zonation between tidemarks on rocky coasts. Journal of Ecology, 38: 289-305.
Stephenson, T.A. & A. Stephenson (1972). Life between tidemarks on rocky shores. W. H. Freeman & Co., San Francisco: 425pp.
Stimson, J. (1970). Territorial behaviour of the owl limpet, Lottia gigantea. Ecology, 51 (1): 113-118.